Abstract
Objective. To test the hypothesis that limb ischemic postconditioning protects the myocardium from reperfusion injury, and examine the mechanism involved. Design. Forty rabbits were randomly divided into four groups: Control, Ischemic Preconditioning, Ischemic Postconditioning and Remote Postconditioning. Myocardial infarct size and tissue myeloperoxidase activity were determined at the end of the experiment. Plasma creatine kinase and malondialdehyde activity were measured at baseline, the end of ischemia, and after 3 h of reperfusion respectively. Results. Myocardial infarct size was significantly reduced in Ischemic Preconditioning, Ischemic Postconditioning and Remote Postconditioning as compared to Control (p < 0.01). Results were confirmed by plasma creatine kinase activity. Plasma malondialdehyde was significantly less at 3 h of reperfusion in Ischemic Preconditioning, Ischemic Postconditioning and Remote Postconditioning than that in Control (p < 0.01). Neutrophil accumulation (myeloperoxidase activity) in the area at risk was less in Ischemic Preconditioning, Ischemic Postconditioning and Remote Postconditioning than that in Control (p < 0.01). Conclusion. Remote postconditioning reduces myocardial infarction in rabbits. The mechanism involved might be reduced oxygen radical-induced injury and improved antioxidant action.
In 1986, Murry et al. Citation1 first introduced the concept of ischemic preconditioning (IPC) as a potent endogenous form of cardioprotection against ischemia-reperfusion injury, in which repetitive brief periods of ischemia protected the myocardium from a subsequent longer ischemic insult. IPC attenuates the incidence and severity of post-ischemic arrhythmias, Citation2–6 enhances the recovery of cardiac function after myocardial ischemia, and reduces infarct size and the appearance of apoptosis in hearts subjected to ischemia/reperfusion injury Citation7, Citation8 This approach, however, crucially depends on intervening before the ischemic event, which is difficult to control, given the unpredictable onset of an acute coronary artery occlusion.
Recently, Zhao et al. Citation9 reported another endogenous form of cardioprotection. In their study, a short series of repetitive cycles of brief reperfusion and re-occlusion of the coronary artery applied immediately at the onset of reperfusion, which was termed postconditioning Citation9, was also cardioprotective for reducing infarct size, coronary artery endothelial dysfunction, and neutrophil accumulation in the area at risk. This protection was, to a certain extent, similar to IPC. To date, cardioprotection by postconditioning has been reported by independent laboratories in several species (including dogs, rabbits, rats and mice Citation10) as well as isolated perfused hearts Citation11. Adenosine was implicated as adenosine blockers could inhibit the benefit. Other mechanisms that have been implicated in the postconditioning phenomenon are epithelial nitric oxide synthase, nitric oxide and guanylyl cyclase, the KATP channel, and closing of the mitochondrial permeability transition pore Citation10.
In 1993, Przyklenk et al. Citation12 first reported that 4 cycles of 5 min left circumflex coronary artery occlusion and 5 min reperfusion reduced infarct size following 1 h sustained left anterior descending coronary artery occlusion and 4.5 h reperfusion in anesthetized dogs. Subsequently, cardioprotection by remote preconditioning from intestine, kidney and limb of several species was reported Citation13–15. This phenomenon was named inter-organ preconditioning or remote preconditioning. Recently, Kerendi et al. Citation16 reported that in an in vivo rat model of myocardial infarction induced by coronary artery occlusion and reperfusion, remote postconditioning produced by a single 5-min episode of renal artery occlusion and reperfusion applied immediately before the onset of coronary artery reperfusion protects the myocardium from reperfusion injury by mechanism involving endogenous adenosine receptor activation. In the present study, we tested the hypothesis that remote postconditioning induced by a single 5-min episode of femoral artery occlusion and reperfusion applied just before the onset of coronary artery reperfusion, protects the myocardium from reperfusion injury, and examined the mechanism involved.
Material and design
Animal preparation
Forty healthy New Zealand white rabbits of either sex weighing 2.5–3.0 kg were purchased from Shandong Provincial Agriculture Academy of Science. The study has been approved by Ethics Committee of Shandong University. All animals were randomized into four groups: Control, Ischemic Preconditioning (Pre-con), Ischemic Postconditioning (Post-con) and Remote Postconditioning (Re Post-con).
