
ABSTRACT
This introduction aims to set out the potential as well as some of the pitfalls of the newly emerging area of the Social Neuroscience of Human Attachment (SoNeAt). To organize and interconnect the burgeoning empirical studies in this line of research, including those in this special issue, we outline a programmatic framework including an extension of our conceptual proposals NAMA and NAMDA to guide future research. We hope that this special issue will act as a stimulus for redoubling our efforts advancing the newly emerging SoNeAt area bridging attachment theory and social neuroscience.
1. General considerations
1.1 Moving from confirmation to integration and conceptual innovation
Ever since the pioneering work on attachment and neurobiology in the early 1990s, scholars have asserted the need for integration of both fields (Coan, Citation2016; Gustison & Phelps, Citation2022; Long et al., Citation2020; Spangler & Grossman, Citation1999; Vrticka & Vuilleumier, Citation2012; Vrtička, Citation2017; White et al., Citation2020). In its early stages, the main objective was to offer a source of objective confirmation for some of attachment theory’s core hypotheses that were intractable using a purely observational-behavioral armamentarium. As a seminal example of this work, Spangler and Grossmann (Citation1993) demonstrated elevations in stress-related physiology (i.e. heart rate and cortisol secretion) despite minimal outward expression of distress among insecure-avoidant infants during the Strange Situation Procedure (SSP). Physiological measures thus served as an elegant route to objectively visualize psychophysiological processes that were otherwise difficult to access, given their “hidden” (i.e. potentially automatic or even non-conscious) nature.
While this “confirmatory” line of work still remains relevant in present-day attachment research, this special issue testifies to the fact that we are now rapidly moving towards a new stage of integration that goes well beyond a purely confirmatory role of neurobiological methods for attachment-related theory. A crucial part of this integration involves newly emerging theoretical cross-fertilization whereby the fields of attachment and neurobiology, and particularly its subfield of social neuroscience (see J. T. Cacioppo & Berntson, Citation1992; J. T. Cacioppo et al., Citation2010; S. Cacioppo & Cacioppo, Citation2020), meet each other on a level footing to promote conceptual innovation – an approach nowadays also known as the social neuroscience of human attachment (SoNeAt). Accordingly, work in the area of SoNeAt not only poses the question of how neurobiology can (dis-)confirm propositions in, but also how it can innovate and bring novel insights to attachment theory.
In what follows, we will spell out some of the key issues to consider for the fields of attachment and social neuroscience before outlining a theoretical perspective that aims to organize emerging empirical work in this burgeoning field using the papers included in this special issue as examples. We will conclude by briefly outlining the key research gaps that we hope will inspire future research agenda for SoNeAt.
1.2 Two heuristics for theoretical cross-fertilization
We believe a new level of integration between attachment theory and social neuroscience is both timely and imperative. Both fields have significantly matured during the last decade or two, procuring ever more robust evidence deriving from conceptual reviews, meta-analyses and rigorous large-scale studies. These insights now afford a novel way to bridge the two disciplines, each with its own firm set of methods and knowledge bases.
We propose two inter-related heuristics for such theoretical cross-fertilization. The first heuristic takes an attachment theory perspective and interrogates what social neuroscience studies may add to our quest for examining many of the core tenets and open issues in attachment theory – as outlined in Thompson et al. (Citation2021) and later summarized in Thompson et al. (Citation2022). One helpful point of departure may be the key hypotheses of attachment theory (IJzendoorn, Citation1990; Mesman et al., Citation2016; Rothbaum et al., Citation2000). Especially with regard to the so-called sensitivity hypothesis (i.e. concerning the child-rearing antecedents of differing attachment classifications) and competence hypothesis (i.e. concerning the interpersonal consequences of differing attachment classifications), SoNeAt research has the potential to specify the biological and brain mechanisms. This particularly pertains to the “how” questions in Tinbergen’s classical terminology (see Gustison & Phelps, Citation2022) regarding the transmission, formation, and maintenance of attachment patterns at the within- and between-person levels (i.e. across time and social contexts). Indeed, SoNeAt with its associated theoretical frameworks has already begun and will undoubtedly continue to help decompose and specify the core ingredients of many attachment-related processes (e.g. internal working models; IWMs), by mapping aspects of their proposed functioning onto specific psychophysiological components or brain modules (see Section 2 below). Such an approach will thus allow us to provide novel neurobiologically grounded insights into many of the key open and contentious questions within the field regarding, among others, definition, measurement, continuity, stability and change. As well, clinical translation and interventions may be strengthened by including psychophysiological measures in the toolbox of measurements of change processes. We believe that SoNeAt holds the promise to do so by offering a common attachment-relevant neurobiological “language” (see also Duschinsky et al., Citation2021; Verhage et al., Citationin press) that allows for more direct comparisons between areas formerly thought of as separate or distinct (see ; e.g. overlap and non-overlap between brain regions involved in formation vs. generalization vs. transmission of attachment patterns).
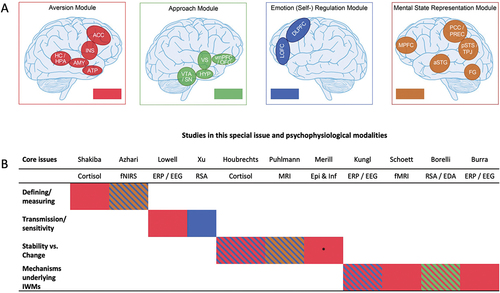
Figure 1. Panel A (top) - depiction of the four neural modules as part of the functional neuro-anatomical model of human attachment (NAMA; adapted from Long et al., Citation2020). Panel B (bottom) - table indicating the core issues in attachment theory and research (rows) and NAMA modules (color coded in accordance with Panel A) to which papers in this special issue (columns listing the respective first author) are making a contribution. *= as reflected in inflammatory processes. fNIRS= functional near-infrared spectroscopy, ERP= event-related potentials, EEG= electroencephalography, RSA= respiratory sinus arrhythmia, MRI= magnetic resonance imaging, Epi= epigenetics, Inf= inflammation, fMRI= functional MRI, EDA= electrodermal activity, IWMs= internal working models of attachment.
However, the aforementioned step can only be achieved if we pay heed to the second heuristic, which involves a sound understanding of the prospects and limits of social neuroscience (J. T. Cacioppo et al., Citation2016). In particular, scholars need to keep abreast of accumulating evidence yielding new developments in theory (see Section 2 below), methods (e.g. transition from first-person to second-person social neuroscience paradigms – see Redcay & Schilbach, Citation2019; Schilbach et al., Citation2013), and analytic strategies (e.g. data quality checks, statistical procedures and thresholds, correction for multiple comparisons, etc., see e.g. Eklund et al., Citation2016; Nguyen, Hoehl, et al., Citation2021), and remain sensitive to the complexities of possible relationships between physiological and psychological processes. Notably, physiological processes are typically somewhat non-specific and thus often merely qualify as so-called concomitants (or correlates) indicating that a one-to-many relationship exists between the underlying physiological index and potential psychological processes. For example, the same physiological measure, such as respiratory sinus arrhythmia (RSA), may reflect emotion regulation or prosocial/empathic tendencies depending on the circumstances in which it is assessed. Inevitably, this forces researchers to consider multiple interpretations of social neuroscience data and justify their preferred interpretation, depending on, among others, the presence/absence of rigorous control conditions and previous work on the chosen physiological modality (J. T. Cacioppo et al., Citation2016). A sound awareness of these issues (that goes beyond mere lip-service) is crucial to ensure high-quality research and reporting of studies as SoNeAt matures.
