
Abstract
COVID-19, a pandemic disease caused by a viral infection, is associated with a high mortality rate. Most of the signs and symptoms, e.g. cytokine storm, electrolytes imbalances, thromboembolism, etc., are related to mitochondrial dysfunction. Therefore, targeting mitochondrion will represent a more rational treatment of COVID-19. The current work outlines how COVID-19’s signs and symptoms are related to the mitochondrion. Proper understanding of the underlying causes might enhance the opportunity to treat COVID-19.
Introduction
COVID-19 is a new emerging pulmonary infection caused by SARS-COV-2. It is characterised by flu-like symptoms often followed by acute pulmonary inflammation. Multiple viruses are known to cause both inflammation and mitochondrial dysregulation (metabolic shifts). The influenza virus H1N1 targets the mitochondria of type II cellsCitation1. Multiple other inflammatory viruses are known to induce metabolic changes, such as the cytomegalovirus (CMV)Citation2, the Epstein-Barr virus (EBV)Citation3, or the hepatitis virus (HCV)Citation4. These viruses interfere with cellular metabolism, increase glucose uptake, and decrease the mitochondrial energy yield resulting in intense glycolysis. In Caco-2 cells, infection with SARS-CoV-2 has been found to up-regulate carbon metabolism and decrease oxidative phosphorylation. I removed it because it is out of context and there is no reference- also no reference for the Caco-2 cells.
The mitochondrion is a doubled-membrane organelle, represents the backbone of the eukaryote cell metabolismCitation5,Citation6. Mitochondrion is the cells' metabolic generator and plays a significant role in determining cellular proliferationCitation7, cellular death pathwaysCitation8 and also plays a crucial role in maintaining the redox state of the cellCitation9.
Many viral diseases disturb the mitochondrial physiologyCitation10–12, e.g. Epstein–Barr virus (EBV) affects mitochondrial fissionCitation13, herpes simplex virus type 1 (HSV-1) and pseudorabies virus (PRV) affect calcium homeostasisCitation14, and many viruses, e.g. influenza viruses, Hepatitis B virus, support and/or encode proapoptotic proteins that lead to programmed cell deathCitation15–17.
Since the occurrence of unidentified pneumonia patients in Wuhan hospitals in China in late 2019 and the labelling of the disease by the World Health Organisation (WHO) as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the disease became a pandemic in less than three months, and as of the beginning of December 2020 the total confirmed cases of COVID-19 reached 65,257,767 worldwide according to a WHO updateCitation18–20.
Despite the increased global incidence records of the COVID-19 cases, most of the infected patients showed either mild infection with no fever or signs of pneumonia or moderate infection with clinical manifestations like cough, sore throat, fever ≥38 °C, fatigue, and shortness of breathCitation21.
Severe infection with increased mortality rate occurs with pneumonia and respiratory failure. At the same time, other complications might present, such as acute respiratory distress syndrome (ARDS), microvascular thrombosis, coagulopathy, liver injury, acute kidney injury, acute cardiac failure and shockCitation22–27. Factors affecting the infection’s severity are not fully understood; however, factors such as the state of the immune system, viral load, and underlying comorbid diseases might play a role in the severity of the infectionCitation28–30.
In the current work, we present COVID-19 as a mitochondriopathy and demonstrate that many of the hallmarks of COVID-19 are driven by mitochondrial injury.
The role of mitochondria and cytokine storm
Hyperinflammation – e.g. cytokine storm – is a hallmark of COVID-19Citation31. Such hyper-inflammation occurs due to a massive increase in Reactive Oxygen Species (ROS)Citation32,Citation33. Increased ROS results in the release of tumour necrosis factor (TNF)-α and interleukin-1β (IL-1β)Citation34,Citation35. The mitochondrion is a significant source of ROS in mammalian cellsCitation36. Therefore, the mitochondrion lies within the cytokine storm's coreCitation37.
The inflammasome is a cytosolic complex composed of multiple proteins of innate immunity to promote and activate the proinflammatory mediators such as IL-1β, IL-18Citation38–41. One protein component is an intracellular pathogen sensor called nucleotide-binding oligomerization domain-like receptors, or NOD-like receptors (NLRs)Citation42. NLRP3 is one NOD-like receptor (NLRs) family member that represents the backbone of the inflammasome. The role of NLRP3 in inflammation and the cytokine storm is crucial and complex. As a consequence of its activation, the cell reprograms its metabolic machinery into increased glycolysis with a subsequent reduction of the Krebs' cycleCitation43, i.e. induces mitochondrial atrophy. ROS also activates the NLRP3 where it is associated with mitochondrial cardiolipinCitation40 and might be correlated with mitochondrial ageing (which stimulates the inflammasome)Citation44.
SARS-COV-2 infection attacks the mitochondrion, especially the phosphorylation (OxPHOS) pathway, e.g. Complex-ICitation45, which results in abnormal ROS production supporting cellular diseases and ageing. SARS-CoV-2 might directly activate the NLRP3 inflammasome, with consequent flaring-up of the inflammation cascadeCitation40. Hence, SARS-COV-2 alters mitochondrial physiologyCitation46,Citation47.
COVID-19 disrupts the possible mitochondrial role in iron homeostasis
Iron is an essential nutrient and its levels differ from one tissue to another and also depend on the tissues pathological stateCitation48. Cellular iron homeostasis is a complexed processCitation49, but generally, it could be described as: the entrance of iron to the cell through: (i) endocytosis of transferrin receptor 1 (TfR1), or (ii) ferrous iron (Fe+2) transporters e.g. divalent metal transporter 1 (DMT1)Citation50 and Zinc transporters 8, 14 (ZIP8, ZIP14)Citation51,Citation52 with the assistance of the iron reductase enzyme Metalloreductase STEAP2Citation53, Duodenal cytochrome B (Dcytb)Citation52, and Stromal cell-derived receptor 2 (SDR-2)Citation54. After being taken-up, the iron is stored in ferritinCitation55–57 for different biochemical functions including the formation of ROSCitation58,Citation59 and managing transcription through regulating the iron-responsive element-binding proteins (IRP1, IRP2)Citation60,Citation61. After that, iron export from the cell occurs via ferroportin-1 (also termed as solute carrier family 40 member 1 (SLC40A1) or iron-regulated transporter 1 (IREG1))Citation62.
