
ABSTRACT
Introduction
Advances in high-throughput sequencing have greatly advanced our understanding of long non-coding RNAs (lncRNAs) in a relatively short period of time. This has expanded our knowledge of cancer, particularly how lncRNAs drive many important cancer phenotypes via their regulation of gene expression.
Areas covered
Men of African descent are disproportionately affected by PC in terms of incidence, morbidity, and mortality. LncRNAs could serve as biomarkers to differentiate low-risk from high-risk diseases. Additionally, they may represent therapeutic targets for advanced and castrate-resistant cancer. We review current research surrounding lncRNAs and their association with PC. We discuss how lncRNAs can provide new insights and diagnostic biomarkers for African American men. Finally, we review advances in computational approaches that predict the regulatory effects of lncRNAs in cancer.
Expert opinion
PC diagnostic biomarkers that offer high specificity and sensitivity are urgently needed. PC specific lncRNAs are compelling as diagnostic biomarkers owing to their high tissue and tumor specificity and presence in bodily fluids. Recent studies indicate that PCA3 clinical utility might be restricted to men of European descent. Further work is required to develop lncRNA biomarkers tailored for men of African descent.
1. Introduction
Prostate cancer (PC) is one of the most prevalent cancers worldwide, particularly affecting men living a western lifestyle and of African descent, suggesting risk factors that are genetic, environmental, and socioeconomic in nature. Despite major advances in the diagnosis and treatment of PC, the disease remains a global health problem. PC is the second most diagnosed non-skin cancer in men worldwide [Citation1,Citation2]. Currently, there is considerable interest in lncRNAs and their regulatory roles in developmental processes and disease/cancer stages [Citation3,Citation4]. Prior to high-throughput sequencing technologies becoming common place, lncRNAs were simply considered transcriptional ‘noise’ and largely ignored by the scientific community [Citation5–7]. With increased sequencing sensitivity in combination with innovative bioinformatic tools, in the past decade, researchers have started to uncover the functional roles of lncRNAs. Two emerging discoveries include the realization that they act upon a wide range of cell processes and are differentially expressed in disease states, including various cancers [Citation8,Citation9].
Computational predictions have identified over 10,000 lncRNAs, and researchers have been keen to characterize them functionally, identify their regulatory roles, and exploit this knowledge in biomedical research, diagnostics, and therapeutic development [Citation10]. In parallel, bioinformatics methods are also being rapidly improved to better predict the regulatory roles of lncRNAs in cancer and determine their full extent as regulatory modulators. This review provides a detailed overview of lncRNAs, particularly in the context of PC. First, we cover lncRNAs, their history, newly discovered functions, and correlation with cancer. We then review PC, racial differences in disease presentation, and why lncRNAs are relevant in this context. Lastly, we provide a summary of tools dedicated to deciphering the functional roles of lncRNAs.
2. What is a long non-Coding RNA?
lncRNAs are defined as transcripts longer than 200 nucleotides that are putatively not translated into proteins [Citation8]. Both short (≤200 base pairs) and lncRNAs may contain coding regions [Citation11]. Short non-coding transcripts, in particular microRNAs (miRNAs), have been studied more extensively than lncRNAs and contribute to the development and progression of many cancers [Citation12]. Prior to the genomic era, lncRNAs were considered ‘junk RNA’ and were largely ignored by the scientific community. The first lncRNA to be identified was h19 in 1984. Although at first mistakenly categorized as a messenger RNA (mRNA) [Citation13], h19 has subsequently been recognized as an epigenetic regulator, differentially expressed in several cancers where it acts as a tumor suppressor [Citation3,Citation14–16]. In 2002, lncRNAs were finally assigned as functional RNA molecules by the Functional Annotation of the Mammalian Genome (FANTOM) project [Citation17]. Like mRNAs, lncRNAs are transcribed by RNA polymerase II, polyadenylated, capped, and spliced [Citation18,Citation19]. Subcellular localization provides a lens to define lncRNA functional classes [Citation7,Citation20]. The identification and number of lncRNAs is continually increasing; however, many remain poorly annotated and are not experimentally validated [Citation12,Citation21].
As of January 2021, LNCBook, a curated lncRNA database contains 268,848 documented lncRNAs (obtained from publicly accessible databases including GENCODE v27, NONCODE v5.0, LNCipedia v4.1 and MiTanscriptome beta). However, there is a large gap between documented lncRNAs and lncRNAs, which have a known link to a molecular or biological function. Of the 268,848 recorded lncRNAs on LNCBook, only 1,867 have been categorized as experimentally validated (linked with functional mechanisms/biological processes and identified in relevant publications) [Citation21,Citation22]. LncRNAs are classified as either 1) intergenic (lincRNAs), transcribed from intergenic regions located between two genes [Citation5,Citation23,Citation24] 2) bidirectional (divergent IncRNAs) transcribed on the reverse strand of a coding transcript located no more than 1000 base pairs away [Citation5,Citation24] 3) intronic lncRNAs, transcribed only from intronic regions of protein coding genes, 4) antisense lncRNAs transcribed from antisense strands of protein coding genes and 5) sense lncRNAs transcribed from sense strands of protein coding genes [Citation6,Citation25,Citation26].
Understanding how lncRNAs are regulated and how they in turn regulate their target genes is an important research area [Citation27]. Thus far, several functional roles of lncRNAs have been established. The most common IncRNA functional groups include guides (e.g. HOTAIR) [Citation6,Citation28], scaffolds (e.g. TERC) [Citation6,Citation29], signals (e.g. XIST) [Citation6,Citation7] and decoys (e.g. MALAT1) [Citation6,Citation30]. There are also a series of documented multifunctional IncRNAs. An example is NEAT1 [Citation31]. NEAT1 was identified as differentially expressed in numerous tumor contexts, including lung, colorectal, and prostate cancer [Citation31]. NEAT1 is an important mediator in the regulation of clear cell renal cell carcinoma (ccRCC) progression and predicts poor prognosis and decreased survival in patients with ccRCC [Citation7,Citation32–34]. Guide IncRNAs can bind and tether epigenetic protein complexes to a target locus directing gene expression either in cis or in trans. Guide IncRNAs can direct chromatin-modifying enzymes (e.g. histone methylases, acetylases, and deacetylases) to a particular gene target locus, which can lead to the activation or repression of local genes [Citation35,Citation36]. Scaffold IncRNAs recruit epigenetic complexes together, thus acting as a molecular scaffold. They recruit RNA-binding factors to form RNA-protein complexes, which promote or inhibit gene transcription [Citation35,Citation36]. Signal IncRNAs possess signaling properties that promote guidance of epigenetic complexes to specific locations. These IncRNAs possess enhancer functions, which alter the structure of chromatin and guide transcriptional complexes to gene targets enabling transcriptional activity [Citation35,Citation36]. Finally, decoy IncRNAs can lure epigenetic complexes to different genomic locations. These IncRNAs can recruit and decoy a transcriptional complex to a distinct location for suppression of gene expression [Citation35,Citation36]. Many reviews have documented the molecular functions of lncRNAs (for example, reference 23), however, none have considered the computational aspects of predicting lncRNA functions and their targets. Our review addresses this shortcoming.
3. Long Non-coding RNAs and Prostate Cancer
The number of patients presenting with urological cancers (malignancies of the genital and urinary organs of men and the urinary organs of women) is on the increase [Citation37,Citation38]. Currently, urological cancers of the prostate, bladder, and kidney stand among the top 10 most diagnosed cancers in men [Citation39]. Despite their frequency, biomarkers for urological cancers are limited. Those that are available typically lack sensitivity in low-grade disease, e.g. urine cytology for bladder cancer and low specificity, e.g. Prostate Specific Antigen (PSA) [Citation39–41]. lncRNAs are now frequently being linked with urological cancers, and many have been designated as playing roles in the development and progression of these malignancies including prostate cancer [Citation42]. summarizes selected lncRNAs that are differentially expressed in urological cancers, which may have potential as biomarkers or therapeutic targets [Citation6,Citation7,Citation31].
Table 1. Urological cancer-associated lncRNAs
Despite major advances in the diagnosis and treatment of PC, the disease remains a global health problem. PC is the second most diagnosed non-skin cancer in men worldwide [Citation60]. Recent estimates indicate that 1 in 7 men will be diagnosed with PC during their lifetime. In 2018 alone, 359,000 recorded deaths from PC occurred globally [Citation61,Citation62]. Racial differences are common in PC. Men of African descent are adversely affected in terms of both incidence and mortality rates compared to European-based counterparts. lncRNAs are now considered to contribute to these racial differences [Citation63,Citation64].
Evidence to date suggests that lncRNAs can alter gene expression at both transcriptional and post-transcriptional levels, thus playing roles in important biological processes including carcinogenesis and metastasis [Citation65]. Some lncRNAs function as tumor suppressors, and possess the ability to interact with oncogenes, inhibit or accelerate proliferation, and influence apoptosis [Citation30,Citation66,Citation67].
Other lncRNAs are commonly aberrantly expressed, deleted, amplified, and/or mutated in multiple cancers [Citation9,Citation68]. lncRNAs have been shown to have roles in numerous forms of cancer [Citation69]. displays lncRNAs correlated with numerous cancer types. Specific cancer types and relevant body tissues are highlighted. provides a summary of lncRNAs, which have been identified as differentially expressed in various malignancies.