Surgical procedure
All animals were anesthetized with intravenous pentobarbital sodium (30 mg/kg) through the left auricular margin vein. During the experiment, additional anesthetic was administered as needed. The rabbits were intubated and mechanically ventilated with room air enriched with 100% oxygen using an animal ventilator (ALC-5, Beijing, China). The left carotid artery was cannulated and connected to a fluid-filled pressure transducer (YP200, Shanghai, China) to monitor mean arterial pressure (MAP) and heart rate (HR). The right femoral artery was exposed below the right inguinal ligament for later induction of femoral artery occlusion. The chest was opened through the left fourth intercostal space to expose the heart. After pericardiotomy, a 4-0 silk suture was placed under the left anterior descending (LAD) artery. The end of the suture was threaded through a piece of tubing, forming a snare for reversible LAD occlusion. The presence of myocardial ischemia was confirmed by significant ST segment elevation indicated by ECG (XDH-3, Shanghai, China). Heparinization was maintained during the experimental period with a bolus injection of 3 mg/kg heparin sodium. The body temperature was monitored by rectal thermometer and maintained constant between 38.5°C and 39.5°C by a heating pad.
Experimental protocol
After a 10-min stabilization period, baseline MAP and HR were measured. All rabbits were subjected to 30 min LAD occlusion, followed by 3 h of reperfusion. The rabbits were randomized to one of four treatment groups (n = 10 in each group): 1) Control: LAD occlusion and reperfusion only, with no other intervention; 2) Pre-con: The LAD was undergone three cycles of 5 min ischemia and 5 min reperfusion before the prolonged LAD occlusion; 3) Post-con: After 30 min of LAD occlusion, reperfusion was initiated for 30 s followed by 30 s reocclusion, repeated for three cycles; 4) Re Post-con: After 24 min of LAD occlusion, the right femoral artery was occluded for 5 min and released for 1 min before 3 h of LAD reperfusion.
Analysis of myocardium at risk of infarction and infarct area
At the end of experiment, the coronary artery was reoccluded in the original position, and diluted Evans blue dye was injected into the aortic root under 80 mmHg perfusion pressure to stain the non-ischemic zone (NIZ) blue and thereby outline the area at risk (AAR). The left ventricle (LV) was sliced transversely into six to eight sections each one of which was about 2 mm thick. And the AAR (uncolored by blue dye) was separated from the NIZ (colored by blue dye). Then the non-stained AAR was incubated in a 1% solution of buffered TTC (pH 7.4) at 37°C for 15 min to differentiate the unstained gray area of necrosis (AN) from the stained AAR (brick red). The AAR was expressed as a percentage of the LV mass (AAR/LV), and the AN was expressed as a percentage of the AAR (AN/AAR). The mass of each area was calculated by tissue weight.
Determination of plasma creatine kinase (CK) and malondialdehyde (MDA) activity
Blood samples were collected from the right auricular margin vein at baseline, the end of ischemia, and after 3 h of reperfusion respectively for measuring plasma CK and MDA activity. The blood samples were centrifuged at 2 500 rpm and 4°C for 10 min. Then the plasma was collected, analyzed CK activity and protein concentration spectrophotometrically at 340 nm absorbance (7230 spectrophotometer, Shanghai, China). The activity of MDA was measured spectrophotometrically at 586 nm absorbance. Plasma CK activity was expressed in international units per gram protein (IU/g) and MDA activity was expressed in µM per ml plasma (µM/ml). The kits were purchased from Nanjing Jiancheng Biological Engineering Institute of China.
Determination of tissue myeloperoxidase (MPO) activity
After determining infarct area, tissue samples were taken from NIZ and AAR for analysis of MPO activity, an enzyme used as a marker of neutrophil accumulation in myocardium. The activity of MPO was measured by spectrophotometrically at 460 nm absorbance and expressed in units per 100 g tissue (U/100 g).
Statistical analysis
All data were expressed as means±standard deviation (SD). A one-way analysis of variance (ANOVA) was used to analyze group differences in a single data such as infarct area and MPO activity data. Time-dependent determinations were analyzed by repeated measures analysis of variance followed by post-hoc analysis with Student-Newman-Keuls multiple comparisons. A p-value less than 0.05 was considered statistically significant.
Results
Hemodynamic data
Hemodynamic data for MAP and HR in the four groups are measured ( and ). There were no significant differences among the four groups at baseline. During coronary occlusion, there were trends for a decrease in MAP and an increase in HR. But these changes were not significant compared with their respective baseline value.
