Abstract
Objective. Fragment Bb is an activator of the alternative pathway of the complement system. Recently, increased first trimester maternal plasma concentrations of this fragment were reported in patients destined to have a spontaneous preterm delivery before 34 weeks of gestation. The aim of this study was to determine whether the amniotic fluid (AF) concentrations of fragment Bb change with gestational age, spontaneous labor (term and preterm) and in the presence of intra-amniotic infection/inflammation (IAI).
Study design. This cross-sectional study included patients in the following groups: (1) mid-trimester (n = 64); (2) term in spontaneous labor (n = 70); (3) term not in labor (n = 43); (4) spontaneous preterm labor (PTL) who delivered at term (n = 76); (5) PTL without IAI who delivered preterm (n = 73); (6) PTL with IAI (n = 76); (7) preterm prelabor rupture of membranes (PROM) without IAI (n = 71); and (8) preterm PROM with IAI (n = 71). Fragment Bb concentration in AF was determined by an enzyme-linked immunoassay. Non-parametric statistics were used for analyses.
Results. (1) Fragment Bb was detected in all AF samples (n = 544); (2) The median AF concentration of fragment Bb in patients at term not in labor was significantly higher than that of those in the mid-trimester [2.42 μg/ml, interquartile range (IQR) 1.78–3.22 vs. 1.64 μg/ml, IQR 1.06–3.49; p < 0.001]; (3) Among patients with PTL, those with IAI had a higher median AF fragment Bb concentration than that of woman without IAI, who delivered preterm (4.82 μg/ml, IQR 3.32–6.08 vs. 3.67 μg/ml, IQR 2.35–4.57; p < 0.001) and than that of women with an episode of PTL, who delivered at term (3.21 μg/ml, IQR 2.39–4.16; p < 0.001); (4) Similarly, among patients with preterm PROM, the median AF fragment Bb concentration was higher in individuals with IAI than in those without IAI (4.24 μg/ml, IQR 2.58–5.79 vs. 2.79 μg/ml, IQR 2.09–3.89; p < 0.001). (5) Among patients at term, the median AF fragment Bb concentration did not differ between women with spontaneous labor and those without labor (term in labor: 2.47 μg/ml, IQR 1.86–3.22; p = 0.97).
Conclusions. (1) Fragment Bb, an activator of the alternative complement pathway, is a physiologic constituent of the AF, and its concentration increases with advancing gestational age; (2) AF concentrations of fragment Bb are higher in pregnancies complicated with IAI; and (3) labor at term is not associated with changes in the AF concentrations of fragment Bb. These findings suggest a role for fragment Bb in the host immune response against IAI.
Introduction
Preterm parturition is syndromic in nature Citation[1],Citation[2], and several mechanisms of disease have been implicated in its pathophysiology, including intrauterine infection, uterine ischemia, uterine overdistension, abnormal allogenic recognition, allergic-like reaction, cervical disease and endocrine disorders Citation[2]. Nevertheless, intrauterine infection Citation[3-13] is the only pathological process for which a firm causal link with preterm birth has been established, and a defined molecular pathophysiology is known Citation[1]. Of note, the pregnancy outcome of patients with spontaneous preterm labor (PTL) with intra-amniotic inflammation defined by elevated pro-inflammatory cytokines [e.g., interleukin (IL)-6 Citation[14-16] and matrix metalloproteinase-8 Citation[17]], is similar to that of those with microbiologically proven intra-amniotic infection Citation[16]. Both intra-amniotic infection and inflammation are associated with the development of a fetal inflammatory response syndrome (FIRS) Citation[18-20], which is a risk factor for fetal injury Citation[14],Citation[21-29].
The complement system is an important component of innate immunity and plays a pivotal role in the process of recognition of foreign antigens and pathogens. In addition, the complement system participates in the inflammatory response elicited against infection and has a role in activating the adaptive immune system Citation[30-32]. Three different pathways can trigger complement activation: the ‘classical’, ‘lectin’ and ‘alternative’. Of note, these pathways converge at the point of C3 convertase generation Citation[32],Citation[33].
The classical and the lectin pathways are initiated by the binding of recognition proteins to specific targets (protein-to-protein and protein-to-carbohydrate interactions, respectively) Citation[32]. In contrast, the alternative pathway does not depend on binding of a protein to a pathogen but is capable of auto-activation by spontaneous hydrolysis of C3 in the plasma generating C3(H2O) Citation[32],Citation[34]. The latter is able to bind factor B, allowing its cleavage by factor D into fragments Ba and Bb. The C3(H2O)Bb complex can cleave additional C3 molecules, generating C3b that, in turn, associates with factor B to generate more C3-convertase Citation[32]. The alternative pathway can also be activated through an ‘amplification loop’, in which fixed C3b generated by the classical or lectin pathways binds factor B Citation[31].
During pregnancy, there is a physiologic activation of the complement system in the maternal circulation, which has been proposed to be a compensatory mechanism aimed to protect the host against infection Citation[35]. However, several pregnancy complications including spontaneous pregnancy losses Citation[36-41], preeclampsia Citation[42],Citation[43], pyelonephritis Citation[44], fetal death Citation[45] and preterm birth Citation[13] have been associated with an excessive systemic maternal complement activation.
Increased activation of components of the complement system in maternal blood (C3a and C5a) Citation[13] and the amniotic fluid (C3) Citation[46] has been reported in patients with PTL and intact membranes. Recently, increased maternal plasma concentrations of fragment Bb in the first trimester were reported in patients who subsequently had a spontaneous preterm delivery before 34 weeks of gestation Citation[47]. However, to date, there is limited information regarding the concentration of fragment Bb in amniotic fluid (AF).
This study was conducted to determine whether the AF concentration of fragment Bb changes with advancing gestational age, spontaneous labor at term, and in the presence of intra-amniotic infection/inflammation (IAI) in patients with spontaneous PTL and intact membranes, as well as in women with preterm prelabor rupture of membranes (preterm PROM).
Materials and methods
Study design and population
A cross-sectional study was conducted by searching our clinical database and bank of biological specimens, and consisted of patients in the following groups: (1) women in the mid-trimester of pregnancy (14–18 weeks) who underwent amniocentesis for genetic indications and delivered a normal neonate at term (n = 64); (2) normal pregnant women at term with spontaneous labor (n = 70); (3) normal pregnant women at term not in labor (n = 43); (4) women with an episode of PTL and intact membranes who delivered at term (n = 76); (5) PTL without IAI who delivered preterm (<37 weeks gestation) (n = 73); (6) PTL with IAI (n = 76); (7) women with preterm PROM without IAI (n = 71) and (8) preterm PROM with IAI (n = 71).
All women involved in the study provided written informed consent before the collection of AF. The collection of AF and its utilization for research purposes were approved by the Institutional Review Boards of participating institutions and the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD/NIH/DHHS). Many of these samples have previously been used to study the biology of inflammation, haemostasis, angiogenesis regulation and growth factor concentrations in normal pregnant women and those with pregnancy complications.
