
ABSTRACT
This paper presents the results of a multi-proxy, palaeolimnological investigation of Lough Inchiquin, a large lake located immediately south of the Burren, a distinctive karst landscape in Co Clare, western Ireland which has been exploited since the Neolithic period. Chironomid sub-fossil analysis and lake sediment geochemistry provides an opportunity to infer lake response to prehistoric farming activity. The results of these new analyses are contextualised by pollen and loss-on-ignition evidence from the same lake catchment. The combined data of chironomid, loss-on-ignition (LOI550, LOI950 and non-combustible fraction (NCF)), organic geochemistry (δ13Corg, δ15N, Corg:N ratio), inorganic geochemistry (Ti, Fe, Mn, S and δ18O) and fossil pollen has allowed for an in-depth understanding of the palaeolimnological and palaeoenvironmental changes from the Mesolithic to the Late Bronze Age (c. 4590–660 BC) in the catchment of Lough Inchiquin. The data highlight the increasing ecological impact of anthropogenic activity through time and provide well-dated evidence for the initiation of Holocene soil erosion from the Burren.
Introduction
Lake sediments are an excellent means through which to investigate human-environment interactions as the accumulation of sediment often provides a continuous record of environmental change through time. The current study aims to investigate prehistoric farming impacts within the catchment of Lough Inchiquin, Co. Clare, Ireland, using a multi-proxy framework which combines chironomid analysis with organic and inorganic geochemistry, contextualised by a previously published high-resolution palynological record from the same lake catchment (Spencer et al. Citation2019). Spencer et al. (Citation2019) highlighted a discrepancy between the archaeological record, which suggested the most intensive human activity occurred during the Chalcolithic to Early Bronze Age (c. 2500–1600 BC), and the palaeoenvironmental results, which saw the Late Bronze Age (c. 1210–560 BC) emerge as the period with the most intensive farming activity with a two-phased Landnam lasting c. 500 years. The current study will carry out a palaeolimnological assessment of the impact of prehistoric farming activity upon the lake system. No study of this sort has previously been reported on from the Burren.
In recent years palaeolimnological studies have focused increasingly upon the effects of human impact with many determining the first anthropogenic disturbance in multi-proxy records (Dubois et al. Citation2018). This is certainly true of chironomid studies which now often focus on the impact of changing land-use associated with human activity (Francis and Foster Citation2001; Heiri and Lotter Citation2003), rather than traditional climatic temperature reconstructions. This has led to successful applications of this methodology within archaeology (O'brien et al. Citation2005; Ruiz, Brown, and Langdon Citation2006; Taylor et al. Citation2013; Citation2017a; Citation2017b). There is a growing body of evidence which suggests that because many Irish lakes have been impacted by anthropogenic land-use change since the Neolithic, these impacted sites may not provide a robust climate signal via chironomid analysis, because anthropogenic land-use has become the dominant environmental control (Mckeown and Potito Citation2016; Potito et al. Citation2014; Chique et al. Citation2018). The addition of organic geochemistry, specifically δ13Corg, δ15N and Corg:N ratios, can provide information on nutrient loading in lakes as a result of agricultural practices and the in-wash of nitrogen enriched soil derived from animal waste (Perren et al. Citation2012) and soil erosion (Talbot Citation2001). In modern Irish lakes, δ15N values are typically higher in agriculturally active catchments than in non-impacted catchments (Woodward, Potito, and Beilman Citation2012). Such nutrient loading subsequently results in a shift in chironomid community composition as a response to changes in food availability, oxygen levels and/or overall lake productivity (Brooks et al. Citation2007). This study attempts to clarify the timing of anthropogenically-induced erosion of Burren soils by using inorganic geochemistry as the influx of catchment soil can result in increased concentrations of trace elements within lake sediments (Mackereth Citation1966; Engstrom and Wright JR Citation1984).
The combined data will include chironomid, loss-on-ignition (LOI550, LOI950 and non-combustible fraction (NCF)), organic geochemistry (δ13Corg, δ15N, Corg:N ratio), inorganic geochemistry (Ti, Fe, Mn, S and δ18O) and pollen. The results allow for an in-depth understanding of palaeoenvironmental and palaeolimnological changes, with a specific focus on anthropogenically-induced change from the Early Neolithic to Late Bronze Age in the catchment of Lough Inchiquin.
Study Site
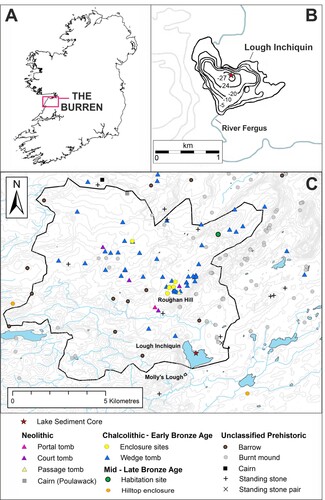
Lough Inchiquin is located immediately south of the Burren (52°57′20″N, 9°5′16″W) at an elevation of 35 m above sea level, covering c. 110 ha and with an average depth of 10.8 m (Cullinane Citation2012). It is hydrologically open with the River Fergus, which drains a considerable amount of the Burren plateau (Drew Citation1988), entering at the northern-most shore and exiting to the south. Previous palaeolimnological studies of Lough Inchiquin have concentrated on the Late Glacial and Early Holocene using a sediment core taken near the southern shore (Diefendorf et al. Citation2006; Citation2008). Water residence time has been calculated as c. 1 month (Diefendorf et al. Citation2008). Forty percent of the catchment is currently classed as pastureland (Waemere Citation2005). The lake catchment has been calculated at 93.2 km2 which encompasses an area rich in archaeological sites (). The Chalcolithic – Early Bronze Age (c. 2500–1600 BC) was a focus of activity on the nearby ridge of Roughan Hill (c. 3 km north of the lake) where wedge tombs, field systems and house sites have been found. The thin rendzinas currently established on Roughan Hill are especially suited to grazing (Finch, Culleton, and Diamond Citation1971).
Figure 1 (A) The location of the Burren within Ireland. (B) Bathymetry map of Lough Inchiquin with depth contours shown in metres. (C) The distribution of archaeological sites within the study area. The catchment of Lough Inchiquin is indicated by the black outline. Archaeological sites are labelled and divided chronologically.
Materials and Methods
Coring and Chronology
A 10.6 m long sediment core (LIQ1) was extracted from the deepest part of Lough Inchiquin (52°57′20″N, 9°5′16″W) at a water depth of 30 m, using a Usinger piston corer in June 2015. Suitable plant material of terrestrial origin was selected for 14C AMS dating carried out at the 14Chrono Centre, Queen's University Belfast, UK. An age-depth model was established using Bacon v. 2.2 Bayesian age-depth modelling software (Blaauw and Christen Citation2011), utilising calibration curve IntCal 13 (Reimer et al. Citation2013) and was presented in Spencer et al. (Citation2019). Dates are expressed as calibrated years BC and rounded to the nearest ten years.
Chironomid Analysis
Chironomid analysis was carried out on 58 samples corresponding with pollen sub-sampling, at 4 cm intervals, between 788–444 cm, an interval which dates from the Mesolithic to Late Bronze Age. Analysis followed standard procedures outlined in Walker (Citation2001). Between 0.5 and 2 ml of sediment was deflocculated in a 10% KOH solution and heated for 30 min at 30°C. The sediment was sieved through a 90 µm mesh and the resultant solution poured into a Bogorov plankton counting tray. Samples were sorted using a Leica MZ12s microscope at x10 – x40 magnification. A minimum of 50 chironomid head capsules, or whole mentum equivalents (WME), per sample (average 65) were retrieved with fine forceps and permanently mounted on slides with Entellan. A Motic B3 Professional Series compound microscope at x100 – x400 magnification was used to identify the chironomid subfossils. Taxa were identified to genus, sub-genus or species-type following Brooks et al. (Citation2007), Wiederholm (Citation1983), and Rieradevall and Brooks (Citation2001). A total of 84 taxa were identified in LIQ1 with 48 designated as common taxa i.e. taxa present in at least two samples with an abundance of ≥2% in at least one sample (Quinlan and Smol Citation2001). A chironomid percentage diagram was constructed in C2 v. 1.7.7 (Juggins Citation2003). Zonation into chironomid assemblage zones (CAZ's) was established using Psimpoll (Bennett Citation2009–Citation1993), based on optimal sum-of-squares partitioning, with the statistical significance of zones determined using BSTICK (Bennett Citation1996).
