Abstract
A synopsis is given of the knowledge of the genus Rhinagrion. The males of two new species are described from the Philippines: R. schneideri sp. nov. (holotype: Samar Island, Hinubangan, San Isidro, 31 March–5 April 1992) and R. reinhardi sp. nov. (holotype: Mindanao Island, Surigao del Sur, Carmen, 24 April 1995). Rhinagrion yokoii is synonymized with R. hainanense and R. viridatum is removed from synonymy with R. mima. A key to the males is given and the distribution of all species is discussed and maps are provided. The scant information available on behaviour and habitat is summarized.
Introduction
Rhinagrion Calvert, 1913 is a small Oriental genus of very colourful damselflies currently placed in the family Megapodagrionidae. The 10 species of Rhinagrion occur in mainland South-east Asia, Sundaland and the Philippines () where they inhabit brooks and rivers in lowland rainforest. Males are typically found perched at a height of one or two metres on the vegetation along brooks; they are far more often encountered than females. All species have extensive and often bright patterns on face, thorax and abdomen. In many species the dorsum of the tip of the abdomen is brightly coloured whilst sternites 8 and 9 are often bright white, violet or red. This colourful appearance suggests that the species may have extensive territorial and mating behaviour; however observations on behaviour are scarce.
Table 1. Species included in Rhinagrion and their distribution
When we studied a large amount of material of Rhinagrion present in the RMNH we discovered two undescribed species of Rhinagrion “hidden” under a known species. These species show relatively few distinct structural characters, but can be distinguished from their congeners based on clear differences in pattern and coloration. The males of these new species are described in the present article, the characters of the males of others species are evaluated and, based on a wealth of new material, the distributions of the species are shown.
Material and methods
Males of Rhinagrion are far more often encountered than females and are easier to recognize, for which reason this study focuses on the males. Terminology largely follows Watson and O'Farrell Citation(1991). All material is in RMNH unless otherwise noted.
Acronyms for collections:
Recognition of Rhinagrion
The males of the genus Rhinagrion are among the most striking and colourful damselflies. In the field they are easily recognized by having a colourful abdomen with red, blue, orange or pink markings and wings that are hyaline and open at rest. The two species of Philosina Ris, 1917 are likely to be their nearest relatives (Kalkman et al. Citation2010, Zhang et al. Citation2011). The adult males of these species are however largely covered with whitish or blue pruinosity which is never the case in Rhinagrion. The only other damselflies that hold their wings open in the area where Rhinagrion occurs are the species of the family Lestidae and, in the Philippines, Argiolestes Selys, Citation1862, also in the Megapodagrionidae. Both of these have however a duller, less patterned abdomen. Specimens can be distinguished from all other Zygoptera except those of Philosina based on the combination of the following three characters: (1) two antenodal crossveins; (2) Pt at least twice as long as broad; (3) median lobe of labium long with a deep median cleft which is about half the length of the labium itself.
Rhinagrion borneense Selys, 1886 (Figures 2, 3d)
Specimens studied
Brunei 2 ♂: surroundings Ingai base camp, along Ingai river, 04°09′ 21 N, 114°42′ 56
E, 14–17 September 1992, leg. D. Thompson. Indonesia, Kalimantan 174 ♂, 17 ♀: Tabang, Bengen River, 125 m, 31 August–1 November 1956, leg. A.M.R. Wegner. — 1 ♂, Tabang, Bengen River, 125 m, 6 September 1956, leg. A.M.R. Wegner, ZSMC. — 8 ♂: Gunung Sari, 95 m, 14–17 August 1956, leg. A.M. Wegener. — 5 ♂, 1 ♀: SE Borneo, S. Mentawir, 2–9 October 1950, leg. unknown. Malaysia, Sarawak ♂: Lio Matu, 1 November 1914, leg. J.C. Moulton. — 1 ♂: Sekerang (Skrang) River, Bunu Longhouse, 01°19.2′ N 111°39.0′ E, 10 July 1997, leg. T. Donnelly. — 1 ♂: Lambir Hills National Park, headquarter area, 5–9 August 1995, leg. H.S. Yong.
Differential diagnosis of male
Face entirely black. Pale marks on dorsum of synthorax large and narrowly divided by middorsal carina. Abdomen largely red with S8–9 blue (rarely small blue dots on the distal part of S7), S10 and appendages black. Sternum of S8 violet in life but can be largely dark reddish to blackish in dead specimens, sternum of S9 white.
Remarks
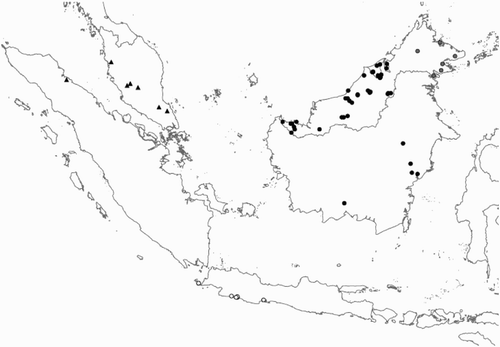
The type of Rhinagrion borneense is said to come from the island of Labuan in the north-west of Sabah (Selys, Citation1886). However it is likely that this was not the actual locality where the specimen was collected, but merely the port from where the material was shipped. The species seems to be fairly common in parts of Borneo () with many records from Brunei (Orr, Citation2001; Thompson, Citation1998; Thompson & van Tol, Citation1993), Sarawak (Donnelly, Citation1994; Dow, Citation2005, Citation2008; Hincks, Citation1930; Kimmins, Citation1936; Laidlaw, Citation1920; Matsuki & Kitagawa, Citation1993; Dow, pers. comm.) and fewer records from Kalimantan (Cleary et al., Citation2004; Lieftinck, Citation1953; Dow, pers. comm.). The scarcity of records from most of Kalimantan may be due to the lesser amount of fieldwork that has been conducted there.
Figure 1. SEM photographs of the genital ligula of: (a) Rhinagrion mima, Thailand, Kanchanaburi, Erawan Falls, 1999, ventral view; (b) same, lateral view; (c) Rhinagrion macrocephalum, Malaysia, Krau Wildlife Reserve, 1995, lateroventral view; (d) R. tricolor, Indonesia, Java, Udjung Kulon, 1958, lateroventral view; (e) R. philippinum, Luzon, Philippines, Apayao Province, Flora, Santa Maria, 1997, lateroventral view; (f) R. schneideri, Philippines, Samar, Las Navas, San Isidro, 1997, lateroventral view. Photos: Dirk Gassmann.
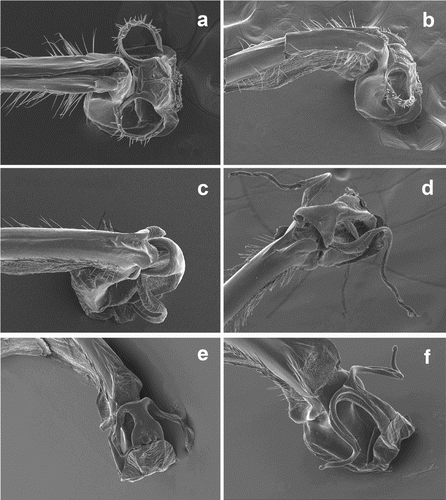
Figure 2. Distribution of the species of the Rhinagrion borneense group based on material from this article and hitherto published records. For Rhinagrion borneense unpublished records from Sarawak made available by R. Dow are also included. Black dots: R. borneense; grey dots on Borneo: R. elopurae; triangles: R. macrocephalum; open circles: R. tricolor.
Lieftinck Citation(1954) recorded the species from sluggish forest brooks in low country and mentioned that larvae were found “concealed among decaying vegetable matter, in pools under the bank of a stream”. The second author collected larvae of R. borneense at a small river by sifting coarse sand. Dow (pers. comm.) has collected and/or observed this species on a large variety of low gradient forest streams in Sarawak, including a stream in an acacia plantation and streams at c.900 m near Long Banga in the upper Baram area and at over 1000 m on Mt Dulit; however, most records are from the lowlands.
