
Abstract
Context: Clove (Eugenia caryophyllata Thunb. [Myrtaceae]) essential oil (CEO) has been shown to possess antimicrobial, antifungal, antiviral, antioxidant, anti-inflammatory and anticancer properties. However, few studies have focused on its topical use.
Objective: We investigated the biological activity of a commercially available CEO in a human skin disease model.
Materials and methods: We evaluated the effect of CEO on 17 protein biomarkers that play critical roles in inflammation and tissue remodelling in a validated human dermal fibroblast system, which was designed to model chronic inflammation and fibrosis. Four concentrations of CEO (0.011, 0.0037, 0.0012, and 0.00041%, v/v) were studied. The effect of 0.011% CEO on genome-wide gene expression was also evaluated.
Results and discussion: CEO at a concentration of 0.011% showed robust antiproliferative effects on human dermal fibroblasts. It significantly inhibited the increased production of several proinflammatory biomarkers such as vascular cell adhesion molecule-1 (VCAM-1), interferon γ-induced protein 10 (IP-10), interferon-inducible T-cell α chemoattractant (I-TAC), and monokine induced by γ interferon (MIG). CEO also significantly inhibited tissue remodelling protein molecules, namely, collagen-I, collagen-III, macrophage colony-stimulating factor (M-CSF), and tissue inhibitor of metalloproteinase 2 (TIMP-2). Furthermore, it significantly modulated global gene expression and altered signalling pathways critical for inflammation, tissue remodelling, and cancer signalling processes. CEO significantly inhibited VCAM-1 and collagen III at both protein and gene expression levels.
Conclusions: This study provides important evidence of CEO-induced anti-inflammatory and tissue remodelling activity in human dermal fibroblasts. This study also supports the anticancer properties of CEO and its major active component eugenol.
Introduction
Clove (Eugenia caryophyllata Thunb. [Myrtaceae]) essential oil (CEO) is topically used for a variety of health purposes. Scientific studies have evaluated its antimicrobial, antifungal, antiviral, antioxidant, anti-inflammatory and anticancer properties in a variety of models. However, research regarding its biological activity in human skin cells is scarce. Prashar et al. (Citation2006) reported that CEO and its major active component eugenol displayed cytotoxicity against human fibroblasts and endothelial cells, at concentrations as low as 0.03% (v/v). Koh et al. (Citation2013) showed the anti-inflammatory activity of eugenol in human gingival fibroblast and pulp cells.
In this study, we investigated the biological activity of a commercially available CEO in a well-validated human skin disease model. We studied the effect of CEO on 17 protein biomarkers that are closely related to inflammation, immune response, and tissue remodelling processes. We also analyzed the effect of CEO on genome-wide gene expression. The data provide important evidence of the biological activity of CEO in human dermal fibroblasts. The study supports the anti-inflammatory and anticancer properties of CEO, and will likely facilitate the future study of its mechanisms of action, clinical efficacy, and safety.
Materials and methods
All the experiments were conducted using a Biologically Multiplexed Activity Profiling (BioMAP) human dermal fibroblast system HDF3CGF (Kunkel et al. Citation2004a, Citation2004b; Berg et al. Citation2010), which was designed to model the pathology of chronic inflammation in a robust and reproducible manner. The system comprises three components: a cell type, stimuli to create the disease environment, and a set of biomarker (protein) readouts to examine how the treatments affected the disease environment (Berg et al. Citation2010). The methodologies used in this study were essentially the same as those previously described (Kunkel et al. Citation2004a, Citation2004b; Han & Parker Citation2017a, Citation2017b).
Reagents
CEO (dōTERRA Intl., Pleasant Grove, UT) was diluted in dimethyl sulfoxide (DMSO) to 8 × the specified concentrations (final DMSO concentration in culture media was no more than 0.1% [v/v]). Then, 25 μL of each 8 × solution was added to the cell culture to obtain a final volume of 200 μL; DMSO (0.1%) served as the vehicle control. Chemical analysis of CEO by gas chromatography-mass spectrometry indicated that its major chemical constitutes (i.e., > 5%) are eugenol (81%), eugenol acetate (10%), and trans-caryophyllene (7%).
