
Abstract
The monophyletic group of species around Dematochroma pilosa Jolivet, Verma & Mille is identified in this work by combining information from mitochondrial DNA data and morphological features. A series of defining traits diagnosing this species assemblage from its closest phylogenetic relatives, including the genera Thasycles Chapuis and Atrichatus Sharp, is used to argue for its taxonomic separation and propose a new genus, named Dematotrichus gen. nov. Both Dematochroma pilosa and Montrouzierella hispida Jolivet, Verma & Mille are transferred to the new genus as D. pilosus (Jolivet, Verma & Mille) comb. nov. and D. hispidus (Jolivet, Verma & Mille) comb. nov., and 11 new species are described: D. capillaris sp. nov., D. capillosus sp. nov., D. comans sp. nov., D. crinitus sp. nov., D. comatulus sp. nov., D. hirtus sp. nov., D. hirsutus sp. nov., D. horridus sp. nov., D. pubescens sp. nov., D. setosus sp. nov. and D. villosus sp. nov. The work includes an identification key for all the species in the new genus.
Introduction
The Eumolpinae, known as leaf monkey beetles in Australia and by extension in the south Pacific, are exceptionally rich in species in New Caledonia, with over 100 species described and estimates that more than double this figure (Papadopoulou et al., Citation2013). Most of this diversity is still unknown and the methodical study of natural groups of species consistently demonstrates that most species are indeed undescribed, often revealing that taxa with wide ranges actually include significant numbers of undescribed, microendemic species (e.g., Gómez-Zurita & Pàmies-Harder, Citation2022; Platania & Gómez-Zurita, Citation2022). Apart from its enormous diversity, the study of this fauna presents an additional challenge, which is accommodating the high species numbers and also high morphological diversity into coherent groups of species that on the one hand reflect the phylogenetic structure of the group and on the other assist communication and referral of newly described species in the future (e.g., Farris, Citation1979). The systematics of New Caledonian Eumolpinae was built around a few problematic genera with a history of disagreement among the handful of authors who occupied themselves with this fauna (e.g., Dematochroma Baly, Citation1864), or recruited from other faunas based on some idea of similarity (e.g., Chalcoplacis Chevrolat, Citation1836 or Colaspoides Laporte, Citation1833), or were proposed without a sound character-based delimitation and diagnosis (e.g., Samuelsonia Jolivet et al., Citation2007a or Montrouzierella Jolivet et al., Citation2007a). Even the endemic genus Taophila Heller, Citation1916, unambiguously defined based on a suite of distinctive morphological traits and appearance, offered problems for a stable delimitation (Gómez-Zurita & Cardoso, Citation2014; Platania & Gómez-Zurita, Citation2022; Samuelson, Citation2010).
Facing this disarray for supraspecific assemblages, the use of molecular phylogenies in recent years helped in recognizing natural groups well characterized from a morphological point of view as well as solving some of the controversies around the generic concepts for New Caledonian Eumolpinae. Among the main advances assisted by phylogenies and the careful examination of morphological traits, some new genera have been described (Gómez-Zurita, Citation2018; Gómez-Zurita et al., Citation2020; Gómez-Zurita & Cardoso, Citation2014) or resurrected (Gómez-Zurita & Pàmies-Harder, Citation2022), and the limits of some old problematic genera have been made explicit (Gómez-Zurita & Pàmies-Harder, Citation2022; Platania & Gómez-Zurita, Citation2022). Examining these limits has been particularly relevant since it paved the route towards subsequent refinements of the genus-level systematics of this group of beetles. One significant finding of previous research was recognizing the phylogenetic position of the type species of the genus Dematochroma, the Lord Howe endemic D. picea Baly, Citation1864, as part of a divergent lineage nested within a clade including New Caledonian endemic genera Taophila and Tricholapita Gómez-Zurita & Cardoso, 2020 (in Gómez-Zurita et al., Citation2020), in turn sister to the clade including all other New Caledonian Eumolpinae and at least some Eumolpinae from New Zealand (Gómez-Zurita & Pàmies-Harder, Citation2022). The latter includes many species that had been ascribed to Dematochroma following a history of disagreements explained elsewhere (Gómez-Zurita & Pàmies-Harder, Citation2022). Given this phylogenetic structure, and in order to avoid the paraphyly of Dematochroma, all species currently known from New Caledonia (but also some from New Zealand) could be transferred to this genus or, alternatively, New Caledonian species currently in Dematochroma should be transferred to other available or newly described genera. The latter option was preferred by Gómez-Zurita and Pàmies-Harder (Citation2022) as it was more compatible with a description of the high morphological diversity observed in the island, and it avoided synonymies difficult to sustain based on marked morphological differences, including those of Taophila Heller or the New Zealand endemic Atrichatus Sharp, Citation1886.
Continuing with the methodical classification attempt initiated in previous studies, one cohesive group of species sharing similarities with Dematochroma pilosa Jolivet et al., Citation2007b and appertaining to the lineages recognized as Clades C and D in Papadopoulou et al. (Citation2013) will be revised in this work. The phylogenetic distance and paraphyly issue mentioned above, the combination of diagnostic traits for this group of species, the lack of defining characters for previously described genera, and the existence of marked differences with the generic types of genera that have not been explicitly and unambiguously defined, advise proposing a new genus to accommodate this natural group of species, which will be described below.
Materials and methods
Collections, specimens and morphological studies
The specimens for this study, including type material, are part of three institutional collections, including the Museum of Natural History of the University of Wroclaw (MNHW, Wroclaw, Poland), the Muséum National d'Histoire Naturelle (MNHN, Paris, France) and the author's research collection in the Botanical Institute of Barcelona (JGZC, CSIC, Barcelona, Spain). High-resolution photographs of the holotype of Dematochroma pilosa Jolivet et al., Citation2007b were made available from the MNHN. Pinned specimens and dry mounted specimens after DNA extraction were studied using a Leica M80 stereomicroscope, including a calibrated eyepiece for measurements and a Leica DFC420 digital camera to take photographs of habiti. Pictures of dissected male genitalia and spermathecae were used as guide for line drawings. Species descriptions followed the same scheme as our previous systematic works on this group, based on the nomenclature for external anatomy established by Lawrence et al. (Citation2010), and that of Lindroth (Citation1957) and Wagner (Citation2007) for male genitalia and spermatheca, respectively. The synthetic detailed description of the genus took into account the morphological diversity observed in all the species known to date, which in this genus tend to be rather constant in shape and proportions, and the invariable traits were not reiterated in the individual species descriptions. Specimen collection data were plotted on maps using ggplot2 3.3.5 (Wickham, Citation2016), elevatr 0.4.2 (Hollister et al., Citation2022) and Natural Earth resources (https://www.naturalearthdata.com/) in R 4.1.2 (R Core Team, Citation2021).