Lieftinck Citation(1956) and Paulson Citation(1981) noted that Rhinagrion borneense is capable of broadening the tip of the abdomen. Lieftinck Citation(1956) states that “While it sat motionless, sunning itself on a prominent Pandanus leaf, the blue tail-end of the abdomen was distended all of a sudden and then curved slightly upwards, the insect holding up this attitude for many minutes so that the bright blue colour was prominently displayed, the wings meanwhile being held motionless in a horizontal position”. Males engage in territorial contests, flying facing each other and holding the end of the abdomen flexed upwards or downwards, displaying the blue dorsal colour and the violet colour of the venter (Dow, pers. comm.). A female was observed apparently ovipositing into a mud bank beside a pebble bottomed stream at Gunung Mulu National Park in Sarawak. Two males were perched within 2 m of this female, but no interaction was observed between the males and the female, or between the males.
Rhinagrion elopurae McLachlan in Selys, 1886 (Figures 2, 3e)
Specimens studied
Malaysia, Sabah1 ♂ (holotype): North Borneo (BMNH). — 23 ♂, 6 ♀: 60 km W of Lahad Datu, Danum Valley Field Centre, near Sungai Segama, 160–180 m, circa 4°58′ N 117°48′ E, 14 March–1 May 1987, leg. J. van Tol & C. Achterberg. — 2 ♂: Danum Valley, Borneo rainforest lodge, stream and waterfall, 500 m NE of the lodge, 200 m, 26 April 1994, leg. M. Hämäläinen. — 1 ♂: 3–4 km WSW of Quoin Hill Tawau, 9–20 July 1962, leg. Y. Hiroshima. — 2 ♂: Danum Valley Field Centre, Sungai Palum Tambun, 150–160 m, 23–28 April 1994, leg. M. Hämäläinen. — 6 ♂: Sepilok Orang Utan Sanctuary, 9 November 1992, leg. D. Paulson.
Additional records
The following records are based on pictures appearing on www.asia-dragonfly.net and have not previously been published.
Malaysia, Sabah 1 ♂: Danum Valley, 9 October 2004, by P. Benstead. — 1 ♂: Danum Valley, 7 October 2008, leg. P. Benstead. — 1 ♂: Tabin Wildlife Reserve, Lahad Datu, 8 July 2007, by J. Pan. — 1 ♂: Madai Waterfall, Kunak, 8 October 2006, by J. Pan. — 1 ♂: Sepilok Orang Utan Sanctuary, 27 January 2010, by T. Termaat. Indonesia, Kalimantan 1 ♂: Kalimantan Timur, Nunukan, Binusan Forest Reserve, 11 July 2010, by J. Pan.
Differential diagnosis of male
Face with extensive pale (blue) pattern. Pale marks on dorsum of synthorax large and narrowly divided by middorsal carina. Abdomen largely red with S8–9 and distal fourth of S7 blue, remainder of S7 and distal part of S6 black, S10 and superior appendages black. Sternum of S8 blackish to reddish pink, sternum of S9 white.
Remarks
The holotype was collected prior to 1886 with the locality “North Borneo”. The only record published since is from Danum Valley, Sabah, Malaysia (Donnelly, Citation1999). Based on at that time unpublished material Orr Citation(2003) was able to state that the species “occurs only on small forest streams in the northeast” of Borneo. The photographic record from Binusan Forest Reserve made by J. Pan is the first record of this species for Indonesia. The new records published here show that R. elopurae has a restricted range and might only occur in Sabah and adjacent parts of Kalimantan (). R. elopurae and R. borneense probably have allopatric ranges; it is unlikely that the range of R. elopurae extends much further south than is currently known. Its western limit is not yet clear, but it probably does not reach Brunei.
Rhinagrion hainanense Wilson & Reels, 2001 (Figures 3a, 4)
Specimens studied
China1 ♂: Guizhou province, Maolan Reserve, 25–28 July 2008, leg. H. Zhang. — 1 ♀: Hainan Island, Tasi-po, Lin-kao district, 25–27 September 1932, leg. F.K. To. Vietnam1 ♂: Dak lak province, Chu Yang Sin National Park, 12°26′ 48 N 108°20′ 10
E, 760 m, 8 June 2007, leg. J. van Tol. — 1 ♂: Thua Thien Hue province, Phong Dien Nature Reserve, 40 km W of Hue, Khe Cá Lóc stream and Ô Lâu river, 60 m a.s.l., 16°31′ 15
N 107°12′45
E, 60 m, 22 March–7 April 2001, leg. J. van Tol. — 1 ♂: Thua Thien Hue province, Phong Dien Nature Reserve, 40 km W of Hue, tributary of Ô Lâu river upstream of KCL camp, 60 m a.s.l., through disturbed forest, 16°31′ 15
N 107°12′ 45
E, 60 m, 22 March–7 April 2001, leg. J. van Tol. — 1 ♂: Khanh Hoa Province, Ba Ho falls, small side stream and main stream below first fall, 12°23′ 22
N 109°08′ 14
E, 25 m, 9 May 2009, leg. M. Hämäläinen.
Figure 3. Males of Rhinagrion: (a) R. hainanense; (b) R. mima, Sumatra; (c) R. viridatum; (d) R. borneense; (e) R. elopurae.

Differential diagnosis of male
Head black with extensive yellow-blue pattern. Antehumeral stripe roughly parallel sided, covering about two-thirds length of synthorax. Side of synthorax with dark stripe along metapleural suture not connected to dark dorsum of synthorax. Abdomen largely yellow-brown to brown, S3–7 with pale band on posterior half of each segment, S8–10 black with pairs of blue dots on S9–10, superior appendages pale. Sternum of S8–9 yellow-brown to reddish.
Remarks
Sasamoto (2003) described R. yokoii from Sekong in south-eastern Laos. In the description the species was compared with R. mima but not with R. hainanense, which was described two years earlier (Wilson & Reels, Citation2001). Based on the original descriptions, photos of R. hainanense from Hainan and the above listed material we concluded that R. yokoii is a junior synonym of R. hainanense. The only clear differences between the material from Hainan and from mainland Asia is that the front of the frons in yokii is black while in hainanense it is pale, thus connecting the lateral pale spots. This difference was deemed too small to warrant species status to R. yokoii. The specimen from Guizhou and the male in a photo from Hainan have brown ground colour of the abdomen while the ground colour in the specimens from Vietnam and Laos is yellow-brown. The amount of material available is too little to tell if this difference in colour can be attributed to the age of the specimens. We studied the genital ligula under the microscope only and no SEM image was made. The genital ligula is most like that of R. mima, sharing the abundant, long hairs on the shaft and the numerous spines on the lateral horns. It differs, however, from R. mima in the lateral horns being longer and the space between them being less broad and rounded (square in R. mima, ).
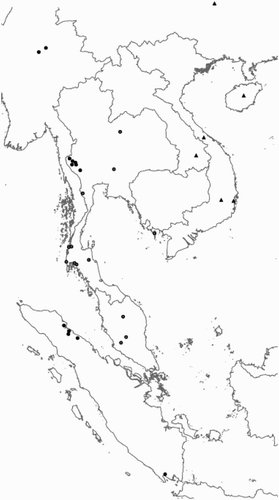
R. hainanense is now known from the Chinese provinces of Hainan (Wilson & Reels, Citation2001) and Guizhou, Laos (Sasomoto, Citation2003) and Vietnam (). The locality at Chu Yang Sin National Park in Vietnam was given as a “sandy forest stream” and that at Phong Dien Nature Reserve as “2 m wide and 20 cm deep through disturbed forest”. In Hainan the species is found in dense vegetation beside shady, slow-flowing streams in lowland areas. They perch low above the ground – no higher than a metre – and both sexes make short flights on the edge of, or within, dense shrubby vegetation along the stream bank and were never seen flying over the open stream (G. Reels, pers. comm.). A female Rhinagrion sp. from Phanom Dongrak mountain range in Si Sa Ket in East Thailand is present in the CUMZ (Hämäläinen & Pinratana, Citation1999). This record falls between the known area of distribution of R. hainanese and R. viridatum and could belong to either species.