Geochemical Analyses
Stable isotope ratios of nitrogen (δ15N) and carbon (δ13Corg) from organic matter, and oxygen (δ18O) from carbonates were carried out at the University of California Davis Stable Isotope Lab. An Elementer Vario EL Cube interfaced to a PDZ Europa 20–20 isotope ratio mass spectrometer (IRMS) was used to measure the δ15N values of a non-acidified portion of the samples. Oxygen (δ18O) from carbonates were run on a ThermoFisher GasBench II device coupled to a Delta V Plus mass spectrometer. Carbon (δ13Corg) isotopes from organic matter were carried out at Amherst College using a Piccaro δ13C-CO2 analyser, connected to a Costech elemental analyser (EA). Following Harris, Horwáth, and Van Kessel (Citation2001) samples were dried, pulverised with a mortar and pestle and passed through a 200 μm sieve. Homogenised material was weighed into silver capsules, and carbonate removed via HCl-fumigation. Corg:N ratios were calculated as the mass ratio of organic carbon to total nitrogen elemental concentrations analysed at Wesleyan University, Connecticut using an EA. Organic geochemistry was carried out on samples at 10 cm intervals with results interpolated to correspond with chironomid sub-sampling for subsequent statistical tests.
Elemental analysis was carried out at every mm and averaged for each cm between 788–444 cm. An ITRAX™ Core Scanner, at the University of Massachusetts, was used with the following settings: voltage of 50 kV, current of 50 mA, exposure time of 1700ms, step size of 1000 microns, XRF exposure time of 10 s, XRF voltage of 30 kV, XRF current of 55 mA and a molybdenum tube. Results were produced as a relative abundance measured in counts per second (cps).
Pollen and Loss-on-ignition
Percentage curves for selected indicative pollen taxa are presented here to contextualise palaeolimnological data. The pollen taxa used include: Plantago lanceolata, used as an indicator of disturbance (Behre Citation1981) often linked to pastoral farming; total non-arboreal pollen indicative of pasture (NAPp) as defined in Spencer et al. (Citation2019); cereal-type pollen (≥40 µm); Myriophyllum spicatum (water milfoil), an aquatic plant of nutrient enriched water and Pediastrum, an algae. Full pollen results for Lough Inchiquin were presented in Spencer et al. Citation2019. Loss-on-ignition (LOI) analysis was carried out at 4 cm intervals with samples dried for 24 h at 101°C and ashed at 550°C (4 h) and 950°C (2 h) (Heiri, Lotter, and Lemcke Citation2001). Estimates for the percentage organic matter and inorganic carbonate content of the samples were determined by weight, and the non-combustible fraction (NCF) calculated from these results.
Statistical Analysis
Ordination analyses were performed using Canoco v. 5.0 (Ter Braak and Šmilauer Citation1997) on square-root transformed chironomid percentage data for all common taxa. Changes in chironomid community composition through time were analysed using principal components analysis (PCA), an unconstrained linear ordination method, and redundancy analysis (RDA), a constrained linear ordination method. The linear-based ordination was chosen by assessing the gradient length (0.7 standard deviations (SD); linear model recommended) of the species composition along the first axis (Birks Citation1995).
A constrained RDA was carried out using 18 environmental variables (LOI550, LOI950, NCF, δ15N, δ13C, Corg:N, δ18O, S, Ti, Mn, Fe, NAPp, Plantago lanceolata, cereal-type, Nymphaea, Pediastrum, Myriophyllum and Botryococcus). Palynological variables associated with land-use were chosen along with geochemical variables which can indicate lake productivity and erosional activities within lake systems. Variables with high co-linearity (>20x) were removed, which led to the rejection of LOI550 (co-linear with Corg:N) and NAPp (co-linear with δ15N), with statistical analysis only bringing forward the most powerful predictors. The statistical significance of each variable (p ≤ 0.05) was assessed using Monte Carlo permutation tests (499 unrestricted permutations) and P value correction including false discovery rate, Holm, and Boneforri tests. Insignificant variables (δ13C, Fe, Mn, S, Botryoccocus and Nymphaea) were removed from further analysis. All significant environmental variables were passively plotted in the PCA to investigate potential associations with chironomid community change. For RDA, environmental variables were forward-selected in order of decreasing variance as long as each additional variable remained a statistically significant predictor of chironomid assemblage data. Pearson correlations of the environmental variables were carried out, which demonstrated that δ15N was highly correlated with other land-use variables and was likely acting as an analogue for land-use in the RDA.
Results
The Lough Inchiquin chironomid dataset spans the Mesolithic to the Late Bronze Age (c. 4590–660 BC). Chironomid data were divided into three statistically significant CAZ; CAZ-1, CAZ-2 and CAZ-3 (). The results of geochemical analysis are presented in . Selected pollen indicator curves are presented in (see Spencer et al. Citation2019 for full pollen data) along with key chironomid and geochemical curves.
Figure 2. Relative percentage chironomid abundance plotted against sediment depth and corresponding age axis and divided into statistically significant chironomid assemblage zones (CAZ). Taxa arrangement follows RDA Axis 1 scores (lake trophic status). The Neolithic Landnam is highlighted in light blue and the period of most significant erosion is highlighted in orange.

Figure 3. Organic and inorganic geochemistry results plotted by age (cal. years BC) with a corresponding depth axis. Grey lines indicate the full sampling resolution of each analysis while black lines indicate values at chironomid sampling depths. Zonation is based on chironomid assemblage zones (CAZ). Archaeological time periods are shown on the right-hand side. The Neolithic Landnam is highlighted in light blue and the period of most significant erosion is highlighted in orange.
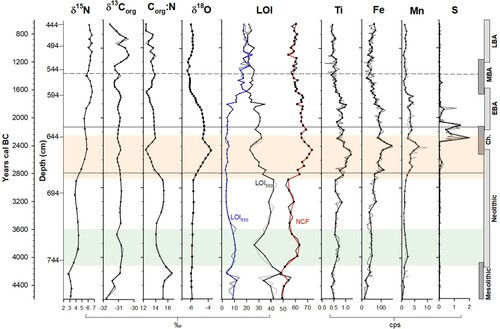
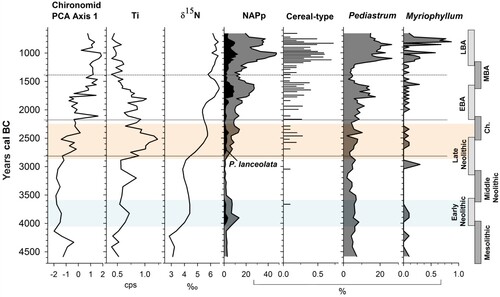
Figure 4. Composite diagram of chironomid, organic geochemistry, inorganic geochemistry and selected pollen data, plotted to a time scale of cal. years BC. Zonation is based on chironomid assemblage zones (CAZ). Archaeological time periods are shown on the right. The Neolithic Landnam is highlighted in light blue and the period of most significant erosion is highlighted in orange. Pollen curves from Spencer et al. (Citation2019).
CAZ-1 788–680 cm; c. 4590–2810 BC (Mesolithic – Late Neolithic)
The highest levels of Micropsectra insignilobus-type (14%), Heterotanytarsus (11%), Pagastiella and Tanytarsus lugens-type were observed in this zone. Well represented taxa include Sergentia coracina-type (14%), Dicrotendipes nervosus-type (20%), Tanytarsus undif. (16%) and Tanytarsus pallidicornis-type (10%). Only low levels of Chironomus anthracinus-type are observed (averaging 1%) increasing slightly between 748–716 cm. In the upper half of the zone Cricotopus intersectus-type increases to a maximum of 14%.
δ15N and δ13Corg values fluctuate but in general rise to end the zone at 4.1‰ and −30.9‰, respectively, while C:N ratios decrease in the middle of the zone. Between 756–732 cm, LOI550 values drop while LOI950 rise. Levels of δ18O, Mn and S are relatively constant while Ti and Fe both increase. NAPp is generally low but expands significantly between 744–720 cm. Cereal-type is recorded in 2 spectra.
CAZ-2 676–636 cm; c. 2810–2170 BC (Late Neolithic – Chalcolithic)
In this zone notable increases occur in Chironomus anthracinus-type (9%), Glyptotendipes pallens-type (8%) and Psectrocladius sordidellus-type (8%). Microtendipes pedellus-type, Parakieferiella bathophila-type, and Polypedilum nubesculosum-type increase to maxima of 7%, 5%, and 8%, respectively. Corynoneura edwardsi-type, Tanytarsus undif. and Tanytarsus mendax-type also increase. Sergentia coracina-type remain well-represented (average 12%) while Heterotanytarsus average 2%.
δ15N values increase to 5.7‰. LOI550 values decrease while NCF increase and LOI950 remains relatively constant. δ18O, Ti, Fe and Mn express similar trends and peak in the middle of this zone, while S increases sharply at the end of the zone. NAPp increases to a maximum of 14.0% by 648 cm (lanceolata at 9.8%) remaining high for the rest of the zone with cereal-type continuously present.
CAZ-3 632–444 cm; c. 2170–660 BC (Early – Late Bronze Age)
CAZ-3 was further subdivided at 548 cm into two subzones which, while not statistically significant, marks the approximate beginnings of the Late Bronze Age and highlights notable qualitative changes in this period.