Figure 1. Cancer associated lncRNAs. Key lncRNAs associated with various cancers are highlighted as to the body tissue and cancer type
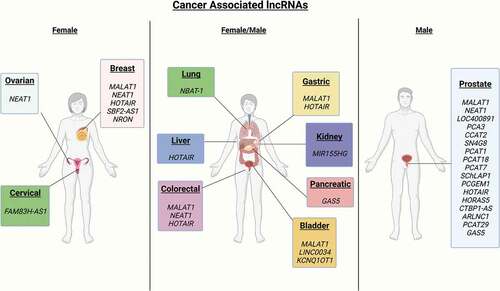
Table 2. Cancer-associated lncRNAs
Presently, three areas that merit particular attention are; 1) Identification of novel diagnostic and prognostic biomarkers for PC, with an emphasis on diagnostic biomarkers for early-stage PC, which differentiates low-risk from high-risk tumors [Citation40,Citation84], 2) An urgent need to better understand biological differences in disease presentation in men of African and Northern European descent [Citation85] and 3) Efficient therapies for advanced and castrate-resistant PC [Citation86]. LncRNAs are emerging as biomarkers or therapeutic targets for these areas, as explained below.
4. lncRNAs as prostate cancer biomarkers
It has been widely accepted that the identification of biomarkers should not exclusively focus upon protein coding genes [Citation87]. A key factor for biomarker utility is whether the proposed biomarker is detectable in bodily fluids. Liquid-based biomarkers provide many benefits over traditional tissue-based biomarkers from a ‘no surgery requirement and recovery time’ perspective, painless non-invasive testing, decreased costs, and more rapid diagnostics [Citation88]. lncRNAs are attractive as PC biomarkers owing to their high tissue and tumor specificity [Citation27] and are detectable in bodily fluids, including PCA3 and PCAT18 [Citation88,Citation89].
Biomarkers with high specificity and sensitivity are particularly needed to help address the over-treatment of clinically insignificant PC. The over diagnosis and treatment of low-risk PC remains a common problem, primarily due to the use of Prostate Cancer Specific Antigen (PSA) tests [Citation90] which offer high sensitivity but low specificity [Citation40]. The introduction of the PSA test in the early 1990s was highly criticized for the over-diagnosis and the peak of PC incidence rates [Citation91–93]. Furthermore, PSA tests have poorer diagnostic performance in men of African descent [Citation94]. Therefore, a current need in PC research is the identification of diagnostic biomarkers, which can not only offer high specificity and sensitivity but are also universally reliable for all races.
PC-specific lncRNAs are compelling as diagnostic biomarkers for clinical use [Citation27]. Possibly, the most well-known is PCA3 (Prostate Cancer Specific Antigen 3). PCA3 was first observed to be highly expressed in PC tumors in 1999 and has since been characterized as overexpressed in 95% of all PC cells [Citation95,Citation96]. Urine-based measurements of PCA3 provide greater PC specificity than PSA testing [Citation12]. In 2012, the PROGENSA PCA3 assay developed by Gen-Probe Inc was the first lncRNA PC biomarker to obtain Food and Drug Administration (FDA) approval. This lncRNA molecular test is used to help guide urologists in determining the need for repeat biopsies in men aged 50 years or older who had previously presented with a negative biopsy result [Citation97,Citation98]. However, its practicability as a first use test remains controversial and it cannot predict PC aggressiveness [Citation99,Citation100]. Several studies have also documented PCA3 expression in PC progression; however, further research is required to confirm its feasibility as a PC prognostic biomarker [Citation101]. Additionally, PCA3’s clinical utility might be restricted to men of European descent. O’Malley, et al. examined the clinical significance of the urinary biomarkers (PCA3 & T2ERG) in 718 African American and non-African American men, and assessed whether they provided increased utility for the detection of PC. Their results indicated that PCA3 and T2ERG enhanced clinical utility; however, this result was only observed in non-African American men [Citation102]. A major limitation of this study was the relatively small number of African American samples [Citation103] in comparison to non-African American men. Further research with larger ethnically diverse datasets would facilitate a more rigorous assessment.
Other studies have yielded varied outcomes in contrast to the initial findings from the O’Malley study. Feibus, et al. conducted a study using 304 patients, 60% of which were African American and noted that measurements of PCA3 enhanced predictability of high-grade PC in African American men [Citation94], while an earlier study by Adam et al. using South African men challenged these results. Of 105 South African men referred for prostate biopsy, Adam et al., found that PCA3 levels were not significantly increased in black men who had tested positive for the disease [Citation103]. Again, larger cohorts would provide greater statistical power.
Several other lncRNAs have been established as possible biomarkers of PC, including PCAT1 (Prostate Cancer Associated Transcript 1). PCAT1 is a highly PC-specific lncRNA [Citation27] and was upregulated in patients presenting with a high-risk (Gleason score ≥7) PC and metastatic disease [Citation27,Citation104]. Prensner et al., 2011 identified 121 unannotated PC ncRNAs through the analysis of 102 prostate tumor tissues and cell lines, among these was PCAT1 [Citation105]. PCAT1 was significantly overexpressed in most PC tumors but particularly in tumors that had metastasized. The group remarked that PCAT1 was likely contributing to cell proliferation and potentially could serve as a urine-based biomarker given the prior success with PCA3 [Citation106]. Interestingly, PCAT1 is located at chromosome 8q24, a chromosome location frequently correlated with PC risk due to its common amplification and the presence of PC susceptibility single nucleotide polymorphisms (SNPs) [Citation107,Citation108]. Yhan et al. and Prensner et al. concluded that PCAT1 supports cell proliferation in PC with Prensner and colleagues noting that it may represent a suitable candidate for biomarker development [Citation104,Citation109]. PCAT18 is overexpressed in PC. Using tissue samples from patients diagnosed at the Stephanshorn clinic in Switzerland and plasma samples collected in Canada, Crea et al. found that PCAT18 was overexpressed in the plasma of patients with localized and advanced forms of the disease compared to healthy individuals. The results from this study displayed a positive correlation between PCAT18 and PC stage, with the highest levels observed in metastatic castrate resistant PC. Thus, PCAT18 could have prognostic value, including to discriminate between localized and metastatic forms of PC, and is a possible therapeutic target for advanced PC [Citation110]. MALAT1 (Metastasis Associated Lung Adenocarcinoma Transcript 1) is a lncRNA originally correlated with lung cancer; however, it has been found to be overexpressed in several cancers including PC. In 2013, Ren et al. identified MALAT1 as detectable in plasma and serum and suggested that it had the potential to be a diagnostic biomarker [Citation111]. Wang et al. identified MALAT1 as overexpressed in urine samples of Chinese PC patients and noted its potential as a urine-based biomarker [Citation112].
lncRNAs correlated with advanced PC include PCAT7 (Prostate Cancer Associated Transcript 7), SChLAP1 (SWI/SNF Complex Antagonist Associated with Prostate Cancer 1), and PCAT18. PCAT7 has previously been correlated with disease progression in nasopharyngeal and non-small cell lung cancer [Citation113,Citation114]. Lang, et al., 2020 noted that PCAT7 has the potential to be a therapeutic target for men presenting with PC bone metastasis via disruption of the constitutive loop between PCA7 and TGF‐β signaling [Citation115]. PCAT7 also has the potential to be a prognostic biomarker. SChLAP1 (SWI/SNF ‘Complex Antagonist Associated With Prostate Cancer’) has been proposed as a biomarker of lethal PC. Cimadamore et al. documented that lncRNA SChLAP1 is critical in the development and progression of PC [Citation116]. Prensner et al. identified that SChLAP1 was highly expressed in ~25% of PCs analyzed (including metastatic PC) and may promote tumor cell invasion (in vitro) as well as metastasis (in vivo) [Citation117–119]. Additionally, Prensner et al. observed SChLAP1 expression in urine and correlated with higher-risk PC patients suggesting its potential as a biomarker to differentiate between low-risk and high-risk tumors [Citation120]. SChLAP1 has also been associated with adverse clinical outcomes in postradical prostatectomy and considered a potential tissue-based prognostic biomarker by Mehra, et al. [Citation118]. Similarly, Chua et al. suggested SChLAP1 expression was linked with increased biochemical relapse and metastasis, cribriform architecture and intraductal carcinoma [Citation121].
PCGEM1 is highly PC specific, and has been linked to PC progression by androgen receptor transcriptional co-regulators and could also decrease apoptosis in PC cell lines [Citation117,Citation122,Citation123]. Petrovics et al. observed that PCGEM1 expression was significantly higher in PC cells of men of African descent compared to men of Northern European descent [Citation122]. An additional study by Lee et al. observed similar results where PCGEM1 was significantly overexpressed in prostate tumors of African American, but not European American men [Citation124].
Despite a better understanding of the roles of lncRNAs and the number identified as potential biomarkers of localized and advanced PC, few lncRNAs have met the success of PCA3. A primary reason for this is that lncRNA research is still in its infancy with thousands of predicted lncRNAs requiring laboratory experimental validation of their utility and confirmation in separate patient cohorts. Moreover, the overwhelming majority of candidate biomarkers fail to achieve clinical utility [Citation89]. The candidate PC biomarkers summarized above require analytical and clinical validation to better assess their utility. To date, numerous articles have been published detailing lncRNAs directly related to PC (refer to references 38, 51, 53, 54, 55, and 68); however, our review differs in two specific ways: 1) more in-depth detail relating to PC and racial differences, and 2) computational methods and software now available for identifying potential lncRNA biomarkers in the disease.