The role of mitochondria in iron homeostasis is one of the most challenging of recently addressed issues. Generally, ferritin is an intracellular protein that can act as an iron-buffering agent to re-equilibrate iron deficiency or iron overloadCitation63. Ferritin is stored in the mitochondrion and imported from the cytoplasm via mitoferrin carriersCitation64,Citation65.
Disruption of mitoferrin leads to hyperferritinemia, accompanied by hyper-inflammation, an additional hallmark of COVID-19 severityCitation64,Citation66,Citation67. Severe iron overload leads to mitochondrial DNA damage that exacerbates the cellular oxidative stressCitation68.
For this reason, the iron-chelating agent, Deferoxamine, has been introduced in the management of COVID-19Citation69,Citation70.
Lactate dehydrogenase in COVID-19
The lactate dehydrogenase (LDH) is an enzyme that catalyses a reversible biochemical reaction that converts pyruvate into lactate. After glucose entry, the hydrogen ions (proton, H+) level is rising, alters the cell's optimum pH to process its chemical pathways. After completing the Krebs' cycle, the cell yields in CO2, energy in ATP, and hydrogen ions. The oxygen reacts with H+ to produce water. Therefore, oxygen in cellular respiration acts as a detoxifying agent (acting as a buffer)Citation71. During transient hypoxia, some tissues, e.g. heart, brain, kidney, are prone to damage.
In contrast, other tissues are slightly adaptable by expressing the lactate dehydrogenase enzyme to shift the cellular metabolism to prevent the Krebs' cycle. Therefore, the glucose utilisation after its entry ends up by forming lactic acid and furthering extracellular acidity via Monocarboxylate Transporters (MCTs)Citation72–74. So, metabolic shifting to end in lactic acid will decrease the possible intracellular acidity and promote the extracellular acidity that exacerbates the cytokine storm as lactate is a signalling molecule that supports inflammationCitation75,Citation76.
The conversion of pyruvate to lactate is associated with the conversion of NADH to NAD+. Increasing of NAD+ level inhibits not only mitochondrial metabolism but also supports the inflammation processCitation77,Citation78.
LDH is correlated with COVID-19 and its severityCitation79 because the lactate synthesis is increased. The level of blood lactate is a prognostic factor for the intensity of the lung's inflammation and decreased survivalCitation80.
Dysregulation of calcium homeostasis during COVID-19 affects mitochondrial biology
Calcium is a vital electrolyte that plays many critical roles in cellular physiologyCitation81. Calcium governs intracellular mitochondrial motility (mitochondrial dynamics)Citation82,Citation83, manages mitophagyCitation84–86, controls ATP productionCitation87, and impacts the role of the mitochondrion in the redox statue of the cellCitation88.
A reduced level of calcium is well-documented in covid-19 infection, and it is thought to have a role in its poor prognosisCitation89. Therefore, hypocalcaemia has a detrimental effect on the mitochondrion, promotes ROS formation, and activates the inflammatory cascade.
The role of the mitochondrion on coagulability
D-dimer
While the term D-dimer reflects the dimerisation process (two subunits), it also seems to be an erroneous name suggested by one of the researchers that discovered itCitation90,Citation91. All in all, D-dimer is fibrin fragments that are crosslinked with polypeptide bonds due to the degradation of fibrinogen via plasminCitation92,Citation93. Higher levels of D-dimer in the blood represent a severe sign of thromboembolismCitation94–96 and recently has become an indicator of how COVID-19 patients develop thromboembolism and the disease severityCitation97–99 since D-dimer level is markedly increased among critical patients and is a significant risk factor for mortalityCitation100
Oxidative stress is associated with thromboembolismCitation101, in that ROS activates urokinase plasminogen activator (UPA)Citation102, subsequently producing plasmin that hydrolyses fibrinogen into D-dimer. The increased Plasmin, in turn, increases ROSCitation103, which produces an out-of-control positive feedback between ROS and plasmin. Furthermore, D-dimer expression also might increase the level of urokinase-type plasminogen activator (plasmin activator), and so it also enters a vicious cycle producing thromboembolism.
There is an inverse relationship between functional mitochondrial and urokinase plasminogen, such that upregulation of the UPA is an indicator of reduced mitochondrial function while, in contrast, downregulation of UPA restores mitochondrial function (e.g. activation of programmed cell death)Citation103.
Troponins
These are a group of proteins found in the heart and skeletal muscle that mediate calcium-dependent muscle contractionCitation104,Citation105. An increased level of troponins in the blood is an indicator of necrosis rather than programmed cell death, i.e. mitochondrial injury or dysfunctionality due to hypoxiaCitation106–112.
COVID-19 is associated with higher troponin levelsCitation113, which might correlate with mortalityCitation114. Indeed, higher troponin levels were confined to cardiac disorder and other diseases, such as sepsis or renal diseaseCitation115, both of which were correlated with COVID-19Citation112,Citation116,Citation117. Also, during cardiac and muscle injury, troponin levels are increased significantly in severe disease patients, leading to progression towards multiple organ failure (MOF) and death.