Figure 4. Distribution of the species of the Rhinagrion mima group based on material from this article and hitherto published records. For Thailand, where both R. mima and R. viridatum can occur, only records based on material studied for this article are used. Triangles: R. hainanense; grey dots: R. mima; black dots: R. viridatum.
Rhinagrion macrocephalum Selys, 1862 (Figures 1c, 2, 6b)
Specimens studied
All leg. M. Hämäläinen unless otherwise noted.
Peninsular Malaysia 1 ♂: Kelansan, East Malacca, 1903, leg. Heijne. — 1 ♂: Selangor, Kerling, Bukit Tarek, Sungai Jerneh, 5–8 June 1997. — 1 ♂: Pahang, Krau Wildlife Reserve, Kuala Gandah, Sungai Gandah, 25 January 1996. — 1 ♀: Pahang, Krau Wildlife Reserve, Kuala Gandah, Sungai Gandah, 22 April 1995. — 2 ♂, 1 ♀: idem, 18 November 1996, leg. M Hämäläinen & H. Olsvik. — 1 ♂: Pahang, Krau Wildlife Reserve, Bukit Rengit, Sungai Rengit, 23 April 1995. — 1 ♀: idem, 21 January 1996.
Differential diagnosis of male
Face with blue pattern. Blue spots on dorsum of synthorax small and broadly divided by middorsal carina. Segments 3–7 largely red, S8–9 blue, S10 and superior appendages black. Sternum of S8 white to pink, sternum of S9 white.
Remarks
R. macrocephalum is known from north and north-east Sumatra (Krüger, Citation1898; Lieftinck, Citation1935) and from Peninsular Malaysia (Donnelly, Citation1994; Kemp & Kemp, Citation1989; Selys Citation1862, Vick, Citation1993; Wilson & Gilbert, 2005). No records of Rhinagrion macrocephalum are known from the south of Sumatra.
Rhinagrion mima Karsch, 1891 (Figures 1a, b, 3b, 4)
Specimens studied
All leg. M. Hämäläinen unless otherwise noted.
Burma 1 ♂, 1 ♀: Upper Burma, Gok Teik, ex coll Fraser, leg. unknown. — 1 ♀: Upper Burma, Maymyo, ex coll Fraser, leg. unknown. — 1 ♂: Upper Burma, Maymyo fluss, 1200 m, 20 August 1977, leg. G. von Rosen, ZSMC. Indonesia, Sumatra 1 ♂: S. Sumatra, Mount Tanggamoes, 19–31 March 1940, leg. M.A. Lieftinck. — 2 ♂: NW Sumatra, W. Atjeh, Seumanjang Estate, 25 December 1953, leg. R. Straatman. — 5 ♂, 1 ♀: NE Sumatra, Laut Tador, 50–90 m, April–May 1948, leg. R. Straatman. — 3 ♂: NE Sumatra, Atjeh, Kuala Simpang, June 1953, leg. A. Sollaart. — 4 ♂: NE Sumatra, Seleleh, Medang Area, 24 May 1953, leg. R. Straatman. — 3 ♂: NE Sumatra, Tandjong Morawa, Serdang, no date, leg. B. Hagen. Thailand 2 ♂: Kanchanaburi, Kroeng Kra Via, downstream Pu Ye and hidden sidestream, 210–330 m, 5 April 2001. — 1 ♂: Kanchanaburi, Kroeng Kra Via National Park, Pu Ye, forest stream, 2 May 2000. — 1 ♂: Kanchanaburi, Huai Mae Kamin, 5 January 1993. — 1 ♂: Kanchanaburi, Sangkhlaburi, streams on way to Ta Khian Thong waterfall, 200 m, 1 November 2000. — 1 ♂: Kanchanaburi, Nang Kroan, 600 m, 4 April 2001. — 2 ♂: Kanchanaburi, Thong Pha Phum area, Nam Tok Nang Kroan, 20 October 1999. — 1 ♀: Kanchanaburi, Erawan Falls, 17–18 October 1982. — 1 ♂: Kanchanaburi, Lam Klong Ngu, 8–9 May 1999.
Differential diagnosis of male
Face black with extensive yellow pattern. Antehumeral stripe parallel sided and extending about two-thirds length of synthorax. Side of synthorax with dark stripe along metapleural suture not connected to dark dorsum of synthorax. S3–5 black with a distinct blue apical spot, S7–9 largely yellow, superior appendages black with the apical third to half yellow-orange. Sterna of S8–9 brown to black.
Remarks
Rhinagrion mima was decribed by Karsch (1891) based on one male from Bindjei (Deli) from the Indonesian island of Sumatra. A redescription of the species was given by Fraser Citation(1933), probably based on material from Maymyo, Upper Burma. Fraser Citation(1938) described R. viridatum from the south of Burma. Without seeing the type Lieftinck Citation(1954) synomized R. viridatum with R. mima as the variation seemed to “fall within the limits of individual variation”. Ever since the two have been regarded as synonyms, although with the large amount of new material becoming available from Peninsular Malaysia and Thailand, it is not difficult to see that they are distinct in pattern and coloration. There are some minor differences between the material of R. mima from Sumatra and that of Thailand and Burma: those from Sumatra are slightly smaller and the pale spots on S3–4 are slightly more distinct. In addition to this the distribution of R. mima is strange; it occurs in two areas broadly separated by an area where R. viridatum occurs (). There is therefore the possibility that R. mima is not monophyletic and that it should be divided into two species, each of which is more closely related to R. viridatum than to the other. There is however no proof of this based on the morphology and coloration; DNA studies are needed to resolve this issue. See R. viridatum for further discussion on the distribution of R. mima and R. viridatum.
Rhinagrion reinhardi sp nov. (Figures 5, 6c)
Figure 5. Distribution of the species of the Rhinagrion philippinum group based on material from this article and hitherto published records. The holotype of R. philippinum comes from Bohol. Details on the location are lacking so that it is not present on the map. Black dots: R. reinhardi; grey dots: R. philippinum; triangles: R. schneideri.
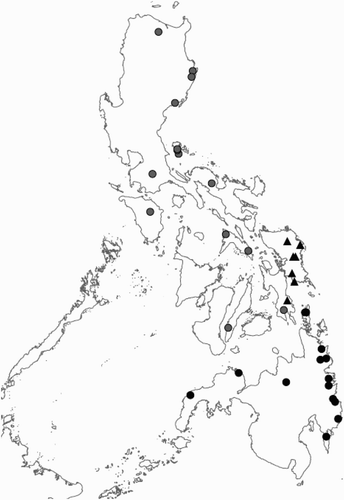
Figure 6. Males of Rhinagrion: (a) R. tricolor; (b) R. macrocephalum; (c) R. reinhardi sp. nov., holotype; (d) R. philippinum; (e) R. schneideri sp. nov., holotype.

Etymology
Noun in the genitive case, named in honour of Reinhard Jödicke for his dedication to the International Journal of Odonatology during his 10-year editorship (2001–2010).
Specimens studied
Holotype ♂: (white printed label) “Philippines, Mindanao Island, Surigao del Sur, Carmen, Upper Tandag River, km 9 Lanang Line, 500 m, 24 April 1995, leg. Buenafe & Gorostiza, coll. Roland A. Müller”. The holotype is deposited in the RMNH.
