Abstract
Objective. Visfatin, a novel adipokine with diabetogenic and immunoregulatory properties, has been implicated in the pathophysiology of insulin resistance, as well as in various acute and chronic inflammatory disorders. We have previously reported that amniotic fluid concentrations of visfatin are higher in patients with preterm labor (PTL) and intra-amniotic infection than in patients with PTL without infection. The aim of this study was to determine whether spontaneous PTL with intact membranes and intra-amniotic infection/inflammation (IAI) is associated with changes in maternal plasma circulating visfatin concentrations.
Study design. This cross-sectional study included patients in the following groups: (1) normal pregnant women (n = 123); (2) patients with an episode of PTL and intact membranes without IAI who delivered at term (n = 57); (3) PTL without IAI who delivered preterm (n = 47); and (4) PTL with IAI who delivered preterm (n = 57). Plasma visfatin concentrations were determined by ELISA. Non-parametric statistics were used for analysis.
Results. (1) PTL with IAI leading to preterm delivery was associated with a higher median maternal plasma concentration of visfatin than normal pregnancy; (2) among patients with PTL, those with IAI had the highest median maternal concentration of visfatin; (3) the changes in maternal plasma visfatin remained significant after adjusting for maternal age, body mass index, gestational age at sampling, and birth weight.
Conclusion. (1) PTL with IAI is characterized by high maternal circulating visfatin concentrations; (2) these findings suggest that visfatin plays a role in the regulation of the metabolic adaptations to insults resulting in PTL in the context of IAI.
Introduction
Infection and/or inflammation has been implicated as a mechanism of disease responsible for preterm parturition, as well as fetal injury Citation[1-14]. Moreover, infection/inflammation is the only pathologic processes for which a solid causal link with preterm parturition has been established Citation[10],Citation[15]. Indeed, there is experimental, epidemiologic and clinical evidence linking intra-amniotic infection and/or inflammation and preterm parturition Citation[2-5],Citation[8],Citation[10],Citation[11],Citation[15-58]. In addition, a large body of evidence supports the same cause and effect relationship between systemic infection/inflammation and preterm parturition: (1) systemic administration of microbial products to pregnant mice can result in spontaneous preterm labor (PTL) and preterm birth Citation[33],Citation[59],Citation[60]; (2) PTL and preterm birth can be induced by the administration of IL-1 to pregnant mice Citation[61] and exposure of these mice to IL-1 receptor antagonist abrogates parturition Citation[62]; (3) systemic infection (e.g. malaria Citation[63-65], pyelonephritis Citation[66-70], pneumonia Citation[71-73], and periodontal disease Citation[74-83]) have been associated with preterm birth; and (4) disorders characterized by a chronic inflammatory state such as systemic lupus erythematosus Citation[84],Citation[85], inflammatory bowel diseases Citation[86],Citation[87], asthma Citation[88],Citation[89], and morbid obesity Citation[90-92] have been associated with preterm parturition.
Adipose tissue has emerged as a highly active endocrine organ Citation[93-96] that can orchestrate a metabolic response to insults, as well as an inflammatory response via the production of soluble factors known as adipocytokines. Indeed, adipocytokines have been implicated in the pathophysiology of inflammatory disorders including asthma Citation[97],Citation[98], ulcerative colitis Citation[99],Citation[100], Crohn's disease Citation[99], rheumatoid arthritis Citation[101],Citation[102], multiple sclerosis Citation[103],Citation[104], and obesity Citation[105-114]. Moreover, these highly active peptide and proteins have immunoregulatory effects on the innate (e.g. resistin Citation[58],Citation[115],Citation[116], visfatin Citation[57],Citation[117],Citation[118]) adaptive or both limbs of the immune response (e.g. leptin Citation[119-122], adiponectin Citation[123-126]). Importantly, adipokines have also been implicated in the adaptation to gestation and complications of pregnancy Citation[57],Citation[108],Citation[110],Citation[111],Citation[113],Citation[114],Citation[127-134].
Visfatin, a 52 kDa newly discovered adipokine, has already been identified more than a decade ago as a growth factor for early B cell, termed pre-B cell colony-enhancing factor (PBEF) Citation[117],Citation[118],Citation[135-137]. The re-discovery of visfatin as an adipokine that is preferentially produced by visceral adipose tissue Citation[138-140], has facilitated the investigation of its metabolic and immunoregulatory effects. Indeed, in addition to its insulin-mimicking effects Citation[139],Citation[141], visfatin plays a key role as a regulator of the innate immune response Citation[117],Citation[137],Citation[142] as well as in inflammation associated with infection Citation[143]. Recently, we have reported that intra-amniotic infection/inflammation (IAI) is characterized by elevated amniotic fluid concentrations of adipocytokines Citation[57],Citation[58]. Specifically, amniotic fluid concentrations of visfatin were higher in patients with PTL and infection than those with PTL without infection.
Currently, there are no reports concerning maternal plasma visfatin in the presence of PTL or infectious disease. Thus, the aim of this study was to determine whether spontaneous PTL with intact membranes and IAI are associated with changes in maternal plasma circulating visfatin concentrations.
Materials and methods
Study design and population
A cross-sectional study was designed by searching our clinical database and bank of biological samples, including 284 patients in the following groups: (1) normal pregnant women (n = 123); (2) patients with an episode of PTL and intact membranes without IAI who delivered at term (n = 57); (3) PTL without IAI who delivered preterm (<37 weeks gestation) (n = 47); and (4) PTL with IAI who delivered preterm (n = 57).
All women provided written informed consent before enrolment and the collection of blood and amniotic fluid. The collection and utilization of blood and amniotic fluid for research purposes was approved by the Institutional Review Boards of the Sotero del Rio Hospital (Chile), the Wayne State University (Detroit, Michigan) and the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) (Bethesda, Maryland). Many of these samples have previously been used to study the biology of inflammation, homeostasis, and growth factor concentrations in normal pregnant women and those with pregnancy complications.
Clinical definitions
Patients were considered to have a normal pregnancy if they had a singleton gestation, a normal oral glucose challenge test Citation[144],Citation[145], and if they did not have any medical, obstetrical, or surgical complications, and delivered a term neonate (≥37 weeks) of appropriate birth weight for gestational age Citation[146],Citation[147] without complications. Spontaneous PTL was defined by the presence of regular uterine contractions occurring at a frequency of at least two every 10 min associated with cervical change before 37 completed weeks of gestation that required hospitalization. Microbial invasion of the amniotic cavity was defined as a positive amniotic fluid culture for micro-organisms. Intra-amniotic inflammation was diagnosed by an amniotic fluid IL-6 concentration ≥2.6 ng/ml Citation[148].
Amniotic fluid collection
Amniotic fluid samples were obtained by trans-abdominal amniocentesis performed for evaluation of microbial status of the amniotic cavity. Samples of amniotic fluid were transported to the laboratory in a sterile capped syringe and cultured for aerobic/anaerobic bacteria and genital mycoplasmas. An amniotic fluid white blood cell (WBC) count, glucose concentration, and Gram-stain were also performed shortly after collection as previously described Citation[149-151]. The results of these tests were used for clinical management. Amniotic fluid IL-6 concentrations were used only for research purposes. Amniotic fluid not required for clinical assessment was centrifuged for 10 min at 4°C and the supernatant was aliquoted and stored at −70°C until analysis.
Maternal plasma sample collection and human visfatin C-terminal immunoassay
Maternal blood samples were collected immediately before or after the amniocentesis into vacutainer tubes. Blood was centrifuged at 1300g for 10 min at 4°C. The plasma obtained was stored at (80°C until analysis. The concentrations of visfatin in maternal plasma were determined using specific and sensitive enzyme immunoassays purchased from Phoenix Pharmaceuticals, (Belmont, CA, USA). Visfatin C-terminal assays were validated in our laboratory for human plasma before the conduction of this study. Validation included spike and recovery experiments, which produced parallel curves indicating that maternal plasma matrix constituents did not interfere with antigen–antibody binding in this assay system. Visfatin enzyme immunoassays are based on the principle of competitive binding and were conducted according to manufacturer recommendations. Briefly, assay plates are pre-coated with a secondary antibody and the non-specific binding sites have been blocked. Standards and samples were incubated in the assay plates along with primary antiserum and biotinylated peptide. The secondary antibody in the assay plates bound to the Fc fragment of the primary antibody whose Fab fragment competitively bound with both the biotinylated peptide and peptide standard or targeted peptide in the samples. Following incubation, the assay plates were repeatedly washed to remove unbound materials and incubated with a streptavidin-horseradish peroxidase (SA-HRP) solution. Following incubation, unbound enzyme conjugate was removed by repeated washing and a substrate solution was added to the wells of the assay plates and color developed in proportion to the amount of biotinylated peptide-SA-HRP complex but inversely proportional to the amount of peptide in the standard solutions or the samples. Color development was stopped with the addition of an acid solution and the intensity of color was read using a programmable spectrophotometer (SpectraMax M2, Molecular Devices, Sunnyvale, CA, USA). Maternal plasma concentrations of visfatin C were determined by interpolation from individual standard curves composed of human visfatin peptide. The calculated inter and intra assay coefficients of variation (CVs) for visfatin C-terminal immunoassays in our laboratory were 5.3 and 2.4%, respectively. The sensitivity was calculated to be 0.04 ng/ml.