Targeting the mitochondrion to treat COVID-19
In 1956, Otto Warburg suggested that cancer occurs due to mitochondrial injury and, in this respect, it seems that COVID-19 could be looked at as an extrapolation of cancerCitation118. At least it could be analysed through Warburg's lens and could stimulate the debate of whether mitochondriopathy is a direct cause of COVID-19 via SARS-COV-2 infection or just a symptom of COVID-19 in which, at least, mitochondrial injury might represent an early step of the SARS-COV-2 disease cascade. In this regard, the administration of pharmacological and non-pharmacological modulators of mitochondrial functionCitation119 could enhance patient recovery and improve patients' quality of life and might boost the vaccine's efficacy in the aged population (mitochondrial is a hub of ageing). An example of those agents includes:
NHE1 inhibitors:
In 2000, Reshkin et al. observed that the over-expression of NHE1 is the first event of carcinogenesis followed by alkaline increases in intracellular pH (alkaline pHi)Citation120,Citation121; and alkaline pHi results in mitochondrial atrophy. Therefore, NHE1 inhibition, and specifically mitochondrial NHE1, will boost the mitochondrial functionalityCitation122 and so decrease the effect of SARS-COV-2.
Amiloride is a potassium-sparing diuretic, and it is a well-known NHE-1 inhibitor. Amiloride perturbs SARS-COV-2 biologyCitation123, and early reports showed that Amiloride inhibited coronavirus replicationCitation123
Amiloride also has potential as an anti-cytokine storm agentCitation124. One of the possible mechanisms of action that explains how Amiloride antagonises the cytokine storm via contrasting the effect of proinflammatory mediators (e.g. the NF-κB transcription factor), by boosting the expression of anti-inflammatory mediators such as Interleukin-10 (IL-10), and nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IκBα)Citation124 (see ).
Significantly, Amiloride also suppresses the urokinase plasminogen activator (UPA), which might have a promising role in preventing thromboembolismCitation125,Citation126 and also prevents heart ischaemiaCitation127other NHE1 inhibitors include Cariporide, Eniporide, etc. (see ).
Fermented wheat germ extract:
Fermented wheat germ extract (FWGE) is a dietary supplement used to treat cancer and to slow ageing. The mode of action of FWGE is a mitochondrial restoration agent as it modulates the activity of the pyruvate dehydrogenase (PDH) complex to support the production of ATP from mitochondriaCitation128. Also, FWGE inhibits LDH and reduces the NAD+ levelsCitation128. Moreover, it shows promising action as an anti-cytokine storm drugCitation129–131.
α-lipoic acid:
The history of α-lipoic dates to the 1950s () when German industry developed this drug. The first use of α-lipoic acid was for peripheral neuropathy due to diabetesCitation132.
A preliminary Chinese study suggests the efficacy of α-Lipoic acid in the treatment of COVID-19Citation133, where α-lipoic acid might act in the same way as FWGE; combined with hydroxycitrate, it synergizes the effect as an acting buffer to correct pHi to restore mitochondrial functionCitation134,Citation135.
Methylene Blue
Methylene Blue is the oldest of synthetic drugs (), even before aspirin. Heinrich Carro manufactured it in 1876 for the German firm BASF. Methylene blue is a simple molecule. The fusion of two benzene rings with one nitrogen and one sulphur atom leads to a tricyclic aromatic compound which has a complex pharmacology and multiple clinical indications. Its mechanism of action involves a stabilising effect on mitochondria. Also, Methylene blue inhibits the replication of SARS-CoV-2Citation136 and we reported a cohort of patients treated for cancer by Methylene Blue in cases without SARS-CoV-2Citation137.
2-deoxy-d-glucose (2DG)
The German scientist Otto Warburg discovered the Warburg effect in the 1920sCitation138. Warburg stated that cancer cells display increased glycolysis and lactic acid secretion and, opposite to normal cells, the presence of oxygen does not inhibit this fermentation. The advent of Positron Emission Tomography (PET) scan combined with radio-labelled fluorodeoxyglucose has revived interest in the Warburg effect as there is an increased uptake of labelled glucose in the primary tumour and its distant metastases. The Warburg effect explains some of the cancer's hallmarksCitation118,Citation135 shift to aerobic glycolysis that has been reported to stimulate cell growth, evade tumour suppression, and resist cell deathCitation139. Increased pressure resulting from unrelenting proliferation in the affected organ's limited space results in cells' extrusion in the vasculature and distant metastases. The release of lactic acid in the extracellular space is a consequence of the Warburg effect. Lactic acid promotes angiogenesis and immune cell modulationCitation140.
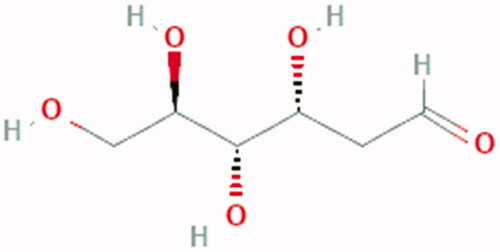
Infection with SARS-CoV-2 in Caco-2 cells has been found to up-regulate glycolytic carbon metabolism and decrease oxidative phosphorylation. In line with this, treatment with the glycolysis inhibitor 2-deoxy-d-glucose (2DG) prevents replication of SARS-CoV-2 in these cellsCitation141 ().
The Warburg hypothesis was based on mitochondrial injury, but the debate is whether it is a cause of malignant transformation or just a consequence. Irrespective of which is correct, mitochondrial damage supports evolutionary tumour trajectoryCitation142. Parallel to this context, COVID-19 is associated with mitochondrial injury and such injury supports SARS-COV-2 pathogenicity and confers its evolutionary advantage. However, a significant concern is whether COVID-19 patients will develop cancer in the future due to such mitochondrial injury?
Figure 1. How does Amiloride re-equilibrate the cytokine storm via boosting the anti-inflammatory cytokines and suppressing the proinflammatory cytokines.

Figure 2. Different chemical formula of some of NHE1 inhibitors.