2. New evidence as part of NAM(D)A
2.1 Functional neuro-anatomical models of human attachment
To organize and make sense of the social neuroscience evidence on attachment available to date, we have recently proposed an extended and refined functional neuro-anatomical model of human attachment (NAMA; see Panel A and Long et al., Citation2020) and its disruption/disorganization (NAMDA; White et al., Citation2020). To structure the extant data in accordance with attachment theory, NAM(D)A draws upon the account of an attachment behavioral system (Mikulincer et al., Citation2003) describing the emotions, cognitive operations, response strategies and dynamics, as well as goals and their psychological manifestations and consequences of different attachment behaviors. NAM(D)A builds upon a prototypical attachment pathway reflecting the most likely sequence of behaviors during attachment-related interactions, and assigns the latter to four distinct, but interconnected affective (i.e. aversion and approach) and cognitive (i.e. emotion regulation and mental state representation) neural modules/networks. NAM(D)A subsequently summarizes the available social neuroscience evidence pertaining to how interindividual differences in organized (i.e. secure, insecure-avoidant and insecure-ambivalentFootnote1) and disorganized attachment modulate the prototypical attachment pathway and corresponding activity and structure within, as well as connectivity between the affective and cognitive evaluation modules. This evidence is extended by also including findings pertaining to physiology (e.g. heart rate, hormone secretion, etc.) and recently emerging data from the fields of genetics and epigenetics. Finally, and particularly in the case of NAMA, new implications of second-person social neuroscience approaches, and especially bio-behavioral and interpersonal neural synchronization are also considered.
2.2 Two key insights of SoNeAt
Before summarizing the new evidence brought forward by the individual reports as part of this special issue, we would like to mention two key insights social neuroscience has already provided to attachment theory (for more details, see Long et al., Citation2020; White et al., Citation2020).
First, there is no dedicated neurobiological attachment system – analogous to a proposed behavioral attachment system (Mikulincer et al., Citation2003) - solely concerned with orchestrating attachment behavior. The four neural modules highlighted in NAM(D)A are part of extended brain networks that are involved in many computations underlying a vast range of behaviors. In other words, there is no dedicated neural “attachment area,” nor is there a dedicated neural “attachment network.” For example, the aversion module as part of NAM(D)A implicated in detecting attachment-related threats and mounting an appropriate fear response linked with a deviation from homeostasis is broadly analogous to a salience neural network for perceiving and responding to domain-general homeostatic demands (Seeley, Citation2019). Likewise, NAM(D)A’s mental state representation module associated with the neurocognitive maintenance of certain aspects of IWMs of attachment is also known more generally as the default mode network (DMN; e.g. Alves et al., Citation2019). The brain basis of human attachment thus consists of extended neural networks subserving many different social, cognitive and emotional computations that are also recruited in an attachment context. Consequently, SoNeAt’s main purpose is to better understand how interindividual differences in attachment are linked to the neurobiological functioning of these extended brain networks in relation to their many other vital functions that are the focus of other social neuroscience domains.
Second, and implicit in the above, we believe that an overarching neurobiological account of human attachment begins to emerge through the combination of SoNeAt’s insights with recent other social neuroscience research and theory. This account links attachment with fundamental neurobiological processes related to energy conservation and co-regulation through social allostasis, mainly drawing upon social baseline theory and an evolutionary theory of social regulation (Atzil et al., Citation2018; Coan & Sbarra, Citation2015). Accordingly, when individuals face a significant environmental challenge (e.g. threat, illness, etc.), an innate interpersonal survival strategy is triggered (see Panel A). This strategy involves checking for the availability of approachable others, and, contingent on their accessibility, reaching out for their help. The underlying mechanism relies on the principle that personal and social resources are neurobiologically treated as interchangeable. In other words, the more individuals can draw upon others’ resources through co-regulation, the less personal bioenergetic resources become depleted and the more they can be conserved (as if they were literally increased from the get-go). Attachment theory nicely captures this intrinsic link between energy conservation and social allostasis (see Panel B). It postulates that individuals engage in proximity seeking under distress as part of their primary attachment strategy (often associated with organized secure attachment), because being close to others, especially significant others, allows them to draw upon others’ resources for co-regulation.
Figure 2. Illustration of the link between fundamental neurobiological processes related to energy conservation and co-regulation through social allostasis from a social neuroscience (Panel A – top) and attachment (Panel B – bottom) perspective. Line style indicates more likely (solid) versus possible alternative (dashed) pathways. IWMs= internal working models of attachment, w/o= without.
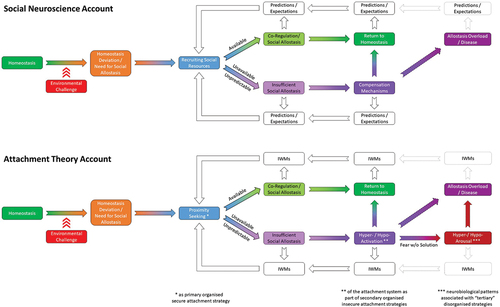
The same account also helps understand what happens in case of a (real or perceived) lack of accessible social resources under environmental challenge. In such circumstances, individuals need to employ compensation mechanisms or, in attachment terms, secondary (organized insecure) attachment strategies. One compensation mechanism involves decreasing proximity seeking and increasing self-regulation to face future challenges independently. This self-focused secondary attachment strategy reflecting an “individual fight” response and attachment system hypo-activation is associated with insecure-avoidant attachment. Another compensation mechanism relies upon intensifying proximity seeking behavior to re-establish social connection. This other-focused secondary attachment strategy reflecting a “social flight” response and attachment system hyper-activation is linked to insecure-ambivalent attachment.
The aforementioned two compensation mechanisms associated with secondary attachment strategies and corresponding neural correlates (see NAMA; Long et al., Citation2020) are initially adaptive because they arise as appropriate responses to specific environmental demands. Moreover, they can typically reduce allostatic load sufficiently to prevent allostasis overload and related pathology. However, if these compensation mechanisms need to be employed over an extended amount of time and/or with increasing allostatic loadFootnote2, they begin to represent risk factors for mental and physical health associated with allostasis overload and attachment disruption/disorganization with its corresponding neural correlates (see NAMDA; White et al., Citation2020). Similar considerations have been put forward by other social neuroscience theories, for example, in association with an evolutionary mechanism for/theory of loneliness (J. T. Cacioppo & Cacioppo, Citation2018; J. T. Cacioppo et al., Citation2014), the link between brain-body interactions, loneliness, and mental and physical health (Quadt et al., Citation2020), a theory of social thermoregulation (IJzerman et al., Citation2015), a new model of social allostatic load (Saxbe et al., Citation2020) and considerations of the neural mechanisms of social homeostasis more generally (Matthews & Tye, Citation2019). Finally, this account also informs our current view of attachment disruption/disorganization where, as a consequence of fear without solution, compensation mechanisms appear to become more rigid and extreme (i.e. linked to neurobiological hyper- and hypo-arousal patterns as “tertiary”, disorganized strategies), or even break down entirely (e.g. as in dissociation).