5. Racial differences in Prostate Cancer
Racial differences are common among cancers, and examples include multiple myeloma, colorectal, and breast cancer. Multiple myeloma is the most common hematologic cancer in patients of African descent [Citation125]. Although it is a rare disease, diagnoses are twice as high in patients of African descent compared to those of European descent. Reasons for these differences are currently unknown and the disease remains incurable [Citation126–128]. In the USA, colorectal cancer patients of African descent experience decreased stage-specific survival in comparison to patients of European descent irrespective of socioeconomical background, age, treatment, and stage at diagnosis [Citation129]. Invasive breast cancer mortality rates also differ between patients of African and Northern European descent. In the USA, African American women are more likely to die of the disease than European American women [Citation130]. African American women also experience worse prognosis and decreased survival times compared to European American women regardless of their socioeconomical background, age, and stage at diagnosis [Citation131].
Prostate cancer has one of the largest racial disparities of all cancer types. In the U.S.A., African Americans have among the highest mortality rates and lowest survival rates of any race or ethnic group [Citation132]. PC is also the top cause of cancer mortality among men of African descent living in Caribbean and sub-Saharan Africa [Citation133]. Similar disparities exist in the UK, where men of African descent are 2–3 times more likely to develop PC than Caucasian men and have a 30% higher mortality rate [Citation134]. Nonbiological factors contributing to PC racial disparities include diminished societal trust in the medical community, level of education attained, and financial concerns. Access to health care plays an important role in PC mortality. However, African American men have higher PSA values and higher tumor grades even at the same cancer stage, compared with non-African American men within the same health-care system, such as the Veterans Administration in the U.S.A. [Citation135]. The reasons underlying these racial differences remain an area of active research. Many biological explanations have been presented, including dietary factors such as alcohol intake, vitamin D deficiency, and lycopene and isoflavone consumption [Citation135–139]. Biological factors include genetic alterations – African Americans have higher rates of variations in tumor suppressor genes, such as EPHB2, and are susceptible to higher prevalence of chromosome 8q24 variants [Citation64,Citation85,Citation107,Citation140–142].
An emerging area of research is the role of lncRNAs in cancer development and progression in the context of racial differences. Frequently, lncRNAs are reported to be differentially expressed in numerous cancers and are known to be involved in tumorigenesis and metastasis [Citation65]. However, there is a lack of research investigating lncRNA differences between racial groups.
Much remains to be uncovered about the mechanisms by which lncRNAs regulate transcriptional and translational processes that drive oncogenesis. The lncRNA previously found to be significantly correlated with both PC and men of African ancestry is PCGEM1 (Prostate Cancer Gene Expression Marker 1). PCGEM1 is highly PC specific and has been linked to PC progression by androgen receptor transcriptional co-regulators and attenuates apoptotic processes in PC cell lines [Citation117,Citation122,Citation123]. Petrovics et al. identified PCGEM1 expression as being significantly higher in PC cells of men of African descent compared to men of Northern European descent [Citation122]. An additional study by Lee et al., 2016, observed similar results where PCGEM1 was significantly overexpressed in prostate tumors of African American but not in European American men [Citation124].
Recently, Yuan, et al., 2020, examined genomic and transcriptomic differences among African American and European American PC patients. The 470 samples that were analyzed consisted of RNA-seq transcriptomes from 57 African American and 413 European American patients from The Cancer Genome Atlas project (TCGA) prostate adenocarcinoma (PRAD) portal. The significantly lower number of African American samples represents a common challenge in health disparities research, with genomic studies typically being constrained to populations of European descent [Citation143].
Yuan, et al., identified 1868 (31%) of the differentially expressed transcripts between African American and European American patients as lncRNAs. Patient and disease characteristics (age and Gleason score) were accounted for [Citation64]. Of these differentially expressed lncRNAs, all were significantly enriched and regulated at least one target gene identified by the Long Noncoding RNA Heterogeneous Regulatory Network integrator (LongHorn) algorithm. LongHorn predictions are based on reverse-engineered canonical interactions, determined experimentally as part of the Encyclopedia of DNA Elements (ENCODE) project including the crosslinking and immunoprecipitation assay (eCLIP) and chromatin immunoprecipitation sequencing (ChIP-seq) data [Citation30,Citation64]. The researchers also showed that nearly 90% of lncRNAs differentially expressed between racial groups remain uncharacterized, underlining the need for continued lncRNA research. Of the 28 characterized lncRNAs differentially expressed between African and European American men, 13 were PC specific including PCA1 and PCAT10. The researchers noted that the differentially expressed lncRNAs potentially contribute to disease progression and influence PC racial differences. Further work is required to understand the regulatory functions of uncharacterized lncRNAs differentially expressed between racial groups [Citation64].
Given the evidence highlighting the potential roles of lncRNAs in PC tumorigenesis, there is an urgent need to understand the functional consequences of lncRNA differences between African and Northern European PC patients. One of the pressing challenges for integrative computational biology and statistical genetics in racial disparities research is predicting genotype-to-phenotypes. The ability to identify the key drivers among a multitude of interacting molecules is challenging. Additional obstacles include the rapid growth of data, the and unavailability of data through issues with incompleteness, inaccuracies, heterogeneity, and data silos [Citation144,Citation145]. LncRNAs may be prioritized according to their impact upon gene regulatory networks through graph theoretic and systems biology approaches [Citation146,Citation147]. Understanding the biological underpinnings of racial differences in PC will ultimately provide an opportunity to help eliminate them by identifying possible PC diagnostic and prognostic biomarkers as well as potential therapeutic targets for men of African descent.
6. LncRNAs as potential therapeutic targets for advanced and castrate resistant Prostate Cancer
Advanced PC is traditionally defined as a disease, which has spread (metastasized) outside the prostate gland [Citation148]. Cancers generally metastasize preferentially to specific distant organ sites [Citation149]. In most cases of advanced PC, metastasis targets the bones [Citation150]. Androgen Deprivation Therapy (ADT) is clinically used as a palliative treatment for patients diagnosed with advanced PC [Citation151–153]. In healthy men, the androgen testosterone and its derivative dihydrotestosterone are required for normal functioning of the prostate, however, researchers have identified it is also vital for PC tumor growth [Citation154]. Despite initial success with ADT, nearly all patients relapse and develop castrate-resistant PC (CRPC) after a mean duration of 2–3 years [Citation155,Citation156]. CRPC is defined as the progression of PC despite ADT and may present as one or any combination of a steady growth in PSA levels, progression of preexisting disease, or the emergence of new metastases [Citation157] Currently CRPC remains incurable and most patients with non-metastatic tumors will develop metastasis within 36 months of diagnosis, 90% of which will experience metastasis to the bone [Citation158,Citation159]. The survival probability of metastatic PC patients depends on a variety of circumstances from which organ sites the disease has metastasized to, the patient response to treatment, and the extent of the metastasis [Citation160]. However, once the disease has spread, survival rates are bleak with 29% five-year survival [Citation161]. Drug resistance in cancer therapies continuously present as a major obstacle [Citation162]. lncRNAs are recognised as possible drivers of drug resistance and may be modulated to enhance drug sensitivity in cancer therapy [Citation162]. Drug resistance refers to tolerance against pharmaceutical drugs [163]; drug sensitivity may be restored after tolerance has been developed Citation163[Citation162].
In PC, there remains an urgent need to identify therapeutic targets for CRPC and advanced forms of the disease. lncRNAs are being proposed as alternative therapeutic targets for slowing the progression of PC, several of which include HOTAIR (Hox Transcript Antisense Intergenic RNA), MALAT1 (Metastasis Associated Lung Adenocarcinoma Transcript 1), and GAS5 (Growth Arrest Specific Transcript 5) [Citation87]. The majority of lncRNAs identified as potential CRPC and metastatic PC therapeutic targets are specifically associated with the androgen receptor (AR) signaling pathway [Citation164,Citation165]. Notably, the AR is vital in all stages of PC because it stimulates cell growth and survival [Citation166]. Thus, most newly suggested therapeutic targets for PC center around this [Citation167,Citation168].
Several lncRNAs proposed as candidate therapeutic targets for CRPC and metastatic PC are primarily associated with the AR, including HORAS5 (LINC00161) [Citation169], CTBP1-AS (C-Terminal Binding Protein 1 Antisense RNA) [Citation170], and PCAT29 (Prostate Cancer Associated Transcript 29) [Citation166]. lncRNAs can be directly influenced by androgen regulation [Citation171], and many have been documented to participate in the advancement of PC through direct contact with the AR [Citation172].