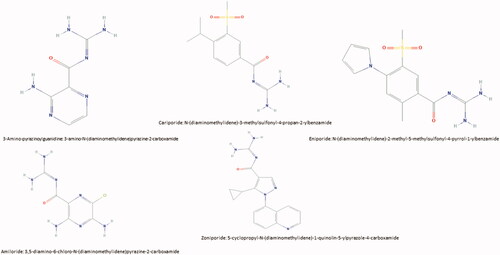
Figure 3. Chemical Structure of lipoic acid: 5-[(3R)-dithiolan-3-yl] pentanoic acid.
![Figure 3. Chemical Structure of lipoic acid: 5-[(3R)-dithiolan-3-yl] pentanoic acid.](/cms/asset/c04451e9-e271-4151-9e99-c56747125a4b/ienz_a_1937144_f0003_c.jpg)
Figure 4. Chemical structure of methylene blue: [7-(dimethylamino) phenothiazin-3-ylidene]-dimethylazanium;chloride.
![Figure 4. Chemical structure of methylene blue: [7-(dimethylamino) phenothiazin-3-ylidene]-dimethylazanium;chloride.](/cms/asset/3c1ed1f9-3cd7-4938-9c6f-76320de44194/ienz_a_1937144_f0004_c.jpg)
Figure 5. Chemical Structure of 2DG: (3 R,4S,5R)-3,4,5,6-tetrahydroxyhexanal.
Recommendations and concluding remarks
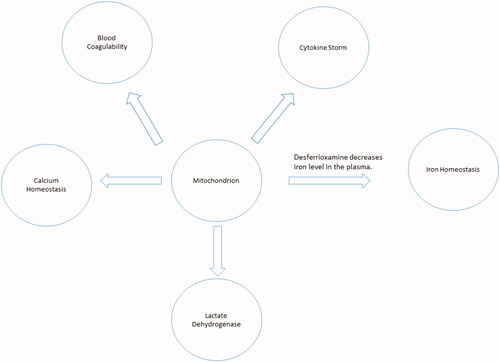
COVID-19 has become a pandemic disease. The biology of the disease is exceptionally intricate, including many overlapping pathways. However, while the mitochondrion lies at the core of these pathways, its importance demands immediate attention and further investigation. A proper understanding of mitochondrial biology in COVID-19 pathogenesis will significantly enhance the strategy of fighting SARS-COV-2 (). This paper has discussed and suggests a couple of pharmacological modulators that might represent potentially promising anti-COVID-19 treatments to block its progression and alleviate its aggressiveness.
Figure 6. The mitochondrion lies within the core of COVID-19 cardinals.
Author contributions
KOA contributed to the conceptualisation, data curation, formal analysis, investigation, resources, software, writing (original draft). SJR contributed to the supervision, conceptualisation, data curation, formal analysis, investigation, resources, software, writing (review and editing). STA, AH, and LS contributed to the conceptualisation, data curation, resources, writing (original draft). ASA, SBMA, AMA, and SSA contributed to methodology, resources, software. AKM, HA, AHHB, and MI contributed to the investigation, methodology, visualisation. SH, MR, and RAC contributed to investigation, methodology, and resources. SH also contributed to review and correct the final text.
Disclosure statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Davis I, Doolittle L, Guttridge D, et al. H1N1 influenza A virus infection of mice induces the warburg effect in ATII cells. Am J Respir Crit Care Med 2020;201:A3842.
- Yu Y, Clippinger AJ, Alwine JC. Viral effects on metabolism: changes in glucose and glutamine utilization during human cytomegalovirus infection. Trends Microbiol 2011;19:360–7.
- Darekar S, Georgiou K, Yurchenko M, et al. Epstein-barr virus immortalization of human B-cells leads to stabilization of hypoxia-induced factor 1 alpha, congruent with the Warburg effect. PLoS One 2012;7:e42072.
- Ripoli M, D'Aprile A, Quarato G, et al. Hepatitis C virus-linked mitochondrial dysfunction promotes hypoxia-inducible factor 1α-mediated glycolytic adaptation. J Virol 2010;84:647–60.
- Gabaldón T, Huynen MA. From endosymbiont to host-controlled organelle: the hijacking of mitochondrial protein synthesis and metabolism. Subramaniam S, ed. PLoS Comput Biol 2007;3:e219.
- Pittis AA, Gabaldón T. Late acquisition of mitochondria by a host with chimaeric prokaryotic ancestry. Nature 2016;531:101–4.
- Yan XJ, Yu X, Wang XP, et al. Mitochondria play an important role in the cell proliferation suppressing activity of berberine. Sci Rep 2017;7:41712.
- Vakifahmetoglu-Norberg H, Ouchida AT, Norberg E. The role of mitochondria in metabolism and cell death. Biochem Biophys Res Commun 2017;482:426–31.
- Handy DE, Loscalzo J. Redox regulation of mitochondrial function. Antioxid Redox Signal 2012;16:1323–67.
- Reshi L, Wang H-V, Hong J-R. Modulation of mitochondria during viral infections. In: Taskin E, Guven C, eds. Mitochondrial diseases. London: InTech; 2018.
- Khan M, Syed GH, Kim SJ, et al. Mitochondrial dynamics and viral infections: a close nexus. Biochim Biophys Acta 2015;1853:2822–33.
- Tiku V, Tan MW, Dikic I. Mitochondrial functions in infection and immunity. Trends Cell Biol 2020;30:263–75.
- Young LS, Rickinson AB. Epstein-Barr virus: 40 years on. Nat Rev Cancer 2004;4:757–68.
- Kramer T, Enquist LW. Alphaherpesvirus infection disrupts mitochondrial transport in neurons. Cell Host Microbe 2012;11:504–14.
- Takada S, Shirakata Y, Kaneniwa N, et al. Association of hepatitis B virus X protein with mitochondria causes mitochondrial aggregation at the nuclear periphery, leading to cell death. Oncogene 1999;18:6965–73.