Additional material, all leg. RJTV unless otherwise noted. Philippines, Mindanao2 ♂: Surigao del Sur, Tago, Meme River, 100–300 m, 12–18 June 1996, leg. A. Buenafe & A. Gorostiza. — 1 ♂: Surigao del Sur, San Miguel, Suba River, 150–250 m, 13–15 April 1995, leg. Müller, Buenafe & Gorostiza. — 1 ♂: Surigao, 27 May 1915, leg. G. Boettcher (SMF). — 1 ♂: Mindanao Id, Bukidnon, Impasub-ong, coll. C. Demayo, April 2007 (MSU-IIT). — 1 ♀: Surigao del Sur, Tagbina, RJTV leg., 23 May 2009 (RJTV). — 1 ♂: Surigao del Sur, Bislig, Tinuy-an Falls, RJTV leg., 24 May 2009 (RJTV). — 1 ♀: Davao Oriental, San Isidro, La Union, Magumilo River, leg. H. Cahilog, 3 July 2008 (RJTV). — 1 ♂: idem, 6 July 2008 (RJTV). — 1 ♂: idem, 13 July 2008 (RJTV). — 1 ♀: idem, 25 September 2008 (RJTV). — 1 ♂: Davao Oriental, Mahu stream, Maglahus, Cateel, N,
E, 3 July 2010, leg. HC, (RJTV). — 1 ♂, 1: Davao Oriental, Uwangan stream, Aliwagwag, Cateel,
N,
E, 3 July 2010, leg. HC, (RJTV). — 1 ♀: Davao Oriental, Buwawan, Manay,
N,
E, 3 July 2010, leg. HC, (RJTV). — 1 ♂: Missamis Occidental, Siloy, Calamba, 8.583° N,
E, 19 May 2010, leg. Hung Nien, (ChungPing Lin collection, Tunghai University Taiwan). — 1 ♀: Zamboanga del Norte, Bagong Silang, Gulatac, 7.971° N,
E, 27 October 2010, leg. HC, (RJTV) — Philippines, Dinagat 2 ♂: Tubajon, Paragua Forest, 12 January 2008 (RJTV). — 1 ♂: idem, 18 January 2008 (RJTV). — 1 ♂: idem, 4 April 2008 (RJTV). — 1 ♂: idem, 7 December 2008 (RJTV). — 1 ♂: Libjo, Paragua Forest, 4 June 2008 (RJTV). — 1 ♂: idem, 25 May 2008 (RJTV). — 1 ♀: idem, 20 October 2007 (RJTV). — 1 ♂: idem, 14 December 2008 (RJTV). — 1 ♂: idem, 27 January 2009 (RJTV).
Differential diagnosis of male
Face with extensive blue pattern. Sides of synthorax pale brown, dorsum of synthorax black with large blue patch crossing median carina, half oval shaped and extending over c. two-thirds of length of dorsum. S1–5 dull orange with some blue spots, S6–10 black with blue marks covering S7 and distal two-fifths of S6, superior appendages black. Sternum of S8–9 brown to black.
Description of holotype
Head
Labium pale yellow, longer (1.35 mm) than wide (1.12 mm), median cleft about half length of labium, about a quarter as wide as deep. Mandibles pale with tip black; labrum black; genae pale yellow-blue; anteclypeus pale blue with black lateral margins; postclypeus largely pale blue; frons as far up as lateral ocelli blue, blue of the front of face running along eye margin to occiput but interrupted at top of head.
Thorax
Anterior lobe of prothorax short, erect, brownish but medially blackened; median lobe gently domed, black with sides yellow-brown; posterior lobe simple, without modifications, yellow except for bluish basal-medial portion. Dorsum of synthorax black, with large blue patch crossing median carina, in shape of a half oval, occupying c. two-thirds of length of mesepisternum. Upper fourth to third of mesepimeron black, remainder of sides of synthorax yellow. – Coxae, trochanters and femora pale yellow-brown except faint blackish streak on femora; tibiae light brown; spines dark brown. – Wing membrane hyaline, veins black. Fw and Hw of equal length, both with two Ax, Hw with 12–14 and Fw with 14–16 Px. Petiolated to Ax2, Ac near to Ax2, 2–3 cells between discoidal cell and subnodus, intercalated vein at level of Px5 and Px3 in Fw and Hw respectively. Pt dark brown, long, equilateral and slightly broader centrally, spanning two or more cells.
Abdomen
S1 and S2 dull orange with narrow dorsoapical blue streak; S3 dorsally dull orange with apical third black; S4 dark orange dorsally with blackened apical half; S5 blackish dorsally; S6 black with large dorsoapical blue patch occupying two-fifths of segment; S7 dorsally blue and ventrally black; S8–10 black. Ventral side of S3–5 pale on apical fourth to third. Sternum of S8–9 dark brown to black. Appendages black. Superior appendages in lateral view with basal fourth robust, then tapered; in dorsal view basal half cylindrical, apical half flattened interiorly and abruptly constricted close to apex, with short spike on dorsal tip. Inferior appendages very short and hardly visible in lateral view.
No SEM image of the genital ligula was made but it was studied using a microscope. It appears to be identical to that of R. schneideri.
Measurements [mm]
Entire length 48, abdomen 36, Fw 29; Fw pterostigma 1.9 (costal length), 2.3 (longest length); Hw pterostigma 2.1 (costal length), 2.4 (longest length).
Remarks
Records of R. reinhardi have been published under the name of R. philippinum from Mindanao (Hämäläinen & Müller, Citation1997) and Dinagat (Villanueva, Citation2009). The species seems to be relatively rare.
Rhinagrion reinhardi has been recorded only in creeks and streams in forest, unlike R. philippinum which has also been found in rivers. One of the authors (RJTV) observed R. reinhardi regularly over a period of two years at a creek in Paragua Forest on Dinagat Island. At a seemingly uniform stretch of nearly 100 metres, males were found to perch only on two branches at a single spot. After a male was collected another male would take its place. These observations suggest that R. reinhardi is territorial.
Rhinagrion philippinum Selys, 1882 (Figures 1e, 5, 6d)
Specimens studied
Philippines, Cebu1 ♂: Badian, Kawasan Falls, 13 February 2001, leg. T. Borromeo. — 10 ♂, 7 ♀: Badian, Kawasan Falls, 12–13 March 2009, leg. RJTV (RJTV). — 1 ♂: Badian, Kawasan Falls, 12–13 May 2009, leg. RJTV (RJTV). — Philippines, Leyte 1 ♂, 1 ♀: Bontoc, 14 September 2009, leg. RJTV & H. Cahilog (RJTV). — Philippines, Luzon 5 ♂, 2 ♀: Apayao Province, Flora, Santa Maria, Lumon River area, 300–500 m, 14–20 April 1997, leg. C. Nazareno. — 1 ♀: Camarines Norte, San Lorenzo Ruiz, Sitio Bay-Bay, Patag, 400–700 m, 24–25 September 1997, leg. C. Nazareno. — 2 ♀: Aurora, Dilalungan, Mount Anaguao, Bungo River, Lomot creek, 1200–1400 m, 20 June 1997, leg. M. Dusayen. — 1 ♂, 1 ♀: Los Banos, 50 m, 28 May 1949, leg. L.B. Uichanco. — 1 ♀: ex coll. Los Banos (without further data). — 3 ♂, 1 ♀: Isabela, Palanan, Dipagsangaan, Dipinantahikan area, 16°53′ 39 N, 122°20′ 47
E, 12–20 September 2008, leg. R. Villanueva. — 1 ♂: Mount Maquiling, 100 m, 16 May 1949, leg. J. dela Paz. — 1 ♀: Los Banos, 20 February 1914, leg. G. Boettcher (SMF). — 1 ♂: Aroroy, 29 August 1917, leg. G. Boettcher (SMF). — 4 ♂, 1 ♀: Isabela, Palanan, Dipagsangaan, Dipinantahikan area (16°53′ 39
N, 122°20′ 47
E), 12–20 September 2008, leg. RJTV (RJTV). — 1 ♀: Isabela, Palanan, Ditalad creek, PLFDP-CI, 19 May 2004, leg. V.P. Gapud (RJTV). — Philippines, Masbate 1 ♂: Cataingan, Toybo, Borabod spring, 26 February–1 March 1993, leg. T. Borromeo. — Philippines, Mindoro 4 ♂: Victoria, Alcate, 6 April 1954, leg unknown. — Philippines, Polillo 2 ♂: Burdeos, Malat River, 23 April 2009, leg. RJTV (RJTV). — 5 ♂, 1 ♀: Salipsip area, 20–21 April 2009, leg. RJTV (RJTV). — 1 ♀: Pinaglubayan area, 19 April 2009, leg. RJTV (RJTV).