DNA extraction, mtDNA markers, sequencing and phylogenetic analyses
The procedures for non-destructive DNA extraction and sequencing were the same as described in previous works (e.g., Gómez-Zurita, Citation2018; Gómez-Zurita & Cardoso, Citation2014; Papadopoulou et al., Citation2013), as were the mtDNA phylogenetic markers of choice for this study: the 3′-end of the cytochrome c oxidase subunit 1 (cox1) and a fragment of the small ribosomal subunit (rrnS). Specimens that yielded useful DNA sequences are shown in , including as ingroup samples all available specimens morphologically similar to D. pilosa and four outgroups in the closely related genera Thasycles and Atrichatus, selected based on their phylogenetic proximity to the group of interest and their morphological similarities (Gómez-Zurita & Pàmies-Harder, Citation2022). Sequences newly generated for this study were deposited in the European Nucleotide Archive database (EBI-EMBL, Hinxton, UK) with accession numbers OW496001–OW496010 (rrnS) and OW496057–OW496068 (cox1). Each gene fragment was subjected to multiple sequence alignment independently using the G-INS-i algorithm in MAFFT 7.3 (Katoh & Standley, Citation2013) and resulting alignments were concatenated in a single phylogenetic matrix. These data were analysed under maximum likelihood (ML) using RAxML 7.2.8 (Stamatakis, Citation2006) and independent GTR + G + I models for three partitions, including rrnS, first and second codon positions of cox1, and third codon positions, the latter treated separately to account for the effects of saturation (Li, Citation1997). The best tree was obtained based on 100 independent thorough searches from as many random starting topologies and selecting the one with the best likelihood score, and node support was based on 500 bootstrap pseudoreplicates and the same analytical conditions as above. This same analytical conditions were also used with a single representative from each species as recognized in this study to produce a species tree hypothesis.
Table 1. Samples, including the species code number in Papadopoulou et al. (Citation2013), JGZC voucher number and provenance of specimens yielding mtDNA sequences (with accession numbers) for phylogenetic assessment in Dematotrichus gen. nov.
Tree-based species delimitation
Objective tree-based species delimitation was based on the Poisson Tree Processes branching model both under ML and in its Bayesian implementation (Zhang et al., Citation2013). The tree topology used for species delimitation was the ML solution of the previous step considering concatenated mtDNA sequence data, which in this case was treated as a single locus given their obligatory linkage and lack of recombination in the mitochondrial genome, and the analysis was run for 100,000 MCMC generations, removing the first 10% of results prior to estimation of posterior probabilities.
Results and discussion
Mitochondrial phylogeny and tree-based species delimitation
The final ML optimization likelihood score for the partitioned analysis of the concatenated cox1-rrnS matrix was −7032.727072 for the tree shown in . In this tree, all the specimens sharing similarities with D. pilosa formed a monophyletic group around this species. Moreover, with a single exception, all the species hypothesized based on morphological differences and represented by more than one individual showed as monophyletic, corroborating their identity as different putative species. The tree separated three main supported lineages: lineage A with a single most divergent species; lineage B (bootstrap support, BS = 88%) corresponding to the group of D. pilosa and including most species of the assemblage; and lineage C (BS = 94%), including data compatible with at least two species, but ultimately resolved as three, thus recognizing one of them as paraphyletic, by incorporating knowledge on objective morphological differences. Objective species delimitation based on the mtDNA gene tree and the underlying phylogenetic species concept, using both Bayesian and ML estimations of species boundaries resolved 11 species in the data set with identical support (; Supplementary Material ). However, lineage C included at least two allopatric and clearly diagnosable groups based on morphological differences and proposed as different species below, thus resulting in an integrative proposition of 12 species in the group on interest, combining information from the mtDNA tree and the analysis of morphological differences. The taxonomic treatment of the group is entirely proposed based on the analysis of morphological traits, including the proposition of a new genus for a monophyletic assemblage of species and the delimitation of the actual species in the group. However, these ideas are entirely compatible with the mtDNA tree topology, serving together as an information-rich hypothesis for the diversity and evolutionary structure of this evolutionary lineage (inset in ).
Fig. 1. Maximum likelihood tree of cox1 and rrnS data obtained from specimens morphologically similar to Dematochroma pilosa Jolivet, Verma & Mille and ascribed to the new genus Dematotrichus gen. nov. in this work. Bootstrap support numbers above 70% are shown next to the respective node and terminals are labelled with the species names proposed in this work. Asterisks identify when these specimens are selected as species holotypes. The inset shows an optimal ML tree with a single representative of the species considered in this work (voucher no. of specimen selected in brackets), except for D. hirtus sp. nov., paraphyletic for mtDNA, and it is treated as the best current hypothesis for the species tree of this group.
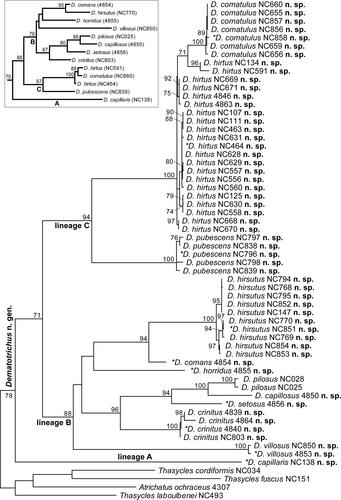
Table 2. Results of species delimitation within Dematotrichus gen. nov. based on an optimal ML topology obtained from mtDNA sequence data and the implementation of a Poisson Tree Processes branching model. The likelihood and posterior probabilities for data partitioning produced identical supports and are given together with the grouped samples, represented by JGZC voucher numbers.
Description of a new genus of New Caledonian Eumolpinae
Genus Dematotrichus gen. nov.
Type species: Dematochroma pilosa Jolivet et al., Citation2007b
urn:lsid:zoobank.org:act:E1EED023-11AF-4279-9E53-D208B7478DF4
Body elongate elliptical, constricted behind pronotum, depressed dorsally
Total body length ranging between 4.9 (in one specimen of D. hirsutus sp. nov.) to 7.5 mm (in one specimen of D. crinitus sp. nov.), and maximum body width ranging between 2.1 (in one specimen of D. pubescens sp. nov.) and 3.7 mm (in one specimen of D. hirtus sp. nov.).