Statistical analysis
Normality of the data was tested using the Shapiro–Wilk or Kolmogorov–Smirnov tests. As plasma visfatin concentrations were not normally distributed, non-parametric tests were used for analyses. Kruskal–Wallis tests with post-hoc analysis by Mann–Whitney U tests were used for comparisons of continuous variables. Comparison of proportions was performed by Chi-square test. Spearman rank correlation was utilized to assess correlations between maternal plasma concentration of visfatin, amniotic fluid WBC count, and amniotic fluid concentrations of glucose IL-6. Multiple linear regression analysis was used to determine which factors were significantly and independently associated with maternal plasma visfatin concentrations (after log transformation). The following parameters were included in the model: maternal age, gestational age at sampling, birth weight, first trimester body mass index (BMI), and the presence of IAI. A p-value < 0.05 was considered statistically significant. Analysis was performed with SPSS, version 14 (SPSS, Chicago, IL, USA).
Results
The demographic and clinical characteristics of women with a normal pregnancy and those with PTL are displayed in . There was no significant difference in the median first trimester BMI or parity among the four groups. Women with a normal pregnancy had a lower gestational age at sampling than those with PTL without IAI who delivered preterm (p = 0.02) but not than patients with PTL and IAI (p = 0.4) or those with PTL who delivered at term (p = 0.051). Women with a normal pregnancy had a higher median birth weight than any of the three PTL groups (p < 0.001 for all comparison). Comparison of the demographics and clinical characteristics among patients with PTL is presented in .
Table I. Clinical and demographic characteristics of the study population.
Maternal plasma visfatin concentration in normal pregnant women and those with preterm labor
The median maternal plasma visfatin concentration was higher in patients with PTL with IAI than in those with PTL without IAI who delivered either preterm (median: 15.5 ng/ml, interquartile range [IQR] 14.2–19.0 vs. 14.4 ng/ml, IQR 11.3–15.9; p < 0.001, ) or at term (15.5 ng/ml, IQR 14.2–19.0 vs. 14.4 ng/ml, IQR 12.2–17.2; p = 0.002, ). Similarly, the median maternal plasma visfatin concentration was higher in patients with PTL with IAI than in those with a normal pregnancy (15.5 ng/ml, IQR 14.2–19.0 vs. 15.1 ng/ml, IQR 12.1–17.7; p = 0.002, ).
Figure 1. Comparison of the median maternal plasma visfatin between women with normal pregnancies and patients with spontaneous PTL. The median maternal plasma visfatin concentration was higher in patients with PTL with IAI than those with PTL without IAI who delivered either preterm or at term. Similarly, the median maternal plasma visfatin concentration was higher in patients with PTL with IAI than those with a normal pregnancy. The median maternal plasma visfatin concentration did not differ significantly between patients with PTL without IAI who delivered preterm and those who delivered at term. In addition, there was no significant difference in the median maternal plasma visfatin concentration between women with a normal pregnancy and those with those with PTL without IAI who delivered either preterm or at term.
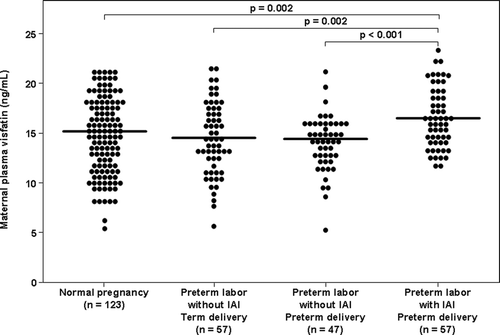
Maternal plasma visfatin concentration in normal pregnant women and those with preterm labor without intra-amniotic infection/inflammation
The median maternal plasma visfatin concentration did not differ significantly between patients with PTL without IAI who delivered preterm and those who delivered at term (p = 0.3). In addition, there was no significant difference in the median maternal plasma visfatin concentration between women with a normal pregnancy and those with PTL without IAI who delivered either preterm (p = 0.6) or at term (p = 0.07).
Multiple regression analysis was employed to examine the relationship between the plasma concentrations of visfatin and PTL adjusting for maternal age, maternal first trimester BMI, gestational age at blood sampling, birthweight, and the presence of IAI. The final regression model suggested that the presence of IAI (p = 0.001) and birthweight (p = 0.04) was independently associated with a higher maternal plasma visfatin concentration. In addition, gestational age at blood sampling (p = 0.001) was independently associated with a lower maternal plasma visfatin concentrations.
A significant correlation was observed between maternal plasma concentrations of visfatin and amniotic fluid concentrations of IL-6 (Spearman rho coefficient: 0.2, p = 0.008).
Discussion
Principal findings of the study
(1) Patients with PTL with IAI leading to preterm delivery had a significantly higher median maternal plasma concentration of visfatin than those with normal pregnancy; (2) among patients with PTL, those with IAI had the highest median maternal concentration of visfatin; and (3) There was a relationship between the maternal plasma concentration of visfatin and the amniotic fluid concentration of IL-6.
What is the physiologic role of visfatin?
Visfatin, a 52 kDa molecule, was originally reported to enhance the effect of IL-7 and stem cell factor on pre-B-cell colony formation, thus named pre-B-cell colony enhancing factor (PBEF) Citation[136]. Subsequently, visfatin was reported to be produced by adipose tissue Citation[139],Citation[152-156], preferentially by visceral fat depot Citation[138],Citation[139],Citation[157], as well as in the placenta, fetal membranes Citation[118],Citation[158-164], myometrium Citation[165], bone marrow, liver, muscle Citation[136], heart, lung, kidney Citation[136], macrophages Citation[166], and neutrophils Citation[117],Citation[136],Citation[137].
The physiologic role of visfatin in humans has not been completely elucidated, however, a growing body of evidence suggests that this adipokine plays a regulatory role in energy homeostasis and in the inflammatory response. The evidence concerning the immunoregulatory role of visfatin includes: (1) treatment of human monocytes with visfatin results in an increased secretion of IL-6, TNF-α, IL-1β in a dose-dependent manner Citation[167]; (2) acute lung injury in humans Citation[137] and animals Citation[142] is associated with increased visfatin expression from cells retrieved by bronchoalveolar lavage from the affected subjects; (3) visfatin expression increases in neutrophils retrieved from septic patients Citation[168]; and (4) chronic inflammatory disorders (e.g. inflammatory bowel disease Citation[167], rheumatoid arthritis Citation[169]) have associated with a higher circulating visfatin concentrations than normal subjects.
The following findings support the role of visfatin in metabolic regulation: (1) treatment of adipocytes with glucose results in increased secretion of visfatin Citation[170]. Consistent with this observation, administration of glucose to human subjects results in increase circulating visfatin concentration Citation[170]; (2) obesity is associated with increased circulating visfatin concentration Citation[117],Citation[152],Citation[171-175], (3) human serum concentration of visfatin are positively correlated with the amount of intra-visceral fat Citation[175]; and (4) patients with type-2 DM Citation[175-178] or metabolic syndrome Citation[179],Citation[180] have higher plasma concentrations of visfatin.
Visfatin in normal gestation and in complications of pregnancy
Visfatin plays a role in normal gestation, as well as in pregnancy complications. The evidence for that includes: (1) normal pregnancy is associated with altered maternal circulating visfatin concentrations Citation[114],Citation[181-183]; (2) gestational diabetes mellitus (GDM) is associated with higher maternal concentrations of visfatin Citation[113],Citation[184],Citation[185] than non-diabetic pregnant women; (3) pre-eclampsia Citation[186] and fetal growth restriction Citation[187] are associated with higher concentrations of visfatin than normal pregnancy; and (4) IAI is associated with higher amniotic fluid concentrations of visfatin than the absence of infection Citation[57]. Collectively, these data support a role for visfatin in the physiologic adaptations to pregnancy as well as in metabolic- and inflammatory-associated complications of pregnancy.
Intra-amniotic infection/inflammation is characterized by high circulating maternal visfatin concentrations
PTL with IAI was associated with a higher maternal plasma visfatin concentration that PTL without IAI either with preterm or term delivery. PTL with IAI was also associated with a higher maternal plasma visfatin than normal pregnant women. These findings are novel. Indeed, to date, there are no reports concerning the association between visfatin and PTL or any other infection-related conditions during pregnancy. The findings reported herein are consistent with our previous study in which median amniotic fluid concentration of visfatin found to be in patients with PTL with IAI, as well as with those reported by Bryant-Greenwood group in which expression and secretion of visfatin from amniotic epithelial were increased in response to various inflammatory stimuli Citation[118],Citation[161],Citation[163],Citation[164],Citation[188],Citation[189].