In sum, we are convinced that there are ample links between SoNeAt and other social neuroscience theories, and that further extending and refining this common ground will greatly benefit advances not only within SoNeAt, but attachment theory and research as a whole.
3. This special issue
SoNeAt has already revealed many crucial insights into the neurobiology of human attachment (see Section 2 above). In this special issue, we would like to highlight some of the most recent methodological developments and insights into SoNeAt based on research from expert teams around the globe. Eleven original empirical studies addressed associations between individual differences in attachment and (i) brain function, structure and connectivity, (ii) interpersonal neural synchronization/coupling, (iii) psychophysiology and (iv) epigenetics. To summarize the main implications of these studies, we would like to briefly illustrate how the new methodology used, and data obtained, can help answer some of the core open questions of attachment theory (see also Panel B above; informed by Thompson et al., Citation2021).
3.1 Definition and assessment of attachment (security)
All articles of this special issue provide crucial insights into the questions of how attachment (security) can be defined and assessed, and whether the different assessment methods yield commensurate neurobiological correlates – thus addressing Questions 1 and 2 in Thompson et al. (Citation2021). A key aim of SoNeAt is to create a common attachment-relevant neurobiological “language” based on converging social neuroscience data patterns. SoNeAt therefore is well-suited to build generalizable conclusions about attachment based on studies with measurement approaches allowing for cross-age and cross-sample comparisons (see Thompson et al., Citation2021, p. 546). In this special issue, and representative of SoNeAt overall, interindividual differences in attachment were assessed with many different means across various samples – in children using the SSP, a Story Stem completion task, the Child Attachment Interview (CAI) and different self-reports; in adolescents using self-reports; and in adults using the Adult Attachment Interview (AAI), Attachment Script Assessment and various self-reports. Nonetheless, the combined data allows for generalizable conclusions about the neurobiological basis of attachment security, insecurity and disorganization. This is due to the fact that outcomes of these different attachment measures were consistently related to objective social neuroscience data. We therefore firmly believe that such an objective SoNeAt approach has strong potential to clarify and unify attachment theory in the upcoming years. Crucially, to achieve this goal, the different attachment assessments employed do not need to measure the same thing. Generalizability is achieved on another level, namely through the link with objectively quantifiable neurobiological data – that can subsequently be compared to data from other social neuroscience domains for even more generalizability and clarity.
Question 1 in Thompson et al. (Citation2021) also inquires which characteristics of close, affectionate relationships should be understood to constitute, or defined as “attachment” relationships. This consideration is also relevant for Question 9 in Thompson et al. (Citation2021) on the relevance of attachment theory and research to systems and services for children and families, including institutional and foster care. One way in which SoNeAt can help address these two questions is related to the link between energy conservation and social allostasis that we think represents a core feature of attachment relationships (see Section 2.2 above). Accordingly, as part of this special issue, Shakiba and Raby (Citation2021) examined salivary cortisol during the SSP in children aged 11–33 months adopted internationally from institutional or foster care. The authors found that forming high-quality (i.e. secure) attachment relationships characterized by proximity seeking and contact with adoptive parents served as a protective factor that facilitated recovery following a stressful situation in these young vulnerable children. Such objective SoNeAt data related to stress co-regulation could thus constitute a central correlate involved in the early formation of secure(−like) attachment relationships with various significant others – which is in line with NAMA that links decreased aversion module activation during distress with the (real or imagined) presence of significant others serving a vital co-regulatory function (Long et al., Citation2020).
A related development within social neuroscience that carries potential for characterizing key variables of attachment relationships and its measurement – and thus speaking to both Questions 1 and 2 in Thompson et al. (Citation2021) - involves second-person social neuroscience data in terms of bio-behavioural and particularly interpersonal neural synchronization (INS). This issues from the fact that co-regulation through allostasis is thought to be tightly linked with patterns of neurophysiological de- and re-synchronisation (Atzil et al., Citation2018; Feldman, Citation2017). SoNeAt investigations have thus recently begun to examine associations between attachment and INS with a special focus on parent-child interaction, including the study by Azhari et al. (Citation2020) as part of this special issue. In their study, the authors asked mother-child dyads (children aged 3–4 years) to passively watch short animation videos together while brain activity was obtained from both parties using functional near-infrared spectroscopy (fNIRS) hyperscanning. Concordance in mothers’ and children’s brain activity in terms of interpersonal neural coupling – similar to INS but more strongly externally than internally driven (see e.g. Hamilton, Citation2021; Nummenmaa et al., Citation2018) - was then related to mothers’ self-reported (Attachment Style Questionnaire; ASQ) interindividual differences in attachment. Results revealed preliminary evidence that interpersonal neural coupling may be diminished for dyads with mothers scoring higher on insecure-ambivalent attachment. These results add to the small, but steadily growing evidence base that INS may constitute a new neurobiologi cal correlate for interaction and relationship quality, particularly so from an attachment perspective (Miller et al., Citation2019; Nguyen et al., Citation2020; Nguyen, Schleihauf, Kayhan, et al., Citation2021, Nguyen, Schleihauf, Kungl, et al., Citation2021). In the future, this line of research may carry important implications for mechanisms underlying intergenerational transmission and the sensitivity hypothesis as well as helping to address the transmission gap (see also Sections 3.2 and 4 below).
3.2 Transmission and sensitivity
Another of the nine fundamental questions of attachment theory discussed by Thompson et al. (Citation2021), more specifically Question 5, concerns the influence of early attachment-related interactions on child socio-emotional outcomes. As part of their discussion, the authors also mention the role of caregiving (partly encapsulated by the sensitivity hypothesis; see Mesman et al., Citation2016; Rothbaum et al., Citation2000) as an important contributing factor. From a somewhat broader perspective, such considerations tap into the question of intergenerational attachment transmission and possible ways to bridge the widening transmission gap (see van IJzendoorn & Bakermans-Kranenburg, Citation2019; Verhage et al., Citation2016). One way in which SoNeAt can help tackle these issues is by revealing the association between parents’ own attachment histories and their brain responses to infant cues as a proxy of caregiving neurobiology. Within this special issue, two studies employed such an experimental strategy. Lowell et al. (Citation2021) examined mothers’ brain responses with electroencephalography (EEG), focusing on event-related potential (ERP) components reflecting early orientation and structural encoding (N100 and N170) and attentional allocation (P300) to infant cues (i.e. emotional facial expressions and vocalizations) as a function of mothers’ AAI classifications. They found latency effects for all three ERP components in that secure mothers were significantly faster to process infant cues, which may facilitate more appropriate (and potentially more sensitive) caregiving responses. In addition, using RSA as an index of parasympathetic nervous system activation, Xu and Groh (Citation2021) assessed the significance of mothers’ attachment in terms of secure base script knowledge (SBSK) for autonomic physiological regulation when interacting with infants in an interpersonally challenging context. Their data revealed that mothers scoring higher on SBSK showed greater change in RSA during the procedure, which may reflect better physiological regulation according to the demands of the caregiving context. The above SoNeAt findings are in line with the notion that the attachment and caregiving behavioral systems are complementary and strongly interconnected (Canterberry & Gillath, Citation2012), with such interconnection increasingly observable by means of objective neurobiological data. What needs to be further addressed, however, is the direct association between neurobiological correlates of caregiving and parental sensitivity and how this association may explain (at least parts of) the complex pattern underlying intergenerational attachment transmission (see also Section 4 below).