Head mediocre, narrower than pronotum, transverse, narrowed anteriorly, subhypognathous; vertex convex and frontoclypeus with frons and at least basal third but most typically basal half of clypeus relatively flat, slightly depressed between eyes above supraantennal calli in D. capillaris sp. nov., D. capillosus sp. nov. and D. villosus sp. nov., and clypeus slightly convex basally in D. pubescens; frontal suture finely impressed posteriorly on frons, interocular distance wide, between 1.8× and 2.1× as wide as transverse diameter of eyes, and transition between frons and clypeus relatively narrow (not so much in D. setosus sp. nov.); clypeus shorter than wide apically, deflexed ventrally at least in apical half, and anterior border with wide arched emargination medially. Labrum transverse, as wide as anterior lobes flanking apical emargination of clypeus, with broadly round anterior angles, mostly flat with deflexed anterior margin, weakly emarginate medially on anterior border; surface finely microreticulate, generally with two setigerous punctures before anterior deflexed margin (but possibly variable: three in the type of D. comans sp. nov. or four in the type of D. hirtus). Maxillary palpi slender; first palpomere short and enlarged apically, second elongate, longest, slightly clavate, and third enlarged preapically, wider than other palpomeres; fourth palpomere as long or slightly longer and slightly broader than second, fusiform, widest at basal third, shortly cut at apex. Eyes large, prominent, laterally and posteriorly bulging, slightly stalked posteriorly, dorsoventrally elongate and weakly emarginate at inner border, with dorsal lobe slightly larger than ventral lobe. Genae short, about half as long as transverse diameter of eyes; finely microsculptured, unpunctured or with very fine punctation and scattered tiny, nearly appressed pale yellow to translucent setae. Antennae filiform, slender, long, reaching middle (males) or at least basal third (females) of elytra; antennomeres 1–6 relatively smooth, finely microreticulate, with few scattered setae on scape and pedicel and progressively more pubescent; antennomeres 7–11 microsculptured and more densely pubescent (slightly so in D. comatulus sp. nov.); antennomeres 3–6 weakly clavate and 7–10 widened apically; scape thick, subcylindrical, enlarged in apical half, flattened and slightly bent posteriorly at middle, longer than wide (from 1.8× in the type of D. pubescens to 2.6× in D. capillosus); pedicel short, not much longer but usually as long or shorter than transverse diameter of scape, bulbous, more dilated apically, about as long or slightly longer than wide (proportionally longer, 1.6× in D. capillosus); antennomeres with different proportions but 2–7 progressively longer; antennomere 11 conical and flattened in apical third. Pronotum transverse, generally 1.4× as wide at widest point as long at middle (slightly narrower in D. comans, D. pubescens and D. villosus, and slightly wider in D. hirtus), constricted in both ends, with anterior border much narrower than posterior border, 0.7× and 0.9× as wide as widest point, respectively; weakly convex in sagittal plane and moderately convex in transverse plane, with convexity accentuated towards anterior angles; anterior border straight and advanced over vertex, markedly curved in anterior view, laterally compressed at angles behind eyes, finely margined, with margin slightly enlarged towards anterior angles, dentiform, weakly protruding laterally, visible from above (except in D. capillaris), with large trichobothrium dorsally at apex; sides strongly curved anteriorly, arched (more regularly round in D. capillaris), widest slightly behind middle, converging towards and slightly sinuous before posterior angles, shortly explanate laterally, with margin thinned near ends; posterior border moderately convex, with thick margin expanded laterally beyond posterior angles as large, blunt teeth with large trichobothrium dorsally at apex (lateral teeth weak in D. capillaris); surface rather uniformly punctured and pubescent with species-specific peculiarities. Anterior border of hypomera short, straight, finely margined and following contour of anterior border of pronotum, but outwardly recurved in D. pubescens and less markedly in D. horridus sp. nov.; surface of hypomera microreticulate, alutaceous or smooth but always with relatively deep punctures at least in some areas. Prosternum transverse, short, residual in front of procoxae, with anterior border well behind mouth, rather regularly concave and finely margined, slightly recurved outwardly at sides in D. hirsutus, D. setosus and D. pubescens; prosternal process elongate, relatively narrow basally, generally half as wide as transverse diameter of procoxae, much narrower in D. pubescens and wider in D. hirtus, with sides thickly margined basally, margins reaching anterior border of prosternum, and slightly divergent posteriorly (exceptionally, D. capillaris with process as long as wide, parallel-sided, about 0.8× as wide as transverse diameter of procoxae), strongly expanded laterally at apex, enclosing procoxae posteriorly, with posterior border straight and as wide or slightly wider than procoxae; surface uneven, with indistinct punctures and sparse long dishevelled fine pale yellow setae. Mesepimera and mesanepisterna finely microreticulate, unpunctured; mesepimera generally with few tiny translucent setae near anterior border (and elsewhere in D. capillaris and D. setosus), and mesanepisterna glabrous. Mesoventrite short, as long as prosternum or slightly shorter, with basal part alutaceous, unpunctured, glabrous; base of process short, about as wide as mid-section of prosternal process, margined at sides, slightly enlarged, transversally spatulate at apex, with posterior border convex or at wide obtuse angle, anterior to posterior border of mesocoxae; surface of process relatively smooth, with sparse minute punctures and fine dishevelled pale yellow setae. Metanepisterna elongate, with wide anterior margin, gradually narrowing in anterior third and rather thin posteriorly, finely shagreened on disc, with relatively abundant short, fine, nearly appressed whitish setae. Metaventrite markedly transverse, short in median part, about as long as prosternum or up to 1.2× longer; anterior border between mesocoxae weakly convex, slightly mucronate premarginally at middle; posterior border between metacoxae explanate, at wide obtuse angle, deeply notched at middle (indented in D. capillaris); disc smooth, glossy, with finely impressed discrimen medially, scattered fine punctures and posteriorly recumbent short, fine pale yellow setae anteriorly, unpunctured and glabrous posteriorly; sides convex, finely shagreened, with abundant fine punctation and posteriorly recumbent short fine pale yellow setae. Scutellum small, as long as wide at base or slightly longer, with sides weakly convergent at base or generally parallel and regularly round or slightly arched apex; surface always finely microreticulate, punctured and pubescent, if scantly in some species. Elytra long, about 0.7× as long as body in most species (proportionally shorter, slightly over 0.6×, in D. capillaris), broader basally than base of pronotum, depressed on dorsum and compressed, nearly vertical at sides around margin, including sutural angle; humeral angles broad, obtuse, generally concealed laterally in dorsal view by callous, prominent humeri (weakly developed in D. capillaris); sides subparallel at base, weakly tapering in posterior half towards broadly round apex; surface with species-specific texture and punctation, but always with rather uniform, dense, posteriorly recumbent short pale yellow setae. Epipleura slender, entirely visible in lateral view, broader and oblique in humeral area, narrower, nearly vertical and gradually thinning posteriorly without reaching sutural angle; surface alutaceous or microreticulate, covered by relatively dense pubescence, slightly shorter than on elytra. Wings fully developed in all species. Femora elongate, inflated medially, finely microreticulate, with scattered tiny punctures and recumbent pale yellow setae ventrally mainly in mid and hind legs. Tibiae straight and nearly as long as corresponding femora in fore and hind legs, slightly and gradually enlarged towards apex with two fine longitudinal dorsal keels largely expanded apically as strong cuspid-like dorsal process in protibiae and sharp paired dorsal teeth flanking tarsal insertion in metatibiae, with outer much larger than inner tooth; mesotibiae slightly shorter than mesofemora, weakly curved ventrally, with three incomplete fine longitudinal keels dorsally, outer keel sharper, slightly enlarged and convex medially and emarginate preapically before short apical tooth; all tibiae with longitudinal rows of numerous long semierect setae and denser pubescence ventrally at apex. Tarsi much shorter than corresponding tibiae; tarsomeres 1–3 progressively shorter, with first tarsomere elongate, subtriangular, 2–3× longer than wide at apex or much longer in metatarsi, as long as or longer than tarsomeres 2–3 combined in most species, and typically shorter in D. capillaris, D. comatulus, D. pubescens and D. hirtus; second tarsomere slightly wider than first, slightly longer than wide at apex, acute at angles; third tarsomere slightly wider than long, deeply bilobate; onychia about as long as first tarsomere, clavate, curved ventrally, carrying divaricate appendiculate claws. First abdominal ventrite 1.2–1.5× longer than metaventrite, with anterior process relatively short, less than half as long as ventrite and wider at base than long, arched, with wide margins; ventrites 2–4 strongly transverse, combined as long or shorter than first ventrite, with second ventrite slightly longer than ventrites 3 or 4; fifth ventrite transverse, as long or slightly longer than fourth, emarginate at apex, with relatively large median fovea preapically. Pygidium short, weakly convex, entirely covered by apices of elytra, finely alutaceous, with tiny punctures and short fine posteriorly recumbent setae, with deep wide median longitudinal furrow apically.
Sexual dimorphism
Strong sexual dimorphism is characteristic of some groups of New Caledonian Eumolpinae (e.g., Acronymolpus Samuelson, Citation2015, Taophila or Tricholapita). However, in the case of Dematotrichus gen. nov., males and females are always very similar in shape, colour, texture and vestiture of teguments, presenting a suite of standard secondary sexual traits that are common among many groups of Chrysomelidae. These typically include larger body size in females, thinner and proportionally shorter tarsi and antennae, slightly rounder apex of elytra, more prominent and ventrally convex abdominal ventrites, and less emarginate apex of fifth abdominal ventrite.