Table I. HR (bpm) during ischemia and reperfusion
Table II. MAP (mmHg) during ischemia and reperfusion
Area at risk and infarct size
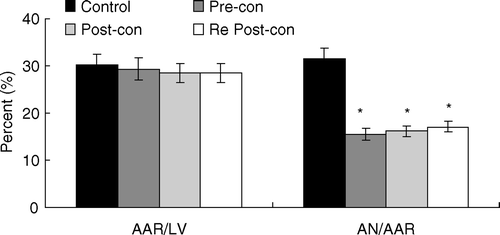
The area placed at risk by LAD occlusion was comparable among the four groups, averaging between 27% and 31% (). Infarct size in Pre-con (15.47±1.65) was reduced by 51%, in Post-con (16.15±2.05) reduced by 49% compared to Control (31.46±1.28). Similarly, infarct size in Re Post-con (17.11±1.70) was reduced by 46% compared to Control. There was no statistical difference in infarct size between Pre-con, Post-con and Re Post-con.
Figure 1. Bar graph showing area at risk (AAR) expressed as a percentage of the left ventricle (LV) and area of necrosis (AN) expressed as a percentage of the AAR. Pre-con = Ischemic Preconditioning; Post-con = Ischemic Postconditioning; Re Post-con = Remote Postconditioning. Re Post-con significantly reduced AN/AAR ratio by 46% compared with Control, showing equivalent cardioprotection to that of Pre-con and Post-con. *p < 0.01 Pre-con, Post-con and Re Post-con vs. Control. Values are means±SD
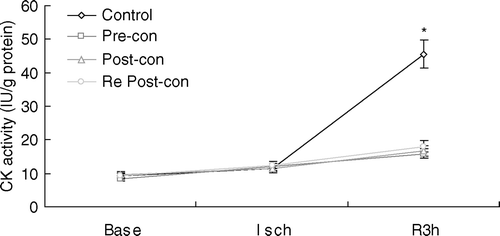
Plasma CK activity
CK activity was used to confirm myocardial infarct size quantified by TTC staining. Plasma CK activity at baseline was comparable among the four groups (). During ischemia, it was slightly increased in all groups. However, it was significantly increased during reprefusion in Control. At 3 h of reperfusion, CK activity in Pre-con (15.89±1.51), Post-con (16.69±1.51) and Re Post-con (18.06±1.63) was significantly less than that in Control (45.60±5.53) (p < 0.01). There was no statistical difference between Pre-con, Post-con and Re Post-con. Group differences in CK activity were consistent with those observed in infarct size.
Figure 2. Line graph showing plasma creatine kinase (CK) activity during the course of experiment. Base = baseline; Isch = end of ischemia; Pre-con = Ischemic Preconditioning; Post-con = Ischemic Postconditioning; R3h = 3 h of reperfusion; Re Post-con = Remote Postconditioning. Plasma CK activity at 3 h of reperfusion was significantly lower in Pre-con, Post-con and Re Post-con than that in Control. There was no statistical difference between Pre-con, Post-con and Re Post-con. *p < 0.01, Control vs. Pre-con, Post-con and Re Post-con at 3 h of reperfusion. Values are means±SD.
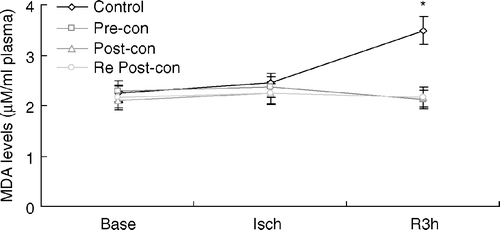
Plasma MDA levels
Appearance of MDA-reactive products in plasma has been used as an indicator of lipid peroxidation. There was no significant difference in plasma MDA levels among all groups at baseline (). During coronary occlusion, the plasma MDA was slightly increased, with no group differences. However, it was significantly increased in Control (3.49±0.32) at 3 h of reperfusion, at which time MDA level in Pre-con (2.12±0.30), Post-con (2.17±0.24) and Re Post-con (2.16±0.33) was significantly lower (p < 0.01). There was no statistical difference between Pre-con, Post-con and Re Post-con.
Figure 3. Line graph showing plasma malondialdehyde (MDA) levels during the course of experiment. Base = baseline; Isch = end of ischemia; Pre-con = Ischemic Preconditioning; Post-con = Ischemic Postconditioning; R3h = 3 h of reperfusion; Re Post-con = Remote Postconditioning. There was no significant difference among the four groups at baseline and after 30 min of ischemia. However, plasma MDA was significantly greater in Control at R3h compared to Pre-con, Post-con and Re Post-con. *p < 0.01, Control vs. Pre-con, Post-con and Re Post-con at 3 h of reperfusion. Values are means±SD.