Cell cultures
Primary human neonatal fibroblasts were prepared as previously described (Bergamini et al. Citation2012) and were plated under low serum conditions for 24 h before stimulation with a mixture of interleukin (IL)-1β, tumour necrosis factor (TNF)-α, interferon (IFN)-γ, basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), and platelet-derived growth factor (PDGF). The HDF3CGF assays were performed in a 96-well plate and cell culture and stimulation conditions have been described in detail elsewhere (Bergamini et al. Citation2012).
Protein-based readouts
An enzyme-linked immunosorbent assay (ELISA) was used to measure the biomarker levels of cell-associated and cell membrane targets. Soluble factors in the supernatants were quantified using either homogeneous time-resolved fluorescence detection, bead-based multiplex immunoassay, or capture ELISA. The adverse effects of the test agents on cell proliferation and viability (cytotoxicity) were measured using the sulforhodamine B (SRB) assay. For proliferation assays, the cells were cultured and measured after 72 h, which is optimal for the HDF3CGF system; the detailed procedure was described by Bergamini et al. (Citation2012). Measurements were performed in triplicate wells, and a glossary of the biomarkers used in this study is provided in Supplementary Table S1.
Quantitative biomarker data are presented as the mean log10 relative expression level (compared to the respective mean vehicle control value) ± standard deviation (SD) of triplicate measurements. Differences in biomarker levels between CEO- and vehicle-treated cultures were tested for significance with the unpaired Student’s t-test. A p-value <0.05, outside of the significance envelope, with an effect size of at least 10% (more than 0.05 log10 ratio units), was regarded as statistically significant.
RNA isolation
Total RNA was isolated from cell lysates using the Zymo Quick-RNA MiniPrep kit (Zymo Research Corp., Irvine, CA) according to the manufacturer’s instructions. RNA concentration was determined using a NanoDrop ND-2000 system (Thermo Fisher Scientific, Waltham, MA). The RNA quality was assessed using a Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA) and an Agilent RNA 6000 Nano kit. All samples had an A260/A280 ratio between 1.9 and 2.1 and an RNA integrity score >8.0.
Microarray analysis of genome-wide gene expression
The effect of 0.011% CEO on the expression of 21,224 genes was evaluated in the HDF3CGF system after treatment for 24 h. Samples for microarray analysis were processed by Asuragen, Inc. (Austin, TX) according to the company’s standard operating procedures. Biotin-labelled cRNA was prepared from 200 ng of total RNA using an Illumina TotalPrep RNA Amplification kit (Thermo Fisher Scientific, Waltham, MA) and one round of amplification. The cRNA yields were quantified using ultraviolet spectrophotometry, and the distribution of the transcript sizes was assessed using the Agilent Bioanalyzer 2100. Labelled cRNA (750 ng) was used to probe Illumina human HT-12 v4 expression bead chips (Illumina, Inc., San Diego, CA). Hybridization, washing, staining with streptavidin-conjugated cyanine-3, and scanning of the Illumina arrays were carried out according to the manufacturer’s instructions. The Illumina BeadScan software was used to produce the data files for each array; the raw data were extracted using Illumina BeadStudio software.
The raw data were uploaded into R (R Development Core Team Citation2011) and analyzed for quality-control metrics using the beadarray package (Dunning et al. Citation2007). The data were normalized using quantile normalization (Bolstad et al. Citation2003), and then re-annotated and filtered to remove probes that were nonspecific or mapped to intronic or intragenic regions (Barbosa-Morais et al. Citation2010). The remaining probe sets comprised the data set for the remainder of the analysis. The fold-change expression for each set was calculated as the log2 ratio of CEO to the vehicle control. These fold-change values were uploaded onto Ingenuity Pathway Analysis (IPA, Qiagen, Redwood City, CA, www.qiagen.com/ingenuity) to generate the networks and pathway analyses.