Diagnosis
One particularly remarkable feature of this genus in the context of the diversity of New Caledonian Eumolpinae, which shall help recognizing it right away, is the relatively homogeneous dorsal pubescence, only thinner perhaps on head. Such type of vestiture is not common at all among members of the tribe Eumolpini, although a few other genera or evolutionary lineages of New Caledonian Eumolpinae exhibit some degree of dorsal pubescence. However, these groups can be told apart very easily based on the shape of their pronota. One of the closest relatives to Dematotrichus gen. nov. is the genus Thasycles Chapuis. These genera show some external similarities, including the presence of setae on pronotum and elytra, but in Thasycles they are sparser and not uniform, mostly at sides and apex of elytra, and have a distinctive pronotum, transverse, with flat anterior angles (Gómez-Zurita & Pàmies-Harder, Citation2022). The species of Taophila Heller have some pubescence on elytra and often on pronotum, but it is also much sparser and/or patchier, and they have an unmistakable body structure, in good part because of the shape of pronotum, as well, narrower than elytra, about as long as wide, narrowing at both ends, and without lateral suture or very finely margined laterally (Platania & Gómez-Zurita, Citation2022). Tricholapita Gómez-Zurita & Cardoso have more generalized pubescence on both pronotum and elytra, but once again, the pronotum is very different from Dematotrichus (or any other known related genus), showing three and most often two prominent teeth at sides (Platania et al., Citation2020). Samuelsonia pilosa Jolivet et al., Citation2007a, S. mayonae Jolivet, Verma & Mille, 2010 (in Jolivet et al., Citation2009) and a group of undescribed species related to them also have pubescent dorsum, but these are immediately recognizable because of their much smaller size (2–3× smaller), typically greenish or cupreous dorsal shine and pronotum not much narrower than elytra, widened posteriorly. The last known hairy representative of New Caledonian Eumolpinae, Dematochroma doiana Jolivet et al., Citation2007b, shows the same dense uniform pubescence on dorsum as all the species of Dematotrichus, and the penis has similar appearance, but the pronotum is very different, transverse, with anterior and posterior borders of similar length and anterior angles not compressed at sides of head.
Derivatio nominis
The generic name is composed of the same Greek root word for Dematochroma, Demato- (perhaps with the original meaning of bundle or bond; Brown, Citation1954), combined with a transliteration derived from the Greek word θρίξ or hair, -trichus, of intended masculine gender and in reference to the hairy dorsum of all species known to belong to this genus.
Taxonomic notes
As will be seen later, the new genus accommodates two species that were previously described and assigned to genera Dematochroma and Montrouzierella, respectively. An alternative to the new combinations in a new genus would be classifying the new species allied to these taxa as part of one of these genera. The large phylogenetic distance with Dematochroma, the lack of operative characters to assist classification and taxonomic conflicts with well-established genera exclude it as an alternative for any known species of New Caledonian Eumolpinae (Gómez-Zurita & Pàmies-Harder, Citation2022). By exclusion of Dematochroma, the oldest available name already used for this particular group could be Montrouzierella. However, the type species of this genus, M. nana Jolivet et al., Citation2007a, is very different anatomically to the species treated here and it belongs to a distant phylogenetic lineage (Platania et al., in prep.), which shall be the most suitable candidate to inherit this particular generic classification. An additional alternative to accommodate the diversity here ascribed to Dematotrichus could be placing the species in the genus Thasycles, which is morphologically similar and phylogenetically close to the lineage of Dematotrichus. Some noticeable similarities include, among others, the presence of dorsal pubescence, even though patchy and sparser in Thasycles, and the presence of a median dark spot in the tibiae of most species, unique among New Caledonian Eumolpinae. However, the pronotal features considered apomorphic for Thasycles (Chapuis, Citation1874; Gómez-Zurita & Pàmies-Harder, Citation2022) do not occur in the group of species treated here, and most importantly, this group is not sister to Thasycles, so that treating them as congeneric would make the genus paraphyletic, at least by inclusion of the New Zealand endemic genus Atrichatus, which also lacks the characteristic shape of pronotum of Thasycles (Gómez-Zurita & Pàmies-Harder, Citation2022). In summary, the most convenient treatment for the evolutionary lineage studied here is placing it in a new genus, as proposed. As for the extent of the genus, there is no conclusive phylogenetic evidence yet disproving the placement of D. doiana in this group of uniformly pubescent Eumolpinae, and it could be tentatively classified as Dematotrichus. However, based on the remarkably different pronotum and to avoid additional future changes in case this species is not monophyletic with this genus, the new combination is not proposed here.
Revision of species in the genus Dematotrichus gen. nov.
Dematotrichus capillaris sp. nov.
Species no. 43 in Papadopoulou et al. (Citation2013)
()
urn:lsid:zoobank.org:act:F26ED66E-F5AF-4239-A1BB-BF96F0C74BBE
Fig. 2. Dorsal views of the male holotype and one female paratype of Dematotrichus hirtus sp. nov. (a), the male holotype of D. pubescens sp. nov. (b), the male holotype of D. capillaris sp. nov. (c), the male holotype and one female paratype of D. comatulus sp. nov. (d), and the male holotype and one female paratype of D. villosus sp. nov. (e).

Fig. 3. Lateral and apical dorsal views of the penises of Dematotrichus capillaris sp. nov. (a), D. comans sp. nov. (b), D. comatulus sp. nov. (c), D. crinitus sp. nov. (d), D. hirtus sp. nov. (e), D. villosus sp. nov. (f), D. pubescens sp. nov. (g) and D. hirsutus sp. nov. (h). All penises drawn at the same scale.
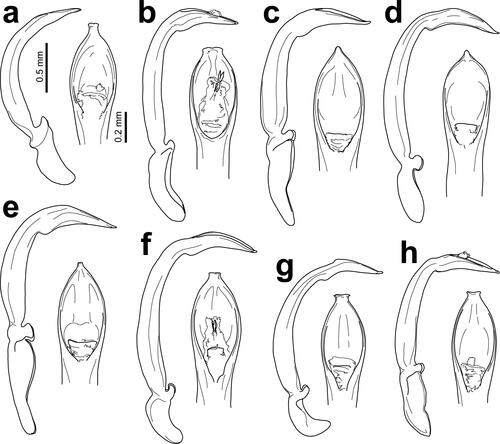
Fig. 4. Distribution maps of the species of Dematotrichus gen. nov.
Holotype: male [JGZC-NC138] (), Nouvelle-Calédonie, Prov. Nord, Baie Ugué, −21.15543 165.54190, 8 m, 30.iii.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Holotype Dematotrichus capillaris sp. nov. J. Gómez-Zurita det. [red label] (JGZC).
Body elongate elliptical, depressed on dorsum, relatively uniformly brown, darker on head, pronotum and scutellum, and testaceous on labrum, antennae, pygidium and legs, darker basally on tibiae. Length: 5.4 mm; width: 2.7 mm.