- Hennet T, Peterhans E, Stocker R. Alterations in antioxidant defences in lung and liver of mice infected with influenza A virus. J General Virol 1992;73:39–46.
- Henkler F, Hoare J, Waseem N, et al. Intracellular localization of the hepatitis B virus HBx protein. J General Virol 2001;82:871–82.
- Heinze G, Schemper M. A solution to the problem of separation in logistic regression. Stat Med 2002;21:2409–19.
- Chen H, Guo J, Wang C, et al. Clinical characteristics and intrauterine vertical transmission potential of COVID-19 infection in nine pregnant women: a retrospective review of medical records. Lancet 2020;395:809–15.
- Anon. WHO Coronavirus Disease (COVID-19) Dashboard | WHO Coronavirus Disease (COVID-19) Dashboard. https://covid19.who.int/ [accessed 5 Dec 2020].
- Guan W, Ni Z, Hu Y, et al. Clinical characteristics of coronavirus disease 2019 in China. New Engl J Med 2020;382:1708–20.
- Sánchez-Recalde Á, Solano-López J, Miguelena-Hycka J, et al. COVID-19 and cardiogenic shock. Different cardiovascular presentations with high mortality. Revista Española de Cardiología 2020;73:669–72.
- Du R-H, Liang L-R, Yang C-Q, et al. Predictors of mortality for patients with COVID-19 pneumonia caused by SARS-CoV-2: a prospective cohort study. Eur Respir J 2020;55:2000524.
- Yang HJ, Zhang YM, Yang M, et al. Predictors of mortality for patients with COVID-19 pneumonia caused by SARS-CoV-2. Eur Respir J 2020;56:2002439.
- Ciceri F, Ruggeri A, Lembo R, et al. Decreased in-hospital mortality in patients with COVID-19 pneumonia. Pathogens Global Health 2020;114:281–2.
- Paek JH, Kim Y, Park WY, et al. Severe acute kidney injury in COVID-19 patients is associated with in-hospital mortality. Hirst JA, ed. PLoS One 2020;15:e0243528.
- Malas MB, Naazie IN, Elsayed N, et al. Thromboembolism risk of COVID-19 is high and associated with a higher risk of mortality: a systematic review and meta-analysis. EClinicalMedicine 2020;29-30:100639.
- Guo T, Fan Y, Chen M, et al. Cardiovascular implications of fatal outcomes of patients with coronavirus disease 2019 (COVID-19). JAMA Cardiol 2020;5:811–8.
- Bhatraju PK, Ghassemieh BJ, Nichols M, et al. Covid-19 in Critically ill patients in the seattle region – case series. New Engl J Med 2020;382:2012–22.
- Zheng S, Fan J, Yu F, et al. Viral load dynamics and disease severity in patients infected with SARS-CoV-2 in Zhejiang province, China, January. Retrospective cohort study. BMJ 2020;369:m1443.
- Coperchini F, Chiovato L, Croce L, et al. The cytokine storm in COVID-19: an overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev 2020;53:25–32.
- Mittal M, Siddiqui MR, Tran K, et al. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal 2014;20:1126–67.
- Naha PC, Davoren M, Lyng FM, et al. Reactive oxygen species (ROS) induced cytokine production and cytotoxicity of PAMAM dendrimers in J774A.1 cells. Toxicol Appl Pharmacol 2010;246:91–9.
- Yang D, Elner SG, Bian ZM, et al. Pro-inflammatory cytokines increase reactive oxygen species through mitochondria and NADPH oxidase in cultured RPE cells. Exp Eye Res 2007;85:462–72.
- Kamata H, Honda SI, Maeda S, et al. Reactive oxygen species promote TNFα-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell 2005;120:649–61.
- Murphy MP. How mitochondria produce reactive oxygen species. Biochem J 2009;417:1–13.
- Saleh J, Peyssonnaux C, Singh KK, et al. Mitochondria and microbiota dysfunction in COVID-19 pathogenesis. Mitochondrion 2020;54:1–7.
- Mariathasan S, Newton K, Monack DM, et al. Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 2004;430:213–8.
- Broz P, Dixit VM. Inflammasomes: mechanism of assembly, regulation and signalling. Nat Rev Immunol 2016;16:407–20.
- Martinon F, Burns K, Tschopp J. The Inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol Cell 2002;10:417–26.
- Sui A, Chen X, Shen J, et al. Inhibiting the NLRP3 inflammasome with MCC950 ameliorates retinal neovascularization and leakage by reversing the IL-1β/IL-18 activation pattern in an oxygen-induced ischemic retinopathy mouse model. Cell Death Dis 2020;11:901.
- Shaw MH, Reimer T, Kim YG, et al. NOD-like receptors (NLRs): bona fide intracellular microbial sensors. Curr Opin Immunol 2008;20:377–82.
- Swanson KV, Deng M, Ting JPY. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol 2019;19:477–89.
- Kauppinen A. Mitochondria-associated inflammasome activation and its impact on aging and age-related diseases. In: Fulop T, Franceschi C, Hirokawa K, Pawelec G, eds. Handbook of immunosenescence. Cham: Springer International Publishing; 2018:1–20.
- Ouyang L, Gong J. Mitochondrial-targeted ubiquinone: a potential treatment for COVID-19. Med Hypotheses 2020;144:110161.
- Singh KK, Chaubey G, Chen JY, et al. Decoding sars-cov-2 hijacking of host mitochondria in covid-19 pathogenesis. Am J Physiol Cell Physiol 2020;319:C258–C267.
- Ganji R, Reddy PH. Impact of COVID-19 on mitochondrial-based immunity in aging and age-related diseases. Front Aging Neurosci 2020;12:614650.
- Zhao N, Enns CA. Iron transport machinery of human cells, players and their interactions. In: Orlov S, ed. Current topics in membranes. Vol. 69. San Diego: Academic Press Inc.; 2012:67–93.