Subzone 3a 632–552 cm; c. 2170– 1380 BC (Early – Middle Bronze Age)
Chironomus anthracinus-type, Microtendipes pedellus-type, and Cricotopus intersectus-type all increase in representation in this subzone to 10%, 12% and 13%, respectively. Tanytarsus undif., Tanytarus pallidicornis-type, and Tanytarsus mendax-type remain well represented and Parakieferiella bathophila-type reaches 9%. Low levels of Stictochironomus and Endochironomus impar-type are present. Dicrotendipes nervosus-type and Sergentia coracina-type remain well-represented. Only low levels of Pagastiella, Micropsectra insignilobus-type and Heterotanytarsus are now observed.
δ15N values increase to 6.7‰ (585 cm). LOI550 and NCF decrease overall while LOI950 increases. δ18O, Ti, Fe and Mn all maintain a period of high levels before steadily declining. Sulphur also maintains high levels initially before declining to former low levels. NAPp rises substantially to 27.5% by 584 cm (lanceolata at 8.5%) before declining to end the zone at 11.3%. Cereal-type is consistently present. Pediastrum increases to 14.3% by 604 cm before also decreasing.
Subzone 3b 548–444 cm; c. 1380–660 BC (Middle – Late Bronze Age)
The highest representations of Microtendipes pedellus-type (maximum 20%) and Crictotopus intersectus-type (maximum 21%) are observed in this zone and Chironomus anthracinus-type increases to a maximum of 11%. Other prevalent taxa include Synorthocladius (maximum 8%), Stempellina (maximum 10%), Ablabesmyia (maximum 9%), and Tanytarsus undif. (average 10%). Dicrotendipes nervosus-type increases to 11% while Paratanytarsus reaches a profile maximum of 12%.
δ15N values remain consistently high averaging 6.3‰, while δ13Corg peaks twice in this subzone. LOI950 increases slightly. NAPp expands rapidly to 47.5% at 500 cm with P. lanceolata contributing up to 10.5%. A second peak is recorded at 456 cm (NAPp at 37.4% with P. lanceolata contributing 4.0%). Cereal-type pollen is consistently present achieving a profile maximum of 0.7%. Pediastrum expands to 26.5% by 512 cm and Myriophyllum, at low levels for much of the profile, also increases significantly.
Ordination Analysis
a and 5b present the passive associations of the chironomid data with ten statistically significant environmental variables (δ15N, Corg:N, LOI950, NCF, Ti, δ18O, Plantago lanceolata, cereal, Pediastrum and Myriophyllum). All variables associated with increased land use and nutrient loading (δ15N, Plantago lanceolata, cereal, Pediastrum and Myriophyllum) and LOI950 group together on the right of Axis 1 while Corg:N is directly opposite. The Ti, δ18O, and to a lesser extent NCF vectors are almost perpendicular to these. In the samples bi-plot (a) those from CAZ-1 are concentrated where Corg:N values are high while those from CAZ-2 are associated with high Ti, δ18O and NCF values. The samples from CAZ-3 are predominantly concentrated to the right of Axis 2 where δ15N and land-use indicator values are high.
Figure 5. PCA bi-plots. (A) Chironomid samples with environmental variables. (B) Chironomid taxa with environmental variables. Chironomid taxa are abbreviated as follows ABLA = Ablabesmyia; CHIA = Chironomus anthracinus-type; CLAD = Cladopelma; CLAM = Cladotanytarsus mancus-type; COAR = Corynoneura arctica-type; COCO = Corynoneura coronata-type; COED = Corynoneura edwardsi-type; CRCY = Cricotopus cylindraceus-type; CRIN = Cricotopus intersectus-type; CRSY = Cricotopus sylvestris-type; DICR = Dicrotendipes nervosus-type; ENDA = Endochironomus albipennis-type; ENDI = Endochironomus impar-type; ENDT = Endochironomus tendens-type; EUKC = Eukieferiella claripennis-type; GLYP = Glyptotendipes pallens-type; GLYS = Glyptotendipes severini-type; GUTT = Guttipelopia; HETE = Heterotanytarsus; HETG = Heterotrissocladius grimshawi-type; LAUT = Lauterborniella; LIMN = Limnophyes; MAI = Micropsectra insignilobus-type; MICP = Microtendipes pedellus-type; MONO = Monopelopia; PAGA = Pagastiella; PVAR = Parachironomus varus-type; PARM = Paramerina; PARA = Paratanytarsus; PTNU = Paratendipes nudisquama-type; PABA = Parakiefferiella bathophila-type; PHFL = Phaenopsectra flavipes-type; POLN = Polypedilum nubesculosum-type; POSO = Polypedilum sordens-type; PROC = Procladius; PSEU = Pseudochironomus; SERG = Sergentia coracina-type; STEM = Stempellina; S/Z = Stempellinella; STIC = Stictochironomus; SYNO = Synorthocladius; TAN = Tanytarsus undifferentiated; TALA = Tanytarsus lacetescens-type; TALU = Tanytarsus lugens-type; TANM = Tanytarsus mendax-type; TANP = Tanytarsus pallidicornis-type; THIE = Thienemannimyia.
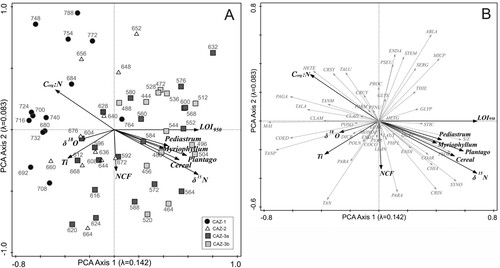
The RDA (a and 6b) has an eigenvalue of 0.101 (10.1%) for Axis 1 and 0.038 (3.8%) for Axis 2, using two statistically significant forward-selected variables (δ15N and Ti). δ15N shows the strongest relationship to RDA Axis 1 (t = 9.06) while Ti has the strongest relationship with Axis 2 (t = 7.24). δ15N exhibits the highest explanatory power followed by Ti. The analysis demonstrates the chironomid community trajectory from a relatively unimpacted lake system in CAZ-1 to an increasingly impacted system by CAZ-3b, with a trend of movement towards high levels of δ15N. Additionally, samples from CAZ-2 are associated with high levels of Ti ( and .).
Figure 6. RDA bi-plots. (A) Chironomid species with forward-selected environmental variables (δ15N and Ti). (B) Chironomid taxa with forward-selected environmental variables. Chironomid abbreviations as in .
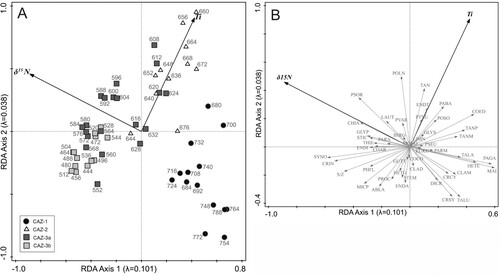
Table 1. Summary statistics for PCA and RDA (*p < 0.05).
Table 2. Partial RDAs for forward selected variables.
Discussion
Minimally impacted lake system from the Mesolithic to Middle Neolithic (c. 4590–3110 BC)
The pollen evidence suggests a predominantly wooded landscape with little evidence for open grassland during the Mesolithic. The combined chironomid and geochemical data highlight the pre-impacted state of Lough Inchiquin prior to the initiation of farming activity. The lake system was oligotrophic with low levels of δ15N prior to c. 4060 BC. Particularly high levels of Micropsectra insignilobus-type and Heterotanytarsus demonstrate a lack of nutrients and Tanytarsus lugens-type, a taxon associated with the profundal of oligotrophic lakes, was present until c. 3980 BC (Brooks et al. Citation2007).
The higher NAPp (13.5%) recorded from c. 4060–3590 BC is reflective of an early farming Landnam in the Early Neolithic at Lough Inchiquin during which δ15N values increased to 4.4‰ (). Nutrient enrichment of the lake is suggested by an increased representation of Myriophyllum. The forward-selected RDA demonstrates a slight response to the Neolithic Landnam with samples having moved along the δ15N vector. Concurrent with this was a decrease in the Corg:N ratio which is suggestive of increased autochthonous productivity (Talbot Citation2001). There was a slight decrease in δ13Corg but the data overall suggest that, as in Taylor et al. (Citation2017b), δ13Corg is not showing a uniform response to land-use. Within the catchment, Early Neolithic monuments associated with the Landnam consist of the excavated Parknabinnia court tomb on Roughan Hill, two additional and one possible court tomb, a court/portal tomb and the Ballycasheen portal tomb while the well-known Poulnabrone portal tomb is located just 1.6 km outside the catchment (Jones Citation2016; Citation2019). Activity within the catchment continued into the Middle Neolithic evidenced by the continued use of the Parknabinnia and Poulnabrone megaliths, habitation evidence at Teeskagh, and the construction of the Poulawack Linkardstown-type cist (Jones Citation2019; Gibson Citation2016).