Differential diagnosis of male
Head with extensive blue pattern. Side of synthorax pale brown, dorsum of synthorax dull orange without large blue patch. Segments 1–10 reddish orange becoming black toward tip of the abdomen, S2–10 each with a pair pale bluish posterior spots of varying size (sometimes absent on the most distal segments), superior appendages reddish. Sternum of S8–9 orange to red.
Remarks
The type of the species comes from the island of Bohol, but R. philippinum has not been found there since. The morphology of the species is identical to that of R. schneideri and R. reinhardi except for the shape of the genital ligula ().
Material of Rhinagrion philippinum from the different islands shows some variation in the size of the blue spots on the head and the abdomen. This variation was deemed too minor to warrant any separate taxonomical status. Rhinagrion philippinum is the most widespread of the three Philippine species, occurring on the islands of Bohol, Cebu, Leyte (western part), Luzon, Masbate, Mindoro, Polillo (Gapud, Citation2006; Hämäläinen & Müller, Citation1997; Villanueva, et al. Citation2009). The material from Mindanao and Samar listed in Hämäläinen and Müller Citation(1997) refers to R. reinhardi and R. schneideri respectively.
Rhinagrion philippinum is found in a relatively wide range of habitats from small streams to shallow rivers. Males are commonly encountered perching on twigs over-hanging, or along, largely stagnant portions of streams or rivers; some are found several metres away inside thickets. Females are more commonly found well away from the water. In Cebu Island females have been found in small groups perching on the branches of Ficus sp., accompanied by one or two males perched slightly higher up in the tree.
During fieldwork in Polillo Island RJTV once recorded a territorial display of this species. Two males faced each other and flew fast in vertical circles. The movement was so fast that only the red ventral side of the apical abdominal segment was visible; it could not be seen if the abdominal tip was curved up as described for R. viridatum. The males flew up to the canopy and out of sight before the contest ended. A few minutes later what was presumably the same pair of males was seen repeating this behaviour.
Rhinagrion schneideri sp nov. (Figures 1f, 5, 6e)
Etymology
Noun in the genitive case, named in honour of Wolfgang Schneider, for his work for the World Dragonfly Association and especially for the help he offered to the senior author during his work on the Turkish dragonfly fauna.
Specimens studied
Holotype ♂: (white printed label) “Philippines, Samar Island, Samar Province, Hinubangan, San Isidro, San Isidro River, 90–200 m, 31 March–5 April 1992, leg. R.A. Müller”. The holotype is deposited in the RMNH.
Additional material: Philippines, Samar2 ♂: Eastern Samar, Oras, leg. T. Borromeo, 23 July 1992. — 1 ♂: Hinubangan, West Samar, 50–150 m, 1–14 April 1992, leg. T. Borromeo jr. — 1 ♂: Samar Province, Hinubangan, Arizona, 100–280 m, 29 March–6 April 1997, leg. R.A. Müller. — 1 ♂, 1 ♀: West Samar Province, Hinubangan, San Isidro area, 90–250 m, 5–12 May 1997, leg. A. Buenafe. — 1 ♀: Samar Province, Hinubangan, San Isidro, San Isidro River, 90–200 m, 31 March–5 April 1992, leg. R.A Müller. — 1 ♀: Eastern Samar, Oras, 10–22 August 1994, leg. T. Borromeo. — 1 ♀: Hinubangan, Bagakay, Arizona, 100–200 m, 29 August–20 September 1996, leg. A. Buenafe. — 1 ♂: Eastern Samar Province, San Rafael, 20–28 May 1993, leg. T. Borromeo. — 1 ♂: Northern Samar, Las Navas, San Isidro, 100–350 m, 22–28 May 1997, leg. A. Buenafe. — 1 ♂: Eastern Samar, Oras, 1–4 August 1992, leg. Th. Borromeo. — 1♂, 1♀: Basey, Sohoton, Balantak Falls, leg. RJTV & H. Cahilog, 13 September 2009 (RJTV).
Additional record: Philippines, Leyte 1♂: Abuyog, Barangay Malasiga, Malasiga Creek, 14 September 2009 (based on field note RJTV).
Differential diagnosis of male
Head with extensive blue pattern. Sides of synthorax pale brown, dorsum of synthorax darker, black along median carina and with ovoid oblique bluish streak on each side. S1–2 dull orange, S3 dull orange with darkened apical half, S4–5 dark brown to black, S6 and S7 black, both with large pinkish spot or pair of spots covering most of dorsum, S8–10 and superior appendages black. Sterna of S8–9 brown to black.
Description of holotype ♂
Head
Labium whitish-brown, longer (1.4 mm) than wide (1.1 mm), median cleft about half length of labium, about a quarter as wide as deep; mandible pale with distal tip black; labrum shiny black; genae bluish cream, except for narrow, black anterior border; anteclypeus black with central third bluish cream; postclypeus largely black; frons and dorsum of head black except for pale blue dots lateral to lateral ocelli and pale blue lines along eye margins, confluent with the pale genae and continuing to occiput, but interrupted on top of head.
Thorax
Anterior lobe of prothorax short, erect, brownish except for yellow central part of medial ridge; median lobe gently domed, brown with yellowish posteroventral sides; posterior lobe simple, fan-shaped, yellow with darkened anteromedial portion. Sides of synthorax pale brown, becoming paler toward venter. Dorsum of synthorax black along the median carina but outer half of mesepisternum brown. Pair of small ovoid pale blue spots situated at ca half length. – Coxae, trochanters and femora pale yellow, except for blackish streak on femora. Tibiae light brown. Spines dark brown. – Wings hyaline with black veins. Fw and Hw with 2 Ax; Fw with 15–16 and Hw with 13 Px. Wings petiolated to Ac; Ac near to Ax2; three cells between discoidal cell and subnodus in Fw and 2 in Hw; intercalated vein at the level of Px5–6. Pterostigma long, dark brown equilateral and broader centrally, spanning two or more cells.
Abdomen
S1–2 dull orange, both with faint, pale blue dorso-subapical streak; S3 dull orange with darkened apical dorsal half; dorsum S4–5 dark brown to black, dorsum S6 and S7 black, both with pair of large pinkish spots covering most of the dorsum, S8–10 black. S4–7 ventrally blackish except for a pale subapical portion. Sternum of S8–9 dark brown to black. Appendages black. Superior appendages in lateral view with basal fourth robust, then tapered; in dorsal view basal half cylindrical, apical half flattened interiorly, and abruptly constricted close to apex, with short spike on dorsal tip. Inferior appendages very short and hardly visible in lateral view. Genital ligula as shown in .
Measurements [mm]
Total length 47, abdomen 38, Fw 27; Fw pterostigma 1.8 (costal length), 2.1 (longest length); Hw pterostigma 2.1 (costal length), 2.4 (longest length).
Remarks
R. schneideri is known from Samar and the eastern part of Leyte. Material listed from Samar as R. philippinum by Hämäläinen and Müller Citation(1997) refers to this species. RJTV observed males and females perching on twigs above small pools along streams on Samar and Leyte. Several rivers were explored, but the species was found only on small tributary streams.