There is paucity of data concerning the association between visfatin and acute infection and/or inflammation Citation[190]. Transcription of the visfatin gene is increased in neutrophils obtained from critically ill, septic patients. Moreover, this increased transcription is mitigated through the use of an antisense oligonucleotide. Of note, neutrophils harvested from the circulation of septic patients are characterized by a marked inhibition of the apoptosis and enhanced respiratory burst capacity Citation[190],Citation[191]. Jia et al. Citation[117] have proposed that visfatin plays a crucial role in these processes. This suggestion is supported by the finding that incubation of non-activated neutrophils (taken from healthy subjects) with visfatin results in dose-dependent inhibition of apoptosis Citation[117]. Another acute condition in which the role of visfatin was investigated is acute lung injury. Ye et al. Citation[137],Citation[142] reported that there is an increased expression of visfatin in cells retrieved by bronchoalveolar lavage from patients with this condition Citation[137] and in lung tissue of animals with acute lung injury Citation[142]. Moreover, patients with the (1001G allele in the visfatin gene have increased risk of developing adult respiratory distress syndrome (ARDS) than wild-type homozygotes, whereas the (1543T allele is associated with decreased risk of developing ARDS in septic shock patients Citation[192].
Why is preterm labor with intra-amniotic infection/inflammation associated with increased maternal plasma concentrations of visfatin?
We can not ascertain from the current study whether increased maternal concentration of visfatin is causally related to PTL with IAI due to the cross-sectional nature of this report. However, several possibilities can be entertained to explain the increased concentration of visfatin, including the presence of labor along with the increased metabolic burden associated with infection.
Labor is associated with a dramatic increase in energy demands as manifested by increased maternal blood glucose Citation[193],Citation[194], free fatty acids Citation[195], ketone bodies Citation[196], high glucose turnover Citation[197]. It is possible that the acute nature of PTL requires prompt metabolic rearrangements to ensure constant flux of macronutrients to the fetus and the mother. Visfatin has been implicated in the pathophysiology of insulin resistance. It is tempting to suggest that the high maternal concentration of visfatin is part of the metabolic adaptations to PTL associated with infection. Indeed, the median maternal visfatin concentrations did not differ between patients in labor (albeit preterm) and in those who are not in labor. Thus, prima facia, it seems that increased circulating visfatin is not associated with labor per se rather than with state of infection. However, the median gestational age of patients with PTL without IAI was significantly higher that this of those with normal pregnancy (not in labor). In addition, acute infection/inflammation is associated with hypermetabolism, enhanced energy expenditure, and insulin resistance Citation[198-200]. Thus, it is likely that the presence of IAI (independent of labor) is associated with increased metabolic demands. Increase in production and/or secretion of visfatin may be part of the adaptive response to the metabolic dysregulation associated with the state of infection.
An additional explanation can be that the alterations in circulating maternal visfatin concentration are aimed at regulating the immune response. Compelling evidence suggests that visfatin has an important role in the regulation of the innate immune limb. In vitro, visfatin induces, in a dose-dependent manner, the production of proinflammatory cytokines (e.g. IL-6, TNF-α, IL-1β) by human monocytes Citation[167]. Moreover, the following pro-inflammatory mediators were reported to increase visfatin expression: TNF-α in monocytes Citation[166], macrophages Citation[201] and neutrophils Citation[117], IL-6 in synovial Citation[202] and amniotic epithelial cells Citation[162] and IL-8 and granulocyte/macrophage colony stimulating factor in neutrophils Citation[117]. Similarly, expression of visfatin can be induced by endotoxin/LPS from neutrophils Citation[117] amniotic epithelial cells Citation[162], and lung microvascular endothelial cells Citation[142]. Thus, we hypothesize that the increased concentrations of visfatin in patients with PTL and IAI can be part of the innate immune response.
In conclusion, the present study is the first to demonstrate that maternal plasma visfatin concentrations are higher in patients with PTL and intra-amniotic infection. The findings reported herein together with our previous report focusing on amniotic fluid visfatin concentration suggest that this adipokine may have a role hitherto unrecognized in the pathogenesis of PTL associated with infection. The data of the present study do not allow us to determine whether the increase in maternal visfatin concentration is secondary to PTL with IAI or whether it is present already before establishment of the disease. However, as visfatin has been implicated in the regulation of metabolic adaptations to insults, as well as in the regulation of the innate immune limb, we suggest that the perturbation in visfatin homeostasis is an adaptive response to the insult imposed by PTL with IAI.
Acknowledgements
This research was supported (in part) by the Perinatology Research Branch, Division of Intramural Research, Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH, DHHS.
References
- Minkoff H. Prematurity: infection as an etiologic factor. Obstet Gynecol 1983; 62: 137–144
- Romero R, Mazor M, Wu Y K, Sirtori M, Oyarzun E, Mitchell M D, Hobbins J C. Infection in the pathogenesis of preterm labor. Semin Perinatol 1988; 12: 262–279
- Romero R, Mazor M. Infection and preterm labor. Clin Obstet Gynecol 1988; 31: 553–584
- Romero R, Brody D T, Oyarzun E, Mazor M, Wu Y K, Hobbins J C, Durum S K. Infection and labor. III. Interleukin-1: a signal for the onset of parturition. Am J Obstet Gynecol 1989; 160: 1117–1123
- Romero R, Manogue K R, Mitchell M D, Wu Y K, Oyarzun E, Hobbins J C, Cerami A. Infection and labor. IV. Cachectin-tumor necrosis factor in the amniotic fluid of women with intraamniotic infection and preterm labor. Am J Obstet Gynecol 1989; 161: 336–341
- Ledger W J. Infection and premature labor. Am J Perinatol 1989; 6: 234–236
- Gibbs R S, Romero R, Hillier S L, Eschenbach D A, Sweet R L. A review of premature birth and subclinical infection. Am J Obstet Gynecol 1992; 166: 1515–1528
- Gomez R, Ghezzi F, Romero R, Munoz H, Tolosa J E, Rojas I. Premature labor and intra-amniotic infection. Clinical aspects and role of the cytokines in diagnosis and pathophysiology. Clin Perinatol 1995; 22: 281–342
- Brocklehurst P. Infection and preterm delivery. BMJ 1999; 318: 548–549
- Romero R, Espinoza J, Mazor M, Chaiworapongsa T. The preterm parturition syndrome. 2004; 28–60
- Romero R, Espinoza J, Kusanovic J P, Gotsch F, Hassan S, Erez O, Chaiworapongsa T, Mazor M. The preterm parturition syndrome. BJOG 2006; 113(Suppl 3)17–42
- Mazaki-Tovi S, Romero R, Kusanovic J P, Erez O, Pineles B L, Gotsch F, Mittal P, Than N G, Espinoza J, Hassan S S. Recurrent preterm birth. Semin Perinatol 2007; 31: 142–158
- Gotsch F, Romero R, Kusanovic J P, Mazaki-Tovi S, Pineles B L, Erez O, Espinoza J, Hassan S S. The fetal inflammatory response syndrome. Clin Obstet Gynecol 2007; 50: 652–683