Definitions
Patients were considered to have a normal pregnancy outcome if they did not have obstetrical complications and delivered a term neonate (≥37 weeks) of appropriate birthweight for gestational age Citation[48],Citation[49] without complications. Spontaneous PTL was defined by the presence of regular uterine contractions occurring at a frequency of at least two every 10 min associated with cervical changes before 37 completed weeks of gestation that required hospitalization. Preterm PROM was diagnosed by sterile speculum examination confirming pooling of AF in the vagina in association with nitrazine and ferning tests when necessary, before 37 weeks of gestation and in the absence of labor. Women at term not in labor underwent amniocentesis for the assessment of fetal lung maturity before cesarean section. Women at term in labor consisted of women who were suspected to have PTL because of uncertain dates and had an amniocentesis for the assessment of fetal lung maturity and microbial invasion of the amniotic cavity. If analysis of AF was consistent with maturity, tocolysis was not used. In addition, if the women delivered a baby heavier than 2500 g without complications of prematurity, they were considered to represent patients in spontaneous labor at term. Intra-amniotic infection was defined as a positive AF culture for microorganisms. Intra-amniotic inflammation was diagnosed in the presence of an AF IL-6 concentration ≥2.6 ng/ml Citation[16]. Acute histologic chorioamnionitis was diagnosed based on the presence of inflammatory cells in the chorionic plate and/or chorioamniotic membranes. Acute funisitis was defined by the presence of neutrophils in the wall of the umbilical vessels and/or Wharton's jelly using the criteria previously described Citation[50].
Sample collection
Amniotic fluid samples were obtained from transabdominal amniocenteses performed for evaluation of microbial status of the amniotic cavity and/or assessment of fetal lung maturity. Sample of AF was transported to the laboratory in a sterile-capped syringe and cultured for aerobic/anaerobic bacteria and genital Mycoplasmas. White blood cell (WBC) count, glucose concentration and Gram stain were also performed shortly after collection. The results of these tests were used for subsequent clinical management. AF not required for clinical assessment was centrifuged for 10 min at 4°C, and the supernatant was aliquoted and stored at –70°C until analysis. Mid-trimester samples were not evaluated for infection. However, all had an AF IL-6 concentration <2.6 ng/ml.
Among patients with spontaneous PTL with intact membranes who delivered within 72 h of amniocentesis, placenta, umbilical cord and chorioamniotic membranes were collected, and the presence or absence of histologic chorioamnionitis and/or funisitis was assessed. The 72-h interval was chosen to preserve a meaningful temporal relationship between AF fragment Bb concentration and placental histopathologic findings.
Determination of fragment Bb concentration in amniotic fluid
AF concentration of human fragment Bb was determined by sensitive enzyme-linked immunoassays (Quidel Corporation, San Diego, CA). Fragment Bb immunoassay was validated for human AF in our laboratory, before the start of this study. Validation included spike and recovery experiments, which produced parallel curves, indicating that AF constituents did not interfere with antigen–antibody binding in this assay. Immunoassays were carried out according to the manufacturer's recommendations. AF samples were incubated in duplicate wells of the microtiter plates that were pre-coated with an antibody specific for the analyte (fragment Bb). During this incubation, the analyte present in the standards or AF samples was bound by the immobilized antibodies in the respective assay plates. After repeated washing and aspiration to remove all unbound substances, an enzyme-linked polyclonal antibody specific for the analyte was added to the wells of the assay plates. Unbound enzyme conjugate was removed by repeated washing, and a substrate solution was added to the wells of the assay plates. Color developed in proportion to the amount of the analyte bound in the initial step, and this development was stopped with the addition of an acid solution. The intensity of color was read using a programmable spectrophotometer (SpectraMax M2, Molecular Devices, Sunnyvale, CA). The concentrations of fragment Bb in AF samples were determined by interpolation from individual standard curves. The calculated inter- and intra-assay coefficients of variation for fragment Bb immunoassays in our laboratory were 3.1% and 2.4%, respectively, and the sensitivity was 0.015 μg/ml.
Statistical analysis
Shapiro–Wilk and Kolmogorov–Smirnov tests were used to test for normal distribution of the data. Since AF fragment Bb concentrations were not normally distributed, non-parametric tests were used for analyses. Correlations between continuous variables were assessed by the Spearman's rank correlation test. Comparisons between proportions were performed with chi-square or Fisher's exact tests. Kruskal–Wallis with post-hoc test (Mann–Whitney U tests) was used for continuous variables. Multiple linear regression analysis was performed to determine the relationship between AF concentration of fragment Bb and the following variables: maternal age, gestational age at amniocentesis and sample storage time. Among patients with PTL and intact membranes, receiver–operating characteristic (ROC) curve analysis was performed to determine cutoffs for the AF fragment Bb concentrations for the identification of patients who had IAI. A p-value of <0.05 was considered statistically significant. The statistical analyses were performed with SPSS package version 12 (SPSS, Chicago, IL).
Results
Fragment Bb was detected in all the AF samples tested (n = 544). The demographic and clinical characteristics of patients with a normal pregnancy (mid-trimester, term not in labor and term in labor), with spontaneous PTL and intact membranes and with preterm PROM are displayed in Tables , respectively. The median gestational age at amniocentesis was significantly lower among patients with PTL with IAI than that of the other two subgroups of PTL (), and among patients with preterm PROM with IAI compared with those with preterm PROM without IAI ().
Table I. Demographic and clinical characteristics of patients with a normal pregnancy in the mid-trimester and those at term, with and without spontaneous labor.
Table II. Demographic and clinical characteristics of patients presenting with spontaneous preterm labor (PTL) and intact membranes.
Table III. Demographic and clinical characteristics of patients presenting with preterm prelabor rupture of membranes (preterm PROM).
Amniotic fluid concentration of fragment Bb in normal pregnancies
Women with a normal pregnancy at term not in labor had a higher median AF concentration of fragment Bb than those in the mid-trimester [2.42 μg/ml, interquartile range (IQR) 1.78–3.22 vs. 1.64 μg/ml, IQR 1.06–3.49; p < 0.001] (). Among women at term, the median AF fragment Bb concentration did not differ significantly between patients with spontaneous labor and those without labor (term in labor: 2.47 μg/ml, IQR 1.86–3.22 vs. term not in labor: 2.42 μg/ml, IQR 1.78–3.22; p = 0.97) ().
Figure 1. Amniotic fluid concentration of fragment Bb in normal pregnancies in the mid-trimester and at term, with and without spontaneous labor: The median amniotic fluid concentration of fragment Bb was higher in women at term not in labor than in those in the mid-trimester [term not in labor: 2.42 μg/ml, IQR 1.78–3.22 vs. mid-trimester: 1.64 μg/ml, IQR 1.06–3.49; p < 0.001]. Among women at term, the median amniotic fluid fragment Bb concentration did not differ between patients with spontaneous labor and those not in labor (term in labor: 2.47 μg/ml, IQR 1.86–3.22; p = 0.97).