Frontoclypeus with frons flat, slightly depressed between eyes before antennal insertions, interocular distance 1.8× as wide as transverse diameter of eyes, narrowly separated from clypeus between supraantennal calli, with finely impressed frontal suture; surface glossy, with fine obliquely transverse ridges and fine punctures, smaller than intervals, of heterogeneous size, much denser in transverse area above supraantennal calli, and sparse fine semierect translucent setae; clypeus flat, glossy, nearly glabrous, with dense relatively large punctures, larger than intervals in basal half, and deflexed ventrally, finely microreticulate, with smaller, shallower punctures apically. Scape relatively long, about 2.3× as long as wide; pedicel as long as transverse diameter of scape; antennomeres 3–6 about 1.8×, 1.9×, 2.2× and 2.2× as long as pedicel; seventh antennomere longest, 2.6× as long as pedicel; antennomeres 8–9 subequal, slightly shorter than seventh, nearly 2.5× as long as pedicel; and antennomeres 10–11 slightly thinner and shorter than previous, 2.4× as long as pedicel. Surface of pronotum rather smooth and glossy, with traces of fine microreticulation mostly at sides near anterior angles, and dense small punctures, as big as smaller punctures on frons, with intervals slightly bigger than diameter of punctures, and abundant short fine recumbent pale yellow pubescence, sparser on disc, mostly oriented posteriorly and laterally near posterior border. Surface of hypomera finely alutaceous with relatively deep punctation medially in posterior third and recumbent short, fine translucent setae near anterior angle. Prosternal process nearly as wide as long, about 0.8× as wide as transverse diameter of procoxae, with sides subparallel, margined at base, strongly expanded apically, transverse at apex, wider than procoxae; surface slightly uneven, with fine punctures. Scutellum as long as wide at base, sides convergent from base to regularly round apex, with numerous tiny punctures and tiny translucent setae. Elytra slightly over 0.6× as long as body, slightly wider at base than base of pronotum, with obtuse humeral angles and weakly developed humeri; surface very finely microreticulate, shiny, with dense shallow punctures slightly bigger than punctures on pronotum and smaller than intervals, and uniformly dense posteriorly recumbent short pale yellow setae. Surface of epipleura relatively smooth, covered by dense pubescence, similar to elytra. First abdominal ventrite 1.5× as long as metaventrite and longer than ventrites 2–4 combined, with anterior process subtrapezoidal with round anterior angles; surface finely shagreened, with scattered fine punctures and posteriorly recumbent fine pale yellow setae; ventrites 2–4 with similar texture and pubescence as first; fifth ventrite with comparatively narrow emargination apically at middle and small shallow fovea near apical border. Penis () long, slender, gradually thinning from base to area before gonopore in ventral view, regularly curved ventrally slightly after middle, with apex nearly perpendicular to base, and ventral curvature regular, without stepped profile in lateral view; apical end around gonopore slightly widened, lanceolate, long, markedly concave dorsally, mucronate at distal end with elongate tooth slightly widened apically, acute at angles and biconvex at apical border; gonopore small, slightly transverse, covered at base by short dorsal flap.
Females
Unknown.
Diagnosis
This species presents marked differences with other species in the genus that correlate with its distant phylogenetic position relative to the other species. The holotype and only known specimen is stockier than any other Dematotrichus (ratio length:width = 2.0 versus 2.1–2.4), with lateral curvature of pronotum less pronounced and base of pronotum nearly as wide as base of elytra, with pronotal angles less prominent, and metatarsi with basitarsomeres much shorter than tarsomeres 2–3 together. A final distinctive trait of the species is the regular ventral curvature of penis, lacking the characteristic stepped profile of the other species. At present, this species could only be confused with D. hispidus (Jolivet, Verma & Mille), but this species from Mont Panié has sparser setae on pronotum, mostly at sides, and slightly corrugate surface of elytra with whitish setae.
Derivatio nominis
The specific epithet for this species is the adjective (m., f.) derived from the noun căpillus (m.) or hair, thus meaning pertaining to the hair.
Distribution
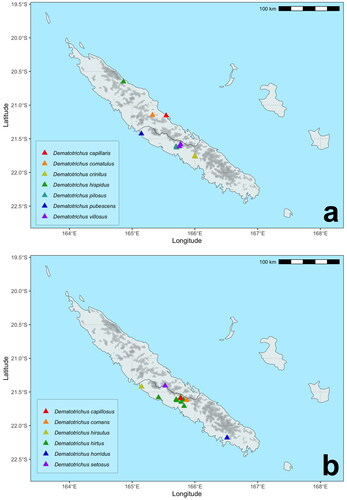
The single known specimen of D. capillaris sp. nov. was collected close to the sea in the northern coast of Grande Terre, in the mouth of the river Ho ().
Dematotrichus capillosus sp. nov.
()
urn:lsid:zoobank.org:act:F336740F-264A-4B6D-8FB6-3A534E166EEA
Fig. 5. Dorsal views of the male holotype and one female paratype of Dematotrichus hirsutus sp. nov. (a), the male holotype and one female paratype of D. crinitus sp. nov. (b), the female holotype of D. capillosus sp. nov. (c), the male holotype of D. comans sp. nov. (d), the female holotype of D. horridus sp. nov. (e), and the female holotype of D. setosus sp. nov. (f).

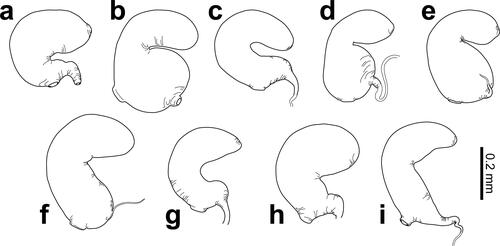
Fig. 6. Spermathecae of Dematotrichus capillosus sp. nov. (a), D. comatulus sp. nov. (b), D. crinitus sp. nov. (c), D. hirsutus sp. nov. (d), D. hirtus sp. nov. (e), D. horridus sp. nov. (f), D. pilosus (Jolivet, Verma & Mille) (g), D. setosus sp. nov. (h), and D. villosus sp. nov. (i). All spermathecae drawn at the same scale.
Holotype: female [JGZC-4849] (), New Caledonia, Parc des Grandes Fougères, N of Pic Vincent, −21.5918 165.7745, 600–680 m, leg. M. Wanat & R. Ruta, Holotype Dematotrichus capillosus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Paratype: female [JGZC-4850], New Caledonia, Parc des Grandes Fougères, N of Pic Vincent, −21.5918 165.7745, 600–680 m, leg. M. Wanat & R. Ruta, Paratype Dematotrichus capillosus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Body elongate elliptical, constricted behind pronotum, depressed dorsally. Dorsum, ventral thoracic parts and abdominal ventrites 1–3 dark brown, slightly darker with weak purplish shine on head, pronotum, scutellum and hypomera; elytra with paler areas laterally and apically; labrum, antennomeres 1–3, tip of antennomeres 4–11, most of legs, pygidium and abdominal ventrites 4–5 brown, with antennomeres 4–11 largely darkened basally and all tibiae with large median dark brown ring; mouth appendages pale ochre. Length of holotype = 6.8 mm [length of paratype = 6.0 mm]; width of holotype = 3.3 mm [width of paratype = 2.8 mm].