- Romero R, Nien J K, Espinoza J, Todem D, Fu W, Chung H, Kusanovic J P, Gotsch F, Erez O, Mazaki-Tovi S, et al. A longitudinal study of angiogenic (placental growth factor) and anti-angiogenic (soluble endoglin and soluble vascular endothelial growth factor receptor-1) factors in normal pregnancy and patients destined to develop preeclampsia and deliver a small for gestational age neonate. J Matern Fetal Neonatal Med 2008; 21: 9–23
- Hirsch E, Wang H. The molecular pathophysiology of bacterially induced preterm labor: insights from the murine model. J Soc Gynecol Investig 2005; 12: 145–155
- Cassell G H, Davis R O, Waites K B, Brown M B, Marriott P A, Stagno S, Davis J K. Isolation of Mycoplasma hominis and Ureaplasma urealyticum from amniotic fluid at 16-20 weeks of gestation: potential effect on outcome of pregnancy. Sex Transm Dis 1983; 10: 294–302
- Romero R, Kadar N, Hobbins J C, Duff G W. Infection and labor: the detection of endotoxin in amniotic fluid. Am J Obstet Gynecol 1987; 157: 815–819
- Romero R, Quintero R, Oyarzun E, Wu Y K, Sabo V, Mazor M, Hobbins J C. Intraamniotic infection and the onset of labor in preterm premature rupture of the membranes. Am J Obstet Gynecol 1988; 159: 661–666
- Romero R, Sirtori M, Oyarzun E, Avila C, Mazor M, Callahan R, Sabo V, Athanassiadis A P, Hobbins J C. Infection and labor. V. Prevalence, microbiology, and clinical significance of intraamniotic infection in women with preterm labor and intact membranes. Am J Obstet Gynecol 1989; 161: 817–824
- Romero R, Shamma F, Avila C, Jimenez C, Callahan R, Nores J, Mazor M, Brekus C A, Hobbins J C. Infection and labor. VI. Prevalence, microbiology, and clinical significance of intraamniotic infection in twin gestations with preterm labor. Am J Obstet Gynecol 1990; 163: 757–761
- Romero R, Avila C, Brekus C A, Morotti R. The role of systemic and intrauterine infection in preterm parturition. Ann NY Acad Sci 1991; 622: 355–375
- Romero R, Ceska M, Avila C, Mazor M, Behnke E, Lindley I. Neutrophil attractant/activating peptide-1/interleukin-8 in term and preterm parturition. Am J Obstet Gynecol 1991; 165: 813–820
- Gray D J, Robinson H B, Malone J, Thomson R B, Jr. Adverse outcome in pregnancy following amniotic fluid isolation of Ureaplasma urealyticum. Prenat Diagn 1992; 12: 111–117
- Romero R, Gonzalez R, Sepulveda W, Brandt F, Ramirez M, Sorokin Y, Mazor M, Treadwell M C, Cotton D B. Infection and labor. VIII. Microbial invasion of the amniotic cavity in patients with suspected cervical incompetence: prevalence and clinical significance. Am J Obstet Gynecol 1992; 167: 1086–1091
- McDuffie R S, Jr, Sherman M P, Gibbs R S. Amniotic fluid tumor necrosis factor-alpha and interleukin-1 in a rabbit model of bacterially induced preterm pregnancy loss. Am J Obstet Gynecol 1992; 167: 1583–1588
- Romero R, Mazor M, Morrotti R, Avila C, Oyarzun E, Insunza A, Parra M, Behnke E, Montiel F, Cassell G H. Infection and labor. VII. Microbial invasion of the amniotic cavity in spontaneous rupture of membranes at term. Am J Obstet Gynecol 1992; 166: 129–133
- Romero R, Salafia C M, Athanassiadis A P, Hanaoka S, Mazor M, Sepulveda W, Bracken M B. The relationship between acute inflammatory lesions of the preterm placenta and amniotic fluid microbiology. Am J Obstet Gynecol 1992; 166: 1382–1388
- Watts D H, Krohn M A, Hillier S L, Eschenbach D A. The association of occult amniotic fluid infection with gestational age and neonatal outcome among women in preterm labor. Obstet Gynecol 1992; 79: 351–357
- Saito S, Kasahara T, Kato Y, Ishihara Y, Ichijo M. Elevation of amniotic fluid interleukin 6 (IL-6), IL-8 and granulocyte colony stimulating factor (G-CSF) in term and preterm parturition. Cytokine 1993; 5: 81–88
- Gomez R, Romero R, Galasso M, Behnke E, Insunza A, Cotton D B. The value of amniotic fluid interleukin-6, white blood cell count, and gram stain in the diagnosis of microbial invasion of the amniotic cavity in patients at term. Am J Reprod Immunol 1994; 32: 200–210
- Gravett M G, Witkin S S, Haluska G J, Edwards J L, Cook M J, Novy M J. An experimental model for intraamniotic infection and preterm labor in rhesus monkeys. Am J Obstet Gynecol 1994; 171: 1660–1667
- Andrews W W, Hauth J C, Goldenberg R L, Gomez R, Romero R, Cassell G H. Amniotic fluid interleukin-6: correlation with upper genital tract microbial colonization and gestational age in women delivered after spontaneous labor versus indicated delivery. Am J Obstet Gynecol 1995; 173: 606–612
- Hirsch E, Saotome I, Hirsh D. A model of intrauterine infection and preterm delivery in mice. Am J Obstet Gynecol 1995; 172: 1598–1603
- Horowitz S, Mazor M, Romero R, Horowitz J, Glezerman M. Infection of the amniotic cavity with Ureaplasma urealyticum in the midtrimester of pregnancy. J Reprod Med 1995; 40: 375–379
- Mazor M, Hershkovitz R, Ghezzi F, Maymon E, Horowitz S, Leiberman J R. Intraamniotic infection in patients with preterm labor and twin pregnancies. Acta Obstet Gynecol Scand 1996; 75: 624–627
- Romero R, Gomez R, Mazor M, Ghezzi F, Yoon B H. The preterm labor syndrome. 1997; 29–49
- Yoon B H, Romero R, Jun J K, Park K H, Park J D, Ghezzi F, Kim B I. Amniotic fluid cytokines (interleukin-6, tumor necrosis factor-alpha, interleukin-1 beta, and interleukin-8) and the risk for the development of bronchopulmonary dysplasia. Am J Obstet Gynecol 1997; 177: 825–830
- Romero R, Gomez R, Ghezzi F, Yoon B H, Mazor M, Edwin S S, Berry S M. A fetal systemic inflammatory response is followed by the spontaneous onset of preterm parturition. Am J Obstet Gynecol 1998; 179: 186–193
- Ghezzi F, Gomez R, Romero R, Yoon B H, Edwin S S, David C, Janisse J, Mazor M. Elevated interleukin-8 concentrations in amniotic fluid of mothers whose neonates subsequently develop bronchopulmonary dysplasia. Eur J Obstet Gynecol Reprod Biol 1998; 78: 5–10
- Gomez R, Romero R, Ghezzi F, Yoon B H, Mazor M, Berry S M. The fetal inflammatory response syndrome. Am J Obstet Gynecol 1998; 179: 194–202
- Yoon B H, Romero R, Kim M, Kim E C, Kim T, Park J S, Jun J K. Clinical implications of detection of Ureaplasma urealyticum in the amniotic cavity with the polymerase chain reaction. Am J Obstet Gynecol 2000; 183: 1130–1137
- Romero R, Maymon E, Pacora P, Gomez R, Mazor M, Yoon B H, Berry S M. Further observations on the fetal inflammatory response syndrome: a potential homeostatic role for the soluble receptors of tumor necrosis factor alpha. Am J Obstet Gynecol 2000; 183: 1070–1077
- Pacora P, Romero R, Maymon E, Gervasi M T, Gomez R, Edwin S S, Yoon B H. Participation of the novel cytokine interleukin 18 in the host response to intra-amniotic infection. Am J Obstet Gynecol 2000; 183: 1138–1143
- Challis J R, Lye S J, Gibb W, Whittle W, Patel F, Alfaidy N. Understanding preterm labor. Ann NY Acad Sci 2001; 943: 225–234
- Yoon B H, Oh S Y, Romero R, Shim S S, Han S Y, Park J S, Jun J K. An elevated amniotic fluid matrix metalloproteinase-8 level at the time of mid-trimester genetic amniocentesis is a risk factor for spontaneous preterm delivery. Am J Obstet Gynecol 2001; 185: 1162–1167
- Gibbs R S, Davies J K, McDuffie R S, Jr, Leslie K K, Sherman M P, Centretto C A, Wolf D M. Chronic intrauterine infection and inflammation in the preterm rabbit, despite antibiotic therapy. Am J Obstet Gynecol 2002; 186: 234–239
- Goldenberg R L, Andrews W W, Hauth J C. Choriodecidual infection and preterm birth. Nutr Rev 2002; 60: S19–S25
- Romero R, Espinoza J, Chaiworapongsa T, Kalache K. Infection and prematurity and the role of preventive strategies. Semin Neonatol 2002; 7: 259–274