![Figure 1. Amniotic fluid concentration of fragment Bb in normal pregnancies in the mid-trimester and at term, with and without spontaneous labor: The median amniotic fluid concentration of fragment Bb was higher in women at term not in labor than in those in the mid-trimester [term not in labor: 2.42 μg/ml, IQR 1.78–3.22 vs. mid-trimester: 1.64 μg/ml, IQR 1.06–3.49; p < 0.001]. Among women at term, the median amniotic fluid fragment Bb concentration did not differ between patients with spontaneous labor and those not in labor (term in labor: 2.47 μg/ml, IQR 1.86–3.22; p = 0.97).](/cms/asset/5f5f29e7-1810-45eb-87c7-fc417f8251e4/ijmf_a_399638_f0001_b.gif)
Amniotic fluid concentration of fragment Bb in women with spontaneous preterm labor and intact membranes and those with preterm prelabor rupture of the membranes
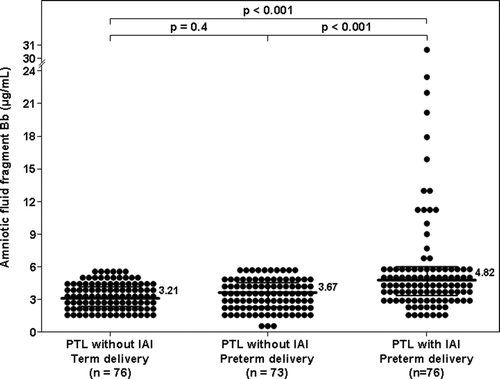
Among women with PTL, the median AF concentration of fragment Bb was higher in patients with IAI than in those without IAI who delivered preterm (4.82 μg/ml, IQR 3.32–6.08 vs. 3.67 μg/ml, IQR 2.35–4.57; p < 0.001) or at term (3.21 μg/ml, IQR 2.39–4.16; p < 0.001) (). Among women with PTL without IAI, there was no significant difference in the median AF concentration of fragment Bb between those who delivered preterm and the ones who delivered at term (p = 0.4; ).
Figure 2. Amniotic fluid concentration of fragment Bb in women with spontaneous preterm labor and intact membranes: The median amniotic fluid concentration of fragment Bb was higher in patients with IAI than in those without IAI who delivered preterm (PTL 4.82 μg/ml, IQR 3.32–6.08 vs. 3.67 μg/ml, IQR 2.35–4.57; p < 0.001), as well as than that of those who delivered at term (3.21 μg/ml, IQR 2.39–4.16; p < 0.001). Among women with PTL without IAI, there was no significant difference in the median amniotic fluid fragment Bb concentration between patients who delivered preterm and those who delivered at term (p = 0.4).
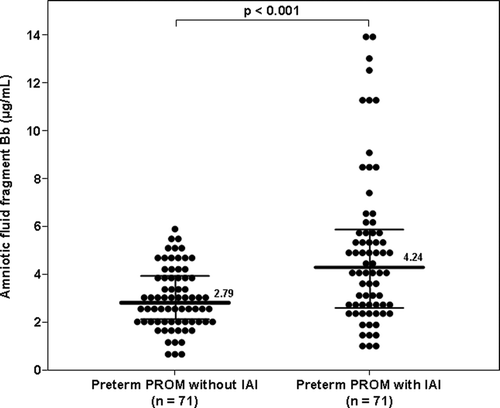
Among women with preterm PROM, the median AF concentration of fragment Bb was higher in patients with IAI than in those without IAI (4.24 μg/ml, IQR 2.58–5.79 vs. 2.79 μg/ml, IQR 2.09–3.89; p < 0.001) ().
Figure 3. Amniotic fluid concentration of fragment Bb in women with preterm prelabor rupture of membranes (preterm PROM): The median amniotic fluid concentration of fragment Bb was higher in patients with IAI than in those without IAI (4.24 μg/ml, IQR 2.58–5.79 vs. 2.79 μg/ml, IQR 2.09–3.89; p < 0.001).
Among patients with PTL and those with preterm PROM, AF fragment Bb concentrations were positively correlated with AF WBC count (Spearman rho coefficient: r = 0.27, p < 0.001) and IL-6 concentrations (r = 0.4, p < 0.001) and were negatively correlated with AF glucose concentrations (r = −0.14, p = 0.006).
To examine the association between AF fragment Bb concentrations, IAI and possible confounding factors, a multiple regression analysis was performed adjusting for maternal age, gestational age at amniocentesis and sample storage time. The model demonstrated that the presence of IAI was independently associated with increased AF fragment Bb concentrations (p < 0.001).
The ROC curve of AF fragment Bb concentration for the identification of IAI among patients with PTL with intact membranes is displayed in (area under the curve 0.72, p < 0.001). An optimized cutoff value (sensitivity and specificity sharing an equal importance) of AF fragment Bb concentration of ≥4.31 μg/ml in patients with PTL and intact membranes had a sensitivity of 61.8% and a specificity of 75.8% for identification of IAI (). Because of the relative poor diagnostic performance of this cutoff, a cutoff value of ≥6.2 μg/ml, which has a specificity of 100%, however, a low sensitivity of 23.7% for the detection of IAI in patients with PTL and intact membranes, has been chosen for the definition of an elevated fragment Bb concentration ().
Figure 4. Receiver–operating characteristic (ROC) curve of amniotic fluid fragment Bb concentration of patients with PTL and intact membranes for the identification of IAI (n = 225, area under the curve 0.72, p < 0.001).
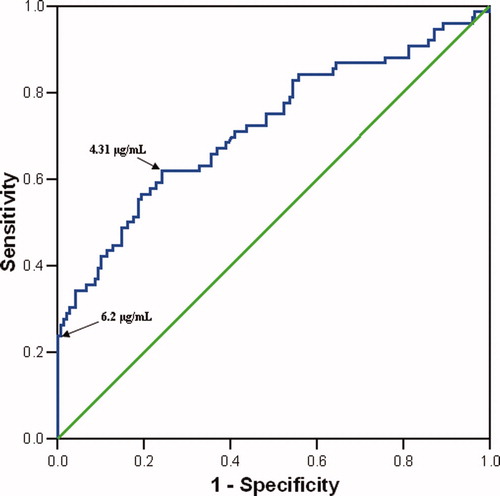
Table IV. Diagnostic indices and likelihood ratios of amniotic fluid fragment Bb concentration for the detection of intra-amniotic infection/inflammation in patients presenting with spontaneous preterm labor with intact membranes (n = 225).
When this cutoff was tested on the entire study population, we found that 21.8% of the women with IAI (32/147) had an elevated fragment Bb concentration (≥6.2 μg/ml), whereas none of those without IAI reached the cutoff (0/397, p < 0.001).