Parolia et al., 2019 found that HORAS5 facilitated the progression of CRPC in an AR-dependent manner by regulating AR mRNA stability [Citation169] and noted that reduction in HORAS5 expression had a knock-on effect on the expression of the AR and its associated cancerous AR targets such as KIAA0101 [Citation169]. lncRNA CTBP1-AS has been observed to have similar results for both CRPC and hormone dependent PC principally by promoting tumor growth [Citation170]. Yakayama, et al. found that CTBP1-AS can suppress expression levels of CTBP1, a transcriptional corepressor known to regulate expression of tumor suppressors, and several genes involved with cell apoptosis [Citation170,Citation173]. CTBP1-AS had androgen-dependent functions, which could hinder the expression of tumor suppressors in both hormone dependent PC and CRPC suggesting potential as a therapeutic target [Citation170]. PCAT29 is a PC-specific lncRNA, which has also been linked to the AR [Citation166] Malik, et al. identified PCAT29, as a tumor suppressor of PC, which is suppressed by the AR. PCAT29 has PC suppressive effects including hindering cellular proliferation, tumor growth, cellular migration, and metastasis [Citation166]. Lower expression of PCAT29 is correlated with decreased survival in PC patients [Citation166].
Several other studies have identified lncRNAs linked to PC and the AR. An associated lncRNA is HOTAIR [Citation174]. HOTAIR is categorized as ubiquitously expressed and associated with multiple illnesses, such as cardiovascular diseases, and cancers including breast, gastric, and colorectal [Citation175]. Zhang, et al., 2015 identified that HOTAIR is repressed by androgens and overexpressed in CRPC [Citation176]. Further research supports the hypothesis that HOTAIR contributes to the progression from hormone dependent to CRPC [Citation177]. Taken together, these results suggest HOTAIR is a key player in numerous cancers including CRPC [Citation176]. Another lncRNA recently associated with the AR is ARLNC1 (AR-regulated long non-coding RNA 1). Zhang, et al., 2018 identified ARLNC1 was strongly correlated with AR signaling in the advancement of PC and deemed it a possible therapeutic target with further detailed study [Citation178]. Several additional lncRNAs have been correlated with the androgen receptor (AR) signaling pathway in PC either to inhibit or promote tumor growth. Therefore, lncRNAs can act as key drivers of PC and have potential as therapeutic targets. presents a schematic overview of AR associated lncRNAs that either inhibit or promote PC pathophysiology [Citation164]. This schematic summarizes key lncRNAs, their role in up- and down-regulation of the AR, the role of these key lncRNAs in promoting and inhibiting carcinogenesis, and interplay with miRNAs and proteins.
Figure 2. Androgen receptor (AR) signaling pathway associated PC IncRNAs (Adapted from Aird et al. [Citation149]). This schematic provides a summary on key lncRNAs and their role in up- and down-regulation of the AR. The role of these key lncRNAs in promoting and inhibiting carcinogenesis and the interplay with miRNAs and proteins is highlighted
![Figure 2. Androgen receptor (AR) signaling pathway associated PC IncRNAs (Adapted from Aird et al. [Citation149]). This schematic provides a summary on key lncRNAs and their role in up- and down-regulation of the AR. The role of these key lncRNAs in promoting and inhibiting carcinogenesis and the interplay with miRNAs and proteins is highlighted](/cms/asset/04f6a780-9ad2-4393-8a9c-a22aa26c5fc6/iero_a_1996227_f0002_oc.jpg)
7. LncRNA Interaction catalogs and prediction tools
Exploratory studies suggest that lncRNAs have an extensive interactome; however, current validation data are insufficient; therefore, many lncRNAs lack known interaction partners and their function remains elusive. lncRNA interactions may be acquired from public databases, some of which are considered below. However, existing annotations do not capture all the possible mechanisms and contexts in which lncRNAs may control gene expression. Experimental determination of lncRNA interactions is labor-intensive and costly, not least due to the large space of potential interactions, and so lncRNA function prediction software is required.
7.1. Databases related to lncRNAs
Presented by Zhou and colleagues, the experimentally validated functional lncRNA database (EVlncRNA) 2.0 is an updated version of the EVlncRNA database, which comprises only functional lncRNAs which have been experimentally validated through low-throughput techniques [Citation179]. The update, which involved the manual curation of over 19,000 lncRNA-associated publications, includes information on 4,010 curated lncRNAs across 124 species, 1,082 diseases, and 6,244 interactions. Importantly, the interaction network integrated into the tool’s online platform (https://www.sdklab-biophysics-dzu.net/EVLncRNAs2/) provides users with the ability to easily browse information retained within the database.
Despite the quality of lncRNA annotated within EVlncRNA 2.0, it could be considered much less comprehensive than alternative databases, such as RNAInter (RNA Interactome Database) or RISE (RNA Interactome from sequencing experiments) which rely heavily on data from high-throughput studies and thus possess substantially more lncRNA interactions [Citation180–182].
lncTarD database (http://bio-bigdata.hrbmu.edu.cn/LncTarD/) is a manually curated resource retaining information corresponding to 2822 lncRNA-target regulatory interactions that have proven relevance in 177 human diseases. For prostate cancer specifically, the database possesses data on 73 lncRNA-target interactions along with the mechanism of interaction and its functional consequences [Citation105]. LIVE (lncRNA Interaction Validated Encyclopedia) is a database of manually curated and experimentally validated lncRNA interactions [Citation183]. Consequently, LIVE is small with just 572 lncRNA interactions at the time of writing. Importantly, the database contains information relating to lncRNA interactions with multiple cellular components, including proteins and transcription factors, in addition to disease associations.
lnCaNet (LncRNA-Cancer gene co-expression Network) was first introduced in 2016 and is currently available as a web-based tool (http://lncanet.bioinfo-minzhao.org/) [Citation184]. It operates as a database containing lncRNA-gene co-expression predictions for 11 cancers (colon, lung, breast, etc.), with co-expression data sourced from Mitranscriptome, an online data repository of long polyadenylated RNA transcripts identified by RNA-seq [Citation185]. lnCaNet includes lncRNA-gene expression correlations for 2,544 cancer genes and 9,641 lncRNAs.
7.2. Prediction tools
In the absence of experimental evidence, predictive algorithms are used; however, benchmarking has revealed room for improved predictive performance [Citation186,Citation187]. Most RNA–RNA prediction algorithms must take into consideration the sequence of the RNA molecule when determining RNA prediction interactions. For example, both IntaRNA and LncRRIsearch methods utilize minimum free energy, a thermodynamic principle affected by the number, type, and arrangement of nucleotides of an RNA molecule, to predict interactions [Citation188–190]. In addition, the RIScoper tool is capable of extracting RNA–RNA interactions through literature text mining, which is highly valuable given that findings are likely to be experimentally validated [Citation191].
lncRNA–DNA interactions mostly occur through RNA–DNA triplex formation, and estimations of such structures are useful substitutes in lieu of experimental evidence. For example, the recently developed Triplex Forming Potential Prediction (TriplexFPP) is a machine learning-based model, which considers triplex formation capacity of a lncRNA based on computational methods in addition to triplex formation sites in DNA based on experimental data in a machine learning model to predict triplex formation [Citation191]. In theory, such a method would be able to outperform more traditional methods such as Triplexator as it is able to consider beyond the limits of the computational model through the inclusion of experimental data [Citation191,Citation192].
Capsule-LPI tool considers sequence, motif, physicochemical and secondary structural features from lncRNAs and proteins within a lncRNA-protein pair to determine lncRNA–protein interactions [Citation193]. Despite its complex mathematical architecture, the authors have compiled Capsule-LPI into a web-based tool (http://csbg-jlu.site/lpc/predict) allowing users to capture lncRNA–protein interactions by providing up to 100 lncRNA and protein sequences in Fasta format. Importantly, under 5-fold cross validation, Capture-LPI was found to possess the highest precision and the second highest sensitivity, highlighting the tool's utility and its room for improvement.
Interestingly, methods are available to translate interaction information into functional predictions. For example, MechRNA is a pipeline for lncRNA functional prediction based on RNA–RNA and RNA–protein interaction data. This approach utilizes the IntaRNA2 algorithm to predict RNA–RNA interactions along with publicly available CLIP-seq (cross-linking immunoprecipitation sequencing) data and the GraphProt computational framework to determine RNA–protein interactions before combining data and predicting function based on several predetermined possibilities [Citation194,Citation195].
Possibly, the most well-known method for lncRNA regulatory predictions is the LongHorn algorithm, which derives lncRNA interactions based on four predefined models of lncRNA regulation.
These models include activities such as decoys, miRNA sponges, cofactors/guides, and switches [Citation30]. In a seminal publication, Chiu et al. applied the algorithm to TCGA data and predicted lncRNA–gene interaction predictions for individual cancers of interest. The lncRNA-gene predictions are available presently for 13 tumor contexts including prostate adenocarcinoma, lung adenocarcinoma, and ovarian serous cystadenocarcinoma. For PC, Longhorn was applied to TCGA data including RNA, miRNA expression and copy number profiles of 371 prostate adenocarcinoma tumors. PLAIDOH (Predicting lncRNA Activity through Integrative Data-driven Omics and Heuristics) was applied to identify lncRNAs associated with non-Hodgkin lymphoma [Citation196]. PLAIDOH offers users the opportunity to analyze both publicly and privately owned datasets, analyze statically small datasets, and ultimately rank lncRNAs in order of importance based on customizable metrics [Citation196]. One limitation is that it only predicts lncRNA targets that are located within a genomic window limit of 800 kb. Using PLAIDOH, researchers have validated predictions by identifying well-recognized lncRNA gene regulatory pairs, such as HOTAIR matching with HOX genes and familiar lncRNA-protein binding pairs such as NEAT1 and NONO [Citation196], new lncRNA interactions have also been uncovered and subsequently validated, demonstrating the utility of this tool.
lncRNA-screen is an interactive platform that operates as a fully functional analysis pipeline which performs all major steps involved with typical RNA-sequencing processing, and its main objective is to facilitate the computational discovery of lncRNA candidates that can be further examined by functional experiments [Citation181]. This pipeline can automatically download and prepare raw FASTQ (sequencing) files from multiple public repositories and performs data quality control, pre-processing, alignment, annotation, differential expression, and novel lncRNA identification and classification. After analysis, lncRNA-screen provides users with an interactive HTML summary report that can be searched and filtered to suit users’ individual needs. Information includes locus, number of exons, a genome snapshot and lncRNA–mRNA interactions. lncRNA-screen is limited to lncRNA-gene predictions by physical location (neighboring genes).