No significant overall change to the chironomid composition occurred with the onset of farming but slight increases in Tanytarsus pallidicornis-type and Tanytarsus mendax-type (both indicative of productive littoral environments) during the Landnam hint at some response to Neolithic farming. Taxa associated with macrophytes, Dicrotendipes nervosus-type and Glyptotendipes pallens- type, increased and low levels of eutrophic taxa were observed. Albeit limited, this indicates the first anthropogenically-induced impact upon the lake system. The chironomid community at Lough Inchiquin did not respond to early farming to the same extent as seen in previous studies in western Ireland where eutrophic taxa were abundant in the stratigraphy during Early Neolithic Landnam (Taylor et al. Citation2013; Citation2017a; Citation2017b). This is likely due to the size and morphometry of Lough Inchiquin. Previous studies were from smaller, shallower, closed systems and so more readily impacted by small-scale farming.
The decrease in NAPp from c. 3510 BC, likely reflecting woodland regeneration as a result of reduced farming pressure, coincides with a decline in δ15N values and an increase in Corg:N, suggestive of a less productive lake system (Woodward, Potito, and Beilman Citation2012). Inorganic geochemistry results were relatively stable during this period but a slight increase in Ti, Fe and Mn values during the Landnam could be linked to the small scale in-wash of soil due to initial woodland clearances (Cohen Citation2003). A decrease in LOI550 supports this, having a strong negative correlation with the land use variables, as does an increase in NCF.
Increased nutrient enrichment during the Late Neolithic, Chalcolithic and Early Bronze Age (c. 3110–1600 BC)
A re-initiation of farming activity in the Late Neolithic occurred with an expansion of NAPp from c. 2890 BC to levels above that of the Neolithic Landnam. Cereal-type pollen recorded from c. 2690 BC suggests an arable component to the farming and both decreased LOI550 and low pollen concentration values indicate an increase in sedimentation reflecting erosion of mineral material. δ15N increased by >1‰ when compared with the Early Neolithic Landnam which suggests that the increasing intensity of farming activity in this later period was having a greater impact on the lake system. Farming intensified in the Early Bronze Age with NAPp reaching 26.8% (c. 1600 BC) occurring contemporaneously with an increase in δ15N values to 6.7‰ (c. 1630 BC). Combined with decreased Corg:N the data suggest increased lake productivity.
This nutrient-enriched environment caused a shift in the chironomid community with increasing macrophyte-dwellers and mesotrophic-eutrophic taxa from c. 2890 BC (Moller Pillot Citation2009; Brooks et al. Citation2007). Chironomus anthracinus-type, common in the profundal zone of deep productive lakes, increased from c. 2750 BC (Moller Pillot Citation2009; Brooks et al. Citation2007; Chique et al. Citation2018). This taxon reached a maximum of 10% by c. 1880 BC suggesting a further increase in lake productivity in the Early Bronze Age ().
Chironomid PCA Axis 1 values are heightened in this period while the forward-selected RDA demonstrates a shift in samples from this period along the δ15N gradient. Nutrient enrichment of the lake suggests that activity was occurring locally which is supported by the high representation of NAPp with a limited dispersal capacity. Archaeological evidence for activity at this time is abundant and varied. At the Parknabinnia court tomb interments continued until the first half of the 3rd millennium, and after an interlude of approximately three centuries, the monument was reused for burials in the Chalcolithic, and then again in the Early Bronze Age (Schulting et al. Citation2012; Snoeck et al. Citation2020). Chalcolithic and Early Bronze Age re-use also occurred at the Poulawack monument (Brindley and Lanting Citation1990), Teeskagh habitation site (Gibson Citation2016) and at Poulnabrone, where a single Early Bronze Age interment was placed (Lynch Citation2014). New burial cairns and wedge tombs were also constructed in the Chalcolithic, with a dense concentration located on Roughan Hill. Here, a distinct farming phase occurred between c. 2300–1500 BC where field systems and four enclosed habitation sites have been identified (Jones Citation2016; Jones Citation2019).
Soil erosion during the Late Neolithic, Chalcolithic and Early Bronze Age (c. 2870–1700 BC)
Significant increases in the relative levels of Ti, Fe and Mn were observed from c. 2870 BC (). Lake sediment geochemistry can be reflective of changes in lake catchments where high levels of trace elements can be indicative of erosion (Mackereth Citation1966). The relative levels of Ti, Fe and Mn follow the same general pattern, but only Ti was selected in the PCA and interactive forward-selected RDA ( and ). This suggests that it exhibits a significant, though somewhat minimal influence, accounting for 5.6% of the variation in the chironomid community composition. After peaking at 1.3 cps at c. 2480 BC, the values of Ti decreased slightly but remained elevated for the remainder of the Chalcolithic and into the Early Bronze Age. Titanium can be used as an erosion indicator in palaeolimnological studies as it is strictly of terrigenous origin and largely unaffected by post-depositional processes (Arnaud et al. Citation2012). Iron and manganese, in contrast, can be heavily affected by a number of independent environmental processes, lake conditions (redox and pH) and within-lake diagenesis (Engstrom and Wright JR Citation1984; Naeher et al. Citation2013). The interpretation of sulphur is similarly complicated by diagenetic processes which can affect its preservation in lake sediments (Cohen Citation2003; Luther et al. Citation2003; Holmer and Storkholm Citation2001). While generally low S levels were elevated between c. 2330–1840 BC, following the maxima in Fe and Mn. As the preservation of sulphur in lake sediment is linked to reduction processes and the levels of iron available (Holmer and Storkholm Citation2001), it is proposed that these elevated values occurred during a period of anoxic benthic conditions in Lough Inchiquin potentially caused by enhanced sedimentation and subsequent eutrophication of the lake system.
Enriched levels of δ18O (maximum of −3.6‰), occur contemporaneously with the increased Ti values, suggesting the influx of terrigenous material in the form of weathered limestone bedrock (Frentzel Citation2017) (). Carboniferous limestone, of which the Burren landscape is formed, has a typical δ18O value of −3‰ to −1‰ (Diefendorf et al. Citation2008; Grossman Citation1994) with the δ18O values in the Lough Inchiquin data moving towards that level at this time. This allochthonous input is further supported by high NCF values (73.5%). The combined data suggest that the beginnings of substantial erosion occurred contemporaneously with the re-initiation of farming activity in the Late Neolithic. The consequences likely included increased sedimentation and productivity in the littoral zone, a change in benthic substrate conditions, increased particle grain size, and decreased oxygen conditions. The composition of the chironomid community clearly changed at this time with samples from this period distinctly clustered towards the Ti vector on the forward-selected RDA (a). Taxa with a preference for sandy sediments, Polypedilum nubesculosum-type, Parakieferiella bathophila-type, Endochironomus impar-type and Stictochironomus (Moller Pillot Citation2009; Brooks et al. Citation2007), all increased suggesting this material influx created more favourable conditions. Substantial increases were also seen in Tanytarsus mendax-type, Tanytarsus pallidicornis-type and Corynoneura edwardsi-type which are characteristic of a productive littoral environment (Brooks et al. Citation2007; Chique et al. Citation2018).
It is proposed that substantial erosion into Lough Inchiquin occurred between c. 2870–2270 BC with continued erosion reflected in the Ti curve until at least c. 1700 BC. By this time, intensive pastoral and arable farming was well-established in the catchment with a gradual increase of anthropogenic indicators throughout the period. A significant increase in pastoral and arable activity occurred from c. 1800 BC. It is likely that this contributed to heightened lake productivity for the remainder of the record both directly, through erosion caused by the destabilisation of catchment soils and, perhaps to a lesser extent, the direct input of animal waste from farming. Although an initial surge in erosion may have been enhanced by substantial rainfall, climate evidence for a shift to cooler and wetter conditions c. 2200 BC is contested in the Irish record (Baillie and Mcaneney Citation2015; Roland et al. Citation2014; Fitzpatrick Citation2015). It has been proposed, however, that if this shift did occur, then the relatively warm limestone region of the Burren, still used for winterage in the present day, and specifically the area of Roughan Hill, would have been advantageous for Chalcolithic communities (Jones Citation2019).