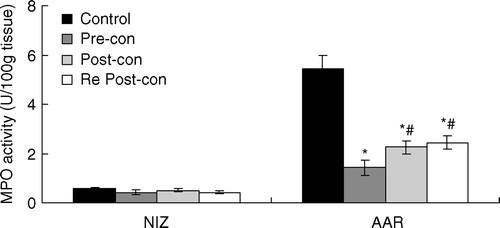
Myocardial MPO activity
MPO activity in the NIZ was low and comparable among the four groups (). After myocardial ischemia and reperfusion, MPO activity in the AAR was significantly greater than that in the NIZ. However, it was significantly less in the AAR in Pre-con (1.43±0.32), Post-con (2.26±0.28) and Re Post-con (2.45±0.28) than that in Control (5.44±0.46) (p < 0.01). There was no statistical difference between Post-con and Re Post-con. However, MPO activity in the AAR of Pre-con was significantly lower than that of Post-con and Re Post-con (p<0.05).
Figure 4. Bar graph showing myeloperoxidase (MPO) activity in the NIZ and AAR. AAR = area at risk; NIZ = nonischemic zone; Pre-con = Ischemic Preconditioning; Post-con = Ischemic Postconditioning; Re Post-con = Remote Postconditioning. MPO activity in the NIZ was low and comparable among the four groups. However, MPO activity in the AAR was significantly less in Pre-con, Post-con and Re Post-con than that in Control. MPO activity in the AAR of Pre-con was significantly lower than that of Post-con and Re Post-con. *p < 0.01 Pre-con, Post-con and Re Post-con vs. Control. #p < 0.05 Post-con and Re Post-con vs. Pre-con. Values are means±SD.
Discussion
The main finding of the present study was that a single 5-min episode of ischemia and reperfusion to the hind limb of rabbit applied immediately prior to the onset of myocardial reperfusion protects the heart against reperfusion injury. The reduction in infarct size with remote postconditioning was confirmed by a decrease in plasma CK activity at the end of reperfusion. In addition, remote postconditioning attenuated neutrophil accumulation in the area at risk of infarction. Furthermore, remote postconditioning was considered contributing to a reduction in plasma MDA activity indicative of oxidant-mediated injury to membrane phospholipids. These data suggested that remote postconditioning could reduce the generation of reactive oxygen species and attenuate the oxidant-mediated injury. The cardioprotective effect achieved with remote postconditioning was similar to that observed in ischemic preconditioning and postconditioning. Therefore, strategically modifying early reperfusion events by transiently occluding and reperfusing the limb provides powerful cardioprotection comparable to a pretreatment strategy such as IPC.
Previous studies have shown that generation of abundant reactive oxygen species during early reperfusion contributes to tissue injury secondary to ischemia and reperfusion. A significant burst of oxygen-derived free radicals occurs within the first minutes of reperfusion and peaks 4–7 min after the onset of reperfusion and increased free radical generation is still detectable during later periods of reperfusion Citation17. Oxygen radicals trigger the release of pro-inflammatory mediators, transcription factors such as NF-κB, and stimulate the surface expression of adhesion molecules on coronary vascular endothelium Citation18. Sun et al. demonstrated that prevention of cardiomyocyte damage by hypoxic postconditioning is associated with a reduction in the generation of reactivr oxygen species Citation19. Kin et al. recently reported that cardioprotection produced by postconditioning at the onset of reperfusion may be mediated by inhibiting oxidant generation and oxidant mediated injury. Remote postconditioning induced by a single 5-min episode of femoral artery occlusion and reperfusion applied immediately before the onset of coronary artery reperfusion was associated with lower levels of MDA-reactive products Citation20. This result suggested that remote postconditioning attenuated lipid peroxidation in the area at risk of myocardium. And this mechanism was also suggested as an important component of the cardioprotection provided by remote postconditioning. Remote postconditioning may have limited the delivery of substrate and thereby directly reduced the generation of oxygen-derived free radicals during the early stage of myocardium reperfusion. However, the source of oxidants has not yet been elucidated. Further studies are needed to clarify the source and the species of free radicals attenuated by remote postconditioning.