Rhinagrion tricolor Krüger, 1898 (Figures 1d, 2, 6a)
Specimens studied
Indonesia, Java1 ♂: Tengger, Java, leg. M.C. Piepers. — 3 ♂, 2 ♀, SW Java, Kalipoetjang, 18 July 1936, 200 m, leg. M.A. Lieftinck. — 1 ♂: W Java, S Coast, Sindangbarang, Salatri-bivak, 14 October 1937, leg. L.J. Toxopeus. — 2 ♂: SW Java, S coast, Tjitoë, (Tjidamar), 28 November 1935, leg. M. Bartels Jr. — 2 ♂: SW Java, S coast, Tjitoë, (Tjipandak), 15 September 1935, leg. M. Bartels. — 1 ♂, 1 ♀: SW Java, S coast, Tjitoë, (Tjipandak), 13 September 1935, leg. M. Bartels. — 1 ♂, 1 ♀: SW Java, S coast, Tjitoë river, (Tjidamar-Tjipandak), 12 October 1935, leg. M. Bartels. — 2 ♂: SW Java, S coast, Tjidaoen, Tjipoenage, 18 November 1935, leg. M. Bartels Jr. — 1 ♂: W Java, Udjung Kulon, 19 June 1958, collector unknown.
Differential diagnosis of male
Face with extensive blue pattern. Dorsum of synthorax black, with medium–large blue marks broadly separated by median carina. S2–3 blackish with small blue marks, S5 reddish black with tiny blue spots, S5–9 completely red without large blue markings, S10 reddish black, superior appendages black. Sterna of S8–9 red.
Remarks
R. tricolor is only recorded from SW Java (Krüger, Citation1898; Lieftinck, Citation1934) and has not been recorded since 1958. The records are from coastal areas in the lowlands. Large parts of this region have been deforested (Whitten, Soeriaatmadja & Afiff, Citation1996); the species is likely to have suffered a decline as a consequence of this. However it was recorded in 1958 in Udjung Kulon which is now a National Park; the species might be safe there. Lieftinck Citation(1956) states that “the rare Javan species R. tricolor, for instance, is a genuine rheobiont, whereas borneense breeds in shady forest brooks with a slow current”.
Rhinagrion viridatum Fraser, 1938 (Figures 3c, 4)
Specimens studied
All leg. M. Hämäläinen unless otherwise noted.
Burma1 ♂ (Holotype), Tenasserim–Bachrup border, 28 May 1932, leg. A. Kerr (BMNH). Malaysia, Peninsular Malaysia1 ♂, 1 ♀: Kelantan (west from Gua Musang) Sungai Lasau, 10 April 1995. — 1 ♂: Kelantan (West from Gua Musang) Sungai Selieh, 13 April 1995. — 1 ♂: Pahang, Krau Wildlife Reserve, Kuala Gandah, Sungai Gandah, 22 April 1995. — 1 ♂: idem, 18 November 1996, leg. M. Hämäläinen & H. Olsvik. — 1 ♂: idem, 26 November 1998. — 1 ♂: Pahang, Krau Wildlife Reserve, Pos Perlok, Sungai Perlok, 28 November 1996, leg. M. Hämäläinen & H. Olsvik. — 3 ♂, 1 ♀: Pahang, Krau Wildlife Reserve, Bukit Rengit, Sungai Rengit, 21–26 January 1996. — 2 ♂: idem, 24 January 1995. — 3 ♂: idem, 5–8 June 1997. — 1 ♂: idem, 60–70 m, 18–19 December 2003. 1 ♂: Pahang, Bilut Valley, 6 October 1960, ex coll Pagden. — 1 ♂: Perak – Pahang, [ex col Forster, published in Handlist]. — 1 ♂: Selangor, Hulu Langat area, Dusun Tua, near Kuala Lumpur, 22 March 1963, M.A. Lieftinck. Thailand1 ♂: Krabi, Than Bokkharani, 26 October 1982. — 1 ♂: Loei, Phu Kradung, 1200–1250 m, 28 October–3 November 1996, leg. M. Hämäläinen & H. Olsvik. — 1 ♂: Nakhon Nayok, Khao Yai, Haew Suwat, 2 November 1986.— Krabi, Khao Phanom Bencha, Huai To Waterfall, 9–10 January 1993. — 1 ♂: Nakhon Si Thammarat, Khao Luang, Huai Mungtam, 17 January 1993. — 1 ♂: Phang Nga, Khao Lampi-Hat Thai Muang National park, Fon Prow waterfall, 28 November 1997. — 3 ♂: Phetchaburi, Kaeng Krachan, Ban Krang camp, Mae Nam Pranburi, 290–320 m, 24–26 October 2000. — 3 ♂: Ranong, Khlong Nakha, Khlong Bang Mun, 9–11 April 2000. — 3 ♂: idem, 24 December 2001. — 2 ♂: idem, 20 m, 8–12 May 2002. — 5 ♂: 1 ♀, Ranong, Khlong Nakha, 8–10 February 1988. — 1 ♂: idem, 7 January 1993. — 1 ♂: Ranong, Kampuan, 15 January 1987, leg. unknown, ex. coll. Pinratani.
Differential diagnosis of male
Face black with extensive yellow pattern. Antehumeral stripe parallel sided and covering about two-thirds length of synthorax. Side of synthorax with dark stripe along metapleural suture not connected to dark dorsum of synthorax. Abdomen yellow and black, S3–5 with no distinct blue apical spot, superior appendages black with apical third to half yellow-orange. Sterna of S8–9 brown to black.
Remarks
In Burma the species is only known from the holotype, which was collected in the southern tip of the country in Tenasserim, which is the present day Burmese province of Tanintharyi. All records previously published as R. mima from Peninsular Malaysia belong to R. viridatum. R. viridatum and R. mima seem to have allopatric ranges in Thailand with R. mima occurring in the north-west and R. viridatum in other parts of the country. The only record of R. viridatum from Laos (as R. mima) is from Phatang, Vang Vieng area (Sasamoto & Honda, Citation2003). Several Vietnamese websites (e.g. www.phuquocconfidential.com; accessed on 1 April 2010) mention the first records of R. viridatum (as R. mima) from Vietnam: Phu Quoc Island, Kien Giang province in South Vietnam. The pictures of a male published on the Internet on 25 December 2008 clearly show R. viridatum. This record makes it clear that R. viridatum undoubtedly also occurs in Cambodia. The border between R. viridatum and R. hainanese is not clear. Probably these species have, like R. viridatum and R. mima, allopatric ranges.
Wilson and Gibert Citation(2005) described the territorial behaviour of R. viridatum (as R. mima): “Males involved in territorial disputes, located at favoured, sunlight spots beside sizable flowing streams, angled the tips of their orange-red abdomens upward and flew side by side for long periods, often slowly rising up toward the adjacent forest canopy. The flights terminated when one of the males would fly rapidly to take up a prominent position at a favoured sunlight spot, with the losing male taking up a position several metres away in a less favoured position. The disputes were often quickly resumed and in one case continued for over twenty minutes.” Wilson and Gibert Citation(2005) also provided information on oviposition by R. viridatum: “one female was observed ovipositing into mossy vegetation, which was growing on a vertical bank, located some 1.5 metres above the stream surface, adjacent to a clump of screwpine (Panadanus sp.) with an extensive aquatic root system”.