3.3 Stability versus change
A further contentious area which SoNeAt may inform is the inter-relation of processes involved in stability and change in attachment-related individual differences across the life span, as reflected in Question 4 in Thompson et al. (Citation2021). Within this special issue, the study by Houbrechts et al. (Citation2021) directly addressed the issue of change in attachment by combining repeated self-reported attachment measurements (Experience in Close Relationships Scale Revised Child Version; ECR-RC) with neurobiological data acquisition as part of a longitudinal experimental design. More specifically, the authors measured attachment in children twice within one year and showed that its change – in particular for insecure-ambivalent attachment – was associated with children’s cortisol stress response and self-reported parental support obtained at the second measurement time point. Another longitudinal study included in this special issue by Puhlmann et al. (Citation2021) also obtained a measure of change, albeit not in attachment itself, but in brain structure as a function of attachment. To do so, in a sample of adolescents, the authors acquired a self-report measure of interindividual differences in attachment at study baseline (Relationship Scales Questionnaire; RSQ) and linked it to changes in brain structure assessed subsequently up to four times over a duration of four years with magnetic resonance imaging (MRI). Findings revealed accelerated cortical thinning in adolescents scoring higher on insecure (i.e. both avoidant and ambivalent) attachment, and especially so in adolescents who were classified as insecurely attached at a younger age.
Finally, a third longitudinal study by Merrill et al. (Citation2016) employed a somewhat different approach that did not primarily focus on change, but rather relative stability by prospectively linking early neurobiological patterns in 3-month-old infants with attachment classified during the SSP carried out 19 months later. The aim of this study was to see whether biological embedding of early life experiences may be reflected in observed attachment behavior at a later developmental stage. As neurobiological variables the authors assessed epigenome-wide blood DNA methylation – a relatively stable process related to changes in gene transcription (Boyce & Kobor, Citation2015) - as well as estimated monocyte proportion differences reflecting inflammatory processes. The data showed differential associations between early DNA methylation and inflammatory marker patterns and secure versus disorganized attachment classifications almost two years later, which suggests that biological embedding of social environments in an attachment-related context can occur in infants as early as 3-months-old. Overall, by employing the above longitudinal methodologies linking neurobiological and attachment data, we believe that SoNeAt can help to significantly advance our understanding of stability versus change in attachment across the entire life span (see also Section 4 below). Crucially, SoNeAt can shed light on this topic from different angles by measuring stability and/or change at several levels of investigation, thereby providing a rich and comprehensive account relating to a wide variety of neurobiological processes.
3.4 Psychophysiological mechanisms underlying IWMs and competence
Finally, several studies as part of this special issue considered various physiological and neural correlates of what might best be summarized as the processes underpinning the influence of (early) attachment on interpersonal outcomes – as addressed by Questions 3 (i.e. what are IWMs, and how do they operate) and 5 (see above) in Thompson et al. (Citation2021). In broader terms, we believe that the results of these studies reflect considerations of the competence hypothesis of attachment (see IJzendoorn, Citation1990). The competence hypothesis suggests that attachment experiences become cumulatively embedded within IWMs of attachment as expectations/predictions (see also ) which, in turn, predispose individuals to respond in certain ways to attachment-relevant information. Accordingly, a first study by Kungl et al. (Citation2022) revealed that 9 to 11-year-olds employing deactivating (i.e. insecure-avoidant) attachment-related strategies, as indexed by story-completions, showed an early ERP pattern (N250) reflective of increased vigilance to angry faces, irrespective of whether the faces belonged to their mothers or strangers. Furthermore, later ERP signals (LPP) from these children were relatively dampened, and, unlike those of their more secure counterparts, did not discriminate between mothers’ facial emotions. In combination, Kungl et al.’s data dovetails with a dual process model of insecure-avoidant attachment, involving an early hyper-vigilance to negative social information followed by a general attentional withdrawal from social stimulus processing. A second EEG study by Burra and Vrtička (Citation2022) assessed social information processing from faces conveyed by eye gaze and head orientation in an adult sample who filled in a self-report questionnaire of attachment (RSQ). The authors found an association between early neural face encoding (N170) and interindividual differences in attachment in that participants with higher insecure-ambivalent attachment scores were more likely to focus on eye gaze irrespective of head orientation and thus more local rather than global facial features.
Using functional MRI, a third study by Schoett et al. (Citation2021) examined the relation between brain activity to pictorial representations of separation and interindividual differences in attachment derived from the CAI in 9 to 11-year-olds. The authors report that activity within an affective subdivision of the anterior cingulate cortex was associated with more insecure-avoidant (and less secure) attachment, presumably reflecting differential experience of negative attachment-related emotions. This is akin to previous findings by Escobar et al. (Citation2013) linking insecure-avoidant attachment in adolescents derived from the Friends and Family Interview (FFI) with a negativity bias for faces in terms of a larger P1 and attenuated N170 component over the right hemisphere. Finally, a fourth study by Borelli et al. (Citation2021) assessed RSA as an index of parasympathetic nervous system activation and electrodermal activity (EDA) as an index of sympathetic nervous system activation. The authors did so in children ages 8–12 during a paradigm aimed to activate attachment needs and help-seeking, and related RSA and EDA data during that paradigm to children’s attachment derived from the CAI. Findings revealed that only during help-seeking, insecure-avoidant attachment was associated with higher EDA and thus greater sympathetic activation, whereas insecure-ambivalent attachment was associated with lower RSA and thus parasympathetic deactivation.
Altogether, this new SoNeAt evidence nicely illustrates that interindividual differences in attachment are linked to a variety of neurobiological processes across the life span. From an attachment theory perspective, it is likely that such neurobiological processes emerge as part of acquired socio-emotional competences reflective of IWMs of attachment in the form of expectations/predictions (see also ). These, in turn, affect emotion regulation strategies and tendencies to reflect on others’ mental states. While an increasing amount of SoNeAt research is concentrating on the behavioral and neurobiological consequences of such competences engraved within IWMs of attachment, it remains relatively under-studied how IWMs are neurobiologically maintained across time and contexts (see also Section 4 below).