- Yoon B H, Romero R, Lim J H, Shim S S, Hong J S, Shim J Y, Jun J K. The clinical significance of detecting Ureaplasma urealyticum by the polymerase chain reaction in the amniotic fluid of patients with preterm labor. Am J Obstet Gynecol 2003; 189: 919–924
- Elovitz M A, Mrinalini C. Animal models of preterm birth. Trends Endocrinol Metab 2004; 15: 479–487
- Keelan J A, Yang J, Romero R J, Chaiworapongsa T, Marvin K W, Sato T A, Mitchell M D. Epithelial cell-derived neutrophil-activating peptide-78 is present in fetal membranes and amniotic fluid at increased concentrations with intra-amniotic infection and preterm delivery. Biology of Reproduction 2004; 70: 253–259
- Menon R, Fortunato S J. Fetal membrane inflammatory cytokines: a switching mechanism between the preterm premature rupture of the membranes and preterm labor pathways. J Perinat Med 2004; 32: 391–399
- Romero R, Espinoza J, Mazor M. Can endometrial infection/inflammation explain implantation failure, spontaneous abortion, and preterm birth after in vitro fertilization?. Fertil Steril 2004; 82: 799–804
- Chaiworapongsa T, Romero R, Espinoza J, Kim Y M, Edwin S, Bujold E, Gomez R, Kuivaniemi H. Macrophage migration inhibitory factor in patients with preterm parturition and microbial invasion of the amniotic cavity. J Matern Fetal Neonatal Med 2005; 18: 405–416
- Esplin M S, Romero R, Chaiworapongsa T, Kim Y M, Edwin S, Gomez R, Mazor M, Adashi E Y. Monocyte chemotactic protein-1 is increased in the amniotic fluid of women who deliver preterm in the presence or absence of intra-amniotic infection. J Matern Fetal Neonatal Med 2005; 17: 365–373
- Vogel I, Thorsen P, Curry A, Sandager P, Uldbjerg N. Biomarkers for the prediction of preterm delivery. Acta Obstet Gynecol Scand 2005; 84: 516–525
- Mazaki-Tovi S, Romero R, Kusanovic J P, Erez O, Gotsch F, Mittal P, Than N G, Nhan-Chang C L, Hamill N, Vaisbuch E, et al. Visfatin/Pre-B cell colony-enhancing factor in amniotic fluid in normal pregnancy, spontaneous labor at term, preterm labor and prelabor rupture of membranes: an association with subclinical intrauterine infection in preterm parturition. J Perinat Med 2008; 36: 485–496
- Kusanovic J P, Romero R, Mazaki-Tovi S, Chaiworapongsa T, Mittal P, Gotsch F, Erez O, Vaisbuch E, Edwin S, Than N G, et al. Resistin in amniotic fluid and its association with intra-amniotic infection and inflammation. J Matern Fetal Neonatal Medi 2008; 21: 902–916
- Fidel P L, Jr, Romero R, Wolf N, Cutright J, Ramirez M, Araneda H, Cotton D B. Systemic and local cytokine profiles in endotoxin-induced preterm parturition in mice. Am J Obstet Gynecol 1994; 170: 1467–1475
- Kaga N, Katsuki Y, Obata M, Shibutani Y. Repeated administration of low-dose lipopolysaccharide induces preterm delivery in mice: a model for human preterm parturition and for assessment of the therapeutic ability of drugs against preterm delivery. Am J Obstet Gynecol 1996; 174: 754–759
- Romero R, Mazor M, Tartakovsky B. Systemic administration of interleukin-1 induces preterm parturition in mice. Am J Obstet Gynecol 1991; 165: 969–971
- Romero R, Tartakovsky B. The natural interleukin-1 receptor antagonist prevents interleukin-1-induced preterm delivery in mice. Am J Obstet Gynecol 1992; 167: 1041–1045
- Gilles H M, Lawson J B, Sibelas M, Voller A, Allan N. Malaria, anaemia and pregnancy. Ann Trop Med Parasitol 1969; 63: 245–263
- Herd N, Jordan T. An investigtion of malaria during pregnancy in Zimbabwe. Afr J Med 1981; 27: 62
- Kalanda B F, Verhoeff F H, Chimsuku L, Harper G, Brabin B J. Adverse birth outcomes in a malarious area. Epidemiol Infect 2006; 134: 659–666
- Hibbard L, Thrupp L, Summeril S, Smale M, Adams R. Treatment of pyelonephritis in pregnancy. Am J Obstet Gynecol 1967; 98: 609–615
- Patrick M J. Influence of maternal renal infection on the foetus and infant. Arch Dis Child 1967; 42: 208–213
- Wren B G. Subclinical renal infection and prematurity. Med J Aust 1969; 2: 596–600
- Cunningham F G, Morris G B, Mickal A. Acute pyelonephritis of pregnancy: A clinical review. Obstet Gynecol 1973; 42: 112–117
- Kaul A K, Khan S, Martens M G, Crosson J T, Lupo V R, Kaul R. Experimental gestational pyelonephritis induces preterm births and low birth weights in C3H/HeJ mice. Infect Immun 1999; 67: 5958–5966
- Benedetti T J, Valle R, Ledger W J. Antepartum pneumonia in pregnancy. Am J Obstet Gynecol 1982; 144: 413–417
- Madinger N E, Greenspoon J S, Ellrodt A G. Pneumonia during pregnancy: has modern technology improved maternal and fetal outcome?. Am J Obstet Gynecol 1989; 161: 657–662
- Munn M B, Groome L J, Atterbury J L, Baker S L, Hoff C. Pneumonia as a complication of pregnancy. J Matern Fetal Med 1999; 8: 151–154
- Offenbacher S, Beck J D, Lieff S, Slade G. Role of periodontitis in systemic health: spontaneous preterm birth. J Den Ed 1998; 62: 852–858
- Madianos P N, Lieff S, Murtha A P, Boggess K A, Auten R L, Jr, Beck J D, Offenbacher S. Maternal periodontitis and prematurity. Part II: Maternal infection and fetal exposure. Ann Periodontol 2001; 6: 175–182
- Jeffcoat M K, Geurs N C, Reddy M S, Goldenberg R L, Hauth J C. Current evidence regarding periodontal disease as a risk factor in preterm birth. Ann Periodontol 2001; 6: 183–188
- Offenbacher S, Lieff S, Boggess K A, Murtha A P, Madianos P N, Champagne C M, McKaig R G, Jared H L, Mauriello S M, Auten, et al. Maternal periodontitis and prematurity. Part I: Obstetric outcome of prematurity and growth restriction. Ann Periodontol 2001; 6: 164–174
- Jeffcoat M K, Geurs N C, Reddy M S, Cliver S P, Goldenberg R L, Hauth J C. Periodontal infection and preterm birth: results of a prospective study. J Am Den Assoc 2001; 132: 875–880
- Goepfert A R, Jeffcoat M K, Andrews W W, Fay -, tersen O, Cliver S P, Goldenberg R L, Hauth J C. Periodontal disease and upper genital tract inflammation in early spontaneous preterm birth. Obstet Gynecol 2004; 104: 777–783
- Jarjoura K, Devine P C, Perez-Delboy A, Herrera-Abreu M, D'Alton M, Papapanou P N. Markers of periodontal infection and preterm birth. Am J Obstet Gynecol 2005; 192: 513–519
- Khader Y S, Ta'ani Q. Periodontal diseases and the risk of preterm birth and low birth weight: A meta-analysis. J of Periodontol 2005; 76: 161–165
- Offenbacher S, Boggess K A, Murtha A P, Jared H L, Lieff S, McKaig R G, Mauriello S M, Moss K L, Beck J D. Progressive periodontal disease and risk of very preterm delivery. Obstet Gynecol 2006; 107: 29–36
- Xiong X, Buekens P, Fraser W D, Beck J, Offenbacher S. Periodontal disease and adverse pregnancy outcomes: a systematic review. BJOG 2006; 113: 135–143
- Johnson M J, Petri M, Witter F R, Repke J T. Evaluation of preterm delivery in a systemic lupus erythematosus pregnancy clinic. Obstet Gynecol 1995; 86: 396–399
- Le H D, Wechsler B, Vauthier-Brouzes D, Seebacher J, Lefebvre G, Bletry O, Darbois Y, Godeau P, Piette J C. Outcome of planned pregnancies in systemic lupus erythematosus: a prospective study on 62 pregnancies. Br J Rheumatol 1997; 36: 772–777
- Dominitz J A, Young J C, Boyko E J. Outcomes of infants born to mothers with inflammatory bowel disease: a population-based cohort study. Am J Gastroenterol 2002; 97: 641–648
- Norgard B, Fonager K, Pedersen L, Jacobsen B A, Sorensen H T. Birth outcome in women exposed to 5-aminosalicylic acid during pregnancy: a Danish cohort study. Gut 2003; 52: 243–247
- Kallen B, Rydhstroem H, Aberg A. Asthma during pregnancy–a population based study. Eur J Epidemiol 2000; 16: 167–171