Amniotic fluid concentration of fragment Bb and placental histopathologic findings
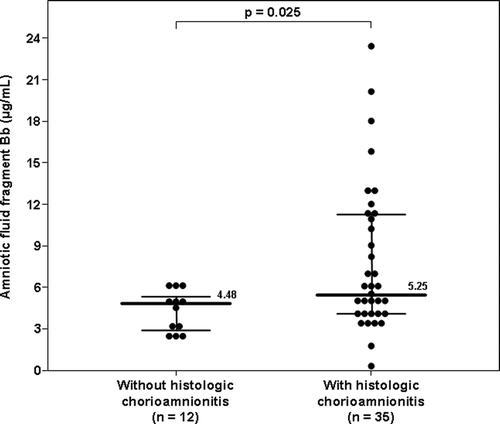
Among patients with spontaneous PTL and intact membranes with IAI, 79% (60/76) delivered within 72 h of amniocentesis, and placental histopathologic diagnoses were available in 78% (47/60) of these individuals. The median AF concentration of fragment Bb in patients with histologic chorioamnionitis was higher than that of women without it (histologic chorioamnionitis: 5.25 μg/ml, IQR 3.97–11.27 vs. no histologic chorioamnionitis: 4.48 μg/ml, IQR 2.80–5.29; p = 0.025) (). Moreover, none of the patients without histologic chorioamnionitis (0/12) had an elevated fragment Bb concentration (≥6.2 μg/ml), whereas 40% (14/35) of those with histologic chorioamnionitis had an elevated AF fragment Bb concentration (Fisher's exact, p = 0.009).
Figure 5. Amniotic fluid concentration of fragment Bb in women with spontaneous PTL, intact membranes and IAI who delivered within 72 h of amniocentesis and have placental histopathologic diagonsis: Among women with IAI, the median amniotic fluid concentration of fragment Bb was higher in patients with histologic chorioamnionitis than in those without (5.25 μg/ml, IQR 3.97–11.27 vs. 4.48 μg/ml, IQR 2.80–5.29; p = 0.025).
Comment
Principal findings of this study
(1) Fragment Bb was detectable in all AF samples and seems to be a physiologic constituent of the AF; (2) patients with IAI, regardless of the membranes status, had a higher median AF concentration of fragment Bb than that of women without IAI; (3) AF fragment Bb concentrations increase with advancing gestation and do not change with labor at term; and (4) elevated AF fragment Bb concentrations were independently associated with IAI.
What is fragment Bb?
The complement system, an important component of the innate immunity, plays a pivotal role in the process of recognition of foreign antigens and pathogens. In addition, the complement system mediates the inflammatory response elicited against infection and has a role in activating the adaptive immune system Citation[30-32]. More than 30 proteins and cell membrane receptors encompass the complement system, which can be activated by three different pathways: the ‘classical’, the ‘lectin’ and the ‘alternative’. Despite the different pathways, all three converge at the point of C3 convertase formation Citation[33]. C3 convertase cleaves C3 to C3a and C3b, and the latter participates in the formation of the C5 convertase, which cleaves C5 to C5a and C5b. C3a and C5a, termed anaphylatoxins, are pleiotropic inflammatory mediators Citation[32].
The classical pathway is initiated by the binding of C1q to antigen–antibody complexes, whereas the lectin pathway begins with the binding of mannose-binding lectin to sugars present on the bacterial cell wall. Increased concentrations of C4 can be regarded as a marker of complement activation by either the classical or lectin pathways. In contrast, the alternative pathway is capable of auto-activation by a process termed ‘tickover’ of C3 Citation[34]. Tickover occurs spontaneously in the absence of a recognizable trigger at a rate of approximately 1% of total C3 per hour, generating a conformationally altered C3, namely C3(H2O) Citation[34]. The latter is capable of binding factor B, a unique component of the alternative pathway, which can then be cleaved by factor D at a single Arg-Lys bond Citation[51-53], generating two unequal fragments, an N-terminal Ba fragment and a C-terminal Bb fragment that carries the active site of factor B Citation[54]. Fragment Bb remains associated with C3(H2O) and the C3(H2O)Bb complex, through its own serine protease domain, can cleave additional C3 molecules, generating C3b that associates with factor B to generate more C3-convertase Citation[32]. The alternative pathway can also be initiated as an ‘amplification loop’ when fixed C3b, generated by the classical or lectin pathways, binds factor B resulting in conformational changes in factor B that allow factor D to cleave it similarly to the tickover process Citation[31]. Thus, activation of the alternative pathway is characterized by increased production of fragment Bb.
The complement system in normal and complicated pregnancies
The complement system is an effector arm of the innate immune system, which is important in the host defense against infections. An excessive or inappropriate activation of the complement system has been implicated in the pathophysiology of many disorders such as rheumatoid arthritis Citation[55],Citation[56], systemic lupus erythematosus Citation[57] and stroke Citation[58],Citation[59]. In addition, perturbation of the complement system homeostasis following exposure of healthy volunteers to stressful conditions has been reported Citation[60-62].
Pregnancy, characterized by physiologic activation of the complement system in the maternal blood, has been proposed to be a compensatory mechanism aimed to protect the host against infection Citation[35]. However, several pregnancy complications, such as spontaneous pregnancy losses Citation[36-41], preeclampsia Citation[42],Citation[43],Citation[63], pyelonephritis Citation[44], fetal death Citation[45] and preterm birth Citation[13] have been associated with excessive systemic maternal complement activation. Components of the complement system have been detected in placenta Citation[64-66], chorioamniotic membranes Citation[67-69], fetal tissues (i.e., liver, spleen and thymus) Citation[64],Citation[70-73] cord blood Citation[74-80] and AF Citation[46],Citation[77],Citation[81],Citation[82].
In 1988, Stabile et al. Citation[77] measured complement factors (C3, C4, C5, Factor B, H and I) in maternal and fetal circulations as well as in AF obtained between 15 and 28 weeks of gestation from 55 women with a retrospectively defined normal pregnancy, who underwent diagnostic fetoscopy or cordocentesis for the exclusion of hematological disorders or chromosomal defects. Concentrations of these proteins were 10 times higher in the maternal than that in the fetal circulation, which, in turn, were 10 times higher than that in the AF. Fetal concentrations of C3, C4 and factor H and AF concentrations of C3 and factor B increased with advancing gestation, but this was not observed in maternal blood. Thus, the authors concluded that the fetus is independently synthesizing proteins of the complement system Citation[77].
Increased activation of components of the complement system has been reported in the AF from patients with PTL Citation[46],Citation[83]. Studying the AF of 104 women with PTL with and without intra-amniotic infection, Elimian et al. Citation[46] found a significantly higher median C3 concentration in the culture-positive group. In addition, our group reported that among 129 patients with PTL, those with intra-amniotic infection had higher median AF concentrations of the anaphylatoxins, C3a, C4a and C5a Citation[83].
Although there are numerous reports regarding activation of the complement system in the maternal and fetal compartments during normal gestation and complications of pregnancy Citation[13],Citation[35-42],Citation[44-46],Citation[81],Citation[83], data concerning fragment Bb during pregnancy are scarce Citation[43],Citation[47],Citation[84]. Recently, Lynch et al. Citation[43],Citation[47] were the first to report the association between elevated maternal plasma fragment Bb concentrations and pregnancy complications. Elevated maternal plasma concentrations of fragment Bb in early pregnancy (before 20 weeks of gestation) were associated with an increased risk for later development of preeclampsia Citation[43]. In a subsequent report, an association between increased maternal plasma concentrations of fragment Bb in early pregnancy and a later spontaneous preterm birth before 34 weeks has been proposed Citation[47].