4. Research gaps and future agenda
In taking stock of an increasingly firm evidence base within SoNeAt, we contend that an exciting turning point is now coming into view which will call for more complex and comprehensive study designs, especially regarding the core tenets or issues already addressed above (see also Panel B). Before considering this in detail for each of the core tenets, we note a disproportionate focus on the aversion and emotion regulation modules as part of NAM(D)A. This tendency not only characterizes papers included in this special issue, but also the field more broadly (see Long et al., Citation2020; White et al., Citation2020) and calls for a greater focus on the approach and mental state representation modules. We wrap up this section by briefly dwelling on the prospects of taking the SoNeAt approach into largely “uncharted territory,” i.e. leveraging its theory and methodology to better understand and examine intercultural differences in attachment (Keller & Bard, Citation2017) as well as mechanisms underpinning attachment-based interventions (Steele & Steele, Citation2018).
The gaps in the extant research bases on core issues involving individual differences in attachment notwithstanding, we deem basic research on the definition/assessment of attachment (i.e. which relationships qualify as “attachment” relationships and what are their defining characteristics) to offer promise. First, SoNeAt may help us gain a deeper understanding of how neural correlates may prove consistent or vary within the hierarchical network of attachment figures (e.g. mothers vs. fathers vs. romantic partners; biological vs. adoptive parents; see Girme & Overall, Citation2021) and compared to others who serve a less clear attachment function (e.g. friends or peers or unfamiliar individuals; see Coan et al., Citation2013 for pioneering work on these issues). Second, given that high-quality neuroscientific data is typically labor- and resource-intensive to collect and evaluate, it is also advisable to maximize the added value of SoNeAt research by pre-registering, collecting, comparing, and reporting all data (i.e. positive and null-findings) on multiple attachment measures (i.e. observations, narratives and self-reports) in the same individuals, even if they are relegated to online supplements.
Turning to the issue of parenting and intergenerational transmission, more complex multi-level SoNeAt designs are now in order including attachment measurements in both adults and children. Only studies encompassing all three levels of data (i.e. child, adult, social neuroscience modalities) will position us sufficiently for clarifying the key neurophysiological correlates involved in actual intergenerational transmission of attachment, as opposed to neurophysiological responses of parents, per se (which may be more or less relevant to their children). Inevitably, this work will require sample sizes powered sufficiently for considering all three variable types in a single statistical model (e.g. mediation analyses). Furthermore, SoNeAt work on transmission will benefit greatly from obtaining attachment-related (e.g. sensitivity, emotional availability) and neurophysiological correlates during caregiver-child interactions unfolding in real time by employing second-person social neuroscience methods. Here, SoNeAt may even guide identification of processes relevant to closing the transmission gap inasmuch as indices of second-person social neuroscience (e.g. INS) may mediate the link between attachment classifications of caregivers and children beyond variance attributable to caregiver sensitivity, per se.
As far as work on stability and change is concerned, future research should take up the challenge of employing multiple (longitudinal) within-person assessments of attachment and/or neuroscientific data to distinguish psychophysiological correlates of IWMs more prone to stability (across time and relationships) from those that are more prone to change. Besides the evident advantage of dismantling IWMs into more stable and more changeable subcomponents (with respective clinical implications, see below), we believe that such work on IWMs will be superior to work merely focusing on single attachment measurements, given that the latter run the risk of confounding the more trait-like neurophysiological correlates of core stable aspects of IWMs with more transient state-like and potentially assessment-specific aspects of IWMs (see Bosmans et al., Citation2020).
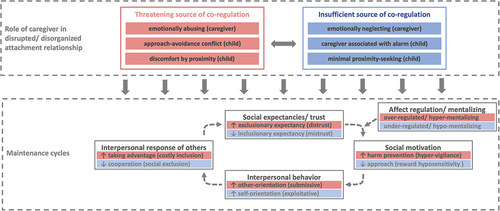
Finally, with regard to psychophysiological mechanisms underlying IWMs of attachment, we envisage future SoNeAt research to increasingly collect multiple psychophysiological indices in the same individuals (e.g. synchronization, brain function and structure, stress-hormones, etc.) - each reflecting different stages of the presumed neurocognitive mechanism of IWMs. Eventually, we hope this will provide the foundation for charting out the aetiological and temporal processes underlying formation, maintenance and consequences of IWMs. We have included a currently somewhat speculative proposal based on NAM(D)A (see Long et al., Citation2020; White et al., Citation2020) in which details two potential maintenance pathways or cycles related to caregivers functioning as threatening and insufficient sources of co-regulation in the wake of emotional abuse and neglect, respectively. These cycles seek to spell out the biobehavioral pathways whereby maltreatment by caregivers exerts deleterious effects on child development through a „pathogenic relationship“. In line with this, emerging neuroscientific, game-theoretical and clinical evidence now suggests that emotional maltreatment may act as the central „culprit“, exceeding and explaining much, if not most, of the variance in social neuroscience data and clinical outcomes over and above other physical forms of abuse and neglect (Schlensog-Schuster et al., Citation2022; Schulz et al., Citation2022). Accordingly, delineates two potential biobehavioral cycles, detailing the process of generalization of IWMs of attachment whereby pathogenic relationships to caregivers may precipitate formation and maintenance of dysfunctional interpersonal relationships across time and contexts. Generalization is thus viewed through the lenses of physiological correlates of expectancies or saliency (Lang et al., Citation1997; White et al., Citation2021), affect regulation and mentalization (Luyten et al., Citation2020), prevention and promotion-focused motivation (Scholer et al., Citation2019), as well as self- and other-orientation (Malti et al., Citation2019), among others.
Figure 3. Biobehavioral cycles based on NAMDA (see White et al., Citation2020) outlining the temporal processes underlying maintenance of individual differences in attachment over time and contexts.
Before concluding, we would like to also highlight the potential of SoNeAt for informing cultural and clinical research in the attachment field related to Questions 6 on cross-cultural differences, 8 on attachment-based (clinical) interventions and 9 on systems and services for families in Thompson et al. (Citation2021). For example, the former field may reap many benefits from comparing neural correlates of attachment classifications across cultures known to differ in the normativity of specific attachment behaviors (e.g. insecure-ambivalent in US vs. Japanese cultures; see Rothbaum et al., Citation2000). Regarding clinical interventions, SoNeAt may help to specify physiological components amenable to specific clinical attachment-focused intervention techniques (“ports of entry”) during certain developmental time-windows (see NAMDA; White et al., Citation2020). Besides advancing clinical attachment-related efforts, this work will also carry important theoretical implications, given that such intervention studies typically involve experimental manipulations of attachment-related constructs with causal implications (see e.g. Garnett et al., Citation2020; Wade et al., Citation2022).
5. Conclusion
This introduction attempts to chart SoNeAt’s future path (along with some „guard rails“) by calling attention to both its prospects and limits and outlining a programmatic framework for organizing current and future research into these lines of inquiry. To this end, we have laid out two crucial extensions of NAM(D)A, proposing a schema linking attachment system activation, co-regulation through social allostasis and energy conservation () as well as a cycle outlining the biobehavioral processes involved in maintaining individual differences in attachment (). We are confident that when we look back in the future, this special issue will demarcate a critical point of departure for ushering in a new era of SoNeAt research.