Results and discussion
Bioactivity profile of CEO in pre-inflamed human dermal fibroblasts
We studied the activity of CEO in a dermal fibroblast system, HDF3CGF, which simulates the disease microenvironment of inflamed human skin cells. None of the four investigated CEO concentrations (0.011, 0.0037, 0.0012, and 0.00041%, v/v) was overly cytotoxic; hence, all were used for further analysis. Key biomarker activities were designated if biomarker values were significantly different (p < 0.05) from vehicle controls at the 0.011% concentration with an effect size of at least 10% (more than 0.05 log ratio units, ).
Figure 1. Bioactivity profile of clove essential oil (CEO, 0.011% v/v) in human dermal fibroblast culture HDF3CGF. X-axis denotes protein-based biomarker readouts. Y-axis denotes the relative expression levels of biomarkers compared to vehicle control values, in log10 form. Vehicle control values are shaded in grey, denoting the 95% significance envelope. Error bars represent the standard deviations from triplicate measurements. A * indicates a biomarker designated with ‘key activity,’ i.e., biomarker values were significantly different (p < 0.05) from vehicle controls, outside of the significance envelope, with an effect size of at least 10% (more than 0.05 log ratio units). MCP-1, monocyte chemoattractant protein; VCAM-1, vascular cell adhesion molecule 1; ICAM-1, intracellular cell adhesion molecule 1; IP-10, interferon γ-induced protein 10; I-TAC, interferon-inducible T-cell α chemoattractant; IL-8, interleukin-8; MIG, monokine induced by γ interferon; EGFR, epidermal growth factor receptor; M-CSF, macrophage colony-stimulating factor; MMP-1, matrix metalloproteinase 1; PAI-1, plasminogen activator inhibitor 1; TIMP, tissue inhibitor of metalloproteinase.
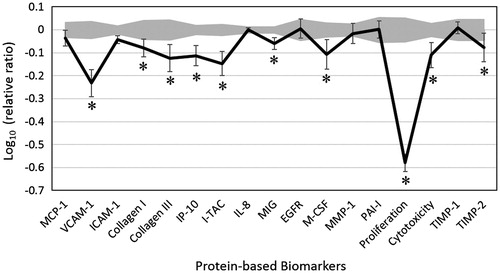
Overall, CEO inhibited many of these 17 important biomarkers. CEO showed significant antiproliferative activity in human dermal fibroblasts. CEO also significantly decreased the levels of inflammatory biomarkers such as vascular cell adhesion molecule-1 (VCAM-1), interferon gamma-induced protein 10 (IP-10), interferon-inducible T-cell α chemoattractant (I-TAC), and monokine induced by γ interferon (MIG). Furthermore, it significantly inhibited tissue remodelling protein molecules, namely, collagen I, collagen III, macrophage colony-stimulating factor (M-CSF), and tissue inhibitor of metalloproteinase 2 (TIMP-2). The effect of CEO on these biomarkers appeared to be concentration-dependent. The significant inhibitory effects exerted by CEO on these biomarkers indicate that CEO may possess anti-inflammatory and pro-wound-healing properties.
A number of studies have reported the anti-inflammatory properties of CEO and its major active component eugenol. Both CEO and eugenol exhibited anti-inflammatory activities in murine macrophages, inhibiting the production of pro-inflammatory cytokines (Rodrigues et al. Citation2009; Bachiega et al. Citation2012). The current study provides further support for the anti-inflammatory properties of CEO and eugenol.
Interestingly, Koh et al. (Citation2013) found that eugenol inhibited increased IL-8 production in human gingival fibroblasts (HGF), but not in periodontal ligament fibroblasts (HPLF) or skin keratinocytes (HaCat) (Koh et al. Citation2013). Our study demonstrated that CEO significantly inhibited many pro-inflammatory cytokines in pre-inflamed human dermal fibroblast cells, but did not significantly affect IL-8 levels. Further research is needed to evaluate the biological mechanism underlying the effect of CEO in different human cells.