Fragment Bb during normal pregnancy
The study presented herein reports, for the first time, the presence of fragment Bb in the AF. Fragment Bb, an activation product of the alternative pathway, was detected in all samples included in this study, suggesting that it is a physiologic constituent of the AF. In addition, its concentration in AF increases with gestational age and does not change during labor at term or preterm, suggesting increasing physiologic activation of the complement system through the alternative pathway with advancing gestation, but without further activation during labor. These results are consistent with the findings of Stabile et al. Citation[77], who reported higher AF concentration of factor B with advancing gestational age, and with a previous study from our group Citation[83] that did not find an association between spontaneous labor at term and changes in the AF concentration of other complement activation products (C3a, C4a and C5a).
Spontaneous labor at term is regarded as an inflammatory process Citation[85]. Supporting this view are its associations with inflammatory cells infiltration in the cervix Citation[86-88], myometrium Citation[88],Citation[89] and chorioamniotic membranes Citation[88],Citation[90] and increased production of proinflammatory cytokines Citation[85],Citation[91] (i.e., IL-1βCitation[88],Citation[92-94], IL-6 Citation[88],Citation[94],Citation[95], tumor necrosis factor αCitation[93],Citation[94] and IL-8 Citation[88],Citation[95-97]) and chemokines Citation[85],Citation[91] (i.e., growth regulated oncogene αCitation[98], granulocyte colony stimulating factor Citation[95], monocyte chemotactic protein 1 Citation[99-101]). However, the results presented herein and those reported by Soto et al. Citation[83] suggest that spontaneous labor at term is not associated with activation of the complement system in the AF.
Fragment Bb in intra-amniotic infection/inflammation
The finding that IAI is associated with a higher median AF concentration of fragment Bb in patients with intact and ruptured membranes is novel and supports the concept of activation of the complement system as part of the fetal inflammatory response to microbial invasion of the amniotic cavity Citation[18],Citation[20]. Evidence in support of this view comes from a report by Hogasen et al. Citation[84], who analyzed the complement activation products in cord blood of neonates born after preterm PROM, and reported a significantly increased concentration of Bb in the cord blood of these neonates when compared with healthy controls. Moreover, higher AF concentration of C3 Citation[46], C3a, C4a and C5a Citation[83] have been found in women with PTL and microbial invasion of the amniotic cavity compared with those with a negative AF culture. Of interest, patients with PTL and IAI who delivered within 72 h of amniocentesis and had histologic evidence of placental inflammation had a higher median AF fragment Bb concentration than women without it. Most cases (29/35) with a positive placental pathology for inflammation showed evidence of fetal involvement manifested as funisitis, which is considered the histological counterpart of the FIRS Citation[50]. Moreover, the association between elevated AF fragment Bb concentrations and IAI is independent of confounding factors, such as gestational age at amniocentesis.
An interesting finding of the present study is that, only women with IAI had an elevated AF fragment Bb concentration (≥6.2 μg/ml). All women with a normal pregnancy (mid-trimester, term with or without labor) and those with PTL who delivered preterm or at term and those with preterm PROM without IAI had AF fragment Bb concentrations below 6.2 μg/ml. Thus, although a cutoff of ≥6.2 μg/ml has a very low sensitivity for the identification of IAI, its specificity is as high as 100%. In other words, if a fragment Bb concentration of ≥6.2 μg/ml is found in the AF, the chance of intra-amniotic infection and/or inflammation is 100%. Moreover, among patients with PTL and intact membranes with IAI who delivered within 72 h from amniocentesis, only those with histologic chorioamnionitis had an elevated AF fragment Bb concentration (). This suggests that AF fragment Bb concentrations above ≥6.2 μg/ml may be almost exclusively attributed to intra-amniotic infection and/or inflammation with histologic chorioamnionitis.
The FIRS, originally defined in fetuses with PTL and preterm PROM by an elevated fetal plasma IL-6 concentration Citation[18], is characterized by systemic activation of the fetal innate immune system Citation[18],Citation[20]. In the presence of invading microorganisms, an inflammatory process is initiated to deliver cells and molecules to suppress the infection, generate a barrier to the spread of the infection and support repair of the injured tissue Citation[32]. This process involves (1) the attraction of cells to the site of injury, including macrophages, neutrophils and lymphocytes; and (2) the release of antimicrobial peptides, cytokines, chemokines and other inflammatory mediators such as prostaglandins, leukotrienes and complement Citation[32]. Some of these molecules are capable of changing the state of activation of macrophages and neutrophils. Indeed, fragment Bb has the ability to induce rapid macrophage spreading Citation[102] and is an effective inhibitor of macrophage migration in a dose-dependent manner Citation[103]. All the while, the complement component C5 has an opposite effect, causing enhancement of macrophage migration where the outcome of the reaction will depend on the predominant peptide Citation[103]. Thus, a role for the alternative pathway of the complement system in the localization of mononuclear phagocytes to areas of inflammation has been suggested Citation[102].
In conclusion, our study demonstrates that fragment Bb can be detected in the AF of all pregnant women, that intra-amniotic infection and/or inflammation is associated with increased AF concentration of fragment Bb, regardless of the membranes status, and that the AF fragment Bb concentration does not change with term or PTL. Collectively, these findings suggest that activation of the alternative pathway of the complement system may be part of the fetal innate immune response to IAI.
Acknowledgment
This research was supported by the Intramural Research Program of the Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH, DHHS.