These databases and computational tools may be applied to understand the regulatory impact of lncRNAs upon oncogenes and tumor suppressors [Citation197,Citation198]. The resultant predictions are useful for the identification of candidate cancer biomarkers and therapeutic targets, in concert with traditional biochemical methods [Citation199].
8. Concluding remarks
lncRNAs are now established regulators of a vast number of biological processes as well key mediators for the development and progression of various cancers. This is remarkable, considering that lncRNAs were once branded as ‘junk’ with no regulatory capabilities. Despite significant progress, there is still much to learn about lncRNAs and their potential value as cancer biomarkers or in therapy.
9. Expert opinion
At present, there are three areas of prostate cancer research that require focus: 1) understanding fundamental biological reasons as to why men of African descent are significantly more prone to disease morbidity and mortality, 2) improved therapeutic treatments for advanced and castrate-resistant PC, and 3) the necessity for biomarkers, which can differentiate low-risk and high-risk diseases and assess PC racial disparities. Elucidating the functional roles of lncRNAs will help drive advances in each of these areas and will lead to better diagnosis and treatment of PC.
Despite these advances, opinions in the scientific community as to whether lncRNAs provide realistic therapeutic, prognostic, or diagnostic targets are varied at this time. This will add to the complexity of implementing clinical testing based on lncRNA diagnostics. The Clinical Laboratory Improvement Amendments (CLIA) federal regulatory standards require that clinical laboratories determine and detail their own performance specifications for laboratory-developed tests to guarantee accurate and precise results prior to implementation of the test. Characteristics that must be established typically include accuracy and precision, range of reporting, analytical sensitivity and specificity, and reference intervals. Although both a positive and negative attribute, the sheer number of lncRNAs typically predicted by computational tools presents challenges for both clinical and research practice. The hundreds to thousands of lncRNAs that may be identified computationally cannot be feasibly validated in a clinical setting. This elevates the need for more sophisticated computational algorithms based on deep learning and other approaches, which not only accurately predict lncRNAs but also facilitate stringent thresholds and intelligent approaches to triage-specific lncRNAs subsets for laboratory-based testing and ultimately clinical implementation.
As precision medicine approaches evolve and become more inclusive, an important impact will be the advancement of knowledge of racial differences among prostate and other cancers. Several studies have determined that lncRNAs may play a role in differences in racial outcomes in cancer. There remain considerable and persistent racial disparities in prostate cancer outcomes. Reduced access to health-care services contributes to racial disparities in PC outcomes, but even in equal access health-care systems, such as the Veterans Administration (VA) in the USA, African American Veterans have higher PSA levels and higher-grade tumors compared to European American Veterans even when presenting with the same stage of disease. Unfortunately, medical mistrust issues and their associated indicators among African American patients have an adverse effect in recruiting sufficient sample numbers of ethnic minorities and their participation in clinical trials. Distrust in the health-care system and fear associated with detection and treatment are responsible for late diagnosis in African American men. There is a paucity of samples from racial minorities in public genomic databases to detect moderately common genomic alterations in African Americans, which is contributing to the gap in PC disparities. We anticipate that as the lncRNA research field develops and matures, greater emphasis will be placed on identifying lncRNAs that play key roles in the development and progression of cancer across all ethnic groups.
This field has already significantly progressed in a relatively short period of time, from the first lncRNA identified (h19) to the realization that lncRNAs do in fact process regulatory capabilities. With novel lncRNAs being identified daily and associated with cancer, the pressure to experimentally validate these emerging candidates is increasing. In the future, we expect lncRNA computational tools to become an essential tool for both research and clinical laboratories and for a variety of diseases beyond cancer. We also expect significant growth in curated lncRNA databases with improved functional predictions of lncRNAs. We anticipate increased commercial activities based around lncRNA therapeutics, prognostics, and diagnostics.
Article highlights
• Racial differences continue to persist in prostate cancer
• lncRNAs possess regulatory functions and can become diagnostic, prognostic, and therapeutic targets
• The vast majority of lncRNAs are yet to be experimentally validated and their molecular function elucidated
• lncRNA candidate biomarkers require analytical and clinical validation to better assess their utility
• There is a clear need for more sophisticated computational algorithms which not only accurately predict lncRNAs but also facilitate stringent thresholds and intelligent approaches to triage specific lncRNAs subsets for laboratory-based testing and ultimately clinical implementation.
Declaration of Interest
IO has undertaken a consultancy for Mevox Ltd. The other authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending, or royalties.
Additional information
Funding
References
- Lee DJ, Mallin K, Graves AJ, et al. Recent changes in prostate cancer screening practices and epidemiology. J Urol. 2017;198(6):1230–1240.
- Wallace TA, et al. Tumor immunobiological differences in prostate cancer between African-American and European-American men. Cancer Res. 2008;68(3):927–936.
- Kung JT, Colognori D, Lee JT. Long noncoding RNAs: past, present, and future. Genetics. 2013;193(3):651–669.
- Zhang J, et al. LncmiRSRN: identification and analysis of long non-coding RNA related miRNA sponge regulatory network in human cancer. Bioinformatics. 2018;34(24):4232–4240.
- Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641.
- Salviano-Silva A, et al. Besides pathology: long non-coding RNA in cell and tissue homeostasis. Noncoding RNA. 2018;4:1.
- Dong P, et al. Long Non-coding RNA NEAT1: a novel target for diagnosis and therapy in human tumors. Front Genet. 2018;9(p):471.
- Hobuss L, Bar C, Thum T. Long Non-coding RNAs: at the heart of cardiac dysfunction? Front Physiol. 2019;10(p):30.
- Sanchez Calle A, et al. Emerging roles of long non-coding RNA in cancer. Cancer Sci. 2018;109(7):2093–2100.
- Jiang Q, Ma R, Wang J, et al. LncRNA2Function: a comprehensive resource for functional investigation of human lncRNAs based on RNA-seq data. BMC Genomics. 2015;16(Suppl 3):S2.
- Choi SW, Kim HW, Nam JW. The small peptide world in long noncoding RNAs. Brief Bioinform. 2019;20(5):1853–1864.
- Walsh AL, et al. Long noncoding RNAs and prostate carcinogenesis: the missing ‘linc’? Trends Mol Med. 2014;20(8):428–436.
- Rao MRS. Long non coding biology. Springer; 2017.
- Ma L, et al. The long noncoding RNA H19 promotes cell proliferation via E2F-1 in pancreatic ductal adenocarcinoma. Cancer Biol Ther. 2016;17(10):1051–1061.
- Hao Y, et al. Tumour-suppressor activity of H19 RNA. Nature. 1993;365(6448):764–767.
- Brannan CI, et al. The product of the H19 gene may function as an RNA. Mol Cell Biol. 1990;10(1):28–36.
- Okazaki Y, et al. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature. 2002;420(6915):563–73.
- Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function. Nat Rev Genet. 2016;17(1):47–62.
- Lam YWF, Cavallari LH. Pharmacogenomics: challenges and opportunities in therapeutic implementation. Academic Press; 2013.
- Chen LL. Linking long noncoding RNA localization and function. Trends Biochem Sci. 2016;41(9):761–772.
- Ma L, et al. LncBook: a curated knowledgebase of human long non-coding RNAs. Nucleic Acids Res. 2019;47(5):2699.
- Sun T, et al. Emerging players in prostate cancer: long non-coding RNAs. Am J Clin Exp Urol. 2014;2(4):294–9.
- Ma L, Bajic VB, Zhang Z. On the classification of long non-coding RNAs. RNA Biol. 2013;10(6):925–933.
- Zhang Q, et al. The complexity of bladder cancer: long noncoding RNAs are on the stage. Mol Cancer. 2013;12(1):101.
- Biosynthesis. What are lncRNAs and lincRNAs. 2013; [cited 2021 Nov 1]. Available from:https//www.biosyn.com/tew/what-are-lncrnas-and-lincrnas.aspx.
- Wu T, Du Y. LncRNAs: from basic research to medical application. Int J Biol Sci. 2017;13(3):295–307.
- Mouraviev V, et al. Clinical prospects of long noncoding RNAs as novel biomarkers and therapeutic targets in prostate cancer. Prostate Cancer Prostatic Dis. 2016;19(1):14–20.
- RNA Central -Homo sapiens (human) HOX transcript antisense RNA. [cited 2021 Nov 1] . Available from: https://rnacentral.org/rna/URS000075C808/9606.
- Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–914.
- Chiu HS, et al. Pan-cancer analysis of lncRNA regulation supports their targeting of cancer genes in each tumor context. Cell Rep. 2018;23(1):297–312. e12.