In the present study, neutrophil accumulation (tissue MPO activity) in the area at risk of myocardium in the ischemic preconditioning and postconditioning and remote postconditioning groups was significantly reduced compared to untreated control group. Previous studies Citation21, Citation22 have shown that neutrophils migrate into and accumulate in ischemic-reperfused myocardium. The process of neutrophil accumulation within the area at risk begins at the onset of reperfusion and continues for 24 h. Moreover, numerous studies Citation23, Citation24 have demonstrated that a reduction in neutrophil activity and accumulation is associated with a reduction in infarct size. However, the observation that remote postconditioning decreases neutrophil accumulation in the present study does not determine whether the reduction in infarct size was a result of attenuation of neutrophil-mediated injury (secondary to neutrophil-derived oxidants and cytokine, i.e. a cause), or whether this reduced accumulation was simply a pedestrian response to less vascular and cardiomyocyte injury (i.e. an effect).
The present study focused on the limb as the remote organ; therefore, it may have important clinical implications, because it would be practical to create limb ischemia by placing a tourniquet on an extremity for a brief period of time before myocardial reperfusion. For example, it can be initiated with thrombolysis and may be applied during percutaneous transluminal coronary angioplasty (PTCA) and coronary artery bypass graft (CABG) to reduce ischemia-reperfusion injury.
As mentioned above, the present study did not identify the species of free radicals, nor its source, that were potentially attenuated by remote postconditioning. Secondly, the present study did not determine whether the radicals involved in remote postconditioning were liberated from the ischemic hind limb or other mechanisms, e.g. the activation of opioid and/or other g-protein-coupled receptors by remote postconditioning, generate free radicals in the heart resulting in cardioprotection. Finally, this study did not determine whether remote postconditioning reduces free radicals by modulating the release of triggers such as endogenous adenosine. We can hypothesize that endogenous adenosine during remote postconditioning may be attenuating the release of oxidants and cytokines by activated coronary vascular endothelium and myocytes, in addition to its well-known inhibitory effects on neutrophils. In addition, in the present study, we perform our experiment on the rabbit because it has sparse native collateral circulation, thereby acute coronary ligation results in rapid progression of injury. However, we did not investigate the effect of remote postconditioning on the myocardial microcirculation and coronary collateral circulation in the present study.
In conclusion, the present study, for the first time, reports that limb ischemic postconditioning reduces myocardial infarct size in an in vivo rabbit model and the mechanism involved might be a reduced oxidant burden from unidentified cell sources. The phenomenon of limb postconditioning implies that ischemic postconditioning has systemic effects to protect distant organ undergoing ischemia-reperfusion injury. Further studies are necessary to investigate whether brief episodes of ischemia to a remote other organ could be cardioprotective and to determine other mechanism involved.
This study was supported by grants from Health Department of Shandong Province (2003 HZ105) and Natural Science Foundation of Shandong Province (Y2005C70), China.
References
- Murry CE, Jennings RB, Reimer KA. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986; 74: 1124–36
- Cohen MV, Yang XM, Downey JM. Conscious rabbits become tolerant to multiple episodes of ischemic preconditioning. Circ Res 1994; 74: 998–1004
- Hagar JM, Hale SL, Kloner RA. Effect of preconditioning ischemia on reperfusion arrhythmias after coronary artery occlusion and reperfusion in the rat. Circ Res 1991; 68: 61–8
- Kita H, Miura T, Tsuchida A, Hasegawa T, Shimamoto K. Suppression of reperfusion arrhythmias by preconditioning is inhibited by an ATP-sensitive potassium channel blocker, 5-hydroxydecanoate, but not by protein kinase C blockers in the rat. J Cardiovasc Pharmacol 1998; 32: 791–7