Acknowledgment
We would like to take the opportunity to thank everyone involved in the inception, conception and execution of this special issue (especially authors, reviewers and the chief editors) for their patience, precious time and continued support. Especially as this special issue had to take shape during what were challenging times, we feel it should fill all those who were part of it with pride to have accomplished this important contribution to SoNeAt research.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Notes
1. For the sake of consistency, we refer to organized attachment as secure, insecure-avoidant and insecure-ambivalent throughout. We are aware, of course, that other nomenclatures exist (e.g. autonomous, insecure-dismissing and insecure-anxious/preoccupied) as a function of attachment measures and reflecting different attachment traditions originating from social versus developmental psychology.
2. This is thought to primarily occur in cases where caregivers are inefficient co-regulators either because they themselves constitute sources of threat (e.g. abusing) or if they are chronically unavailable (e.g. neglectful caregiving) resulting in “fear without solution.”
References
- Alves, P. N., Foulon, C., Karolis, V., Bzdok, D., Margulies, D. S., Volle, E., & Thiebaut de Schotten, M. (2019). An improved neuroanatomical model of the default-mode network reconciles previous neuroimaging and neuropathological findings. Communications Biology, 2(1), Article 1. https://doi.org/10.1038/s42003-019-0611-3
- Atzil, S., Gao, W., Fradkin, I., & Barrett, L. F. Growing a social brain. (2018). Nature Human Behaviour, 2(9), 624–636. Article 9. https://doi.org/10.1038/s41562-018-0384-6
- Azhari, A., Gabrieli, G., Bizzego, A., Bornstein, M. H., & Esposito, G. (2020). Probing the association between maternal anxious attachment style and mother-child brain-to-brain coupling during passive co-viewing of visual stimuli. Attachment & Human Development, 0(0), 1–16. https://doi.org/10.1080/14616734.2020.1840790
- Borelli, J. L., Gaskin, G., Smiley, P., Chung, D., Shahar, B., & Bosmans, G. (2021). Multisystem physiological reactivity during help-seeking for attachment needs in school-aged children: Differences as a function of attachment. Attachment & Human Development, 0(0), 1–15. https://doi.org/10.1080/14616734.2021.1913874
- Bosmans, G., Bakermans-Kranenburg, M. J., Vervliet, B., Verhees, M. W. F. T., & van IJzendoorn, M. H. (2020). A learning theory of attachment: Unraveling the black box of attachment development. Neuroscience & Biobehavioral Reviews, 113, 287–298. https://doi.org/10.1016/j.neubiorev.2020.03.014
- Boyce, W. T., & Kobor, M. S. (2015). Development and the epigenome: The ‘synapse’ of gene–environment interplay. Developmental science, 18(1), 1–23. https://doi.org/10.1111/desc.12282
- Burra, N., & Vrtička, P. (2022). Association between attachment anxiety and the gaze direction-related N170. Attachment & Human Development, 0(0), 1–18. https://doi.org/10.1080/14616734.2022.2091337
- Cacioppo, J. T., & Berntson, G. G. (1992). Social psychological contributions to the decade of the brain: Doctrine of multilevel analysis. The American Psychologist, 47(8), 1019–1028. https://doi.org/10.1037/0003-066X.47.8.1019
- Cacioppo, J. T., Berntson, G. G., & Decety, J. (2010). Social neuroscience and its relationship to social psychology. Social cognition, 28(6), 675–685. https://doi.org/10.1521/soco.2010.28.6.675
- Cacioppo, J. T., & Cacioppo, S. (2018). Chapter three - loneliness in the modern age: An evolutionary theory of loneliness (ETL). In J. M. Olson (Ed.), Advances in experimental social psychology (Vol. 58, pp. 127–197). Academic Press. https://doi.org/10.1016/bs.aesp.2018.03.003
- Cacioppo, S., & Cacioppo, J. T. (2020). Introduction to social neuroscience. Princeton University Press.
- Cacioppo, J. T., Cacioppo, S., & Boomsma, D. I. (2014). Evolutionary mechanisms for loneliness. Cognition & Emotion, 28(1), 10.1080/02699931.2013.837379. https://doi.org/10.1080/02699931.2013.837379
- Cacioppo, J. T., Tassinary, L. G., & Berntson, G. G. (Eds.). (2016). Handbook of psychophysiology (4th ed.). Cambridge University Press. https://doi.org/10.1017/9781107415782
- Canterberry, M., & Gillath, O. (2012). Attachment and caregiving. In P. Noller & G. C. Karantzas (Eds.), The Wiley-Blackwell handbook of couples and family relationships (pp. 207–219). Wiley-Blackwell. https://doi.org/10.1002/9781444354119.ch14
- Coan, J. A. (2016). Toward a neuroscience of attachment. In Cassidy, J., & Shaver, P. R. (eds.), Handbook of attachment: Theory, research, and clinical applications, 3rd Edition (pp. 242–269). New York: Guilford Press.
- Coan, J. A., Beckes, L., & Allen, J. P. (2013). Childhood maternal support and social capital moderate the regulatory impact of social relationships in adulthood. International Journal of Psychophysiology, 88(3), 224–231. https://doi.org/10.1016/j.ijpsycho.2013.04.006
- Coan, J. A., & Sbarra, D. A. (2015). Social baseline theory: The social regulation of risk and effort. Current Opinion in Psychology, 1, 87–91. https://doi.org/10.1016/j.copsyc.2014.12.021
- Duschinsky, R., Bakkum, L., Mannes, J. M. M., Skinner, G. C. M., Turner, M., Mann, A., Coughlan, B., Reijman, S., Foster, S., & Beckwith, H. (2021). Six attachment discourses: Convergence, divergence and relay. Attachment & Human Development, 23(4), 355–374. https://doi.org/10.1080/14616734.2021.1918448
- Eklund, A., Nichols, T. E., & Knutsson, H. (2016). Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proceedings of the National Academy of Sciences, 113(28), 7900–7905. https://doi.org/10.1073/pnas.1602413113
- Escobar, M. J., Rivera-Rei, A., Decety, J., Huepe, D., Cardona, J. F., Canales-Johnson, A., Sigman, M., Mikulan, E., Helgiu, E., Baez, S., Manes, F., Lopez, V., & Ibañez, A. (2013). Attachment Patterns Trigger Differential Neural Signature of Emotional Processing in Adolescents. PLOS ONE. 8(8), e70247. https://doi.org/10.1371/journal.pone.0070247
- Feldman, R. (2017). The neurobiology of human attachments. Trends in Cognitive Sciences, 21(2), 80–99. https://doi.org/10.1016/j.tics.2016.11.007
- Garnett, M., Bernard, K., Hoye, J., Zajac, L., & Dozier, M. (2020). Parental sensitivity mediates the sustained effect of attachment and biobehavioral catch-up on cortisol in middle childhood: A randomized clinical trial. Psychoneuroendocrinology, 121, 104809. https://doi.org/10.1016/j.psyneuen.2020.104809
- Girme, Y. U., & Overall, N. C. (2021). A functional account of multiple internal working models: Flexibility in ranking, structure, and content across contents and time. In R. A. Thompson, J. A. Simpson, & L. J. Berlin (Eds.), Attachment: The fundamental questions (pp. 136–143). The Guilford Press.