- Yu X, et al. NEAT1: a novel cancer-related long non-coding RNA. Cell Prolif. 2017;50:2.
- Klec C, Prinz F, Pichler M. Involvement of the long noncoding RNA NEAT1 in carcinogenesis. Mol Oncol. 2019;13(1):46–60.
- An J, Lv W, Zhang Y. LncRNA NEAT1 contributes to paclitaxel resistance of ovarian cancer cells by regulating ZEB1 expression via miR-194. Onco Targets Ther. 2017;10(p):5377–5390.
- Shin VY, et al. Long non-coding RNA NEAT1 confers oncogenic role in triple-negative breast cancer through modulating chemoresistance and cancer stemness. Cell Death Dis. 2019;10(4):270.
- Bhat SA, et al. Long non-coding RNAs: mechanism of action and functional utility. Noncoding RNA Res. 2016;1(1):43–50.
- Blewitt M. Noncoding RNAs - long noncoding RNAs introduction. In: Blewitt M, editor. Epigenetic control of gene expression. Melbourne Australia: Coursera.
- Morrison AS. Epidemiology and environmental factors in urologic cancer. Cancer. 1987;60(3 Suppl):632–4.
- Fujita K. Urinary biomarkers of urological malignancies. Transl Androl Urol. 2021;10(4):1827–1828.
- Wu P, Cao Z, Wu S. New progress of epigenetic biomarkers in urological cancer. Dis Markers. 2016;2016:9864047.
- Filella X, et al. Emerging biomarkers in the diagnosis of prostate cancer. Pharmgenomics Pers Med. 2018;11(p):83–94.
- Lee HH, Kim SH. Review of non-invasive urinary biomarkers in bladder cancer. Transl Cancer Res. 2020;9(10):6554–6564.
- Jiang G, et al. Long non-coding RNAs in prostate tumorigenesis and therapy (review). Mol Clin Oncol. 2020;13(6):76.
- Amodio N, et al. MALAT1: a druggable long non-coding RNA for targeted anti-cancer approaches. J Hematol Oncol. 2018;11(1):63.
- Sun Y, Ma L. New insights into long non-coding RNA MALAT1 in cancer and metastasis. Cancers (Basel). 2019;11(2). DOI:https://doi.org/10.3390/cancers11020216
- Fan Y, et al. TGF-beta-induced upregulation of malat1 promotes bladder cancer metastasis by associating with suz12. Clin Cancer Res. 2014;20(6):1531–1541.
- Nitusca D, et al. Long noncoding RNA NEAT1 as a potential candidate biomarker for prostate cancer. Life (Basel). 2021;11(4).
- Ye T, et al. Long noncoding RNA linc00346 promotes the malignant phenotypes of bladder cancer. Biochem Biophys Res Commun. 2017;491(1):79–84.
- Pandey RR, et al. Kcnq1ot1 antisense noncoding RNA mediates lineage-specific transcriptional silencing through chromatin-level regulation. Mol Cell. 2008;32(2):232–246.
- Mohammad F, et al. Long noncoding RNA-mediated maintenance of DNA methylation and transcriptional gene silencing. Development. 2012;139(15):2792–2803.
- Li Y, et al. LncRNA KCNQ1OT1 facilitates the progression of bladder cancer by targeting MiR-218-5p/HS3ST3B1. Cancer Gene Ther. 2021;28(3–4):212–220.
- Wang J, et al. Overexpression of long non-coding RNA LOC400891 promotes tumor progression and poor prognosis in prostate cancer. Tumour Biol. 2016;37(7):9603–9613.
- Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981.
- Gao N, et al. Long non-coding RNAs: the regulatory mechanisms, research strategies, and future directions in cancers. Front Oncol. 2020;10(p):598817.
- Maass PG, Luft FC, Bahring S. Long non-coding RNA in health and disease. J Mol Med (Berl). 2014;92(4):337–346.
- Xu T, et al. Pathological bases and clinical impact of long noncoding RNAs in prostate cancer: a new budding star. Mol Cancer. 2018;17(1):103.
- Al Aameri RFH, et al. Tonic suppression of PCAT29 by the IL-6 signaling pathway in prostate cancer: reversal by resveratrol. PLoS One. 2017;12(5):e0177198.
- Sakurai K, et al. The lncRNA DRAIC/PCAT29 locus constitutes a tumor-suppressive nexus. Mol Cancer Res. 2015;13(5):828–838.
- Frank F, et al. The lncRNA growth arrest specific 5 regulates cell survival via distinct structural modules with independent functions. Cell Rep. 2020;32(3):107933.
- Pickard MR, Mourtada-Maarabouni M, Williams GT. Long non-coding RNA GAS5 regulates apoptosis in prostate cancer cell lines. Biochim Biophys Acta. 2013;1832(10):1613–1623.
- Bray F, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
- Schatten H. Cell & molecular biology of prostate cancer. Vol. 1095. Cham, Switzerland: Springer; 2018.
- Culp MB, et al. Recent global patterns in prostate cancer incidence and mortality rates. Eur Urol. 2020;77(1):38–52.
- Dovey ZS, Nair, SS, and Chakravarty, D, et al. Racial disparity in prostate cancer in the African American population with actionable ideas and novel immunotherapies. Cancer Rep (Hoboken). 2021;e1341.
- Yuan J, et al. Integrative comparison of the genomic and transcriptomic landscape between prostate cancer patients of predominantly African or European genetic ancestry. PLoS Genet. 2020;16(2):e1008641.
- Jiang MC, et al. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am J Cancer Res. 2019;9(7):1354–1366.
- Mendell JT. Targeting a long noncoding RNA in breast cancer. N Engl J Med. 2016;374(23):2287–2289.
- He Q, et al. Emerging roles of lncRNAs in the formation and progression of colorectal cancer. Front Oncol. 2019;9(p):1542.
- Arun G, Diermeier SD, Spector DL. Therapeutic targeting of long non-coding RNAs in cancer. Trends Mol Med. 2018;24(3):257–277.
- Carlevaro-Fita J, et al. Cancer LncRNA census reveals evidence for deep functional conservation of long noncoding RNAs in tumorigenesis. Commun Biol. 2020;3(1):56.
- Zheng HT, et al. High expression of lncRNA MALAT1 suggests a biomarker of poor prognosis in colorectal cancer. Int J Clin Exp Pathol. 2014;7(6):3174–3181.
- Zhu K, Ren Q, Zhao Y. lncRNA MALAT1 overexpression promotes proliferation, migration and invasion of gastric cancer by activating the PI3K/AKT pathway. Oncol Lett. 2019;17(6):5335–5342.
- Kim J, et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat Genet. 2018;50(12):1705–1715.
- Liu Y, et al. Long non-coding RNA NEAT1 promoted ovarian cancer cells’ metastasis through regulation of miR-382-3p/ROCK1 axial. Cancer Sci. 2018;109(7):2188–2198.
- Luo Y, et al. Long non-coding RNA NEAT1 promotes colorectal cancer progression by competitively binding miR-34a with SIRT1 and enhancing the Wnt/beta-catenin signaling pathway. Cancer Lett. 2019;440-441(p):11–22.
- Gupta RA, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076.
- Niinuma T, et al. Upregulation of miR-196a and HOTAIR drive malignant character in gastrointestinal stromal tumors. Cancer Res. 2012;72(5):1126–1136.
- Kogo R, et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011;71(20):6320–6326.
- Yang Z, et al. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann Surg Oncol. 2011;18(5):1243–1250.
- Barr JA, et al. Long non-coding RNA FAM83H-AS1 is regulated by human papillomavirus 16 E6 independently of p53 in cervical cancer cells. Sci Rep. 2019;9(1):3662.
- Xia W, et al. Down-regulated lncRNA SBF2-AS1 inhibits tumorigenesis and progression of breast cancer by sponging microRNA-143 and repressing RRS1. J Exp Clin Cancer Res. 2020;39(1):18.
- Niu L, et al. LncRNA NRON down-regulates lncRNA snaR and inhibits cancer cell proliferation in TNBC. Biosci Rep. 2019;39(5). DOI:https://doi.org/10.1042/BSR20190468
- Lei T, et al. LncRNA NBAT-1 is down-regulated in lung cancer and influences cell proliferation, apoptosis and cell cycle. Eur Rev Med Pharmacol Sci. 2018;22(7):1958–1962.
- Lu X, et al. Downregulation of gas5 increases pancreatic cancer cell proliferation by regulating CDK6. Cell Tissue Res. 2013;354(3):891–896.
- Malik B, Feng FY. Long noncoding RNAs in prostate cancer: overview and clinical implications. Asian J Androl. 2016;18(4):568–574.
- Rebbeck TR. Prostate cancer genetics: variation by race, ethnicity, and geography. Semin Radiat Oncol. 2017;27(1):3–10.
- Das R, Feng FY, Selth LA. Long non-coding RNAs in prostate cancer: biological and clinical implications. Mol Cell Endocrinol. 2019;480(p):142–152.
- Arriaga-Canon C, et al. The use of long non-coding RNAs as prognostic biomarkers and therapeutic targets in prostate cancer. Oncotarget. 2018;9(29):20872–20890.
- Marrugo-Ramirez J, Mir M, Samitier J. Blood-based cancer biomarkers in liquid biopsy: a promising non-invasive alternative to tissue biopsy. Int J Mol Sci. 2018;19:10.