- Collins JF, Anderson GJ. Molecular mechanisms of intestinal iron transport. In: Johnson LR, ed. Physiology of the gastrointestinal tract. Vol. 2. London: Elsevier Inc.; 2012:1921–1947.
- Garrick MD. Human iron transporters. Genes Nutr 2011;6:45–54.
- Liuzzi JP, Aydemir F, Nam H, et al. Zip14 (Slc39a14) mediates non-transferrin-bound iron uptake into cells. Proc Natl Acad Sci USA 2006;103:13612–7.
- McKie AT, Barrow D, Latunde-Dada GO, et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science 2001;291:1755–9.
- Ji C, Kosman DJ. Molecular mechanisms of non-transferrin-bound and transferring-bound iron uptake in primary hippocampal neurons. J Neurochem 2015;133:668–83.
- Vargas JD, Herpers B, McKie AT, et al. Stromal cell-derived receptor 2 and cytochrome b561 are functional ferric reductases. Biochim Biophys Acta 2003;1651:116–23.
- Mackenzie EL, Iwasaki K, Tsuji Y. Intracellular iron transport and storage: from molecular mechanisms to health implications. Antioxid Redox Signal 2008;10:997–1030.
- Arosio P, Elia L, Poli M. Ferritin, cellular iron storage and regulation. IUBMB Life 2017;69:414–22.
- De Domenico I, McVey Ward D, Kaplan J. Regulation of iron acquisition and storage: consequences for iron-linked disorders. Nat Rev Mol Cell Biol 2008;9:72–81.
- Bystrom LM, Guzman ML, Rivella S. Iron and reactive oxygen species: friends or foes of cancer cells? Antioxid Redox Signal 2014;20:1917–24.
- Dixon SJ, Stockwell BR. The role of iron and reactive oxygen species in cell death. Nat Chem Biol 2014;10:9–17.
- Cairo G, Recalcati S. Iron-regulatory proteins: molecular biology and pathophysiological implications. Exp Rev Mol Med 2007;9:1–13.
- Zhou ZD, Tan EK. Iron regulatory protein (IRP)-iron responsive element (IRE) signaling pathway in human neurodegenerative diseases. Mol Neurodegener 2017;12:1–12.
- Yanatori I, Richardson DR, Imada K, et al. Iron export through the transporter ferroportin 1 is modulated by the iron chaperone PCBP2. J Biol Chem 2016;291:17303–18.
- Daru J, Colman K, Stanworth SJ, et al. Serum ferritin as an indicator of iron status: what do we need to know? Am J Clin Nutr 2017;106:1634S–9S. Vol
- Paradkar PN, Zumbrennen KB, Paw BH, et al. Regulation of mitochondrial iron import through differential turnover of mitoferrin 1 and mitoferrin 2. Mol Cell Biol 2009;29:1007–16.
- Shaw GC, Cope JJ, Li L, et al. Mitoferrin is essential for erythroid iron assimilation. Nature 2006;440:96–100.
- Gao G, Chang YZ. Mitochondrial ferritin in the regulation of brain iron homeostasis and neurodegenerative diseases. Front Pharmacol 2014;5:19.
- Chen C, Paw BH. Cellular and mitochondrial iron homeostasis in vertebrates. Biochim Biophys Acta 2012;1823:1459–67.
- Walter PB, Knutson MD, Paler-Martinez A, et al. Iron deficiency and iron excess damage mitochondria and mitochondrial DNA in rats. Proc Natl Acad Sci USA 2002;99:2264–9.
- Vlahakos VD, Marathias KP, Arkadopoulos N, et al. Hyperferritinemia in patients with COVID‐19: an opportunity for iron chelation? Artif Organs 2021;45:163–7. aor.
- Abobaker A. Can iron chelation as an adjunct treatment of COVID-19 improve the clinical outcome? Eur J Clin Pharmacol 2020;76:1619–20.
- Alfarouk KO, Verduzco D, Rauch C, et al. Glycolysis, tumor metabolism, cancer growth and dissemination. A new pH-based etiopathogenic perspective and therapeutic approach to an old cancer question. Oncoscience 2014;1:777–802.
- Alfarouk KO. Tumor metabolism, cancer cell transporters, and microenvironmental resistance. J Enzyme Inhibit Med Chem 2016;31:859–8.
- Hertz L, Dienel GA. Lactate transport and transporters: general principles and functional roles in brains cells. J Neurosci Res 2005;79:11–8.
- Bosshart PD, Kalbermatter D, Bonetti S, et al. Mechanistic basis of L-lactate transport in the SLC16 solute carrier family. Nature Commun 2019;10:2649.
- Haas R, Smith J, Rocher-Ros V, et al. Lactate regulates metabolic and proinflammatory circuits in control of T cell migration and effector functions. PLoS Biol 2015;13:e1002202.
- Pucino V, Bombardieri M, Pitzalis C, et al. Lactate at the crossroads of metabolism, inflammation, and autoimmunity. Eur J Immunol 2017;47:14–21.
- Gerner RR, Klepsch V, Macheiner S, et al. NAD metabolism fuels human and mouse intestinal inflammation. Gut 2018;67:1813–23.
- Adriouch S, Hubert S, Pechberty S, et al. NAD + released during inflammation participates in T cell homeostasis by inducing ART2-mediated death of naive T cells in vivo. J Immunol 2007;179:186–94.
- Henry BM, Aggarwal G, Wong J, et al. Lactate dehydrogenase levels predict coronavirus disease 2019 (COVID-19) severity and mortality: a pooled analysis. Am J Emerg Med 2020;38:1722–6.
- Booth AL, Abels E, McCaffrey P. Development of a prognostic model for mortality in COVID-19 infection using machine learning. Modern Pathol 2021;34:522–31.