The farming communities that populated Roughan Hill between c. 2300–1550 BC probably contributed both to the evidence of farming and erosion identified within the Lough Inchiquin sediment. It is likely that erosion continued beyond c. 1700 BC (contributing to high levels of δ15N) but that the higher productivity of the lake, evidenced by the values of LOI950 and a lower minerogenic component (NCF), could have weakened the Ti erosion signal at this point in the record. Past research has led to a significant body of evidence for soil erosion. Increased sedimentation accumulation rates and elevated magnetic susceptibility in Molly's Lough (c. 415 m southwest of Lough Inchiquin) was interpreted as erosion between 2542–639 BC (Thompson Citation1997). Additional evidence has been found in caves across the Burren and on the slopes of Slieve Rua, Mullaghmore and Knockanes Mountain, predominantly dating to the Bronze Age (Moles, Moles, and Leahy Citation1999; Moles and Moles Citation2002; Drew Citation1983). Remnants of prehistoric soils have also been sealed by archaeological structures (Jones Citation2004). Evidence for later anthropogenically induced erosion has been found in grykes (rock fissures) at Gortaclare and Slieve Rua where colluvial deposition occurred in the Iron Age and medieval period, respectively (Feeser and O’connell Citation2009). Until the current study, however, there was a lack of conclusive evidence from lake sediments in this area. Previous investigations have focused on both climate change and terrestrial landscape change in the Inchiquin catchment through multi-proxy palaeolimnological analyses of lake sediment (Diefendorf et al. Citation2006; Citation2008). Diefendorf et al. (Citation2008) determined that carbon isotopes reflected landscape change from bare limestone bedrock to a forested ecosystem during the late Glacial to Early Holocene (up to c. 3590 BC). Similarly, climate and vegetation change during the Younger Dryas and Early Holocene at An Loch Mór, an island off the coast of the Burren region, was investigated by Holmes et al. (Citation2019), using carbon and oxygen isotopes. Clearly such change is of a greater magnitude than that identified in the current study, but this new geochemical dataset continues the story of landscape evolution in this region and highlights the anthropogenic influence. The current study has been the first to apply lake sediment geochemistry providing unambiguous evidence for the significant erosion of Burren soils from as early as the Late Neolithic as a result of human activity.
Farming intensification and increased lake productivity from the Middle to Late Bronze Age (c. 1600–660 BC)
The pollen record indicates that farming (pastoral with a lesser arable component) continued through the Middle Bronze Age (c. 1560–1320 BC) but less intensively than the preceding period (see reduced NAPp and cereal-type – ). Decreased lake productivity at this time is indicated by lower δ15N values, increased Corg:N, and a significant decrease in Pediastrum suggestive of reduced anthropogenic nutrient influx. This may indicate that farming activity occurred at a greater distance from the lake or on a smaller scale while an increase in LOI550 likely reflects the stabilisation of catchment soils which is supported by a reduced minerogenic signal (Ti at 0.7 cps).
Farming activity within the catchment of Lough Inchiquin increased substantially in the Late Bronze Age with a Landnam lasting c. 500 years recorded between c. 1210–740 BC (). Two periods of particularly intensive farming occurred (c. 1200–1000 BC and c. 900–750 BC) with NAPp reaching its highest representation (47.5%) in the record by c. 1000 BC presumably reflecting the significant expansion of species-rich grasslands. Maxima for cereal-type representation were recorded at c. 1070 BC (0.6%) and c. 830 BC (0.7%) with the data indicative of a largely pastoral landscape with a prevalence of small-scale arable farming. δ15N levels increased to a maximum of 6.6‰ by c. 1060 BC and remained above 5‰ for the remainder of the Late Bronze Age. It is proposed that the mechanism driving this increase in Lough Inchiquin was the input of nutrients from pastoral agriculture (Woodward, Potito, and Beilman Citation2012; Botrel, Gregory-Eaves, and Maranger Citation2014; Vander Zanden et al. Citation2005) combined with any continuing small-scale erosion.
The rapid and significant increase in algae and Myriophyllum from c. 1230 BC implies that the lake system was becoming more productive as a result of anthropogenic nutrient loading (Chase and Knight Citation2006; Chique et al. Citation2018). The presence of Chara oospores and shells in the Inchiquin sediments at this point (see Spencer et al. (Citation2019)) supports both increased productivity and increased biogenic calcium carbonate. δ18O and NCF levels remain depleted in comparison to the Late Neolithic to Early Bronze Age trend and so do not appear to indicate significant erosion at the same scale as seen earlier in prehistory, although this may be influenced by heightened LOI950. Significantly enriched δ13Corg in the Late Bronze Age corresponds with an expansion of Pediastrum (>20%) from c. 1230 BC. As the δ13Corg of algae is −25‰ to −30‰ (Anderson et al. Citation2008), a high abundance in the lake may have caused the δ13Corg of the lake sediment to move towards this range. The values of δ13Corg and Corg:N were examined in an attempt to apportion the source of δ13Corg but no systematic relationship could be identified.
Chironomid response follows the catchment changes and increased lake productivity evident in the pollen record and geochemistry. The chironomid community composition during the Middle to Late Bronze Age was dominated by eutrophic taxa including Cricotopus intersectus-type, Microtendipes pedellus-type and Chironomus anthracinus-type (Brooks et al. Citation2007; Moller Pillot Citation2009, Citation2013) which reach maximum representations in the profile (). Increases in Stempellinella/Zavrelia and Tanytarsus undifferentiated indicate there was a productive littoral environment which could support the development of macrophytes (expansion of Myriophyllum - ) and subsequently taxa such as Dicrotendipes nervosus-type, Paratanytarsus and Ablabesmyia (Brooks et al. Citation2007; Vallenduuk and Pillot Citation2007). Only low occurrences of oligotrophic taxa such as Pagastiella and Micropsectra insignilobus-type were identified suggesting oxygen levels within the lake were relatively low (Brooks et al. Citation2007). The continued representation of Sergentia coracina-type in combination with farming, contrary to the trend observed in Taylor et al. (Citation2017a), suggests that the large size of Lough Inchiquin may have prevented it from becoming severely eutrophic, although it was clearly more productive in the Late Bronze Age.
Ordinations demonstrate that samples from the Late Bronze Age are associated with enriched δ15N levels on both the RDA and PCA and are not as strongly associated with Ti (RDA Axis 2) as those from the Late Neolithic to Early Bronze Age. δ15N exhibits an influence of almost 10% of the variance within the chironomid data suggesting it is the more substantial of the two forward-selected variables and agrees with past studies which highlighted δ15N as a dominant control in Irish lakes during prehistory (Taylor et al. Citation2013; Citation2017a; Citation2017b). Microtendipes pedellus-type, Cricotopus intersectus-type and Chironomus anthracinus-type are taxa associated with modern pastoral farming in Ireland (Potito et al. Citation2014; Chique et al. Citation2018) and are predominant in Lough Inchiquin at this time. Late Bronze Age farming activity had a much greater impact on the lake system than any preceding period and nitrogen may have been the main driving force affecting the lake system. Indeed, the timing and magnitude of the shift seen in chironomid community composition, following the most substantial increase in NAPp in the record, establishes prehistoric farming as a significant control.
This shift towards intensified pastoral farming in the Bronze Age and a concomitant increase in eutrophic chironomid taxa has been demonstrated in previous studies (Taylor et al. Citation2017a). At all three western Ireland study sites examined by Taylor et al. (Citation2017a) there was a shift in the abundance of eutrophic taxa following the largest peaks of NAPp, with the scale of change roughly proportionate to the scale of farming activity. Although an independent climate variable was not examined in the current study, it is unlikely that it could account for such a significant shift in chironomid data as seen at Lough Inchiquin given the relatively subdued temperature variability in Ireland during the Holocene (see Swindles et al., Citation2013). Furthermore, in more isolated lake catchments (e.g. Lough Meenachrina, northwest Ireland) with little to no evidence of human activity, and where climate was the dominant control, large shifts in chironomid data were not identified during the Holocene (Taylor et al. Citation2018). Human impact can be readily discerned from that of climate in Irish lakes and a moderate level can be sufficient to overwhelm the climate signal (Mckeown and Potito Citation2016).
The trends shown in the geochemical, chironomid and pollen data indicate that pastoral farming in the catchment of Inchiquin increased significantly at c. 1000 BC which may be part of a more widespread trend. Pollen evidence from lowland areas adjacent to the Burren suggest significant tree clearance and hillforts were constructed to the northeast, southeast, and west (Cotter Citation2012; Jones Citation2019; Molloy Citation2005; Molloy, Feeser, and O'connell Citation2014; Molloy and O'connell Citation2012, 206; Plunkett Citation2009). Furthermore, recent study of the δ15N values of archaeologically-derived animal bone show that Ireland as a whole experienced a significant intensification of settlement and farming activity at this time (Guiry et al. Citation2018). Faunal assemblages of cattle, sheep/goat, and pig have been excavated from Neolithic and Chalcolithic sites on Roughan Hill (Jones Citation2004; Jones and Gilmer Citation1999; Ó Maoldúin Citation2015) and it is likely that Late Bronze Age communities within the area relied on these animals in addition to small-scale cereal-cultivation.