- Helsmoortel H, et al. Detecting long non-coding RNA biomarkers in prostate cancer liquid biopsies: hype or hope? Noncoding RNA Res. 2018;3(2):64–74.
- Etzioni R, et al. Overdiagnosis due to prostate-specific antigen screening: lessons from U.S. prostate cancer incidence trends. J Natl Cancer Inst. 2002;94(13):981–990.
- Wilt TJ, et al. Prostate-specific antigen screening in prostate cancer: perspectives on the evidence. J Natl Cancer Inst. 2014;106(3):dju010.
- O’Sullivan J. Controversies in PSA screening. Evid Based Med. 2017;22(6):198.
- Brawley OW. Trends in prostate cancer in the United States. J Natl Cancer Inst Monogr. 2012;2012(45):152–156.
- Feibus AH, et al. Clinical use of PCA3 and TMPRSS2:ERG urinary biomarkers in African-American men undergoing prostate biopsy. J Urol. 2016;196(4):1053–60.
- Bussemakers MJ, et al. DD3: a new prostate-specific gene, highly overexpressed in prostate cancer. Cancer Res. 1999;59(23):5975–9.
- Crawford ED, et al. Diagnostic performance of PCA3 to detect prostate cancer in men with increased prostate specific antigen: a prospective study of 1,962 cases. J Urol. 2012;188(5):1726–31.
- Yao Y, Ranade K, Jannal B. Genomic biomarkers for pharmaceutical development. Academic Press; 2013.
- Gray SG. Epigenetic cancer therapy. Academic Press; 2015.
- Misawa A, Takayama KI, Inoue S. Long non-coding RNAs and prostate cancer. Cancer Sci. 2017;108(11):2107–2114.
- Dijkstra S, et al. Personalized management in low-risk prostate cancer: the role of biomarkers. Prostate Cancer. 2012;2012:327104.
- Mitobe Y, et al. Prostate cancer-associated lncRNAs. Cancer Lett. 2018;418(p):159–166.
- O’Malley PG, et al. Racial variation in the utility of urinary biomarkers PCA3 and T2ERG in a large multicenter study. J Urol. 2017;198(1):42–49.
- Adam A, Engelbrecht MJ, Bornman MS, et al. The role of the PCA3 assay in predicting prostate biopsy outcome in a South African setting. BJU International. 2011;108(11):1728–1733.
- Prensner JR, et al. PCAT-1, a long noncoding RNA, regulates BRCA2 and controls homologous recombination in cancer. Cancer Res. 2014;74(6):1651–1660.
- Zhao X, et al. PCAT1 is a poor prognostic factor in endometrial carcinoma and associated with cancer cell proliferation, migration and invasion. Bosn J Basic Med Sci. 2019;19(3):274–281.
- Prensner JR, et al. Transcriptome sequencing across a prostate cancer cohort identifies PCAT-1, an unannotated lincRNA implicated in disease progression. Nat Biotechnol. 2011;29(8):742–9.
- Matejcic M, et al. Germline variation at 8q24 and prostate cancer risk in men of European ancestry. Nat Commun. 2018;9(1):4616.
- Xiong T, et al. PCAT-1: a novel oncogenic long non-coding RNA in human cancers. Int J Biol Sci. 2019;15(4):847–856.
- Yuan Q, et al. LncRNA PCAT1 and its genetic variant rs1902432 are associated with prostate cancer risk. J Cancer. 2018;9(8):1414–1420.
- Crea F, et al. Identification of a long non-coding RNA as a novel biomarker and potential therapeutic target for metastatic prostate cancer. Oncotarget. 2014;5(3):764–774.
- Ren S, et al. Long non-coding RNA metastasis associated in lung adenocarcinoma transcript 1 derived miniRNA as a novel plasma-based biomarker for diagnosing prostate cancer. Eur J Cancer. 2013;49(13):2949–2959.
- Wang F, et al. Development and prospective multicenter evaluation of the long noncoding RNA MALAT-1 as a diagnostic urinary biomarker for prostate cancer. Oncotarget. 2014;5(22):11091–11102.
- Liu Y, et al. Long non-coding RNA PCAT7 regulates ELF2 signaling through inhibition of miR-134-5p in nasopharyngeal carcinoma. Biochem Biophys Res Commun. 2017;491(2):374–381.
- Liu Q, et al. Long non-coding RNA prostate cancer-associated transcript 7 (PCAT7) induces poor prognosis and promotes tumorigenesis by inhibiting mir-134-5p in non-small-cell lung (NSCLC). Med Sci Monit. 2017;23(p):6089–6098.
- Lang C, et al. SMAD3/SP1 complex-mediated constitutive active loop between lncRNA PCAT7 and TGF-beta signaling promotes prostate cancer bone metastasis. Mol Oncol. 2020;14(4):808–828.
- Cimadamore A, et al. Long non-coding RNAs in prostate cancer with emphasis on second chromosome locus associated with prostate-1 expression. Front Oncol. 2017;7(p):305.
- Parolia A, et al. The long non-coding RNA PCGEM1 is regulated by androgen receptor activity in vivo. Mol Cancer. 2015;14(p):46.
- Mehra R, et al. A novel RNA in situ hybridization assay for the long noncoding RNA SChLAP1 predicts poor clinical outcome after radical prostatectomy in clinically localized prostate cancer. Neoplasia. 2014;16(12):1121–1127.
- Prensner JR, et al. The long noncoding RNA SChLAP1 promotes aggressive prostate cancer and antagonizes the SWI/SNF complex. Nat Genet. 2013;45(11):1392–1398.
- Prensner JR, et al. RNA biomarkers associated with metastatic progression in prostate cancer: a multi-institutional high-throughput analysis of SChLAP1. Lancet Oncol. 2014;15(13):1469–1480.
- Chua MLK, et al. A prostate cancer “nimbosus”: genomic instability and SChLAP1 dysregulation underpin aggression of intraductal and cribriform subpathologies. Eur Urol. 2017;72(5):665–674.
- Petrovics G, et al. Elevated expression of PCGEM1, a prostate-specific gene with cell growth-promoting function, is associated with high-risk prostate cancer patients. Oncogene. 2004;23(2):605–611.
- Fu X, et al. Regulation of apoptosis by a prostate-specific and prostate cancer-associated noncoding gene, PCGEM1. DNA Cell Biol. 2006;25(3):135–141.
- Lee JEA. Increased expression of PCGEM1 lncRNA in prostate cancer of African American men.2016; 6(6).
- Waxman AJ, et al. Racial disparities in incidence and outcome in multiple myeloma: a population-based study. Blood. 2010;116(25):5501–5506.
- Smith CJ, Ambs S, Landgren O. Biological determinants of health disparities in multiple myeloma. Blood Cancer J. 2018;8(9):85.
- Cui YS, Song YP, Fang BJ. The role of long non-coding RNAs in multiple myeloma. Eur J Haematol. 2019;103(1):3–9.
- Kazandjian D. Multiple myeloma epidemiology and survival: a unique malignancy. Semin Oncol. 2016;43(6):676–681.
- Wallace K, et al. Platelet and hemoglobin count at diagnosis are associated with survival in African American and Caucasian patients with colorectal cancer. Cancer Epidemiol. 2020;67(p):101746.
- Chatterjee NA, He Y, Keating NL. Racial differences in breast cancer stage at diagnosis in the mammography era. Am J Public Health. 2013;103(1):170–176.
- Gupta V, Haque I, Chakraborty J, et al. Racial disparity in breast cancer: can it be mattered for prognosis and therapy. J Cell Commun Signal. 2018;12(1):119–132.
- DeSantis CE, Miller KD, Goding Sauer A, et al. Cancer statistics for African Americans, 2019. CA Cancer J Clin. 2019;69(3):211–233.
- Rebbeck TR, et al. Global patterns of prostate cancer incidence, aggressiveness, and mortality in men of African descent. Prostate Cancer. 2013;2013(p):560857.
- Jones AL, Chinegwundoh F. Update on prostate cancer in black men within the UK. Ecancermedicalscience. 2014;8:455.
- Hardiman G, et al. Systems analysis of the prostate transcriptome in African-American men compared with European-American men. Pharmacogenomics. 2016;17(10):1129–1143.
- Richards Z, et al. Prostatic compensation of the vitamin D axis in African American men. JCI Insight. 2017;2(2):91054.
- Benafif S, et al. A review of prostate cancer genome-wide association studies (GWAS). Cancer Epidemiol Biomarkers Prev. 2018;27(8):845–857.
- Barrington WE, et al. Difference in association of obesity with prostate cancer risk between US African American and non-Hispanic white men in the selenium and vitamin E cancer prevention trial (SELECT). JAMA Oncol. 2015;1(3):342–349.
- Xiao J, et al. Mitochondrial biology and prostate cancer ethnic disparity. Carcinogenesis. 2018;39(11):1311–1319.
- Freedman ML, et al. Admixture mapping identifies 8q24 as a prostate cancer risk locus in African-American men. Proc Natl Acad Sci U S A. 2006;103(38):14068–14073.
- Robbins CM, et al. EphB2 SNPs and sporadic prostate cancer risk in African American men. PLoS One. 2011;6(5):e19494.
- Rawla P. Epidemiology of Prostate Cancer. World J Oncol. 2019;10(2):63–89.