- Lu H, Remeysen P, De Clerck F. The protection by ischemic preconditioning against myocardial ischemia- and reperfusion-induced arrhythmias is not mediated by ATP-sensitive potassium channels in rats. Cor Art Dis 1993; 4: 649–57
- Miura T, Ishimoto R, Sakamoto J, Tsuchida A, Suzuki K, Ogawa T, et al. Suppression of reperfusion arrhythmia by ischemic preconditioning in the rat: Is it mediated by the adenosine receptor, prostaglandin or bradykinin receptor?. Basic Res Cardiol 1995; 90: 240–6
- Liu YG, Downey JM. Ischemic preconditioning protects against infarction in Lirat-heart. Am J Physiol 1992; 263: H1107–11
- Lott FD, Guo P, Toombs CF. Reduction in infarct size by ischemic preconditioning persists in a chronic rat model of myocardial ischemia–reperfusion injury. Pharmacology 1996; 52: 113–8
- Zhao ZQ, Corvera JS, Halkos ME, Kerendi F, Wang NP, Guyton RA, et al. Inhibition of myocardial injury by ischemic postconditioning during reperfusion: Comparison with ischemic preconditioning. Am J Physiol Heart Circ Physiol 2003; 285: H579–88
- Vinten-Johansen J, Zhao ZQ, Zatta AJ, Kin H, Halkos ME, Kerendi F. Postconditioning. A new link in nature's armor against myocardial ischemia-reperfusion injury. Basic Res Cardiol 2005; 100: 295–310
- Tsang A, Hausenloy DJ, Mocanu MM, Yellon DM. Postconditioning: A form of “modified reperfusion” protects the myocardium by activating the phosphatidylinositol 3-kinase-akt pathway. Circ Res 2004; 95: 230–2
- Przyklenk K, Bauer B, Ovize M, Kloner RA, Whittaker P. Regional ischemic preconditioning protects remote virgin myocardium from subsequent sustained coronary occlusion. Circulation 1993; 87: 893–9
- Gho BC, Schoemaker RG, van den Doel MA, Duncker DJ, Verdouw PD. Myocardial protection by brief ischemia in noncardiac tissue. Circulation 1996; 94: 2193–200
- Li G, Labruto F, Sirsjo A, Chen F, Vaage J, Valen G. Myocardial protection by remote preconditioning: The role of nuclear factor kappa-B p105 and inducible nitric oxide synthase. Eur J Cardio-thorac Surg 2004; 26: 968–73
- Kharbanda RK, Mortensen UM, White PA, Kristiansen SB, Schmidt MR, Hoschtitzky JA, et al. Transient limb ischemia induces remote ischemic preconditioning in vivo. Circulation 2002; 106: 2881–3
- Kerendi F, Kin H, Halkos ME, Jiang R, Zatta AJ, Zhao ZQ, et al. Remote postconditioning. Brief renal ischemia and reperfusion applied before coronary artery reperfusion reduces myocardial infarct size via endogenous activation of adenosine receptors. Basic Res Cardiol 2005; 100: 404–12
- Bolli R, Patel BS, Jeroudi MO, Lai EK, McCay PB. Demonstration of free radical generation in “stunned” myocardium of intact dogs with the use of the spin trap a-phenyl-N-tertbutyl nitrone. J Clin Invest 1988; 82: 476–85
- Miki T, Cohen MV, Downey JM. Failure of N-2- mercaptopropionyl glycine to reduce myocardial infarction after 3 days of reperfusion in rabbits. Basic Res Cardiol 1999; 94: 180–7
- Sun HY, Wang NP, Kerendi F, Halkos M, Kin H, Guyton RA, et al. Hypoxic postconditioning reduces cardiomyocyte loss by inhibiting ROS generation and intracellular Ca2 + overload. Am J Physiol Heart Circ Physiol 2005; 288: H1900–8
- Kin H, Zhao ZQ, Sun HY, Wang NP, Corvera JS, Halkos ME, et al. Postconditioning attenuates myocardial ischemia-reperfusion injury by inhibiting events in the early minutes of reperfusion. Cardiovasc Res 2004; 62: 74–85
- Chatelain P, Latour JG, Tran D, de Lorgeril M, Dupras G, Bourassa M. Neutrophil accumulation in experimental myocardial infarcts: Relation with extent of injury and effect of reperfusion. Circulation 1987; 75: 1083–90
- Dreyer WJ, Michael LH, West MS, Smith CW, Rothlein R, Rossen RD, et al. Neutrophil accumulation in ischemic canine myocardium: Insights into time course, distribution, and mechanism of localization during early reperfusion. Circulation 1991; 84: 400–11
- Curtis WE, Gillinov AM, Wilson IC, Bator JM, Burch RM, Cameron DE, et al. Inhibition of neutrophil adhesion reduces myocardial infarct size. Ann Thorac Surg 1993; 56: 1069–72
- Hartman JC, Anderson DC, Wiltse AL, Lane CL, Rosenbloom CL, Manning AM, et al. Protection of ischemic/reperfused canine myocardium by CL18/6, a monoclonal antibody to adhesion molecule ICAM-1. Cardiovasc Res 1995; 30: 47–54