Archaeological evidence for Late Bronze Age activity in the catchment is varied. Immediately west of Lough Inchiquin, a leaf-shaped sword and a metalworking hoard were found while the spectacular sheet-gold Gleninsheen gorget was found near the centre of the Burren (Jones Citation2004). Various burnt mounds in the catchment likely date to the Bronze Age, with one example at Fahee South (c. 9 km north of Lough Inchiquin) dated to c. 1413–1213 BC (Brindley and Lanting Citation1990). Habitation is again evidenced at Teeskagh which has been dated to c. 1208–976 and c. 1043–835 BC while comparable dates of c. 1210–931 BC and c. 1022–836 BC were obtained from a very large drystone enclosure at Carran, c. 8.5 km northeast of the Lough Inchiquin (Gibson Citation2007; Citation2016). Several radiocarbon determinations from Glencurran Cave (c. 7 km northeast of Lough Inchiquin) indicate funerary and ritual activity occurred there during the Middle and Late Bonze Age (Dowd Citation2009). Middle and Late Bronze Age activity has also been recently discovered in the far north of the Burren in Moneen Cave (Dowd Citation2016) and at the enigmatic hilltop site of Turlough Hill (Ó Maoldúin and Mccarthy Citation2016; Ó Maoldúin personal communication Citation2018).
It had previously been proposed that the lowland areas between Roughan Hill and Lough Inchiquin may have gained importance in later prehistory because of the earlier effects of soil erosion on the Burren uplands (Grogan Citation2005a). Settlement activity on Roughan Hill itself is thought to have ended c. 1550 BC, and although this may signal a move to lower ground, the continued exploitation of the Burren at sites of different form and location than those of Roughan Hill seems a likely scenario (cf. Jones Citation2019). On the basis of the high representation of poorly-dispersed NAPp taxa and cereals, Spencer et al. (Citation2019) concluded that the Late Bronze Age Landnam was likely the result of continued activity in close proximity to the lake. The current study supports this view with localised and within-lake change in the Late Bronze Age, as demonstrated in the palaeolimnological results, linked to human activity at the lakeside. The area surrounding Lough Inchiquin should thus be considered an important area of Late Bronze Age activity based on the combined palaeoenvironmental and limnological results, which adds to the fuller archaeological picture of the Burren during this period.
Conclusion
Through a multi-proxy approach, this study has assessed the intensity and limnological impacts of farming activity from the Early Neolithic to the Late Bronze Age. The results highlight the increasing ecological impact of anthropogenic activity upon the lake through time. Small-scale Early Neolithic farming had little consequence on the lake itself while intensified Late Bronze Age agriculture resulted in a significant shift in productivity and chironomid community composition favouring eutrophic taxa. This shift, seen in such a large lake, highlights the sensitivity of chironomid taxa to prehistoric farming. The nutrient enrichment of the lake identified via palaeolimnology can be linked to land-use in the surrounding catchment with a strong correspondence between increased NAPp and increased δ15N levels. The level of δ15N was shown through statistical analysis to be a dominant control on the chironomid community composition, through time, followed by titanium. The latter was shown to have the most significant effect on the chironomid data in the Late Neolithic to Early Bronze Age, when the geochemical results suggest significant erosion of Burren soils was occurring. The geochemical data from Lough Inchiquin has provided some of the best-dated evidence for Holocene soil erosion from the Burren which occurred at an early stage in the long history of human-environment interactions in this area. Furthermore, the combined data demonstrate localised and intensive farming in the Late Bronze Age which had a significant impact on the lake system and was likely concentrated in the landscape surrounding Lough Inchiquin.
Acknowledgements
With thanks to the Irish Research Council for funding this research. Help obtained with equipment and coring from Dr Siubhán Comer, Pat O’Rafferty, Dr Carlos Chique, Lynda McCormick, Dr Eugene Costello, Mary Gavaghan and Seamus McGinley is gratefully acknowledged. Thank you to Dr Karen Taylor for assistance with chironomid identification. Support with GIS maps was provided by Joe Fenwick and Dr Benjamin Thébaudeau.
Disclosure Statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
Daisy Eleanor Spencer
Daisy Spencer is an environmental archaeologist and pollen specialist with a research focus on prehistoric Ireland and Britain, specifically Neolithic to Bronze Age human-environment interactions. Her research interests include the use of multi-proxy palaeoenvironmental data, which when linked to the archaeological record can allow for the further development of archaeological narratives. Past research has focused on anthropogenic landscape change via pollen analysis, chironomid analysis, soil micromorphology and DNA studies.
Aaron Potito
Aaron Potito’s research focuses on the use of lake sedimentary records, especially chironomid sub-fossils, to reconstruct past environments. Current research interests include temperature reconstruction of late Pleistocene and Holocene climates, assessment of historic and pre-historic human impacts on lake systems, and impacts of recent climate change on lake ecosystems. Research locations include field sites in Ireland, China and North America.
Karen Molloy
Karen Molloy, a graduate of Trinity College Dublin and University College Galway, specialises in reconstructing landscape change primarily through the use of pollen analysis. Her research, on lake sediments and peat cores, has focussed on long-term climate and landscape change during the Late-glacial period, and the role people have played in shaping the Irish landscape from the Neolithic period through to present time.
Anna Martini
Anna Martini is Professor of Geology and Environmental Studies at Amherst College, Massachusetts, USA. Her work focuses on applying stable isotope geochemistry to understanding processes recorded in sediments and pore waters. Specifically, recent work has examined the microbial and thermogenic effects on the isotopologues of methane and carbon dioxide in coalbed and shale unconventional reservoirs.
Henry Frentzel
Henry Frentzel is an environmental geologist whose past research focused on low-temperature stable isotope geochemistry, including work in rivers, lakes, and a natural gas play. He currently works as a geologist in New York state, where he specialises in contaminant delineation in lake and river sediments.
Carleton Jones
Carleton Jones is a lecturer in prehistoric archaeology at NUI Galway. Dr Jones's research is primarily concerned with investigating the organisation and dynamics of prehistoric societies in Ireland with a focus on the Neolithic, Chalcolithic, and Bronze Age periods. To that end, he has over many years conducted surveys and excavations on the Burren in western Ireland which were supported in part by the Heritage Council and the Royal Irish Academy. In addition to prehistoric social organisation, Dr Jones has also published on megalithic monuments, the archaeology of karst landscapes, and climate change in the past, as well as contributing to studies focused on isotopic evidence, ancient DNA, and human-environment interactions.
References
- Anderson, R. S., R. B. Jass, J. L. Toney, C. D. Allen, L. M. Cisneros-Dozal, M. Hess, J. Heikoop, and J. Fessenden. 2008. “Development of the Mixed Conifer Forest in Northern New Mexico and its Relationship to Holocene Environmental Change.” Quaternary Research 69: 263–275.
- Arnaud, F., S. Révillon, M. Debret, M. Revel, E. Chapron, J. Jacob, C. Giguet-Covex, J. Poulenard, and M. Magny. 2012. “Lake Bourget Regional Erosion Patterns Reconstruction Reveals Holocene NW European Alps Soil Evolution and Paleohydrology.” Quaternary Science Reviews 51: 81–92.
- Baillie, M., and J. Mcaneney. 2015. “Why we Should not Ignore the mid-24th Century BC When Discussing the 2200–2000BC Climate Anomaly.” In 2200 BC—Ein Klimasturz als Ursache für den Zerfall der Alten Welt? 2200 BC—A Climatic Breakdown as a Cause for the Collapse of the Old World?, edited by H. Meller, H. Wolfgang Arz, R. Jung, and R. Risch, 23–26. Halle (Saale): Landesamt fü r Denkmalpflege und Archä ologie Sachsen-Anhalt, Landesmuseum fü r Vorgeschichte.
- Behre, K.-E. 1981. “The Interpretation of Anthropogenic Indicators in Pollen Diagrams.” Pollen et Spores 23: 225–245.
- Bennett, K. D. 1996. “Determination of the Number of Zones in a Biostratigraphical Sequence.” New Phytologist 132: 155–170.
- Bennett, K. D. 2009–1993. psimpoll. Version 4.27. Version 4.27 ed.: Queen's University Belfast, Belfast.
- Birks, H. J. B. 1995. “Quantitative Paleoenvironmental Reconstructions.” Statistical Modelling of Quaternary Science Data. Quaternary Research Association, edited by D. Maddy and J. S. Brew, 161–254. London: Quaternary Research Association.
- Blaauw, M., and J. A. Christen. 2011. “Flexible Paleoclimate age-Depth Models Using an Autoregressive Gamma Process.” Bayesian Analysis 6: 457–474.
- Botrel, M., I. Gregory-Eaves, and R. Maranger. 2014. “Defining Drivers of Nitrogen Stable Isotopes (δ15N) of Surface Sediments in Temperate Lakes.” Journal of Paleolimnology 52: 419–433.
- Brindley, A., and J. Lanting. 1990. “Radiocarbon Dates for Neolithic Single Burials.” The Journal of Irish Archaeology 5: 1–7.
- Brooks, S. J., P. G. Langdon, O. Heiri, Q. R. Association, and G. Britain. 2007. The Identification and use of Palaearctic Chironomidae Larvae in Palaeoecology. London: Quaternary Research Association.