References
- Romero R, Mazor M, Munoz H, Gomez R, Galasso M, Sherer D M. The preterm labor syndrome. Ann NY Acad Sci 1994; 734: 414–429
- Romero R, Espinoza J, Kusanovic J P, Gotsch F, Hassan S, Erez O, Chaiworapongsa T, Mazor M. The preterm parturition syndrome. BJOG 2006; 113(Suppl 3)17–42
- Naeye R L, Ross S M. Amniotic fluid infection syndrome. Clin Obstet Gynaecol 1982; 9: 593–607
- Minkoff H. Prematurity: infection as an etiologic factor. Obstet Gynecol 1983; 62: 137–144
- Romero R, Mazor M, Wu Y K, Sirtori M, Oyarzun E, Mitchell M D, Hobbins J C. Infection in the pathogenesis of preterm labor. Semin Perinatol 1988; 12: 262–279
- Romero R, Sirtori M, Oyarzun E, Avila C, Mazor M, Callahan R, Sabo V, Athanassiadis A P, Hobbins J C. Infection and labor. V. Prevalence, microbiology, and clinical significance of intraamniotic infection in women with preterm labor and intact membranes. Am J Obstet Gynecol 1989; 161: 817–824
- Ledger W J. Infection and premature labor. Am J Perinatol 1989; 6: 234–236
- Gibbs R S, Romero R, Hillier S L, Eschenbach D A, Sweet R L. A review of premature birth and subclinical infection. Am J Obstet Gynecol 1992; 166: 1515–1528
- Brocklehurst P. Infection and preterm delivery. BMJ 1999; 318: 548–549
- Goldenberg R L, Hauth J C, Andrews W W. Intrauterine infection and preterm delivery. N Engl J Med 2000; 342: 1500–1507
- Goncalves L F, Chaiworapongsa T, Romero R. Intrauterine infection and prematurity. Ment Retard Dev Disabil Res Rev 2002; 8: 3–13
- Hirsch E, Wang H. The molecular pathophysiology of bacterially induced preterm labor: insights from the murine model. J Soc Gynecol Investig 2005; 12: 145–155
- Soto E, Romero R, Richani K, Espinoza J, Nien J K, Chaiworapongsa T, Santolaya-Forgas J, Edwin S S, Mazor M. Anaphylatoxins in preterm and term labor. J Perinat Med 2005; 33: 306–313
- Yoon B H, Romero R, Kim C J, Jun J K, Gomez R, Choi J H, Syn H C. Amniotic fluid interleukin-6: a sensitive test for antenatal diagnosis of acute inflammatory lesions of preterm placenta and prediction of perinatal morbidity. Am J Obstet Gynecol 1995; 172: 960–970
- Wenstrom K D, Andrews W W, Hauth J C, Goldenberg R L, DuBard M B, Cliver S P. Elevated second-trimester amniotic fluid interleukin-6 levels predict preterm delivery. Am J Obstet Gynecol 1998; 178: 546–550
- Yoon B H, Romero R, Moon J B, Shim S S, Kim M, Kim G, Jun J K. Clinical significance of intra-amniotic inflammation in patients with preterm labor and intact membranes. Am J Obstet Gynecol 2001; 185: 1130–1136
- Yoon B H, Oh S Y, Romero R, Shim S S, Han S Y, Park J S, Jun J K. An elevated amniotic fluid matrix metalloproteinase-8 level at the time of mid-trimester genetic amniocentesis is a risk factor for spontaneous preterm delivery. Am J Obstet Gynecol 2001; 185: 1162–1167
- Gomez R, Romero R, Ghezzi F, Yoon B H, Mazor M, Berry S M. The fetal inflammatory response syndrome. Am J Obstet Gynecol 1998; 179: 194–202
- Romero R, Gomez R, Ghezzi F, Yoon B H, Mazor M, Edwin S S, Berry S M. A fetal systemic inflammatory response is followed by the spontaneous onset of preterm parturition. Am J Obstet Gynecol 1998; 179: 186–193
- Gotsch F, Romero R, Kusanovic J P, Mazaki-Tovi S, Pineles B L, Erez O, Espinoza J, Hassan S S. The fetal inflammatory response syndrome. Clin Obstet Gynecol 2007; 50: 652–683
- Dammann O, Leviton A. Maternal intrauterine infection, cytokines, and brain damage in the preterm newborn. Pediatr Res 1997; 42: 1–8
- Yoon B H, Romero R, Jun J K, Park K H, Park J D, Ghezzi F, Kim B I. Amniotic fluid cytokines (interleukin-6, tumor necrosis factor-alpha, interleukin-1 beta, and interleukin-8) and the risk for the development of bronchopulmonary dysplasia. Am J Obstet Gynecol 1997; 177: 825–830
- Yoon B H, Jun J K, Romero R, Park K H, Gomez R, Choi J H, Kim I O. Amniotic fluid inflammatory cytokines (interleukin-6, interleukin-1beta, and tumor necrosis factor-alpha), neonatal brain white matter lesions, and cerebral palsy. Am J Obstet Gynecol 1997; 177: 19–26
- Yoon B H, Romero R, Park J S, Kim C J, Kim S H, Choi J H, Han T R. Fetal exposure to an intra-amniotic inflammation and the development of cerebral palsy at the age of three years. Am J Obstet Gynecol 2000; 182: 675–681
- Dammann O, Leviton A. Role of the fetus in perinatal infection and neonatal brain damage. Curr Opin Pediatr 2000; 12: 99–104
- Gibbs R S. The relationship between infections and adverse pregnancy outcomes: an overview. Ann Periodontol 2001; 6: 153–163
- Patrick L A, Smith G N. Proinflammatory cytokines: a link between chorioamnionitis and fetal brain injury. J Obstet Gynaecol Can 2002; 24: 705–709
- Hagberg H, Mallard C, Jacobsson B. Role of cytokines in preterm labour and brain injury. BJOG 2005; 112(Suppl 1)16–18
- Bashiri A, Burstein E, Mazor M. Cerebral palsy and fetal inflammatory response syndrome: a review. J Perinat Med 2006; 34: 5–12
- Holers V M. The complement system as a therapeutic target in autoimmunity. Clin Immunol 2003; 107: 140–151
- Thurman J M, Holers V M. The central role of the alternative complement pathway in human disease. J Immunol 2006; 176: 1305–1310
- Murphy K, Travers P, Walport M. Innate immunity 2008; Seventh: 39–103
- Walport M J. Complement. First of two parts. N Engl J Med 2001; 344: 1058–1066
- Muller-Eberhard H J. Molecular organization and function of the complement system. Annu Rev Biochem 1988; 57: 321–47
- Richani K, Soto E, Romero R, Espinoza J, Chaiworapongsa T, Nien J K, Edwin S, Kim Y M, Hong J S, Mazor M. Normal pregnancy is characterized by systemic activation of the complement system. J Matern Fetal Neonatal Med 2005; 17: 239–245
- Tichenor J R, Bledsoe L B, Opsahl M S, Cunningham D S. Activation of complement in humans with a first-trimester pregnancy loss. Gynecol Obstet Invest 1995; 39: 79–82
- Mellor A L, Sivakumar J, Chandler P, Smith K, Molina H, Mao D, Munn D H. Prevention of T cell-driven complement activation and inflammation by tryptophan catabolism during pregnancy. Nat Immunol 2001; 2: 64–68
- Holers V M, Girardi G, Mo L, Guthridge J M, Molina H, Pierangeli S S, Espinola R, Xiaowei L E, Mao D, Vialpando C G, et al. Complement C3 activation is required for antiphospholipid antibody-induced fetal loss. J Exp Med 2002; 195: 211–220
- Caucheteux S M, Kanellopoulos-Langevin C, Ojcius D M. At the innate frontiers between mother and fetus: linking abortion with complement activation. Immunity 2003; 18: 169–172