Frontoclypeus flattened, slightly depressed medially on frons and clypeus deflexed anteriorly, wider than long, with weak apical emargination; interocular distance 2.1× transverse diameter of eye; surface with strong microreticulation and heterogeneous small punctation on frons, with punctures generally smaller than intervals except in transition towards clypeus, smaller punctures bearing dishevelled long pale recumbent yellow setae, and clypeus with tiny punctures, setose in basal half only. Scape relatively long, about 2.6× longer than wide; pedicel half as long as scape, enlarged anteriorly towards apex, about 1.6× longer than wide; antennomeres 3–6 1.4×, 1.8×, 2.0× and 2.0× longer than pedicel; antennomeres 7–10 of similar length, 2.2× longer than pedicel; antennomere 11 slightly longer than seventh, slightly mucronate at apex. Surface of pronotum strongly microreticulate, with abundant small punctures, as big as smaller punctures on frons and smaller than intervals, many bearing long recumbent pale yellow setae, mainly oriented posteriorly, except near base and sides posteriorly and scattered setae oriented laterally at sides of disc. Surface of hypomera finely microreticulate with scattered small punctures in posterior half. Prosternal process half as wide as procoxae basally, gradually widened posteriorly, enlarged to width larger than diameter of procoxae posteriorly; surface uneven, finely microsculptured, unpunctured. Scutellum 1.1× longer than wide at base, parallel-sided at base and regularly round apically; surface with abundant small punctures and short recumbent pale yellow setae. Elytra slightly more than 0.7× as long as body; surface strongly microreticulate, matt, slightly uneven, rather uniformly punctured with punctures clearly larger than on pronotum, with intervals more than three times bigger than punctures and abundant tiny setigerous punctures with relatively short posteriorly recumbent fine pale yellow setae. Epipleura finely microreticulate and pubescent, with setae mostly aligned in single row except in humeral area. First abdominal ventrite longer than metaventrite at middle, with anterior process regularly arched; surface finely shagreened, with fine punctation and sparse posteriorly recumbent short pale yellow setae; ventrites 2–4 combined shorter than first ventrite, progressively shorter; fifth ventrite slightly longer than second, slightly emarginate apically, with deep round premarginal fovea. Spermatheca () small, with cornu inflated, weakly curved and blunt at apex, twice as long and more than twice as wide as nodulus, both connected through U-shaped turn; spermathecal duct thick, basal to nodulus, curved at straight angle opposite to cornu; ramus indicated by weak prebasal enlargement of nodulus opposite to cornu receiving spermathecal gland.
Males
Unknown.
Diagnosis
This species belongs to a group of closely related species presenting many similarities, including the presence of dark interstitial rings on all tibiae, elongate basitarsomeres, as long or longer than tarsomeres 2–3 combined, conspicuously microreticulate dorsum, variegate and slightly bumpy elytra at sides and apically, with setae clustering on weak bumps, and spermathecae with cornu larger than nodulus. D. capillosus seems to have particularly developed dark rings in protibiae compared with the other species in its group, covering 5–6 setae, while in the others they typically cover 3–5 setae, but discrete characters allow separating each other: microreticulate scutellum from D. crinitus sp. nov., with shiny scutellum; sparse finer punctation on disc of pronotum from D. pilosus (Jolivet, Verma & Mille), with stronger, dense punctation on disc; and finer, sparser pubescence on elytra and finer basal hypomeral punctures from D. setosus sp. nov., more densely pubescent and with deep punctures throughout hypomera.
Derivatio nominis
Latin adjective (m.) derived from the noun căpillus (m.), or hair, thus meaning full of hair or very hairy.
Distribution
This species is currently known from a single locality in the Central Massif ().
Dematotrichus comans sp. nov.
()
urn:lsid:zoobank.org:act:C88710CB-12DE-47ED-9071-261F54A8223D
Holotype: male [JGZC-4854] (), New Caledonia, Sarramea, trail to Dogny, −21.6229 165.8684, 300–560 m, 9.xi.2010, leg. M. Wanat & R. Ruta, Holotype Dematotrichus comans sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Body elongate elliptical, markedly constricted at junction between pronotum and elytra, depressed on dorsum. Head, including most of mandibles, pronotum, hypomera and scutellum black with very faint bronzy metallic reflection; most of elytra, including base of epipleurae, thoracic ventrites, first abdominal ventrite and disc of abdominal ventrites 2–3 deep dark brown; antennomeres 3–10, sides of prosternum, coxae, trochanters, thoracic sterna, sides of abdominal ventrites 2–3, abdominal ventrite 4, narrow base of tibiae and interstitial ring on tibiae brown; ventral surface of head, antennomeres 1, 2 and 11, labrum, humeri, apex of elytra, most of legs, pygidium and fifth abdominal ventrite orange to pale brown. Length: 6.0 mm; width: 2.5 mm.
Frontoclypeus with frons flat and clypeus triangular, weakly convex, deflexed anteriorly, with wide arched emargination in anterior border between relatively prominent lateral lobes; interocular distance slightly more than twice transverse diameter of eyes; surface of frons and most of clypeus before deflexed area densely punctured, with moderate punctures larger than intervals and scattered long dishevelled pale yellow setae; anterior deflexed margin of clypeus microreticulate, with shallow punctures and sparse long pale yellow setae. Scape about twice as long as wide; pedicel slightly longer than wide, half as long as scape, cup-shaped; antennomeres 3–6 1.5×, 1.7×, 2.1× and 2.1× longer than pedicel; antennomeres 7–9 subequal, 2.3× longer than pedicel; 10th antennomere subequal to fifth and 11th antennomere short, as long as third antennomere. Surface of pronotum glossy and punctured as densely as frons, with intervals smaller than punctures and with abundant, relatively uniform golden recumbent pubescence with most setae oriented posteriorly, except at sides of disc anteriorly and along midline. Surface of hypomera smooth, glossy, with dense large punctures, larger near anterior angles. Prosternal process relatively narrow at base, half as wide as transverse diameter of procoxae, slightly enlarged medially and strongly expanded laterally at apex, as wide as transverse diameter of procoxae; surface uneven, with irregular punctures and abundant dishevelled long fine pale yellow setae. Scutellum slightly longer than wide at base, parallel-sided basally and broadly round apically, with few small setigerous punctures in basal half and short, fine recumbent setae. Elytra about 0.65× as long as body, slightly wider at base than base of pronotum, with round humeral angles and slightly callous humeri; surface without apparent microreticulation, glossy, but densely and disorderly punctured with punctures about as big as on pronotum and slightly larger intervals, about twice as wide as punctures, corrugated and with abundant tiny setigerous punctures and dense, posteriorly recumbent long, golden setae. Epipleura smooth, with small punctures and abundant, posteriorly recumbent long golden setae. First abdominal ventrite about 1.5× as long as metaventrite, with anterior process acute with round apex and thick margins, finely microreticulate, relatively densely punctured, with posteriorly recumbent, fine long pale yellow setae; ventrites 2–4 combined as long as first ventrite, with finer punctation than first ventrite and similar texture and pubescence; fifth ventrite as long as fourth, with shallow apical emargination and apical impression medially. Penis () long, slender, nearly straight in basal half in lateral view and strongly arched at nearly right angle in apical half, enlarged ventrally in area of maximum curvature; sides subparallel in ventral view, slightly narrowed in area of maximum curvature and weakly expanded laterally around elongate elliptical, dorsally excavate gonopore; apical border of penis with broad subrectangular apical expansion, acute at weakly recurved angles and feebly emarginate at middle of apical border.
Females
Unknown.
Diagnosis
This species belongs to the group of species with ringed tibiae and elongate basitarsomeres, specifically to those with smooth and glossy teguments of pronotum and elytra. Among them, D. comans sp. nov. is almost indistinguishable from D. hirsutus sp. nov., with which it shares the strong, dense punctation on pronotum. However, at least males of both species can be recognized by dorsal pubescence, golden in D. comans and whitish in D. hirsutus, as well as the shape of the apical border of the distal projection of penis, biconvex in D. comans and concave in D. hirsutus.
Derivatio nominis
The species epithet for this species is the Latin adjective (m.) comāns, participle of the verb comō, -āre, and in this case meaning covered with hair, hairy.