Key to the males of Rhinagrion
1. Antehumeral stripe absent or not parallel sided and clearly broader on front of mesepisternum. Lateral horns of genital ligula without spines, at most with minute denticles ……………… 2 | |||||
1′. Antehumeral stripe parallel sided and covering about two-thirds the length of mesepisternum. Lateral horns of genital ligula with numerous small spines (, b) … 3 – mima group | |||||
2. Main colour of S8–9 blue or red. Dark line along metapleural suture present (can be faint) and connected with dark dorsum of thorax; together these form a Z-shape. Apex of genital ligula is, at the place where the two lateral horns meet, strongly bent (, d). Sundaland species. …………………… …………………………………………… 5 – borneense group | |||||
2′. Main colour of S8–9 black, sometimes with small dorsal blue spot. Dark line along metapleural suture is absent. Apex of the genital ligula is, at the place where the two lateral horns meet, not strongly bent (, f). Philippine species ………………… 8 – philippinum group | |||||
3. Dorsum of S8–9 largely brown to black. Sternites S8–9 orange to red. Superior appendages entirely pale ……………………………………… ……………………………R. hainanense | |||||
3′. Dorsum of S8–9 largely yellow. Sternites S8–9 dark brown to black. Basal half of superior appendages black ……………………………… …………………………………………… 4 | |||||
4. S3–4 brown to black with apical blue spot covering a third to half of the segment. Dorsum of S5–6 largely black …………………………… …………………………………R. viridatum | |||||
4′. S3–4 yellow without apical blue spot although pale white-yellow spots can be present. Dorsum of S5–6 black with some orange pattern and with or without an apical black ring …R. mima | |||||
5. Spots on dorsum of synthorax large and narrowly divided by middorsal carina ……………6 | |||||
5′. Spots on dorsum of synthorax small and broadly divided by middorsal carina ………………7 | |||||
6. Dorsum of S8–9 blue. Sternum of S8 pinkish white, sternum of S9 white ..R. macrocephalum | |||||
6′. Dorsum of S8–9 red. Sternum of S8–9 red ……………………… …………………R. tricolor | |||||
7. Face and head with extensive pale pattern. S7 red with apical fourth blue ……… R. elopurae | |||||
7′. Face and head black. S7 red, occasionally with small blue apical spots ………… R. borneense | |||||
8. Dorsum of synthorax black with large blue, half oval shaped patch extending over median carina, occupying c. two-thirds of the length of synthorax ………………………R. reinhardi | |||||
8′. Dorsum of synthorax yellow-brown to brown with pair of small and sometimes faint blue marks which are less than a fourth of the length of synthorax and are divided by median carina …………………………… ………………………………………………………………….9 | |||||
9. Dorsum of S6–7 with large pink spots; spot on S6 covers over four-fifths of the segment. Dorsum of thorax dark brown. Distal part of genital ligula gradually tapering toward apex ) ………………………………… ……………………………………R. schneideri | |||||
9′. Dorsum of S6–7 with small blue spots; spot on S6 covers less than half of the segment. Dorsum of thorax pale brown. Distal part of genital ligula with broad base and a clear constriction in the apical half () ………………………………… …………………R. philippinum | |||||
Discussion
Taxonomic relations within Rhinagrion
In many Zygoptera the prothorax of the female forms a lock, which can only be “opened” by the right key, the anal appendages of a conspecific male. There seems to be a trend in the Zygoptera that families in which this lock-key mechanism is strongly developed, for instance the Platystictidae, have less pronounced patterns of coloration. On the other hand families in which the prothorax of the female and the appendages of the male show little variation between species often show pronounced patterns of coloration of body or wings often correlated with display behaviour of the males, as is for instance found in Calopterygidae and Chlorocyphidae. Rhinagrion seems to falls in the latter group, where differences in prothorax of the female and the appendages of the male are small between species and mate recognition is probably largely dependent on patterns of coloration probably in combination with display behaviour. In Zygoptera the secondary genitalia of the male and the vagina of the female form a second key-lock system ensuring that only conspecifics can mate. Here the same trend seems to be visible with brightly coloured families showing relatively little variation in the secondary genitalia (for instance Chlorocyphidae) and far less colourful families showing distinct variation in the build of the ligula (for instance Platystictidae). In Rhinagrion the initial mate recognition seems to be served by the differences in coloration, as the prothorax and anal appendages vary only marginally different between species. In line with the above-mentioned trends it would be expected that Rhinagrion would also show little variation in the genital ligula. This is however only partially true, with well-defined differences in the ligula shape in the borneense group but small differences in the mima group. The most problematic species pair from a taxonomical point of view is that of R. mima and R. Viridatum, which are morphologically the same and have to be recognized based on differences in coloration and pattern. The former occurs in two areas separated by a wide area where the closely related R. viridatum occurs. It is not unlikely that the two areas where R. mima is found represent two taxa, each more closely related to R. viridatum than to each other.
Three different groups can be recognized within Rhinagrion. The mima group, containing R. mima, R. viridatum and R. hainanese, share the same pattern on the thorax and have a good apomorphic character in the numerous spines on the horns of the genital ligula. The other two groups, the borneense group (borneense, elopurae, tricolor, macrocephalum) and the philippinum group (philippinum, schneideri, reinhardi), lack these spines and are probably more closely related to each other than to the mima group. There are no structural differences between the borneense group and the philippinum group except for small differences in the shape of the genital ligula head (see key). The members of the borneense group have a brighter red and blue pattern on thorax and abdomen and a more pronounced black pattern on the thorax.
Habitat and behaviour
The species of Rhinagrion are mainly found at slower running stretches of brooks and, in some species, rivers. They are confined to forest and largely limited to the lowlands, although R. borneense has been found above 1000 m. A preference for places with Pandanus has been mentioned (Orr, Citation2003), but most species have also been found in places with no Pandanus and Rhinagrion is certainly not limited to habitats with Pandanus.
The scarcity of observations on behaviour of these conspicuous damselflies is probably not due to a lack of behaviour but to the fact that they seldom display it. Males are far more common at the waterside than females, which is nicely illustrated by the 174 males and 17 females of R. borneense collected at Tabang, Kalimantan, in the period 31 August to 1 November 1956. Males are mostly found sitting on trunks or twigs at one or two metre height, but not often higher, facing the stream. They sit with open wings in most cases with the abdomen slightly above or below horizontal. The observations of R. reinhardi show that some perches are preferred over others, but it is not clear if they occupy the same territory for several days.
The bright colours of the species of Rhinagrion probably play a role in courtship and agonistic behaviour. For R. borneense it was noted that males can expand the blue tip of their abdomen and it seems likely that other species are also capable of doing so. This expanding of the tip of the abdomen might be to attract females, to scare off other males or both. Courtship and/or copulation have not been observed; the role of the coloration in courtship is unknown. Agonistic behaviour was observed for R. borneense, R. viridatum and R. philippinum. The scant information available suggests that this is different in each of these three species. In R. borneense the males fly facing each other and hold the end of the abdomen flexed upwards or downwards, in R. viridatum the males fly side by side with the tip of the abdomen curved upwards, and in R. philippinum the males fly in circles, quickly gaining height. In R. borneense and R. viridatum it was noted that the tip of the abdomen was flexed, probably in order to show the colourful upper- or underside of the tip. This could not be seen in R. philippinum due to the speed of the action, but it was noted that the red of the sternum of the last segments was very much visible suggesting that it was also expanded and turned up. Apparent oviposition was only observed for R. borneense, which used a mud bank beside a pebble-bottomed stream, and R. viridatum, which used mossy vegetation some 1.5 metres above the stream surface.
Distribution
The three different groups of Rhinagrion all have their own areas of distribution. The mima group is found in mainland South-East Asia and Sumatra, the borneense group in Sundaland and the philippinum group in the Philippines. The only species known to overlap in distribution are R. mima and R. macrocephalum in Sumatra and R. viridatum and R. macrocephalum in Peninsular Malaysia. In both cases the pairs comprise one species each from the well-differentiated mima group and borneense group, allowing their sympatric occurrence. Thus far no species of a single species group have found to co-occur, although R. schneideri and R. philippinum are both found on the island of Leyte. The limits of distribution are largely clear for most species. It would, however, be interesting to get better information on the border between R. viridatum and R. hainanese and on the southernmost occurrence of R. macrocephalum on Sumatra, as it is not completely impossible that R. tricolor occurs in South Sumatra. The later is endemic to south-west Java and has not been found since 1958. This species is now listed as Data Deficient on the IUCN Red List; fieldwork in this area is needed in order to assess its threat status.
Acknowledgements
Rory Dow and Dennis Paulson contributed strongly to this paper by making information and unpublished records available and by reviewing the paper. Matti Hämäläinen, Jan van Tol and Graham Reels helped with comments, pictures and material. Wolfgang Schneider and Stefan Ober helped during visits to the SMF and ZSMC, respectively.