- Gustison, M. L., & Phelps, S. M. (2022). Individual differences in social attachment: A multi-disciplinary perspective. Genes, Brain, and Behavior, 21(3), e12792. https://doi.org/10.1111/gbb.12792
- Hamilton, A. F. D. C. (2021). Hyperscanning: Beyond the hype. Neuron, 109(3), 404–407. https://doi.org/10.1016/j.neuron.2020.11.008
- Houbrechts, M., Cuyvers, B., Goossens, L., Bijttebier, P., Bröhl, A. S., Calders, F., Chubar, V., Claes, S., Geukens, F., Van Leeuwen, K., Noortgate, W. V. D., Weyn, S., & Bosmans, G. (2021). Parental support and insecure attachment development: The cortisol stress response as a moderator. Attachment & Human Development, 0(0), 1–13. https://doi.org/10.1080/14616734.2021.1907968
- IJzendoorn, M. H. V. (1990). Developments in cross-cultural research on attachment: Some methodological notes. Human development, 33(1), 3–9. https://doi.org/10.1159/000276498
- IJzerman, H., Coan, J. A., Wagemans, F. M. A., Missler, M. A., van Beest, I., Lindenberg, S., & Tops, M. (2015). A theory of social thermoregulation in human primates. Frontiers in Psychology, 6, 464. https://doi.org/10.3389/fpsyg.2015.00464
- Keller, H., & Bard, K. A. (Eds.). (2017). The cultural nature of attachment: Contextualizing relationships and development. The MIT Press. https://doi.org/10.7551/mitpress/11425.001.0001
- Kungl, M., Vrticka, P., Heinisch, C., Beckmann, M. W., Fasching, P. A., Ziegler, C., & Spangler, G. (2022). Deactivating attachment strategies associate with early processing of facial emotion and familiarity in middle childhood: An ERP study. Attachment & Human Development, 0(0), 1–19. https://doi.org/10.1080/14616734.2022.2132050
- Lang, P. J., Simons, R., Balaban, M. (1997). Attention and Orienting: Sensory and Motivational Processes . Psychology Press. https://doi.org/10.4324/9780203726457
- Long, M., Verbeke, W., Ein-Dor, T., & Vrtička, P. (2020). A functional neuro-anatomical model of human attachment (NAMA): Insights from first- and second-person social neuroscience. Cortex, 126, 281–321. https://doi.org/10.1016/j.cortex.2020.01.010
- Lowell, A. F., Dell, J., Potenza, M. N., Strathearn, L., Mayes, L. C., & Rutherford, H. J. V. (2021). Adult attachment is related to maternal neural response to infant cues: An ERP study. Attachment & Human Development, 0(0), 1–18. https://doi.org/10.1080/14616734.2021.1880057
- Luyten, P., Campbell, C., Allison, E., & Fonagy, P. (2020). The mentalizing approach to psychopathology: State of the art and future directions. Annual Review of Clinical Psychology, 16(1), 297–325. https://doi.org/10.1146/annurev-clinpsy-071919-015355
- Malti, T., Zhang, L., Myatt, E., Peplak, J., & Acland, E. L. (2019). Emotions in contexts of conflict and morality: Developmental perspectives. In Handbook of emotional development (pp. 543–567). Springer Nature Switzerland AG. https://doi.org/10.1007/978-3-030-17332-6_21
- Matthews, G. A., & Tye, K. M. (2019). Neural mechanisms of social homeostasis. Annals of the New York Academy of Sciences, 1457(1), 5–25. https://doi.org/10.1111/nyas.14016
- Merrill, S. M., Gladish, N., Fu, M. P., Moore, S. R., Konwar, C., Giesbrecht, G. F., MacIssac, J. L., Kobor, M. S., & Letourneau, N. L. (2021). Associations of peripheral blood DNA methylation and estimated monocyte proportion differences during infancy with toddler attachment style. Attachment & Human Development, 0(0), 1–30. https://doi.org/10.1080/14616734.2021.1938872
- Mesman, J., van IJzendoorn, M. & Sagi-Schwartz, A. …(2016). Cross-cultural patterns of attachment. In Cassidy, J., & Shaver, P. R. (eds.), The handbook of attachment: Theory, research, and clinical applications, 3rd Edition. New York: Guilford Press, 852–877.
- Mikulincer, M., Shaver, P. R., & Pereg, D. (2003). Attachment theory and affect regulation: The dynamics, development, and cognitive consequences of attachment-related strategies. Motivation and Emotion, 27(2), 77–102. https://doi.org/10.1023/A:1024515519160
- Miller, J. G., Vrtička, P., Cui, X., Shrestha, S., Hosseini, S. M. H., Baker, J. M., & Reiss, A. L. (2019). Inter-brain synchrony in mother-child dyads during cooperation: An fNIRS hyperscanning study. Neuropsychologia, 124, 117–124. https://doi.org/10.1016/j.neuropsychologia.2018.12.021
- Nguyen, T., Hoehl, S., & Vrtička, P. A guide to parent-child fNIRS hyperscanning data processing and analysis. (2021). Sensors, 21(12), 4075. Article 12. https://doi.org/10.3390/s21124075
- Nguyen, T., Schleihauf, H., Kayhan, E., Matthes, D., Vrtička, P., & Hoehl, S. (2020). The effects of interaction quality on neural synchrony during mother-child problem solving. Cortex, 124, 235–249. https://doi.org/10.1016/j.cortex.2019.11.020
- Nguyen, T., Schleihauf, H., Kayhan, E., Matthes, D., Vrtička, P., & Hoehl, S. (2021). Neural synchrony in mother–child conversation: Exploring the role of conversation patterns. Social cognitive and affective neuroscience, 16(1–2), 93–102. https://doi.org/10.1093/scan/nsaa079
- Nguyen, T., Schleihauf, H., Kungl, M., Kayhan, E., Hoehl, S., & Vrtička, P. (2021). Interpersonal neural synchrony during father–child problem solving: An fNIRS hyperscanning study. Child Development, 92(4), e565–580. https://doi.org/10.1111/cdev.13510
- Nummenmaa, L., Lahnakoski, J. M., & Glerean, E. (2018). Sharing the social world via intersubject neural synchronisation. Current Opinion in Psychology, 24, 7–14. https://doi.org/10.1016/j.copsyc.2018.02.021
- Puhlmann, L. M., Derome, M., Morosan, L., Kilicel, D., Vrtička, P., & Debbané, M. (2021). Longitudinal associations between self-reported attachment dimensions and neurostructural development from adolescence to early adulthood. Attachment & Human Development, 0(0), 1–19. https://doi.org/10.1080/14616734.2021.1993628
- Quadt, L., Esposito, G., Critchley, H. D., & Garfinkel, S. N. (2020). Brain-body interactions underlying the association of loneliness with mental and physical health. Neuroscience and Biobehavioral Reviews, 116, 283–300. https://doi.org/10.1016/j.neubiorev.2020.06.015