Distribution
This species is only known from the holotype, captured in dense rainforest at mid elevations of the ascension to the Plateau de Dogny, in the Central Massif ().
urn:lsid:zoobank.org:act:60A1F361-E324-4E3C-A790-6881E63A71C8
Holotype: male [JGZC-NC858] (), Nouvelle-Calédonie, Prov. Nord, Aoupinié, refuge, −21.14780 165.32447, 413 m, 4.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Holotype Dematotrichus comatulus sp. nov. J. Gómez-Zurita det. [red label] (JGZC).
Paratypes: (1) 1 male [JGZC-NC660] and 3 females [JGZC-NC655, JGZC-NC656, JGZC-NC659 ()], idem, Paratype Dematotrichus comatulus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (2) 1 male [JGZC-NC856], idem, Paratype Dematotrichus comatulus sp. nov. J. Gómez-Zurita det. [red label] (MNHW); (3) 1 female [JGZC-NC857], idem, Paratype Dematotrichus comatulus sp. nov. J. Gómez-Zurita det. [red label] (MNHN).
Body elongate elliptical, constricted at base of pronotum, depressed on dorsum. Head, pronotum, scutellum, hypomera and base of epipleura black with slight bronzy reflections; elytra deep brown with irregular black areas and slight bronzy reflections; labrum and ventral surfaces dark brown; antennae with antennomeres 1–3 brown, 4–10 dark brown except near tip, and 11 dark basally and conspicuously paler at tip; most of legs and pygidium brown, with apex of femora, narrow base of tibiae and median spot on tibiae blackish. Length of holotype = 6.0 mm (range of paratypes: 6.0–7.0 mm); width of holotype = 2.7 mm (range of paratypes = 2.7–3.3 mm).
Frontoclypeus with frons and base of clypeus flat, and apical half of clypeus deflexed; interocular distance twice as large as transverse diameter of eyes; surface of frons and basal half of clypeus very finely microreticulate, nearly smooth, glossy, with sparse punctures, smaller than intervals, except anteriorly on frons, near narrow transition to clypeus, and on basal half of clypeus, with smaller, tighter punctures; some punctures on frons with recumbent, relatively short, fine, almost translucent setae; anterior half of clypeus finely microreticulate with sparse tiny punctures and anteriorly appressed short fine translucent setae; anterior border of clypeus with wide, relatively deep apical emargination. Scape nearly half as wide as long; pedicel half as long as scape, globose, slightly longer than wide; antennomeres 3–7 1.4×, 1.7×, 2.2×, 2.4× and 2.6× longer than pedicel; antennomeres 8–9 subequal, about as long as sixth antennomere; antennomeres 10–11 shorter than previous antennomeres, about as long as fifth antennomere; antennomeres 6–8 more markedly enlarged apically and antennomeres 10–11 thinner. Surface of pronotum very finely microreticulate with dense punctures, smaller than punctures on frons, as big as intervals, and abundant, dishevelled long pale yellow setae. Surface of hypomera smooth, nearly entirely covered with dense, deep punctures. Prosternal process narrow at base, half as wide as transverse diameter of procoxae, margined at sides and gradually enlarged posteriorly, prominently behind procoxae, slightly wider than transverse diameter of procoxae; surface finely microsculptured, with sparse fine punctures. Scutellum nearly 1.2× as long as wide at base, with sides parallel and arched apically, surface with sparse shallow punctures and recumbent tiny translucent setae. Elytra nearly 0.7× as long as body; surface microreticulate with abundant punctures, larger and sparser than on pronotum, with intervals more than twice as big as diameter of punctures, slightly uneven, and dense, posteriorly recumbent relatively long pale yellow setae. Epipleura microreticulate with abundant pubescence, slightly shorter and finer than on elytra. First abdominal ventrite 1.2× longer than metaventrite at middle, nearly as long as ventrites 2–4 combined, with wide arched anterior process; surface finely microreticulate, with abundant fine punctures and posteriorly recumbent fine pale yellow setae, sparser along middle; ventrites 2–4 with similar texture and pubescence as first; fifth ventrite slightly longer than fourth, with narrow apical median emargination and glabrous on disc, with small apical shallow fovea. Penis () long, slender, nearly straight and gradually narrowing apically in basal 2/3, regularly curved before apical third, perpendicular to base and enlarged ventrally at level with base of gonopore; apical third transversally concave dorsally, slightly widened, lanceolate in dorsal view with tip acute, shortly mucronate; gonopore small, elliptical, transverse, covered basally by short subtrapezoidal dorsal flap.
Females
Spermatheca () small, with cornu elongate, round at apex, connected to nodulus at right angle through narrower cylindrical short segment; nodulus larger than cornu, subspherical with narrow spermathecal duct opposite to neck connecting to cornu, and ramus indistinguishable, prebasal, opposite to cornu.
Diagnosis
One striking feature of this species that should allow distinguishing it from any other known species of Dematotrichus gen. nov. is the dense dorsal yellowish vestiture on pronotum and elytra, in contrast with a glossy black head with sparse, very fine setae. These traits are particularly useful to separate this species from D. hirtus sp. nov., the only other known species with relatively short first metatarsomeres and black apex of femora. The latter are helpful to distinguish D. comatulus sp. nov. from most other species of Dematotrichus.
Derivatio nominis
The Latin word cŏmātŭlus is the diminutive form of the adjective cŏmātus (m.), derived from the noun cŏma (f.), or hair, and it has the specific meaning of having luxurious hair, for a species with particularly abundant dorsal pubescence.
Distribution
The specimens were collected in a single mid elevation locality in rainforests of the northern slopes of the Aoupinié range ().
urn:lsid:zoobank.org:act:9885D597-CCB3-4C8D-8D65-356EEAF90EDF
Holotype: male [JGZC-4840] (), New Caledonia, Mt. Do, subsummit forest, at light, −21.76674 166.00540, 850 m, 5.xi.2008, leg. M. Wanat, Holotype Dematotrichus crinitus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Paratypes: (1) 1 male [JGZC-NC803], Mont Do, 21°45.684'S 166°00.054'E, 795 m, 4.i-10.v.2007, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus crinitus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (2) 1 female [JGZC-4839], Mt. Do, roadside, night beating, −21.76674 166.00540, 820-920 m, 6.xi.2008, leg. M. Wanat, Paratype Dematotrichus crinitus sp. nov. J. Gómez-Zurita det. [red label] (MNHW); (3) 1 male and 1 female [JGZC-4864] (), Mt. Do, roadside, night beating, −21.75706 165.99894, 900–1000 m, 6.xi.2008, leg. M. Wanat, Paratype Dematotrichus crinitus sp. nov. J. Gómez-Zurita det. [red label] (MNHW); (4) 1 female [JGZC-4861], Mt. Do, summit maquis, −21.7544 165.9995, 1025 m, 4.xi.2010, leg. M. Wanat & R. Ruta, Paratype Dematotrichus crinitus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Body elongate elliptical, constricted at junction of pronotum and elytra. Most of head except anterior half of clypeus, mandibles, pronotum, scutellum, hypomera, irregular markings on elytra, base of epipleura, narrow base and median dark spot externally of tibiae dark brown; most of elytra, most of epipleura and of ventral surfaces chestnut brown; labrum, antennae, most of legs, apex of abdomen and pygidium brown; palpi ochre brown. Length of holotype = 5.9 mm [range of paratypes: 5.7–7.5 mm]; width of holotype = 2.6 mm [range of paratypes: 2.5–3.5 mm].