- Bentley AR, Callier SL, Rotimi CN. Evaluating the promise of inclusion of African ancestry populations in genomics. NPJ Genom Med. 2020;5:5.
- Hardiman G. An Introduction to systems analytics and integration of big omics data. Genes (Basel). 2020;11(3):245.
- Frey LJ. Artificial intelligence and integrated genotype–phenotype identification. Genes (Basel). 2019;10(1):18.
- Koutrouli M, et al. A guide to conquer the biological network era using graph theory. Front Bioeng Biotechnol. 2020;8(34). DOI:https://doi.org/10.3389/fbioe.2020.00034
- Pavlopoulos GA, et al. Using graph theory to analyze biological networks. BioData Min. 2011;4(1):10.
- Moul JW. The evolving definition of advanced prostate cancer. Rev Urol. 2004;6(Suppl 8):S10–7.
- Obenauf AC, Massague J. Surviving at a distance: organ-Specific metastasis. Trends Cancer. 2015;1(1):76–91.
- Weidle UH, et al. The functional role of prostate cancer metastasis-related micro-RNAs. Cancer Genomics Proteomics. 2019;16(1):1–19.
- Society AC. Hormone Therapy for Prostate Cancer [cited 2021 Nov 1] . 2019; Available from: https://www.cancer.org/cancer/prostate-cancer/treating/hormone-therapy.html.
- Service NH. Treatment Prostate cancer [cited 2021 Nov 1] . 2018; Available from: https://www.nhs.uk/conditions/prostate-cancer/treatment/.
- Wadosky KM, Koochekpour S. Molecular mechanisms underlying resistance to androgen deprivation therapy in prostate cancer. Oncotarget. 2016;7(39):64447–64470.
- Crawford ED, et al. Androgen-targeted therapy in men with prostate cancer: evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019;22(1):24–38.
- Dong L, et al. Metastatic prostate cancer remains incurable, why? Asian J Urol. 2019;6(1):26–41.
- Karantanos T, Corn PG, Thompson TC. Prostate cancer progression after androgen deprivation therapy: mechanisms of castrate resistance and novel therapeutic approaches. Oncogene. 2013;32(49):5501–5511.
- Hotte SJ, Saad F. Current management of castrate-resistant prostate cancer. Curr Oncol. 2010;17(Suppl 2):S72–9.
- Moreira DM, et al. Predictors of time to metastasis in castration-resistant prostate cancer. Urology. 2016;96(p):171–176.
- Saad F, Hotte SJ. Guidelines for the management of castrate-resistant prostate cancer. Can Urol Assoc J. 2010;4(6):380–384.
- Mehtala J, et al. Overall survival and second primary malignancies in men with metastatic prostate cancer. PLoS One. 2020;15(2):e0227552.
- Damodaran S, Kyriakopoulos CE, Jarrard DF. Newly Diagnosed Metastatic Prostate Cancer: has the Paradigm Changed? Urol Clin North Am. 2017;44(4):611–621.
- Barth DA, et al. lncRNA and mechanisms of drug resistance in cancers of the genitourinary system. Cancers (Basel. 2020;12(8). DOI:https://doi.org/10.3390/cancers12082148.
- Housman G, et al. Drug resistance in cancer: an overview. Cancers (Basel). 2014;6(3):1769–1792.
- Aird J, et al. Carcinogenesis in prostate cancer: the role of long non-coding RNAs. Noncoding RNA Res. 2018;3(1):29–38.
- Shih JW, et al. Non-coding RNAs in castration-resistant prostate cancer: regulation of androgen receptor signaling and cancer metabolism. Int J Mol Sci. 2015;16(12):28943–28978.
- Malik R, et al. The lncRNA PCAT29 inhibits oncogenic phenotypes in prostate cancer. Mol Cancer Res. 2014;12(8):1081–1087.
- Bajpai P, et al. Mitochondrial localization, import, and mitochondrial function of the androgen receptor. J Biol Chem. 2019;294(16):6621–6634.
- Altschuler J, Stockert JA, and Kyprianou N. Non-coding RNAs set a new phenotypic frontier in prostate cancer metastasis and resistance. Int J Mol Sci. 2021;22(4). DOI:https://doi.org/10.3390/ijms22042100
- Parolia A, et al. The long noncoding RNA HORAS5 mediates castration-resistant prostate cancer survival by activating the androgen receptor transcriptional program. Mol Oncol. 2019;13(5):1121–1136.
- Takayama K, et al. Androgen-responsive long noncoding RNA CTBP1-AS promotes prostate cancer. EMBO J. 2013;32(12):1665–1680.
- Scaravilli M, Koivukoski S, Latonen L. Androgen-driven fusion genes and chimeric transcripts in prostate cancer. Front Cell Dev Biol. 2021;9:623809.
- Yang Y, et al. Androgen receptor-related non-coding RNAs in prostate cancer. Front Cell Dev Biol. 2021;9(p):660853.
- Wang R, et al. Role of transcriptional corepressor CtBP1 in prostate cancer progression. Neoplasia. 2012;14(10):905–914.
- Rinn JL, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323.
- Carrion K, et al. The long non-coding HOTAIR is modulated by cyclic stretch and WNT/β-CATENIN in human aortic valve cells and is a novel repressor of calcification genes. PLoS One. 2014;9(5):e96577.
- Zhang A, et al. LncRNA HOTAIR enhances the androgen-receptor-mediated transcriptional program and drives castration-resistant prostate cancer. Cell Rep. 2015;13(1):209–221.
- Mather RL, Wang Y, Crea F. Is HOTAIR really involved in neuroendocrine prostate cancer differentiation? Epigenomics. 2018;10(10):1259–1261.
- Zhang Y, et al. Analysis of the androgen receptor-regulated lncRNA landscape identifies a role for ARLNC1 in prostate cancer progression. Nat Genet. 2018;50(6):814–824.
- Zhou B, et al. EVLncRNAs 2.0: an updated database of manually curated functional long non-coding RNAs validated by low-throughput experiments. Nucleic Acids Res. 2021;49(D1):D86–D91.
- Gong J, et al. RISE: a database of RNA interactome from sequencing experiments. Nucleic Acids Res. 2018;46(D1):D194–D201.
- Gong Y, et al. lncRNA-screen: an interactive platform for computationally screening long non-coding RNAs in large genomics datasets. BMC Genomics. 2017;18(1):434.
- Lin Y, et al. RNAInter in 2020: RNA interactome repository with increased coverage and annotation. Nucleic Acids Res. 2020;48(D1):D189–D197.
- An G, Sun, J, and Ren, C, et al. LIVE: a manually curated encyclopedia of experimentally validated interactions of lncRNAs. Database (Oxford). 2019;2019.
- Liu Y, Zhao M. lnCaNet: pan-cancer co-expression network for human lncRNA and cancer genes. Bioinformatics. 2016;32(10):1595–1597.
- Iyer MK, et al. The landscape of long noncoding RNAs in the human transcriptome. Nat Genet. 2015;47(3):199–208.
- Umu SU, Gardner PP. A comprehensive benchmark of RNA-RNA interaction prediction tools for all domains of life. Bioinformatics. 2017;33(7):988–996.
- Lai D, Meyer IM. A comprehensive comparison of general RNA-RNA interaction prediction methods. Nucleic Acids Res. 2016;44(7):e61.
- Pinkney HR, Wright BM, Diermeier SD. The lncrna toolkit: databases and in silico tools for lncrna analysis. Noncoding RNA. 2020;6(4):49.
- Fukunaga T, et al. LncRRIsearch: a web server for lncRNA-RNA interaction prediction integrated with tissue-specific expression and subcellular localization data. Front Genet. 2019;10(p):462.
- Mann M, Wright PR, Backofen R. IntaRNA 2.0: enhanced and customizable prediction of RNA–RNA interactions. Nucleic Acids Res. 2017;45(W1):W435–W439.
- Zhang Y, Long Y, Kwoh CK. Deep learning based DNA: RNA triplex forming potential prediction. BMC Bioinformatics. 2020;21(1):1–13.
- Buske FA, et al. Triplexator: detecting nucleic acid triple helices in genomic and transcriptomic data. Genome Res. 2012;22(7):1372–1381.
- Li Y, et al. Capsule-LPI: a LncRNA–protein interaction predicting tool based on a capsule network. BMC Bioinformatics. 2021;22(1):1–19.
- Maticzka D, et al. GraphProt: modeling binding preferences of RNA-binding proteins. Genome Biol. 2014;15(1):1–18.
- Gawronski AR, et al. MechRNA: prediction of lncRNA mechanisms from RNA-RNA and RNA-protein interactions. Bioinformatics. 2018;34(18):3101–3110.
- Pyfrom SC, Luo H, Payton JE. PLAIDOH: a novel method for functional prediction of long non-coding RNAs identifies cancer-specific LncRNA activities. BMC Genomics. 2019;20(1):137.
- Ji H, et al. Identification, functional prediction, and key lncRNA verification of cold stress-related lncRNAs in rats liver. Sci Rep. 2020;10(1):521.
- da Silveira WA, et al. miRmapper: a tool for interpretation of miRNA⁻mRNA interaction networks. Genes (Basel). 2018;9(9). DOI:https://doi.org/10.3390/genes9090458
- Beerenwinkel N, Greenman CD, Lagergren J. Computational cancer biology: an evolutionary perspective. PLoS Comput Biol. 2016;12(2):e1004717.