- Dombrowski M P, Schatz M, Wise R, Momirova V, Landon M, Mabie W, Newman R B, McNellis D, Hauth J C, Lindheimer M, et al. Asthma during pregnancy. Obstet Gynecol 2004; 103: 5–12
- Adams M M, Sarno A P, Harlass F E, Rawlings J S, Read J A. Risk factors for preterm delivery in a healthy cohort. Epidemiology 1995; 6: 525–532
- Nohr E A, Bech B H, Vaeth M, Rasmussen K M, Henriksen T B, Olsen J. Obesity, gestational weight gain and preterm birth: a study within the Danish National Birth Cohort. Paediatr Perinat Epidemiol 2007; 21: 5–14
- Bhattacharya S, Campbell D M, Liston W A, Bhattacharya S. Effect of body mass index on pregnancy outcomes in nulliparous women delivering singleton babies. BMC Public Health 2007; 7: 168
- Matsuzawa Y, Funahashi T, Nakamura T. Molecular mechanism of metabolic syndrome X: contribution of adipocytokines adipocyte-derived bioactive substances. Ann NY Acad Sci 1999; 892: 146–154
- Kahn S E, Hull R L, Utzschneider K M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006; 444: 840–846
- Tilg H, Moschen A R. Adipocytokines: mediators linking adipose tissue, inflammation and immunity. Nat Rev Immunol 2006; 6: 772–783
- Catalano P M, Hoegh M, Minium J, Huston-Presley L, Bernard S, Kalhan S, Hauguel-De M S. Adiponectin in human pregnancy: implications for regulation of glucose and lipid metabolism. Diabetologia 2006; 49: 1677–1685
- Gurkan F, Atamer Y, Ece A, Kocyigit Y, Tuzun H, Mete N. Serum leptin levels in asthmatic children treated with an inhaled corticosteroid. Ann Allergy Asthma Immunol 2004; 93: 277–280
- Guler N, Kirerleri E, Ones U, Tamay Z, Salmayenli N, Darendeliler F. Leptin: does it have any role in childhood asthma?. J Allergy Clin Immunol 2004; 114: 254–259
- Barbier M, Vidal H, Desreumaux P, Dubuquoy L, Bourreille A, Colombel J F, Cherbut C, Galmiche J P. Overexpression of leptin mRNA in mesenteric adipose tissue in inflammatory bowel diseases. Gastroenterol Clin Biol 2003; 27: 987–991
- Tuzun A, Uygun A, Yesilova Z, Ozel A M, Erdil A, Yaman H, Bagci S, Gulsen M, Karaeren N, Dagalp K. Leptin levels in the acute stage of ulcerative colitis. J Gastroenterol Hepatol 2004; 19: 429–432
- Schaffler A, Ehling A, Neumann E, Herfarth H, Tarner I, Scholmerich J, Muller-Ladner U, Gay S. Adipocytokines in synovial fluid. JAMA 2003; 290: 1709–1710
- Bernotiene E, Palmer G, Talabot-Ayer D, Szalay-Quinodoz I, Aubert M L, Gabay C. Delayed resolution of acute inflammation during zymosan-induced arthritis in leptin-deficient mice. Arthritis Res Ther 2004; 6: R256–R263
- Matarese G, Sanna V, Di G A, Lord G M, Howard J K, Bloom S R, Lechler R I, Fontana S, Zappacosta S. Leptin potentiates experimental autoimmune encephalomyelitis in SJL female mice and confers susceptibility to males. Eur J Immunol 2001; 31: 1324–1332
- Batocchi A P, Rotondi M, Caggiula M, Frisullo G, Odoardi F, Nociti V, Carella C, Tonali P A, Mirabella M. Leptin as a marker of multiple sclerosis activity in patients treated with interferon-beta. J Neuroimmunol 2003; 139: 150–154
- Ahima R S, Flier J S. Adipose tissue as an endocrine organ. Trends Endocrinol Metab 2000; 11: 327–332
- Yang W S, Lee W J, Funahashi T, Tanaka S, Matsuzawa Y, Chao C L, Chen C L, Tai T Y, Chuang L M. Weight reduction increases plasma levels of an adipose-derived anti-inflammatory protein, adiponectin. J Clin Endocrinol Metab 2001; 86: 3815–3819
- Esposito K, Pontillo A, Di Palo C, Giugliano G, Masella M, Marfella R, Giugliano D. Effect of weight loss and lifestyle changes on vascular inflammatory markers in obese women: a randomized trial. JAMA 2003; 289: 1799–1804
- Nien J K, Mazaki-Tovi S, Romero R, Erez O, Kusanovic J P, Gotsch F, Pineles B L, Gomez R, Edwin S, Mazor M, et al. Plasma adiponectin concentrations in non-pregnant, normal and overweight pregnant women. J Perinat Med 2007; 35: 522–531
- Klok M D, Jakobsdottir S, Drent M L. The role of leptin and ghrelin in the regulation of food intake and body weight in humans: a review. Obes Rev 2007; 8: 21–34
- Nien J K, Mazaki-Tovi S, Romero R, Kusanovic J P, Erez O, Gotsch F, Pineles B L, Friel L A, Espinoza J, Goncalves L, et al. Resistin: a hormone which induces insulin resistance is increased in normal pregnancy. J Perinat Med 2007; 35: 513–521
- Nien J K, Mazaki-Tovi S, Romero R, Erez O, Kusanovic J P, Gotsch F, Pineles B L, Gomez R, Edwin S, Mazor M, et al. Adiponectin in severe preeclampsia. J Perinat Med 2007; 35: 503–512
- Mazaki-Tovi S, Romero R, Kusanovic J P, Erez O, Vaisbuch E, Gotsch F, Mittal P, Than G N, Nhan-Chang C, Chaiworapongsa T, et al. Adiponectin multimers in maternal plasma. J Matern Fetal Neonatal Med 2008; 21: 796–815
- Mazaki-Tovi S, Romero R, Kusanovic J P, Vaisbuch E, Erez O, Than G N, Chaiworapongsa T, Nhan-Chang C, Pacora P, Gotsch F, et al. Maternal visfatin concentration in normal pregnancy. J Perinat Med 2009; 37: 206–217
- Mazaki-Tovi S, Romero R, Kusanovic J P, Vaisbuch E, Erez O, Than G N, Chaiworapongsa T, Nhan-Chang C, Pacora P, Gotsch F, et al. Visfatin in human pregnancy: maternal gestational diabetes vis-a-vis neonatal birthweight. J Perinat Med 2009; 37: 218–231
- Verma S, Li S H, Wang C H, Fedak P W, Li R K, Weisel R D, Mickle D A. Resistin promotes endothelial cell activation: further evidence of adipokine-endothelial interaction. Circulation 2003; 108: 736–740
- Bokarewa M, Nagaev I, Dahlberg L, Smith U, Tarkowski A. Resistin, an adipokine with potent proinflammatory properties. J Immunol 2005; 174: 5789–5795
- Jia S H, Li Y, Parodo J, Kapus A, Fan L, Rotstein O D, Marshall J C. Pre-B cell colony-enhancing factor inhibits neutrophil apoptosis in experimental inflammation and clinical sepsis. J Clin Invest 2004; 113: 1318–1327
- Ognjanovic S, Bryant-Greenwood G D. Pre-B-cell colony-enhancing factor, a novel cytokine of human fetal membranes. Am J Obstet Gynecol 2002; 187: 1051–1058
- Gainsford T, Willson T A, Metcalf D, Handman E, McFarlane C, Ng A, Nicola N A, Alexander W S, Hilton D J. Leptin can induce proliferation, differentiation, and functional activation of hemopoietic cells. Proc Natl Acad Sci USA 1996; 93: 14564–14568
- Matarese G, Moschos S, Mantzoros C S. Leptin in immunology. J Immunol 2005; 174: 3137–3142
- Tian Z, Sun R, Wei H, Gao B. Impaired natural killer (NK) cell activity in leptin receptor deficient mice: leptin as a critical regulator in NK cell development and activation. Biochem Biophys Res Commun 2002; 298: 297–302
- Lord G M, Matarese G, Howard J K, Baker R J, Bloom S R, Lechler R I. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature 1998; 394: 897–901
- Wolf A M, Wolf D, Rumpold H, Enrich B, Tilg H. Adiponectin induces the anti-inflammatory cytokines IL-10 and IL-1RA in human leukocytes. Biochem Biophys Res Commun 2004; 323: 630–635
- Yamaguchi N, Argueta J G, Masuhiro Y, Kagishita M, Nonaka K, Saito T, Hanazawa S, Yamashita Y. Adiponectin inhibits Toll-like receptor family-induced signaling. FEBS Lett 2005; 579: 6821–6826
- Yokota T, Oritani K, Takahashi I, Ishikawa J, Matsuyama A, Ouchi N, Kihara S, Funahashi T, Tenner A J, Tomiyama Y, et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood 2000; 96: 1723–1732
- Okamoto Y, Folco E J, Minami M, Wara A K, Feinberg M W, Sukhova G K, Colvin R A, Kihara S, Funahashi T, Luster A D, et al. Adiponectin inhibits the production of CXC receptor 3 chemokine ligands in macrophages and reduces T-lymphocyte recruitment in atherogenesis. Circ Res 2008; 102: 218–225
- Mazaki-Tovi S, Romero R, Kusanovic J P, Erez O, Vaisbuch E, Gotsch F, Mittal P, Than N G, Nhan-Chang C L, Chaiworapongsa T, et al. Maternal serum adiponectin multimers in preeclampsia. J Perinat Med 2009, In Press