- Romero-Garcia S, Prado-Garcia H. Mitochondrial calcium: transport and modulation of cellular processes in homeostasis and cancer (Review). Int J Oncol 2019;54:1155–67.
- Paupe V, Prudent J. New insights into the role of mitochondrial calcium homeostasis in cell migration. Biochem Biophys Res Commun 2018;500:75–86.
- Contreras L, Drago I, Zampese E, et al. Mitochondria: the calcium connection. Biochim Biophys Acta 2010;1797:607–18.
- Gandhi S, Wood-Kaczmar A, Yao Z, et al. PINK1-associated Parkinson’s disease is caused by neuronal vulnerability to calcium-induced cell death. Mol Cell 2009;33:627–38.
- Dagda RK, Cherra SJ, Kulich SM, et al. Loss of PINK1 function promotes mitophagy through effects on oxidative stress and mitochondrial fission. J Biol Chem 2009;284:13843–55.
- Gelmetti V, De Rosa P, Torosantucci L, et al. PINK1 and BECN1 relocalize at mitochondria-associated membranes during mitophagy and promote ER-mitochondria tethering and autophagosome formation. Autophagy 2017;13:654–69.
- Jouaville LS, Pinton P, Bastianutto C, et al. Regulation of mitochondrial ATP synthesis by calcium: evidence for a long-term metabolic priming. Proc Natl Acad Sci USA 1999;96:13807–12.
- Kennedy ED, Rizzuto R, Theler JM, et al. Glucose-stimulated insulin secretion correlates with changes in mitochondrial and cytosolic Ca2+ in aequorin-expressing INS-1 cells. J Clin Invest 1996;98:2524–38.
- Yang C, Ma X, Wu J, et al. Low serum calcium and phosphorus and their clinical performance in detecting COVID‐19 patients. J Med Virol 2021;93:1639–51.
- Gaffney PJ, Brasher M. Subunit structure of the plasmin-induced degradation products of crosslinked fibrin. BBA 1973;295:308–13.
- Gaffney PD. dimer. History of the discovery, characterisation and utility of this and other fibrin fragments. Fibrinol Proteol 1993;7:2–8.
- Haverkate F, Timan G. Protective effect of calcium in the plasmin degradation of fibrinogen and fibrin fragments D. Thrombosis Res 1977;10:803–12.
- Favresse J, Lippi G, Roy P-M, et al. D-dimer: preanalytical, analytical, postanalytical variables, and clinical applications. Crit Rev Clin Lab Sci 2018;55:548–77.
- Kelly J, Rudd A, Lewis RR, et al. Plasma D-dimers in the diagnosis of venous thromboembolism. Arch Intern Med 2002;162:747–56.
- Pulivarthi S, Gurram MK. Effectiveness of D-dimer as a screening test for venous thromboembolism: an update. North Am J Med Sci 2014;6:491–9.
- Matsuo H, Nakajima Y, Ogawa T, et al. Evaluation of D-dimer in screening deep vein thrombosis in hospitalized japanese patients with acute medical diseases/Episodes. Ann Vasc Dis 2016;9:193–200.
- Cho ES, McClelland PH, Cheng O, et al. Utility of D-dimer for diagnosis of deep vein thrombosis in coronavirus disease-19 infection. J Vasc Surg 2021;9:47–53.
- Yu HH, Qin C, Chen M, et al. D-dimer level is associated with the severity of COVID-19. Thrombosis Res 2020;195:219–25.
- Yao Y, Cao J, Wang Q, et al. D-dimer as a biomarker for disease severity and mortality in COVID-19 patients: a case control study. J Intens Care 2020;8:49.
- Zhou F, Yu T, Du R, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet 2020;395:1054–62.
- Wang Q, Zennadi R. Oxidative stress and thrombosis during aging: the roles of oxidative stress in RBCS in venous thrombosis. Int J Mol Sci 2020;21:1–22.
- Lee KH, Kim JR. Reactive oxygen species regulate the generation of urokinase plasminogen activator in human hepatoma cells via MAPK pathways after treatment with hepatocyte growth factor. Exp Mol Med 2009;41:180–8.
- Tykhomyrov AA, Zhernosekov DD, Guzyk MM, et al. Plasminogen modulates formation of reactive oxygen species in human platelets. Ukrain Biochem J 2018;90:31–40.
- Wijnker PJM, Sequeira V, Foster DB, et al. Length-dependent activation is modulated by cardiac troponin I bisphosphorylation at Ser23 and Ser24 but not by Thr143 phosphorylation. Am J Physiol 2014;306:H1171–H1181.
- Sun YB, Lou F, Irving M. Calcium- and myosin-dependent changes in troponin structure during activation of heart muscle. J Physiol 2009;587:155–63.
- Korff S, Katus HA, Giannitsis E. Differential diagnosis of elevated troponins. Heart 2006;92:987–93.
- Skeik N, Patel DC. A review of troponins in ischemic heart disease and other conditions. Int J Angiol 2007;16:53–8.
- Daubert MA, Jeremias A. The utility of troponin measurement to detect myocardial infarction: review of the current findings. Vasc Health Risk Manage 2010;6:691–9.
- Okamoto R, Hirashiki A, Cheng XW, et al. Usefulness of serum cardiac troponins T and I to predict cardiac molecular changes and cardiac damage in patients with hypertrophic cardiomyopathy. Int Heart J 2013;54:202–6.
- Pankuweit S, Richter A. Mitochondrial disorders with cardiac dysfunction: an under-reported aetiology with phenotypic heterogeneity. Eur Heart J. 36:2894–7.
- Amgalan D, Pekson R, Kitsis RN. Troponin release following brief myocardial ischemia: apoptosis versus necrosis. JACC 2017;2:118–21.
- Park KC, Gaze DC, Collinson PO, et al. Cardiac troponins: from myocardial infarction to chronic disease. Cardiovasc Res 2017;113:1708–18.