- Chase, J. M., and T. M. Knight. 2006. “Effects of Eutrophication and Snails on Eurasian Watermilfoil (Myriophyllum Spicatum) Invasion.” Biological Invasions 8: 1643.
- Chique, C., A. P. Potito, K. Molloy, and J. Cornett. 2018. “Tracking Recent Human Impacts on a Nutrient Sensitive Irish Lake: Integrating Landscape to Water Linkages.” Hydrobiologia 807: 207–231.
- Cohen, A. S. 2003. Palaeolimnology: The History and Evolution of Lake Systems. Oxford: Oxford University Press.
- Cotter, C. 2012. The Western Stone Forts Project Exacvations at Dun Aonghasa and Dun Eoganachta Volume 1 & 2, Dublin, Wordwell Ltd (The Discovery Programme).
- Cullinane, S. 2012. An Assessment of Algal Biodiversity and Water Quality in Loughs Atedaun, Cullaun and Inchiquin, Three Lakes on the River Fergus. Limerick: Mary Immaculate College, University of Limerick.
- Diefendorf, A. F., W. P. Patterson, C. Holmden, and H. T. Mullins. 2008. “Carbon Isotopes of Marl and Lake Sediment Organic Matter Reflect Terrestrial Landscape Change During the Late Glacial and Early Holocene (16,800 to 5,540 cal yr BP): A Multiproxy Study of Lacustrine Sediments at Lough Inchiquin, Western Ireland.” Journal of Paleolimnology 39: 101–115.
- Diefendorf, A. F., W. P. Patterson, H. T. Mullins, N. Tibert, and A. Martini. 2006. “Evidence for High-Frequency Late Glacial to mid-Holocene (16,800 to 5500 cal yr B.P.) Climate Variability from Oxygen Isotope Values of Lough Inchiquin, Ireland.” Quaternary Research 65: 78–86.
- Dowd, M. 2009. “Middle and Late Bronze Age Ritual Activity at Glencurran Cave, Co. Clare.” In From Bann Flakes to Bushmills: Papers in Honour of Professor Peter Woodman, edited by N. Finlay, S. Mccartan, N. Milner, and C. Wickham-Jones, 89–100. Oxford: Oxbow Books.
- Dowd, M. 2016. Archaeological Excavations in Moneen Cave, The Burren, Co. Clare. Insights Into Bronze Age and Post-Medieval Life in the West of Ireland. Oxford: Archaeopress.
- Drew, D. 1983. “Accelerated Soil Erosion in a Karst Area: The Burren, Western Ireland.” Journal of Hydrology 61: 113–124.
- Drew, D. 1988. “The Hydrology of the Upper Fergus River Catchment, Co. Clare.” Proceedings of the University Bristol Spelaeological Society 18: 265–277.
- Dubois, N., É Saulnier-Talbot, K. Mills, P. Gell, R. Battarbee, H. Bennion, S. Chawchai, et al. 2018. “First Human Impacts and Responses of Aquatic Systems: A Review of Palaeolimnological Records from Around the World.” The Anthropocene Review 5 (1): 28–68.
- Engstrom, D. R., and H. Wright JR. 1984. “Chemical Stratigraphy of Lake Sediments as a Record of Environmental Change.” Lake Sediments and Environmental History: Studies in Palaeolimnology and Palaeoecology in Honour of Winifred Tutin/Edited by EY Haworth and JWG Lund 1: 11–67.
- Feeser, I., and M. O’connell. 2009. “Fresh Insights Into Long-Term Changes in Flora, Vegetation, Land use and Soil Erosion in the Karstic Environment of the Burren, Western Ireland.” Journal of Ecology 97: 1083–1100.
- Finch, T., E. Culleton, and S. Diamond. 1971. Soils of County Clare. Dublin, Ireland: An Foras Talúntais Dublin.
- Fitzpatrick, A. P. 2015. “Great Britain and Ireland in 2200 BC.” In 2200 BC—Ein Klimasturz als Ursache für den Zerfall der Alten Welt? 2200 BC—A Climatic Breakdown as a Cause for the Collapse of the Old World?, edited by H. Meller, H. Wolfgang Arz, R. Jung, and R. Risch, 805–832. Halle (Saale): Landesamt fü r Denkmalpflege und Archä ologie Sachsen-Anhalt, Landesmuseum fü r Vorgeschichte.
- Francis, D. R., and D. R. Foster. 2001. “Response of Small New England Ponds to Historic Land use.” The Holocene 11: 301–312.
- Frentzel, H. 2017. Marl Support: Climate and Landscape Changes in the Burren, Western Ireland, as Reflected in Lough Inchiquin lake sediment. Unpublished Undergraduate Thesis, Amherst College.
- Gibson, D. B. 2007. The Hill-slope Enclosures of the Burren, Co. Clare. Proceedings of the Royal Irish Academy. Section C: Archaeology, Celtic Studies, History, Linguistics, Literature, 1–29.
- Gibson, D. B. 2016. “Chalcolithic Beginnings and Ecological Change Spanning 1,000 Years as Glimpsed from a Doline in the Burren, Co. Clare.” Proceedings of the Royal Irish Academy. Section C: Archaeology, Celtic Studies, History, Linguistics, Literature 116: 33–91.
- Grogan, E., ed. 2005a. The North Munster Project Vol. 2: The Prehistoric Landscape of North Munster. Bray: Wordwell.
- Grossman, E. L. 1994. “. The Carbon and Oxygen Isotope Record During the Evolution of Pangea: Carboniferous to Triassic.” Geological Society of America Special Papers 288: 207–228. doi:10.1130/SPE288-p207.
- Guiry, E., F. Beglane, P. Szpak, R. Schulting, F. Mccormick, and M. P. Richards. 2018. “Anthropogenic Changes to the Holocene Nitrogen Cycle in Ireland.” Science Advances 4: eaas9383.
- Harris, D., W. R. Horwáth, and C. Van Kessel. 2001. “Acid Fumigation of Soils to Remove Carbonates Prior to Total Organic Carbon or CARBON-13 Isotopic Analysis.” Soil Science Society of America Journal 65: 1853–1856.
- Heiri, O., and A. F. Lotter. 2003. “9000 Years of Chironomid Assemblage Dynamics in an Alpine Lake: Long-Term Trends, Sensitivity to Disturbance, and Resilience of the Fauna.” Journal of Paleolimnology 30: 273–289.
- Heiri, O., A. F. Lotter, and G. Lemcke. 2001. “Loss on Ignition as a Method for Estimating Organic and Carbonate Content in Sediments: Reproducibility and Comparability of Results.” Journal of Paleolimnology 25: 101–110.
- Holmer, M., and P. Storkholm. 2001. “Sulphate Reduction and Sulphur Cycling in Lake Sediments: A Review.” Freshwater Biology 46: 431–451.
- Holmes, J. A., M. Leuenberger, K. Molloy, and M. O’connell. 2019. “Younger Dryas and Holocene Environmental Change at the Atlantic Fringe of Europe Derived from Lake-Sediment Stable-Isotope Records from Western Ireland.” BOREAS 49 (2): 233–247.
- Jones, C. 2004. The Burren and the Aran Islands: Exploring the Archaeology. Cork: Collins.
- Jones, C. 2016. “Dating Ancient Field Walls in Karst Landscapes Using Differential Bedrock Lowering.” Geoarchaeology 77–100.
- Jones, C. 2019. “Climate Change and Farming Response in a Temperate Oceanic Zone - the Exploitation of a Karstic Region in Western Ireland in the Third and Second Millennia BC.” Journal of Island and Coastal Archaeology. doi:10.1080/15564894.2019.1614115.
- Jones, C., and A. Gilmer. 1999. “Roughan Hill, a Final Neolithic/Early Bronze Age Landscape Revealed.” Archaeology Ireland 13: 30–32.
- Juggins, S. 2003. Program C2 Data Analysis. Newcastle, UK: University of Newcastle.
- Luther, G. W., B. Glazer, M. A. S, R. Trouwborst, B. R. Shultz, G. Druschel, and C. Kraiya. 2003. “Iron and Sulfur Chemistry in a Stratified Lake: Evidence for Iron-Rich Sulfide Complexes.” Aquatic Geochemistry 9: 87–110.
- Lynch, A. 2014. Poulnabrone: An Early Neolithic Portal Tomb in Ireland. Dublin: The Stationary Office.
- Mackereth, F. J. H. 1966. “Some Chemical Observations on Post-Glacial Lake Sediments.” Phil. Trans. R. Soc. Lond. B 250: 165–213.
- Mckeown, M., and A. P. Potito. 2016. “Assessing Recent Climatic and Human Influences on Chironomid Communities from two Moderately Impacted Lakes in Western Ireland.” Hydrobiologia 765: 245–263.
- Moles, N., and R. Moles. 2002. “Influence of Geology, Glacial Processes and Land use on Soil Composition and Quaternary Landscape Evolution in The Burren National Park, Ireland.” Catena 47: 291–321.