Effects of CEO on genome-wide gene expression
To further explore the biological activities of CEO in human skin cells, we studied the effect of 0.011% CEO (the highest tested non-cytotoxic concentration) on the RNA expression of 21,224 genes in the HDF3CGF system. The results showed that CEO exerted robust and diverse effects on gene regulation, with many genes being downregulated and others being upregulated. Of the 200 most affected genes (log2 [expression fold-change ratio relative to vehicle control] ≥ |1.5|), the majority (142 out of 200 genes) were significantly downregulated, and the rest were upregulated (Table S2). A cross comparison of protein and gene expression data revealed that CEO significantly inhibited collagen III and VCAM-1 at both protein and gene expression levels.
IPA studies showed that CEO bioactivity significantly matched that of many canonical signalling pathways from the literature-validated database (). Many of these pathways are involved in inflammation, tissue remodelling, stress response, cell cycle regulation, cancer signalling, or cellular metabolism. This indicates that CEO may play an important role in a variety of biological and physiological processes. For example, the top matched pathway was hepatic fibrosis activation, and this was inhibited by CEO, further supporting its anti-inflammatory properties.
Figure 2. Top 20 canonical pathways matching the bioactivity profile of clove essential oil (CEO, 0.011% v/v) in gene expression in the HDF3CGF system produced using Ingenuity Pathway Analysis (IPA, Qiagen, www.qiagen.com/ingenuity). Each p-value was calculated using right-tailed Fisher's exact test. The p-value measures the likelihood that the observed association between a specific pathway and the dataset is due to random chance. The smaller p-value (the bigger - ln [p-value], indicated by the black bars) the pathway has, the more significantly it matches the bioactivity of CEO. A ratio, indicated by each grey bar, was calculated by taking the number of genes from the CEO dataset that participate in a canonical pathway, and dividing it by the total number of genes in that pathway. GADD45: growth arrest and DNA-damage-inducible protein 45.
![Figure 2. Top 20 canonical pathways matching the bioactivity profile of clove essential oil (CEO, 0.011% v/v) in gene expression in the HDF3CGF system produced using Ingenuity Pathway Analysis (IPA, Qiagen, www.qiagen.com/ingenuity). Each p-value was calculated using right-tailed Fisher's exact test. The p-value measures the likelihood that the observed association between a specific pathway and the dataset is due to random chance. The smaller p-value (the bigger - ln [p-value], indicated by the black bars) the pathway has, the more significantly it matches the bioactivity of CEO. A ratio, indicated by each grey bar, was calculated by taking the number of genes from the CEO dataset that participate in a canonical pathway, and dividing it by the total number of genes in that pathway. GADD45: growth arrest and DNA-damage-inducible protein 45.](/cms/asset/066ba2b7-ba1d-4660-8511-46832ffc2da0/iphb_a_1314513_f0002_b.jpg)
In addition to their anti-inflammatory properties, CEO and eugenol have been shown to possess anticancer properties against breast, colorectal, lung, and leukaemia cancer cells (Yoo et al. Citation2005; Kouidhi et al. Citation2010; Kumar et al. Citation2014). These previous findings are largely consistent with the findings of the current study, which indicate that CEO affected cell cycle control and cancer biology.
Conclusions
Taken together, the results of this study demonstrated the anti-inflammatory, immune-modulating, and tissue remodelling activities of CEO in a human skin disease model. The microarray study also showed evidence suggesting the role of CEO in modulating important signalling pathways related to immune function, cell cycle control, cellular stress responses, and even cancer biology. These data largely support the anti-inflammatory and anticancer properties of CEO and its major active component eugenol.
Xuesheng_Han_and_Tory_Parker_supplemental_content.zip
Download Zip (572.2 KB)Disclosure statement
X.H. and T.P. are employees of dōTERRA, where the study agent CEO was manufactured.
Additional information
Funding
References
- Bachiega TF, de Sousa JPB, Bastos JK, Sforcin JM. 2012. Clove and eugenol in noncytotoxic concentrations exert immunomodulatory/anti-inflammatory action on cytokine production by murine macrophages. J Pharm Pharmacol. 64:610–616.