- Tersalvi G, Vicenzi M, Calabretta D, et al. Elevated troponin in patients with coronavirus disease 2019: possible mechanisms. J Cardiac Fail 2020;26:470–5.
- Lombardi CM, Carubelli V, Iorio A, et al. Association of troponin levels with mortality in italian patients hospitalized with coronavirus disease 2019: results of a multicenter study. JAMA Cardiol 2020;5:1274–E7.
- Mannu GS. The non-cardiac use and significance of cardiac troponins. Scott Med J 2014;59:172–8.
- Collado S, Arenas MD, Barbosa F, et al. COVID-19 in grade 4–5 chronic kidney disease patients. Kidney Blood Pressure Res 2020;45:768–74.
- Ajaimy M, Melamed ML. Covid-19 in patients with kidney disease. Clin J Am Soc Nephrol 2020;15:1087–9.
- Alfarouk KO, Shayoub MEA, Muddathir AK, et al. Evolution of tumor metabolism might reflect carcinogenesis as a reverse evolution process (dismantling of multicellularity). Cancers 2011;3:3002–17.
- Andreux PA, Houtkooper RH, Auwerx J. Pharmacological approaches to restore mitochondrial function. Nat Rev Drug Discovery 2013;12:465–83.
- Reshkin SJ, Bellizzi A, Caldeira S, et al. Na+/H+ exchanger-dependent intracellular alkalinization is an early event in malignant transformation and plays an essential role in the development of subsequent transformation-associated phenotypes. FASEB J 2000;14:2185–97.
- Cardone RAR, Alfarouk KOK, Elliott RLR, et al. The role of sodium hydrogen exchanger 1 in dysregulation of proton dynamics and reprogramming of cancer metabolism as a sequela. Int J Mol Sci 2019;20:3694.
- Alvarez BV, Villa-Abrille MC. Mitochondrial NHE1: a newly identified target to prevent heart disease. Front Physiol 2013;4:152.
- Wilson L, Gage P, Ewart G. Hexamethylene amiloride blocks E protein ion channels and inhibits coronavirus replication. Virology 2006;353:294–306.
- Haddad JJ, Land SC. Amiloride blockades lipopolysaccharide-induced proinflammatory cytokine biosynthesis in an IκB-α/NF-κB-dependent mechanism: evidence for the amplification of an antiinflammatory pathway in the alveolar epithelium. Am J Respir Cell Mol Biol 2002;26:114–26.
- Jankun J, Skrzypczak-Jankun E. Molecular basis of specific inhibition of urokinase plasminogen activator by amiloride. Cancer Biochem Biophys 1999;17:109–23.
- Vassalli JD, Belin D. Amiloride selectively inhibits the urokinase-type plasminogen activator. FEBS Lett 1987;214:187–91.
- Javadov S, Choi A, Rajapurohitam V, et al. NHE-1 inhibition-induced cardioprotection against ischaemia/reperfusion is associated with attenuation of the mitochondrial permeability transition. Cardiovasc Res 2007;77:416–24.
- Bencze G, Bencze S, Rivera KD, et al. Mito-oncology agent: fermented extract suppresses the Warburg effect, restores oxidative mitochondrial activity, and inhibits in vivo tumor growth. Sci Rep 2020;10:14174.
- Mueller T, Voigt W. Fermented wheat germ extract – nutritional supplement or anticancer drug? Nutr J 2011;10:89.
- Sukkar SG, Cella F, Rovera GM, et al. A multicentric prospective open trial on the quality of life and oxidative stress in patients affected by advanced head and neck cancer treated with a new benzoquinone-rich product derived from fermented wheat germ (Avemar). Mediterranean J Nutr Metab 2008;1:37–42.
- Jeong H-Y, Choi Y-S, Lee J-K, et al. Anti-inflammatory activity of citric acid-treated wheat germ extract in lipopolysaccharide-stimulated macrophages. Nutrients 2017;9:730.
- Mijnhout GS, Kollen BJ, Alkhalaf A, et al. Alpha lipoic acid for symptomatic peripheral neuropathy in patients with diabetes: a meta-analysis of randomized controlled trials. Int J Endocrinol 2012;2012:2012.
- Zhong M, Sun A, Xiao T, et al. A randomized, single-blind, group sequential, active-controlled study to evaluate the clinical efficacy and safety of α-Lipoic acid for critically ill patients with coronavirus disease 2019 (COVID-19). medRxiv 2020:2020.04.15.20066266.
- Schwartz L, Seyfried T, Alfarouk KO, et al. Out of Warburg effect: an effective cancer treatment targeting the tumor specific metabolism and dysregulated pH. Semin Cancer Biol 2017;43:134–8.
- Schwartz L, Supuran CT, Alfarouk KO. The Warburg effect and the hallmarks of cancer. Anti-Cancer Agents Med Chem 2017;17:164–70.
- Gendrot M, Andreani J, Duflot I, et al. Methylene blue inhibits replication of SARS-CoV-2 in vitro. Int J Antimicrob Agents 2020;56:106202.
- Henry M, Summa M, Patrick L, et al. A cohort of cancer patients with no reported cases of SARS-CoV-2 infection: the possible preventive role of Methylene Blue. Substantia 2020;4:888.
- Warburg O. On the origin of cancer cells. Science 1956;123:309–14.
- Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 2009;324:1029–1033.
- Dhup S, Dadhich RK, Porporato PE, et al. Multiple biological activities of lactic acid in cancer: influences on tumor growth, angiogenesis and metastasis. Curr Pharm Des 2012;18:1319–30.
- Ayres JS. A metabolic handbook for the COVID-19 pandemic. Nat Metab 2020;2:572–585.
- Alfarouk KO, Muddathir AK, Shayoub MEA. Tumor acidity as evolutionary spite. Cancers 2011;3:408–414.