- Girardi G, Salmon J B. The role of complement in pregnancy and fetal loss. Autoimmunity 2003; 36: 19–26
- Girardi G. Complement inhibition keeps mothers calm and avoids fetal rejection. Immunol Invest 2008; 37: 645–659
- Richani K, Soto E, Romero R, Espinoza J, Chaiworapongsa T, Nien J K, Edwin S, Kim Y M, Hong J S, Goncalves L, et al. Preeclampsia and SGA differ in the maternal plasma complememt split products profile. J Soc Gynecol Investig 2005; 12(Suppl 2)148A
- Lynch A M, Murphy J R, Byers T, Gibbs R S, Neville M C, Giclas P C, Salmon J E, Holers V M. Alternative complement pathway activation fragment Bb in early pregnancy as a predictor of preeclampsia. Am J Obstet Gynecol 2008; 198: 385–389
- Soto E, Richani K, Romero R, Espinoza J, Chaiworapongsa T, Nien J K, Edwin S, Kim Y M, Hong J S, Goncalves L, et al. Increased concentration of the complement split product C5a in acute pyelonephritis during pregnancy. J Matern Fetal Neonatal Med 2005; 17: 247–252
- Richani K, Romero R, Soto E, Espinoza J, Nien J K, Chaiworapongsa T, Refuerzo J, Blackwell S, Edwin S S, Santolaya-Forgas J, et al. Unexplained intrauterine fetal death is accompanied by activation of complement. J Perinat Med 2005; 33: 296–305
- Elimian A, Figueroa R, Canterino J, Verma U, guero-Rosenfeld M, Tejani N. Amniotic fluid complement C3 as a marker of intra-amniotic infection. Obstet Gynecol 1998; 92: 72–76
- Lynch A M, Gibbs R S, Murphy J R, Byers T, Neville M C, Giclas P C, Salmon J E, Van Hecke T M, Holers V M. Complement activation fragment Bb in early pregnancy and spontaneous preterm birth. Am J Obstet Gynecol 2008; 199: 354–358
- Alexander G R, Kogan M, Martin J, Papiernik E. What are the fetal growth patterns of singletons, twins, and triplets in the United States?. Clin Obstet Gynecol 1998; 41: 114–125
- Gonzalez R P, Gomez R M, Castro R S, Nien J K, Merino P O, Etchegaray A B, Carstens M R, Medina L H, Viviani P G, Rojas I T. [A national birth weight distribution curve according to gestational age in Chile from 1993 to 2000]. Rev Med Chil 2004; 132: 1155–1165
- Pacora P, Chaiworapongsa T, Maymon E, Kim Y M, Gomez R, Yoon B H, Ghezzi F, Berry S M, Qureshi F, Jacques S M, et al. Funisitis and chorionic vasculitis: the histological counterpart of the fetal inflammatory response syndrome. J Matern Fetal Neonatal Med 2002; 11: 18–25
- Lesavre P H, Hugli T E, Esser A F, Muller-Eberhard H J. The alternative pathway C3/C5 convertase: chemical basis of factor B activation. J Immunol 1979; 123: 529–534
- Kerr M A. Limited proteolysis of complement components C2 and factor B. Structural analogy and limited sequence homology. Biochem J 1979; 183: 615–622
- Niemann M A, Volanakis J E, Mole J E. Amino-terminal sequence of human factor B of the alternative complement pathway and its cleavage fragments, Ba and Bb. Biochemistry 1980; 19: 1576–1583
- Gotze O, Muller-Eberhard H J. The C3-activator system: an alternate pathway of complement activation. J Exp Med 1971; 134: 90s–108s
- Nakagawa K, Sakiyama H, Tsuchida T, Yamaguchi K, Toyoguchi T, Masuda R, Moriya H. Complement C1s activation in degenerating articular cartilage of rheumatoid arthritis patients: immunohistochemical studies with an active form specific antibody. Ann Rheum Dis 1999; 58: 175–181
- Neumann E, Barnum S R, Tarner I H, Echols J, Fleck M, Judex M, Kullmann F, Mountz J D, Scholmerich J, Gay S, et al. Local production of complement proteins in rheumatoid arthritis synovium. Arthritis Rheum 2002; 46: 934–945
- Walport M J. Complement and systemic lupus erythematosus. Arthritis Res 2002; 4(Suppl 3)S279–S293
- del Zoppo G J. In stroke, complement will get you nowhere. Nat Med 1999; 5: 995–996
- Huang J, Kim L J, Mealey R, Marsh H C, Jr, Zhang Y, Tenner A J, Connolly E S, Jr, Pinsky D J. Neuronal protection in stroke by an sLex-glycosylated complement inhibitory protein. Science 1999; 285: 595–599
- Endresen I M, Relling G B, Tonder O, Myking O, Walther B T, Ursin H. Brief uncontrollable stress and psychological parameters influence human plasma concentrations of IgM and complement component C3. Behav Med 1991; 17: 167–176
- Maes M, Hendriks D, Van G A, Demedts P, Wauters A, Neels H, Janca A, Scharpe S. Effects of psychological stress on serum immunoglobulin, complement and acute phase protein concentrations in normal volunteers. Psychoneuroendocrinology 1997; 22: 397–409
- Burns V E, Edwards K M, Ring C, Drayson M, Carroll D. Complement cascade activation after an acute psychological stress task. Psychosom Med 2008; 70: 387–396
- Haeger M, Bengtson A, Karlsson K, Heideman M. Complement activation and anaphylatoxin (C3a and C5a) formation in preeclampsia and by amniotic fluid. Obstet Gynecol 1989; 73: 551–556
- Kohler P F. Maturation of the human complement system. I. Onset time and sites of fetal C1q, C4, C3, and C5 synthesis. J Clin Invest 1973; 52: 671–677
- Holmes C H, Simpson K L. Complement and pregnancy: new insights into the immunobiology of the fetomaternal relationship. Baillieres Clin Obstet Gynaecol 1992; 6: 439–460
- Goldberg M, Luknar-Gabor N, Keidar R, Katz Y. Synthesis of complement proteins in the human chorion is differentially regulated by cytokines. Mol Immunol 2007; 44: 1737–1742
- Holmes C H, Simpson K L, Okada H, Okada N, Wainwright S D, Purcell D F, Houlihan J M. Complement regulatory proteins at the feto-maternal interface during human placental development: distribution of CD59 by comparison with membrane cofactor protein (CD46) and decay accelerating factor (CD55). Eur J Immunol 1992; 22: 1579–1585
- Vanderpuye O A, Labarrere C A, McIntyre J A. Expression of CD59, a human complement system regulatory protein, in extraembryonic membranes. Int Arch Allergy Immunol 1993; 101: 376–384
- Richani K, Soto E, Romero R, Han Y, Pineles B, Kim Y M, C E, Yoon B H, Kusanovic J, Kim C J. Decreased mRNA expression of complement regulatory proteins in chorioamnionitis. Am J Obstet Gynecol 2006; 195(Suppl 1)S71
- Adinolfi M, Gardner B, Wood C B. Ontogenesis of two components of human complement: beta1E and beta1C-1A globulins. Nature 1968; 219: 189–191
- Colten H R, Gordon J M, Borsos T, Rapp H J. Synthesis of the first component of human complement in vitro. J Exp Med 1968; 128: 595–604