- Barbosa-Morais NL, Dunning MJ, Samarajiwa SA, Darot JFJ, Ritchie ME, Lynch AG, Tavaré S. 2010. A re-annotation pipeline for Illumina BeadArrays: improving the interpretation of gene expression data. Nucleic Acids Res. 38:e17.
- Berg EL, Yang J, Melrose J, Nguyen D, Privat S, Rosler E, Kunkel EJ, Ekins S. 2010. Chemical target and pathway toxicity mechanisms defined in primary human cell systems. J Pharmacol Toxicol Methods. 61:3–15.
- Bergamini G, Bell K, Shimamura S, Werner T, Cansfield A, Müller K, Perrin J, Rau C, Ellard K, Hopf C, et al. 2012. A selective inhibitor reveals PI3Kγ dependence of T(H)17 cell differentiation. Nat Chem Biol. 8:576–582.
- Bolstad BM, Irizarry RA, Astrand M, Speed TP. 2003. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinforma Oxf Engl. 19:185–193.
- Dunning MJ, Smith ML, Ritchie ME, Tavaré S. 2007. Beadarray: R classes and methods for Illumina bead-based data. Bioinforma Oxf Engl. 23:2183–2184.
- Han X, Parker TL. 2017a. Biological activity of vetiver (Vetiveria zizanioides) essential oil in human dermal fibroblasts. Cogent Med. 4:1298176.
- Han X, Parker TL. 2017b. Anti-inflammatory, tissue remodeling, immunomodulatory, and anticancer activities of oregano (Origanum vulgare) essential oil in a human skin disease model. Biochimie Open. 4:73–77.
- Koh T, Murakami Y, Tanaka S, Machino M, Sakagami H. 2013. Re-evaluation of anti-inflammatory potential of eugenol in IL-1β-stimulated gingival fibroblast and pulp cells. In Vivo. 27:269–273.
- Kouidhi B, Zmantar T, Bakhrouf A. 2010. Anticariogenic and cytotoxic activity of clove essential oil (Eugenia caryophyllata) against a large number of oral pathogens. Ann Microbiol. 60:599–604.
- Kumar PS, Febriyanti RM, Sofyan FF, Luftimas DE, Abdulah R. 2014. Anticancer potential of Syzygium aromaticum L. in MCF-7 human breast cancer cell lines. Pharmacogn Res. 6:350–354.
- Kunkel EJ, Dea M, Ebens A, Hytopoulos E, Melrose J, Nguyen D, Ota KS, Plavec I, Wang Y, Watson SR, et al. 2004a. An integrative biology approach for analysis of drug action in models of human vascular inflammation. FASEB J. 18:1279–1281.
- Kunkel EJ, Plavec I, Nguyen D, Melrose J, Rosler ES, Kao LT, Wang Y, Hytopoulos E, Bishop AC, Bateman R, et al. 2004b. Rapid structure-activity and selectivity analysis of kinase inhibitors by BioMAP analysis in complex human primary cell-based models. Assay Drug Dev Technol. 2:431–441.
- Prashar A, Locke IC, Evans CS. 2006. Cytotoxicity of clove (Syzygium aromaticum) oil and its major components to human skin cells. Cell Prolif 39:241–248.
- R Development Core Team. 2011. R: A Language and Environment for Statistical Computing. Vienna, Austria: The R Foundation for Statistical Computing.
- Rodrigues TG, Fernandes A, Sousa JPB, Bastos JK, Sforcin JM. 2009. In vitro and in vivo effects of clove on pro-inflammatory cytokines production by macrophages. Nat Prod Res. 23:319–326.
- Yoo C-B, Han K-T, Cho K-S, Ha J, Park H-J, Nam J-H, Kil UH, Lee KT. 2005. Eugenol isolated from the essential oil of Eugenia caryophyllata induces a reactive oxygen species-mediated apoptosis in HL-60 human promyelocytic leukemia cells. Cancer Lett. 225:41–52.