- Mazaki-Tovi S, Kanety H, Sivan E. Adiponectin and human pregnancy. Curr Diab Rep 2005; 5: 278–281
- Mazaki-Tovi S, Kanety H, Pariente C, Hemi R, Schiff E, Sivan E. Cord blood adiponectin in large-for-gestational age newborns. Am J Obstet Gynecol 2005; 193: 1238–1242
- Mazaki-Tovi S, Kanety H, Pariente C, Hemi R, Wiser A, Schiff E, Sivan E. Maternal serum adiponectin levels during human pregnancy. J Perinatol 2007; 27: 77–81
- Mazaki-Tovi S, Kanety H, Pariente C, Hemi R, Efraty Y, Schiff E, Shoham A, Sivan E. Determining the source of fetal adiponectin. J Reprod Med 2007; 52: 774–778
- Mazaki-Tovi S, Kanety H, Pariente C, Hemi R, Yinon Y, Wiser A, Schiff E, Sivan E. Adiponectin and leptin concentrations in dichorionic twins with discordant and concordant growth. J Clin Endocrinol Metab 2009; 94: 892–8
- Mazaki-Tovi S, Romero R, Kusanovic J P, Vaisbuch E, Erez O, Than N G, Chaiworapongsa T, Nhan-Chang C L, Pacora P, Gotsch F, et al. Visfatin in human pregnancy: maternal gestational diabetes vis-a-vis neonatal birthweight. J Perinat Med 2009; 37: 218–231
- Sivan E, Mazaki-Tovi S, Pariente C, Efraty Y, Schiff E, Hemi R, Kanety H. Adiponectin in human cord blood: relation to fetal birth weight and gender. J Clin Endocrinol Metab 2003; 88: 5656–5660
- Marvin K W, Keelan J A, Eykholt R L, Sato T A, Mitchell M D. Use of cDNA arrays to generate differential expression profiles for inflammatory genes in human gestational membranes delivered at term and preterm. Mol Hum Reprod 2002; 8: 399–408
- Samal B, Sun Y, Stearns G, Xie C, Suggs S, McNiece I. Cloning and characterization of the cDNA encoding a novel human pre-B-cell colony-enhancing factor. Mol Cell Biol 1994; 14: 1431–1437
- Ye S Q, Simon B A, Maloney J P, Zambelli-Weiner A, Gao L, Grant A, Easley R B, McVerry B J, Tuder R M, Standiford T, et al. Pre-B-cell colony-enhancing factor as a potential novel biomarker in acute lung injury. Am J Respir Crit Care Med 2005; 171: 361–370
- Hug C, Lodish H F. MedicineVisfatin: a new adipokine. Science 2005; 307: 366–367
- Sethi J K, Vidal-Puig A. Visfatin: the missing link between intra-abdominal obesity and diabetes?. Trends Mol Med 2005; 11: 344–347
- Tanaka M, Nozaki M, Fukuhara A, Segawa K, Aoki N, Matsuda M, Komuro R, Shimomura I. Visfatin is released from 3T3-L1 adipocytes via a non-classical pathway. Biochem Biophys Res Commun 2007; 359: 194–201
- Xie H, Tang S Y, Luo X H, Huang J, Cui R R, Yuan L Q, Zhou H D, Wu X P, Liao E Y. Insulin-like effects of visfatin on human osteoblasts. Calcif Tissue Int 2007; 80: 201–210
- Ye S Q, Zhang L Q, Adyshev D, Usatyuk P V, Garcia A N, Lavoie T L, Verin A D, Natarajan V, Garcia J G. Pre-B-cell-colony-enhancing factor is critically involved in thrombin-induced lung endothelial cell barrier dysregulation. Microvasc Res 2005; 70: 142–151
- Bajwa E K, Yu C L, Gong M N, Thompson B T, Christiani D C. Pre-B-cell colony-enhancing factor gene polymorphisms and risk of acute respiratory distress syndrome. Crit Care Med 2007; 35: 1290–1295
- Carpenter M W, Coustan D R. Criteria for screening tests for gestational diabetes. Am J Obstet Gynecol 1982; 144: 768–773
- ACOG Practice Bulletin. Clinical management guidelines for obstetrician-gynecologists. Number 30, September 2001 (replaces Technical Bulletin Number 200, December 1994). Gestational diabetes. Obstet Gynecol 2001; 98: 525–538
- Alexander G R, Himes J H, Kaufman R B, Mor J, Kogan M. A United States national reference for fetal growth. Obstet Gynecol 1996; 87: 163–168
- Gonzalez R P, Gomez R M, Castro R S, Nien J K, Merino P O, Etchegaray A B, Carstens M R, Medina L H, Viviani P G, Rojas I T. [A national birth weight distribution curve according to gestational age in Chile from 1993 to 2000]. Rev Med Chil 2004; 132: 1155–1165
- Yoon B H, Romero R, Moon J B, Shim S S, Kim M, Kim G, Jun J K. Clinical significance of intra-amniotic inflammation in patients with preterm labor and intact membranes. Am J Obstet Gynecol 2001; 185: 1130–1136
- Romero R, Emamian M, Quintero R, Wan M, Hobbins J C, Mazor M, Edberg S. The value and limitations of the Gram stain examination in the diagnosis of intraamniotic infection. Am J Obstet Gynecol 1988; 159: 114–119
- Romero R, Jimenez C, Lohda A K, Nores J, Hanaoka S, Avila C, Callahan R, Mazor M, Hobbins J C, Diamond M P. Amniotic fluid glucose concentration: a rapid and simple method for the detection of intraamniotic infection in preterm labor. Am J Obstet Gynecol 1990; 163: 968–974
- Romero R, Quintero R, Nores J, Avila C, Mazor M, Hanaoka S, Hagay Z, Merchant L, Hobbins J C. Amniotic fluid white blood cell count: a rapid and simple test to diagnose microbial invasion of the amniotic cavity and predict preterm delivery. Am J Obstet Gynecol 1991; 165: 821–830
- Berndt J, Kloting N, Kralisch S, Kovacs P, Fasshauer M, Schon M R, Stumvoll M, Bluher M. Plasma visfatin concentrations and fat depot-specific mRNA expression in humans. Diabetes 2005; 54: 2911–2916
- Choi K C, Ryu O H, Lee K W, Kim H Y, Seo J A, Kim S G, Kim N H, Choi D S, Baik S H, Choi K M. Effect of PPAR-alpha and -gamma agonist on the expression of visfatin, adiponectin, and TNF-alpha in visceral fat of OLETF rats. Biochem Biophys Res Commun 2005; 336: 747–753
- Frydelund-Larsen L, Akerstrom T, Nielsen S, Keller P, Keller C, Pedersen B K. Visfatin mRNA expression in human subcutaneous adipose tissue is regulated by exercise. Am J Physiol Endocrinol Metab 2007; 292: E24–E31
- Hammarstedt A, Pihlajamaki J, Rotter S V, Gogg S, Jansson P A, Laakso M, Smith U. Visfatin is an adipokine, but it is not regulated by thiazolidinediones. J Clin Endocrinol Metab 2006; 91: 1181–1184
- Sethi J K. Is PBEF/visfatin/Nampt an authentic adipokine relevant to the metabolic syndrome?. Curr Hypertens Rep 2007; 9: 33–38
- Tanaka M, Nozaki M, Fukuhara A, Segawa K, Aoki N, Matsuda M, Komuro R, Shimomura I. Visfatin is released from 3T3-L1 adipocytes via a non-classical pathway. Biochem Biophys Res Commun 2007; 359: 194–201
- Marvin K W, Keelan J A, Eykholt R L, Sato T A, Mitchell M D. Use of cDNA arrays to generate differential expression profiles for inflammatory genes in human gestational membranes delivered at term and preterm. Mol Hum Reprod 2002; 8: 399–408
- Nemeth E, Tashima L S, Yu Z, Bryant-Greenwood G D. Fetal membrane distention: I. Differentially expressed genes regulated by acute distention in amniotic epithelial (WISH) cells. Am J Obstet Gynecol 2000; 182: 50–59
- Nemeth E, Millar L K, Bryant-Greenwood G. Fetal membrane distention: II. Differentially expressed genes regulated by acute distention in vitro. Am J Obstet Gynecol 2000; 182: 60–67
- Kendal-Wright C E, Hubbard D, Bryant-Greenwood G D. Chronic stretching of amniotic epithelial cells increases pre-B Cell colony-enhancing factor (PBEF/Visfatin) expression and protects them from apoptosis. Placenta 2008; 29: 255–265
- Ognjanovic S, Bao S, Yamamoto S Y, Garibay-Tupas J, Samal B, Bryant-Greenwood G D. Genomic organization of the gene coding for human pre-B-cell colony enhancing factor and expression in human fetal membranes. J Mol Endocrinol 2001; 26: 107–117
- Ognjanovic S, Tashima L S, Bryant-Greenwood G D. The effects of pre-B-cell colony-enhancing factor on the human fetal membranes by microarray analysis. Am J Obstet Gynecol 2003; 189: 1187–1195
- Ognjanovic S, Ku T L, Bryant-Greenwood G D. Pre-B-cell colony-enhancing factor is a secreted cytokine-like protein from the human amniotic epithelium. Am J Obstet Gynecol 2005; 193: 273–282