- Gitlin D, Biasucci A. Development of gamma G, gamma A, gamma M, beta IC-beta IA, C 1 esterase inhibitor, ceruloplasmin, transferrin, hemopexin, haptoglobin, fibrinogen, plasminogen, alpha 1-antitrypsin, orosomucoid, beta-lipoprotein, alpha 2-macroglobulin, and prealbumin in the human conceptus. J Clin Invest 1969; 48: 1433–1446
- Colten H R. Ontogeny of the human complement system: in vitro biosynthesis of individual complement components by fetal tissues. J Clin Invest 1972; 51: 725–730
- Fireman P, Zuchowski D A, Taylor P M. Development of human complement system. J Immunol 1969; 103: 25–31
- Ballow M, Fang F, Good R A, Day N K. Developmental aspects of complement components in the newborn. The presence of complement components and C3 proactivator (properdin factor B) in human colostrum. Clin Exp Immunol 1974; 18: 257–266
- Miyano A, Nakayama M, Fujita T, Kitajima H, Imai S, Shimizu A. Complement activation in fetuses: assessment by the levels of complement components and split products in cord blood. Diagn Clin Immunol 1987; 5: 86–90
- Stabile I, Nicolaides K H, Bach A, Teisner B, Rodeck C, Westergaard J G, Grudzinskas J G. Complement factors in fetal and maternal blood and amniotic fluid during the second trimester of normal pregnancy. Br J Obstet Gynaecol 1988; 95: 281–285
- Zilow G, Zilow E P, Burger R, Linderkamp O. Complement activation in newborn infants with early onset infection. Pediatr Res 1993; 34: 199–203
- Enskog A, Bengtsson A, Bengtson J P, Heideman M, Andreasson S, Larsson L. Complement anaphylatoxin C3a and C5a formation in premature children with respiratory distress. Eur J Pediatr 1996; 155: 41–45
- Sonntag J, Brandenburg U, Polzehl D, Strauss E, Vogel M, Dudenhausen J W, Obladen M. Complement system in healthy term newborns: reference values in umbilical cord blood. Pediatr Dev Pathol 1998; 1: 131–135
- Sharma A, Prabhakar P, Sharma D P, Jayasinghe R G. Immunoglobulin and C3 levels in normal human amniotic fluid. West Indian Med J 1983; 32: 140–146
- Huffaker J, Witkin S S, Cutler L, Druzin M L, Ledger W J. Total complement activity in maternal sera, amniotic fluids and cord sera in women with premature labor, premature rupture of membranes or chorioamnionitis. Surg Gynecol Obstet 1989; 168: 397–401
- Soto E, Romero R, Richani K, Chaiworapongsa T, Yoon B H, Nien J K, Edwin S, Kusanovic J P, Espinoza J. Evidence for complement activation in premature labor associated with intra-amniotic infection. Am J Obstet Gynecol 2006; 195(Suppl 1)S74
- Hogasen A K, Overlie I, Hansen T W, Abrahamsen T G, Finne P H, Hogasen K. The analysis of the complement activation product SC5 b-9 is applicable in neonates in spite of their profound C9 deficiency. J Perinat Med 2000; 28: 39–48
- Romero R, Espinoza J, Goncalves L F, Kusanovic J P, Friel L A, Nien J K. Inflammation in preterm and term labour and delivery. Semin Fetal Neonatal Med 2006; 11: 317–326
- Junqueira L C, Zugaib M, Montes G S, Toledo O M, Krisztan R M, Shigihara K M. Morphologic and histochemical evidence for the occurrence of collagenolysis and for the role of neutrophilic polymorphonuclear leukocytes during cervical dilation. Am J Obstet Gynecol 1980; 138: 273–281
- Liggins G C. Cervical ripening as an inflammatory reaction. The Cervix in pregnancy and labor, clinical and biochemical investigation, D A Ellwood, A MB Anderson. Churchill Livingstone, Edinburgh 1981; 1–9
- Osman I, Young A, Ledingham M A, Thomson A J, Jordan F, Greer I A, Norman J E. Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol Hum Reprod 2003; 9: 41–45
- Thomson A J, Telfer J F, Young A, Campbell S, Stewart C J, Cameron I T, Greer I A, Norman J E. Leukocytes infiltrate the myometrium during human parturition: further evidence that labour is an inflammatory process. Hum Reprod 1999; 14: 229–236
- Keski-Nisula L, Aalto M L, Katila M L, Kirkinen P. Intrauterine inflammation at term: a histopathologic study. Hum Pathol 2000; 31: 841–846
- Keelan J A, Blumenstein M, Helliwell R J, Sato T A, Marvin K W, Mitchell M D. Cytokines, prostaglandins and parturition–a review. Placenta 2003; 24(Suppl A)S33–S46
- Romero R, Brody D T, Oyarzun E, Mazor M, Wu Y K, Hobbins J C, Durum S K. Infection and labor. III. Interleukin-1: a signal for the onset of parturition. Am J Obstet Gynecol 1989; 160: 1117–1123
- Romero R, Mazor M, Sepulveda W, Avila C, Copeland D, Williams J. Tumor necrosis factor in preterm and term labor. Am J Obstet Gynecol 1992; 166: 1576–1587
- Opsjln S L, Wathen N C, Tingulstad S, Wiedswang G, Sundan A, Waage A, Austgulen R. Tumor necrosis factor, interleukin-1, and interleukin-6 in normal human pregnancy. Am J Obstet Gynecol 1993; 169: 397–404
- Saito S, Kasahara T, Kato Y, Ishihara Y, Ichijo M. Elevation of amniotic fluid interleukin 6 (IL-6), IL-8 and granulocyte colony stimulating factor (G-CSF) in term and preterm parturition. Cytokine 1993; 5: 81–88
- Romero R, Ceska M, Avila C, Mazor M, Behnke E, Lindley I. Neutrophil attractant/activating peptide-1/interleukin-8 in term and preterm parturition. Am J Obstet Gynecol 1991; 165: 813–820
- Osmers R G, Blaser J, Kuhn W, Tschesche H. Interleukin-8 synthesis and the onset of labor. Obstet Gynecol 1995; 86: 223–229
- Cohen J, Ghezzi F, Romero R, Ghidini A, Mazor M, Tolosa J E, Goncalves L F, Gomez R. GRO alpha in the fetomaternal and amniotic fluid compartments during pregnancy and parturition. Am J Reprod Immunol 1996; 35: 23–29
- Esplin M S, Romero R, Chaiworapongsa T, Kim Y M, Edwin S, Gomez R, Gonzalez R, Adashi E Y. Amniotic fluid levels of immunoreactive monocyte chemotactic protein-1 increase during term parturition. J Matern Fetal Neonatal Med 2003; 14: 51–56
- Esplin M S, Peltier M R, Hamblin S, Smith S, Fausett M B, Dildy G A, Branch D W, Silver R M, Adashi E Y. Monocyte chemotactic protein-1 expression is increased in human gestational tissues during term and preterm labor. Placenta 2005; 26: 661–671
- Shynlova O, Tsui P, Dorogin A, Lye S J. Monocyte chemoattractant protein-1 (CCL-2) integrates mechanical and endocrine signals that mediate term and preterm labor. J Immunol 2008; 181: 1470–1479
- Gotze O, Bianco C, Cohn Z A. The induction of macrophage spreading by factor B of the properdin system. J Exp Med 1979; 149: 372–386
- Bianco C, Gotze O, Cohn Z A. Regulation of macrophage migration by products of the complement system. Proc Natl Acad Sci USA 1979; 76: 888–891