- Moles, R., N. Moles, and J. J. Leahy. 1999. “Radiocarbon Dated Episode of Bronze Age Slope Instability in the South-Eastern Burren, County Clare.” Irish Geography 32: 52–57.
- Moller Pillot, H. K. M. 2009. Chironomidae Larvae of the Netherlands and Adjacent Lowlands: Biology and Ecology of the Chironomini. Zeist, Netherlands: KNNV Publishing.
- Moller Pillot, H. 2013. Chironomidae Larvae, Volume 3: Biology and Ecology of the Aquatic Orthocladiinae. Zeist: KNNV Publishing.
- Molloy, K. 2005. “Holocene Vegetation and Land-use History at Mooghaun, SE Clare, with Particular Reference to the Bronze Age.” In The North Munster Project, Vol. 1 The Later Prehistoric Landscape of South-East Clare. Discovery Programme Monographs, edited by E. Grogan, 255–287. Dublin, Bray: Wordwell.
- Molloy, K., I. Feeser, and M. O'connell. 2014. Farming and Woodland Dynamics: A New Pollen Diagram from Ballinphuill Bog.
- Molloy, K., and M. O'connell. 2012. “Prehistoric Farming in Western Ireland: Pollen Analysis at Caheraphuca, Co. Clare.” In Borderlands. Archaeological Investigations on the Route of the M18 Gort to Crusheen Road Scheme, NRA Scheme Monographs, edited by Delaney, 109–122. Dublin: National Roads Authority.
- Naeher, S., A. Gilli, R. P. North, Y. Hamann, and C. J. Schubert. 2013. “Tracing Bottom Water Oxygenation with Sedimentary Mn/Fe Ratios in Lake Zurich, Switzerland.” Chemical Geology 352: 125–133.
- O'brien, C., K. Selby, Z. Ruiz, A. Brown, M. Dinnin, C. Caseldine, P. Langdon, and I. Stuijts. 2005. “A Sediment-Based Multiproxy Palaeoecological Approach to the Environmental Archaeology of Lake Dwellings (Crannogs), Central Ireland.” The Holocene 15: 707–719.
- Ó Maoldúin, R. 2015. Preliminary Report on the Excavation of Wedge Tomb CL017–180002, Parknabinnia, Co. Clare. Unpublished.
- Ó Maoldúin, R. 2018. RE: Turlough Hill dating. Personal communication with SPENCER, D.
- Ó Maoldúin, R., and N. Mccarthy. 2016. Preliminary Report on excavation 16E0130 Turlough Hill, Co. Clare. Unpublished.
- Perren, B. B., C. Massa, V. Bichet, É Gauthier, O. Mathieu, C. Petit, and H. Richard. 2012. “A Paleoecological Perspective on 1450 Years of Human Impacts from a Lake in Southern Greenland.” The Holocene 22: 1025–1034.
- Plunkett, G. 2009. “Land-use Patterns and Cultural Change in the Middle to Late Bronze Age in Ireland: Inferences from Pollen Records.” Vegetation History and Archaeobotany 18: 273–295.
- Potito, A. P., C. A. Woodward, M. Mckeown, and D. W. Beilman. 2014. “Modern Influences on Chironomid Distribution in Western Ireland: Potential for Palaeoenvironmental Reconstruction.” Journal of Paleolimnology 52 (4): 1–20.
- Quinlan, R., and J. P. Smol. 2001. “Setting Minimum Head Capsule Abundance and Taxa Deletion Criteria in Chironomid-Based Inference Models.” Journal of Paleolimnology 26: 327–342.
- Reimer, P. J., E. Bard, A. Bayliss, J. W. Beck, P. G. Blackwell, C. B. Ramsey, C. E. Buck, H. Cheng, R. L. Edwards, and M. Friedrich. 2013. “IntCal13 and Marine13 Radiocarbon age Calibration Curves 0–50,000 Years cal BP.” Radiocarbon 55: 1869–1887.
- Rieradevall, M., and S. J. Brooks. 2001. “An Identification Guide to Subfossil Tanypodinae Larvae (Insecta: Diptera: Chrironomidae) Based on Cephalic Setation.” Journal of Paleolimnology 25: 81–99.
- Roland, T. P., C. Caseldine, D. Charman, C. Turney, and M. Amesbury. 2014. “Was There a ‘4.2 ka Event’in Great Britain and Ireland? Evidence from the Peatland Record.” Quaternary Science Reviews 83: 11–27.
- Ruiz, Z., A. G. Brown, and P. G. Langdon. 2006. “The Potential of Chironomid (Insecta: Diptera) Larvae in Archaeological Investigations of Floodplain and Lake Settlements.” Journal of Archaeological Science 33: 14–33.
- Schulting, R. J., E. Murphy, C. Jones, and G. Warren. 2012. New Dates from the North and a Proposed Chronology for Irish Court Tombs. Proceedings of the Royal Irish Academy, Section C. The Royal Irish Academy, 1–60.
- Snoeck, C., C. Jones, S. Goderis, P. Claeys, N. Mattielli, A. Zazzo, M. Lebon, P. J. Reimer, J. Lee-Thorp, and R. Schulting. 2020. “Isotopic Evidence for a Change in Mobility and Landscape use Patterns Between the Neolithic and Early Bronze Age in Western Ireland.” Journal of Archaeological Science: Reports, 1–11. doi:10.1016/j.jasrep.2020.102214.
- Spencer, D. E., K. Molloy, A. Potito, and C. Jones. 2019. New Insights Into Late Bronze Age Settlement and Farming Activity in the Southern Burren, Western Ireland. Vegetation History and Archaeobotany. doi:10.1007/s00334-019-00746-1.
- Swindles, G. T., I. T. Lawson, I. P. Matthews, M. Blaauw, T. J. Daley, D. J. Charman, T. P. Roland, G. Plunkett, G. Schettler, and B. R. Gearey. 2013. “Centennial-Scale Climate Change in Ireland During the Holocene.” Earth-Science Reviews 126: 300–320.
- Talbot, M. R. 2001. “Nitrogen Isotopes in Palaeolimnology.” In Tracking Environmental Change Using Lake Sediments Volume 2: Geochemical Methods, edited by W. M. Last and J. P. Smol, 401–439. The Netherlands: Kluwer Academic Publishers.
- Taylor, K., S. Mcginley, A. P. Potito, and K. Molloy. 2018. “A mid to Late Holocene Chironomid-Inferred Temperature Record from Northwest Ireland.” Palaeogeography, Palaeoclimatology, Palaeoecology 505: 274–286.
- Taylor, K. J., A. P. Potito, D. W. Beilman, B. Ghilardi, and M. O'connell. 2013. “Palaeolimnological Impacts of Early Prehistoric Farming at Lough Dargan, County Sligo, Ireland.” Journal of Archaeological Science 40: 3212–3221.
- Taylor, K. J., A. P. Potito, D. W. Beilman, B. Ghilardi, and M. O’connell. 2017a. “Impact of Early Prehistoric Farming on Chironomid Communities in Northwest Ireland.” Journal of Paleolimnology 57: 227–244.
- Taylor, K. J., S. Stolze, D. W. Beilman, and A. P. Potito. 2017b. “Response of Chironomids to Neolithic Land-use Change in North-West Ireland.” The Holocene 6: 0959683616675935.
- Ter Braak, C., and P. Šmilauer. 1997. Canoco for Windows (version 4.54): Software for Canonical Community Ordination. Copyright.
- Thompson, A. D. 1997. A Lake Sediment Record of Late Glacial and Holocene Environmental Change from Molly's Lough, Co. Clare, Ireland. PhD, University of Wales.
- Vallenduuk, H. J., and H. K. M. Pillot. 2007. Chironomidae Larvae, Vol. 1: Tanypodinae.
- Vander Zanden, M. J., Y. Vadeboncoeur, M. W. Diebel, and E. Jeppesen. 2005. “Primary Consumer Stable Nitrogen Isotopes as Indicators of Nutrient Source.” Environmental Science & Technology 39: 7509–7515.
- Waemere, A. 2005. Influence of Catchment Characteristics on the Relationship between Land Use and Lake Water Quality in County Clare. PhD, Trinity College Dublin, Dublin.
- Walker, I. R. 2001. “Midges: Chironomidae and Related Diptera.” In Tracking Environmental Change Using Lake Sediments, edited by J. P. Smol, H. J. B. Birks, and W. M. Last, 43–66. The Netherlands: Kluwer Academic Publishers.
- Wiederholm, T. 1983. “Chironomidae of the Holarctic Region. Keys and Diagnoses. Part 1-Larva.” Entomologica Scandinavica Supplement 19: 1–457.
- Woodward, C. A., A. P. Potito, and D. W. Beilman. 2012. “Carbon and Nitrogen Stable Isotope Ratios in Surface Sediments from Lakes of Western Ireland: Implications for Inferring Past Lake Productivity and Nitrogen Loading.” Journal of Paleolimnology 47: 167–184.