References
- Cleary , D. F.R. , Mooers , A. O. , Eichhorn , K. A.O. , van Tol , J. , de Jong , R. and Menken , S. B.J. 2004 . Diversity and community composition of butterflies and odonates in an ENSO-induced fire affected habitat mosaic: a case study from East Kalimantan, Indonesia . Oikos , 105 : 426 – 448 .
- Donnelly , N. 1994 . Through darkest Borneo [and Malaysia] with net and camera . Malangpo , 11 : 123 – 125 .
- Donnelly , T. W. 1999 . Back to Borneo . Argia , 11 : 8 – 11 .
- Dow , R. 2005 . Odonata, burglary and ballistic cicadas in South-east Asia . Agrion , 9 : 10 – 12 .
- Dow , R. 2008 . Odonata of the “Glen Forest” at Samarakan, Bintulu Division, Sarawak, Malaysian Borneo . Agrion , 12 : 46 – 48 .
- Fraser , F. C. 1933 . The fauna of British India, including Ceylon and Burma. Odonata , Vol. 1 , London : Taylor and Francis .
- Fraser , F. C. 1938 . Two new species of Oriental Odonata . Proceedings Royal Entomological Society London Series B , 7 : 197 – 198 .
- Gapud , V. P. 2006 . Damselflies (Odonata: Zygoptera) of Greater Luzon, Philippines with description of two new species . Philippine Entomologist , 19 : 1 – 42 .
- Hämäläinen , M. and Müller , R. A. 1997 . Synopsis of the Philippine Odonata, with lists of species recorded from forty Islands . Odonatologica , 26 : 249 – 315 .
- Hämälänen , M. and Pinratana , A. 1999 . “ Atlas of the dragonflies of Thailand: distribution maps by provinces ” . Bangkok : Brothers of St. Gabriel in Thailand .
- Hincks , W. D. 1930 . Some notes on a collection of Sarawak Odonata . Sarawak Museum Journal , 4 : 49 – 56 .
- Kalkman , V. J. , Choong , C. Y. , Orr , A. G. and Schütte , K. 2010 . Remarks on the taxonomy of Megapodagrionidae with emphasis on the larval gills (Odonata) . International Journal of Odonatology , 13 : 119 – 135 .
- Kemp , R. G. and Kemp , G. S. 1989 . Some dragonfly records from the states of Pahang and Perak, West Malaysia . Notulae Odonatologicae , 3 : 37 – 40 .
- Kimmins , D. E. 1936 . The Odonata of the Oxford University Sarawak expedition . Journal of the Federated Malay States Museum , 18 : 65 – 108 .
- Krüger , L. 1898 . Die Odonaten von Sumatra. I. Theil. Familie Agrioniden. [Including ‘Anhang’: Neue Calopteryginen und Agrioninen aus der südostasiatischen Fauna ausserhalb Sumatra] . Stettiner Entomologische Zeitung , 59 : 64 – 139 .
- Laidlaw , F. F. 1920 . Contributions to the study of the dragonfly fauna of Borneo. Part IV. A list of species known to occur in the island . Proceedings of the Zoological Society of London , 1920 : 311 – 342 .
- Lieftinck , M. A. 1934 . An annotated list of the Odonata of Java, with notes on their distr., habits and life-history . Treubia , 14 : 377 – 462 .
- Lieftinck , M. A. 1935 . A synopsis of the Odonata (Dragonflies) of Sumatra . Miscellanea Zoologica Sumatrana , 92/93 : 1 – 23 .
- Lieftinck , M. A. 1953 . New dragonflies (Odonata) from Borneo, with notes on their habits and larvae . Treubia , 22 : 381 – 406 .
- Lieftinck , M. A. 1954 . Handlist of Malaysian Odonata. A catalogue of the dragonflies of the Malay peninsula, Sumatra, Java and Borneo, including the adjacent small islands . Treubia (Supplement) , 22 : 1 – 202 .
- Lieftinck , M. A. 1956 . Revision of the genus Argiolestes Selys (Odonata) in New Guinea and the Moluccas, with notes on the larval forms of the family Megapodagrionidae . Nova Guinea (N.S.) , 7 : 59 – 121 .
- Matsuki , K. and Kitagawa , K. 1993 . Bornean Odonata taken by Dr. T. Ueda. II. Zygoptera . Aeschna , 27 : 1 – 10 .
- Orr , A. G. 2001 . An annotated checklist of the Odonata of Brunei with ecological notes and descriptions of hitherto unknown males and larvae . International Journal of Odonatology , 4 : 167 – 220 .
- Orr , A. G. 2003 . Dragonflies of Borneo , Kota Kinabalu : Natural History Publications .
- Paulson , D. R. 1981 . Anatomical modifications for displaying bright coloration in megapodagrionid and chlorocyphid dragonflies (Zygoptera) . Odonatologica , 10 : 301 – 310 .
- Sasomoto , A. 2003 . Description of a Rhinagrion species from Laos (Megapodagrionidae; Zygoptera) . Tombo , 46 : 17 – 19 .
- Sasamoto , A. and Honda , T. 2003 . Collecting records of Odonata in the Laos in the spring of 2002 . Aohada , 2 : 1 – 21 . [in Japanese]
- de Selys Longchamps , E. 1862 . Synopsis des Agrionines. Troisième légion: Podagrion . Bulletin Academie royale Belgique Serie , 2 ( 14 ) : 5 – 44 .
- de Selys Longchamps , E. 1882 . Odonates des Philippines . Anales de la Sociedad Española de Historia Natural , 11 : 1 – 32 .
- de Selys Longchamps , E. 1886 . Révision du synopsis des Agrionines. Première partie comprenant les légions Pseudostigma – Podagrion – Platycnemis et Protoneura . Memoires couronnes Academie royale Belgique , 38 : 1 – 233 .
- Thompson , D. J. 1998 . On the biology of the damselfly Euphaea ameeka van Tol & Norma-Rashid in Borneo (Zygoptera: Euphaeidae) . Odonatologica , 27 : 259 – 265 .
- Thompson , D. J. and van Tol , J. 1993 . Damselflies and dragonflies from four forest types in Brunei . Brunei Museum Journal , 8 : 57 – 72 .
- Vick , G. S. 1993 . A visit to Malaysia in July 1993 . Kimminsia , 4 : 37 – 39 .
- Villanueva , R. J.T. 2009 . Odonata of Dinagat Island, Philippines: Updated species lists and notes on conservation of species and habitats . Notulae Odonatologicae , 7 : 27 – 35 .
- Villanueva , R. J. , van der Ploeg , J. and van Weerd , M. 2009 . Some Odonata from the Northern Sierra Madre Natural Park . Agrion , 13 : 72 – 74 .
- Watson , J. A.L. and O'Farrell , A. F. 1991 . “ Odonata (dragonflies and damselflies) ” . In The insects of Australia , 2 , Edited by: Naumann , I. D. , Carne , P. B. , Lawrence , J. F. , Nielsen , E. S. , Sprabery , J. P. , Taylor , R. W. , Whitten , M. J. and Littlejohn , M. J. 294 – 310 . Melbourne : Melbourne University Press .
- Whitten , T. , Soeriaatmadja , R. E. and Afiff , S. A. 1996 . The Ecology of Java and Bali , Hong Kong : Periplus Editions .
- Wilson , K. D.P. and Reels , E. 2001 . Odonata of Hainan . Odonatologica , 30 : 145 – 208 .
- Wilson , K. D.P. and Gibert , E. 2005 . Endau-Rompin 20–24 July ’04 . Agrion , 9 : 26 – 28 .
- Zhang , H. , Kalkman , V. J. and Tong , X. 2011 . A synopsis of the genus Philosina with descriptions of the larvae of P. alba and P. buchi (Odonata: Megapodagrionidae) . International Journal of Odonatology , 14 : 55 – 68 .