- Redcay, E., & Schilbach, L. (2019). Using second-person neuroscience to elucidate the mechanisms of social interaction. Nature reviews: Neuroscience, 20(8), 495–505. https://doi.org/10.1038/s41583-019-0179-4
- Rothbaum, F., Weisz, J., Pott, M., Miyake, K., & Morelli, G. (2000). Attachment and culture: Security in the United States and Japan. The American Psychologist, 55(10), 1093–1104. https://doi.org/10.1037/0003-066X.55.10.1093
- Saxbe, D. E., Beckes, L., Stoycos, S. A., & Coan, J. A. (2020). Social allostasis and social allostatic load: A new model for research in social dynamics, stress, and health. Perspectives on Psychological Science: A Journal of the Association for Psychological Science, 15(2), 469–482. https://doi.org/10.1177/1745691619876528
- Schilbach, L., Timmermans, B., Reddy, V., Costall, A., Bente, G., Schlicht, T., & Vogeley, K. (2013). Toward a second-person neuroscience. The Behavioral and Brain Sciences, 36(4), 393–414. https://doi.org/10.1017/S0140525X12000660
- Schlensog-Schuster, F., Keil, J., Von Klitzing, K., Gniewosz, G., Schulz, C. C., Schlesier-Michel, A., Mayer, S., Stadelmann, S., Döhnert, M., Klein, A. M., Sierau, S., Manly, J. T., Sheridan, M. A., & White, L. O. (2022). From maltreatment to psychiatric disorders in childhood and adolescence: The relevance of emotional maltreatment. Child Maltreatment, 10775595221134248. https://doi.org/10.1177/10775595221134248
- Schoett, M. J. S., Basten, U., Deichmann, R., Fiebach, C. J., & Fischmann, T. (2021). Brain responses to social cues of attachment in mid-childhood. Attachment & Human Development, 0(0), 1–15. https://doi.org/10.1080/14616734.2020.1840791
- Scholer, A. A., Cornwell, J. F. M., & Higgins, E. T. (2019). Regulatory focus theory and research: Catching up and looking forward after 20 years. In R. M. Ryan (Ed.), The oxford handbook of human motivation, 2nd ed (pp. 47–66). Oxford University Press.
- Schulz, C. C., von Klitzing, K., Deserno, L., Sheridan, M. A., Crowley, M. J., Schoett, M. J. S., Hoffmann, F., Villringer, A., Vrtička, P., & White, L. O. (2022). Emotional maltreatment and neglect impact neural activation upon exclusion in early and mid-adolescence: An event-related fMRI study. Development and Psychopathology, 34(2), 573–585. https://doi.org/10.1017/S0954579421001681
- Seeley, W. W. (2019). The salience network: A neural system for perceiving and responding to homeostatic demands. Journal of Neuroscience, 39(50), 9878–9882. https://doi.org/10.1523/JNEUROSCI.1138-17.2019
- Shakiba, N., & Raby, K. L. (2021). Attachment dimensions and cortisol responses during the strange situation among young children adopted internationally. Attachment & Human Development, 0(0), 1–15. https://doi.org/10.1080/14616734.2021.1896445
- Spangler, G., & Grossman, K. (1999). Individual and physiological correlates of attachment disorganization in infancy. In J. Solomon & C. George (Eds.), Attachment disorganization (pp. 95–124). The Guilford Press.
- Spangler, G., & Grossmann, K. E. (1993). Biobehavioral organization in securely and insecurely attached infants. Child Development, 64, 1439–1450. https://doi.org/10.2307/1131544
- Steele, H., & Steele, M. (2018). Handbook of attachment-based interventions. The Guilford Press.
- Thompson, R. A., Simpson, J. A., & Berlin, L. J. (Eds.). (2021). Attachment: The fundamental questions. Guilford Press.
- Thompson, R. A., Simpson, J. A., & Berlin, L. J. (2022). Taking perspective on attachment theory and research: Nine fundamental questions. Attachment & Human Development, 24(5), 543–560. https://doi.org/10.1080/14616734.2022.2030132
- van IJzendoorn, M. H., & Bakermans-Kranenburg, M. J. (2019). Bridges across the intergenerational transmission of attachment gap. Current Opinion in Psychology, 25, 31–36. https://doi.org/10.1016/j.copsyc.2018.02.014
- Verhage, M. L., Schuengel, C., Madigan, S., Fearon, R. M. P., Oosterman, M., Cassibba, R., Bakermans-Kranenburg, M. J., & van IJzendoorn, M. H. (2016). Narrowing the transmission gap: A synthesis of three decades of research on intergenerational transmission of attachment. Psychological Bulletin, 142(4), 337–366. https://doi.org/10.1037/bul0000038
- Verhage, M. L., Tharner, A., Duschinsky, R., Bosmans, G., & Fearon, R. M. P. (in press). Editorial perspective: On the need for clarity about attachment terminology. Journal of Child Psychology and Psychiatry. https://doi.org/10.1111/jcpp.13675
- Vrtička, P. (2017). The social neuroscience of attachment. In A. Ibáñez, L. Sedeño, & A. M. García (Eds.), Neuroscience and social science: The missing link (pp. 95–119). Springer International Publishing. https://doi.org/10.1007/978-3-319-68421-5_5
- Vrticka, P., & Vuilleumier, P. (2012). Neuroscience of human social interactions and adult attachment style. Frontiers in human neuroscience, 6, 6. https://doi.org/10.3389/fnhum.2012.00212
- Wade, M., Parsons, J., Humphreys, K. L., McLaughlin, K. A., Sheridan, M. A., Zeanah, C. H., Nelson, C. A., & Fox, N. A. (2022). The Bucharest early intervention project: Adolescent mental health and adaptation following early deprivation. Child Development Perspectives, 16(3), 157–164. https://doi.org/10.1111/cdep.12462
- White, L. O., Bornemann, B., Crowley, M. J., Sticca, F., Vrtička, P., Stadelmann, S., Otto, Y., Klein, A. M., & Klitzing, K. (2021). Exclusion expected? Cardiac slowing upon peer exclusion links preschool parent representations to school‐age peer relationships. Child Development, 92(4), 1274–1290. https://doi.org/10.1111/cdev.13494
- White, L. O., Schulz, C. C., Schoett, M. J. S., Kungl, M. T., Keil, J., Borelli, J. L., & Vrtička, P. (2020). Conceptual analysis: A social neuroscience approach to interpersonal interaction in the context of disruption and disorganization of attachment (NAMDA). Frontiers in Psychiatry, 11. https://doi.org/10.3389/fpsyt.2020.517372
- Xu, N., & Groh, A. M. (2021). The significance of mothers’ attachment representations for vagal responding during interactions with infants. Attachment & Human Development, 0(0), 1–21. https://doi.org/10.1080/14616734.2021.1876615