- Esplin M S, Fausett M B, Peltier M R, Hamblin S, Silver R M, Branch D W, Adashi E Y, Whiting D. The use of cDNA microarray to identify differentially expressed labor-associated genes within the human myometrium during labor. Am J Obstet Gynecol 2005; 193: 404–413
- Dahl T B, Yndestad A, Skjelland M, Oie E, Dahl A, Michelsen A, Damas J K, Tunheim S H, Ueland T, Smith C, et al. Increased expression of visfatin in macrophages of human unstable carotid and coronary atherosclerosis: possible role in inflammation and plaque destabilization. Circulation 2007; 115: 972–980
- Moschen A R, Kaser A, Enrich B, Mosheimer B, Theurl M, Niederegger H, Tilg H. Visfatin, an adipocytokine with proinflammatory and immunomodulating properties. J Immunol 2007; 178: 1748–1758
- Jia S H, Li Y, Parodo J, Kapus A, Fan L, Rotstein O D, Marshall J C. Pre-B cell colony-enhancing factor inhibits neutrophil apoptosis in experimental inflammation and clinical sepsis. J Clin Invest 2004; 113: 1318–1327
- Otero M, Lago R, Gomez R, Lago F, Dieguez C, Gomez-Reino J J, Gualillo O. Changes in plasma levels of fat-derived hormones adiponectin, leptin, resistin and visfatin in patients with rheumatoid arthritis. Ann Rheum Dis 2006; 65: 1198–1201
- Haider D G, Schaller G, Kapiotis S, Maier C, Luger A, Wolzt M. The release of the adipocytokine visfatin is regulated by glucose and insulin. Diabetologia 2006; 49: 1909–1914
- Haider D G, Holzer G, Schaller G, Weghuber D, Widhalm K, Wagner O, Kapiotis S, Wolzt M. The adipokine visfatin is markedly elevated in obese children. J Pediatr Gastroenterol Nutr 2006; 43: 548–549
- Zahorska-Markiewicz B, Olszanecka-Glinianowicz M, Janowska J, Kocelak P, Semik-Grabarczyk E, Holecki M, Dabrowski P, Skorupa A. Serum concentration of visfatin in obese women. Metabolism 2007; 56: 1131–1134
- Filippatos T D, Derdemezis C S, Kiortsis D N, Tselepis A D, Elisaf M S. Increased plasma levels of visfatin/pre-B cell colony-enhancing factor in obese and overweight patients with metabolic syndrome. J Endocrinol Invest 2007; 30: 323–326
- Filippatos T D, Derdemezis C S, Gazi I F, Lagos K, Kiortsis D N, Tselepis A D, Elisaf M S. Increased plasma visfatin levels in subjects with the metabolic syndrome. Eur J Clin Invest 2008; 38: 71–72
- Sandeep S, Velmurugan K, Deepa R, Mohan V. Serum visfatin in relation to visceral fat, obesity, and type 2 diabetes mellitus in Asian Indians. Metabolism 2007; 56: 565–570
- Chen M P, Chung F M, Chang D M, Tsai J C, Huang H F, Shin S J, Lee Y J. Elevated plasma level of visfatin/pre-B cell colony-enhancing factor in patients with type 2 diabetes mellitus. J Clin Endocrinol Metab 2006; 91: 295–299
- Fernandez-Real J M, Moreno J M, Chico B, Lopez-Bermejo A, Ricart W. Circulating visfatin is associated with parameters of iron metabolism in subjects with altered glucose tolerance. Diabetes Care 2007; 30: 616–621
- Lopez-Bermejo A, Chico-Julia B, Fernandez-Balsells M, Recasens M, Esteve E, Casamitjana R, Ricart W, Fernandez-Real J M. Serum visfatin increases with progressive beta-cell deterioration. Diabetes 2006; 55: 2871–2875
- Filippatos T D, Derdemezis C S, Kiortsis D N, Tselepis A D, Elisaf M S. Increased plasma levels of visfatin/pre-B cell colony-enhancing factor in obese and overweight patients with metabolic syndrome. J Endocrinol Invest 2007; 30: 323–326
- Filippatos T D, Derdemezis C S, Gazi I F, Lagos K, Kiortsis D N, Tselepis A D, Elisaf M S. Increased plasma visfatin levels in subjects with the metabolic syndrome. Eur J Clin Invest 2008; 38: 71–72
- Mastorakos G, Valsamakis G, Papatheodorou D C, Barlas I, Margeli A, Boutsiadis A, Kouskouni E, Vitoratos N, Papadimitriou A, Papassotiriou I, et al. The role of adipocytokines in insulin resistance in normal pregnancy: visfatin concentrations in early pregnancy predict insulin sensitivity. Clin Chem 2007; 53: 1477–1483
- Morgan S A, Bringolf J B, Seidel E R. Visfatin expression is elevated in normal human pregnancy. Peptides 2008; 29: 1382–1389
- Katwa L C, Seidel E R. Visfatin in pregnancy: proposed mechanism of peptide delivery. Amino Acids 2008, DOI 10.1007
- Krzyzanowska K, Krugluger W, Mittermayer F, Rahman R, Haider D, Shnawa N, Schernthaner G. Increased visfatin concentrations in women with gestational diabetes mellitus. Clin Sci (Lond) 2006; 110: 605–609
- Lewandowski K C, Stojanovic N, Press M, Tuck S M, Szosland K, Bienkiewicz M, Vatish M, Lewinski A, Prelevic G M, Randeva H S. Elevated serum levels of visfatin in gestational diabetes: a comparative study across various degrees of glucose tolerance. Diabetologia 2007; 50: 1033–1037
- Fasshauer M, Waldeyer T, Seeger J, Schrey S, Ebert T, Kratzsch J, Lossner U, Bluher M, Stumvoll M, Faber R, et al. Serum levels of the adipokine visfatin are increased in preeclampsia. Clin Endocrinol (Oxf) 2008; 69: 69–73
- Fasshauer M, Bluher M, Stumvoll M, Tonessen P, Faber R, Stepan H. Differential regulation of visfatin and adiponectin in pregnancies with normal and abnormal placental function. Clin Endocrinol (Oxf) 2007; 66: 434–439
- Kendal-Wright C E. Stretching, mechanotransduction, and proinflammatory cytokines in the fetal membranes. Reprod Sci 2007; 14: 35–41
- Kendal C E, Bryant-Greenwood G D. Pre-B-cell colony-enhancing factor (PBEF/Visfatin) gene expression is modulated by NF-kappaB and AP-1 in human amniotic epithelial cells. Placenta 2007; 28: 305–314
- Luk T, Malam Z, Marshall J C. Pre-B cell colony-enhancing factor (PBEF)/visfatin: a novel mediator of innate immunity. J Leukoc Biol 2008; 83: 804–816
- Taneja R, Parodo J, Jia S H, Kapus A, Rotstein O D, Marshall J C. Delayed neutrophil apoptosis in sepsis is associated with maintenance of mitochondrial transmembrane potential and reduced caspase-9 activity. Crit Care Med 2004; 32: 1460–1469
- Bajwa E K, Yu C L, Gong M N, Thompson B T, Christiani D C. Pre-B-cell colony-enhancing factor gene polymorphisms and risk of acute respiratory distress syndrome. Crit Care Med 2007; 35: 1290–1295
- Kashyap M L, Sivasamboo R, Sothy S P, Cheah J S, Gartside P S. Carbohydrate and lipid metabolism during human labor: free fatty acids, glucose, insulin, and lactic acid metabolism during normal and oxytocin-induced labor for postmaturity. Metabolism 1976; 25: 865–875
- Katz M, Kroll D, Shapiro Y, Cristal N, Meizner I. Energy expenditure in normal labor. Isr J Med Sci 1990; 26: 254–257
- Whaley W H, Zuspan F P, Nelson G H, Ahlquist R P. Alterations of plasma free fatty acids and glucose during labor. Am J Obstet Gynecol 1967; 97: 875–880
- Felig P, Lynch V. [Starvation in human pregnancy: hypoglycemia, hypoinsulinemia, and hyperketonemia.]. Science 1970; 170: 990–992
- Maheux P C, Bonin B, Dizazo A, Guimond P, Monier D, Bourque J, Chiasson J L. Glucose homeostasis during spontaneous labor in normal human pregnancy. J Clin Endocrinol Metab 1996; 81: 209–215
- Goldstein S A, Elwyn D H. The effects of injury and sepsis on fuel utilization. Annu Rev Nutr 1989; 9: 445–473
- Tappy L, Chiolero R. Substrate utilization in sepsis and multiple organ failure. Crit Care Med 2007; 35: S531–S534
- Trager K, DeBacker D, Radermacher P. Metabolic alterations in sepsis and vasoactive drug-related metabolic effects. Curr Opin Crit Care 2003; 9: 271–278
- Iqbal J, Zaidi M. TNF regulates cellular NAD+ metabolism in primary macrophages. Biochem Biophys Res Commun 2006; 342: 1312–1318
- Nowell M A, Richards P J, Fielding C A, Ognjanovic S, Topley N, Williams A S, Bryant-Greenwood G, Jones S A. Regulation of pre-B cell colony-enhancing factor by STAT-3-dependent interleukin-6 trans-signaling: implications in the pathogenesis of rheumatoid arthritis. Arthritis Rheum 2006; 54: 2084–2095