Frontoclypeus with frons flat, narrowly connected with clypeus between large, glabrous, unpunctured supraantennal calli, and clypeus gradually deflexed ventrally; frons microreticulate, with abundant punctures of heterogeneous size and most of smaller punctures with relatively long fine whitish setae, oriented centrifugally from middle of frons; clypeus markedly microreticulate, with abundant tiny punctures on disc and near base, and weakly emarginate anterior border. Scape relatively short and thick, half as broad as long; pedicel short, stocky, slightly longer than wide and about half as long as scape; antennomeres 3–7 1.6×, 1.9×, 2.1×, 2.1× and 2.5× as long as pedicel; antennomeres 8–9 subequal to seventh, and 10–11 slightly longer than fifth. Surface of pronotum finely microreticulate, relatively shiny, with dense small punctures, generally smaller than punctures on frons except near sides, and typically smaller than intervals, and many punctures bearing long fine, mainly posteriorly recumbent pale yellow setae, oriented anteriorly across base. Surface of hypomera finely alutaceous with few scattered small punctures on disc. Prosternal process half as broad as procoxae with sides thickened at base and strongly enlarged posteriorly, wider than procoxae posteriorly. Scutellum slightly longer than wide at base, parallel at sides and arched at apex; surface unpunctured with few isolated minute translucent setae. Elytra 0.7× as long as body; surface weakly uneven at sides and apical declivity, with irregular depressed dark and glabrous areas among weak bumps, finely microreticulate, with abundant small punctures, larger than on pronotum and 2–3× smaller than intervals, and intervals with rather uniform tiny punctures bearing long, fine recumbent pale yellow setae. First abdominal ventrite longer than metaventrite and as long as ventrites 2–4 at middle, with anterior intercoxal process long, arched; abdominal ventrites 2–4 progressively shorter and fifth ventrite as long as fourth, broadly emarginate at apex and impressed medially on disc; surface of ventrites finely shagreened, with abundant shallow small punctures and sparse fine appressed pale yellow setae mainly at sides and near posterior border. Penis () long, slender, gradually narrowing and moderately curved ventrally before strong ventral curvature basal to gonopore, with marked ventral protuberance at level with point of maximum curvature, and long lanceolate apical end with blunt mucronate apex; gonopore small, transverse, with short subtrapezoidal dorsal flap.
Females
Spermatheca () small, compact, reniform, with thick, bulbous cornu regularly curved towards slightly narrower and shorter nodulus; spermathecal duct attached to nodulus prebasally, oriented towards cornu, enlarged basally; ramus weakly indicated by slight enlarged attachment of spermathecal gland prebasally on nodulus, opposite to duct.
Diagnosis
D. crinitus sp. nov. can be distinguished from all the other species in its group (characteristics given in the diagnosis of D. capillosus sp. nov.) because of a particular texture of scutellum, which is finely microreticulate but shiny, contrasting with the texture of elytra and pronotum. In the other species, microreticulation of scutellum is stronger, and the surface texture is more similar to surrounding areas.
Derivatio nominis
The specific epithet for this species is the Latin adjective (m.) crīnītus, related to the noun (m.) crīnis or hair, thus meaning hairy or downy.
Distribution
This species is only known from high elevations (> 800 m asl), reaching the summit of Mont Do, a rather isolated mountain and nature reserve south of the Massif Central and separated from the Humboldt Massif by the valley of the River Kuenthio ().
urn:lsid:zoobank.org:act:1808A871-CEDE-474A-B8C7-E2C1BE0F0423
Holotype: male [JGZC-NC851] (), Nouvelle-Calédonie, Poya, Mépouiri, Station Beaupré (M. Merzdorf), 21°25.453'S 165°08.807'E, 4 m, 10.i-1.iv.2008, Malaise trap, St. rech. fruit. Pocquereux staff, Holotype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (JGZC).
Paratypes: (1) 1 female [JGZC-NC852], Poya, Mépouiri, Station Beaupré (M. Merzdorf), 21°25.453'S 165°08.807'E, 4 m, 10.i-1.iv.2008, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (2) 1 female [JGZC-NC853], Poya, Station Beaupré (M. Dalstein), 21°25.305'S 165°09.084'E, 15 m, 31.vii-5.ix.2007, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (MNHW); (3) 1 female [JGZC-NC854], Poya, Station Beaupré (M. Dalstein), 21°25.305'S 165°09.084'E, 15 m, 31.vii-5.ix.2007, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (MNHN); (4) 3 females [JGZC-NC793, JGZC-NC794 (), JGZ-NC795], Poya, Mépouiri, Station Beaupré (M. Merzdorf), 21°25.453'S 165°08.807'E, 4 m, 31.vii-5.ix.2007, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (5) 3 males [JGZC-NC768, JGZC-NC769, JGZC-NC770], Poya, Mépouiri, Station Beaupré (M. Merzdorf), 21°25.453'S 165°08.807'E, 4 m, 6.xii.2007-10.i.2008, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (6) 1 female [JGZC-NC147], Poya, Station de Beaupré, −21.42392 165.14715, 26 m, 1.iv.2008, leg. J. Gómez-Zurita & A. Cardoso, Paratype Dematotrichus hirsutus sp. nov. J. Gómez-Zurita det. [red label] (JGZC).
Body elongate elliptical, constricted at junction of pronotum and elytra, depressed on dorsum. Head, mandibles, pronotum, scutellum, hypomera, thoracic ventrites, diffuse irregular areas on elytra, base and median spot on tibiae (markedly reduced in holotype) dark brown with slight bronzy reflections on head and pronotum; most of antennae, elytra, epipleura and abdominal ventrites chestnut brown; labrum, scape, maxillary palpi, most of legs and pygidium ochre brown. Length of holotype = 5.1 mm [range of paratypes: 4.9–6.2 mm]; width of holotype = 2.3 mm [range of paratypes: 2.2–3.0 mm].
Frontoclypeus flat on frons and base of clypeus, deflexed in apical 2/3 of clypeus; interocular distance about twice as wide as transverse diameter of eyes; surface mostly smooth and glossy on frons and base of clypeus, with dense large punctures, larger than intervals, and sparse long translucent setae; clypeus shorter than wide anteriorly, with anterior border weakly emarginate, finely microreticulate and densely punctured in anterior deflexed area. Scape about twice as long as wide; pedicel thick, enlarged apically, slightly longer than wide at apex, about 0.6× as long as scape; third antennomere 1.4× as long as pedicel, antennomeres 4–6 and 10 of similar length, about twice as long as pedicel, antennomeres 7–9 longest, some 2.3× longer than pedicel, and 11th antennomere slightly longer than 10th. Surface of pronotum smooth, glossy, with dense, tight punctures, smaller than punctures on frons, larger than slightly vermiculate intervals, and abundant fine, long recumbent pale yellow setae mainly oriented posteriorly, except near posterior border. Surface of hypomera smooth, glossy, with abundant large punctures mainly in basal half and near anterior border. Prosternal process relatively narrow, half as wide as transverse diameter of procoxae, transversely expanded apically behind procoxae, as wide as diameter of coxae; surface finely rugose and unpunctured basally, glossy at apex. Scutellum 1.4× longer than wide at base, sides parallel basally and regularly curved apically, finely microreticulate but glossy, with 2–3 tiny punctures on disc and short, fine, translucent setae. Elytra nearly 0.7× as long as body; surface smooth, glossy, with dense punctures, larger than punctures on pronotum and more or less as large as intervals, and intervals slightly uneven, with abundant tiny punctures and uniformly distributed relatively long posteriorly recumbent fine pale yellow setae. Epipleura smooth, entirely covered by similar pubescence as on elytra. First abdominal ventrite 1.4× longer than metaventrite at middle and longer than ventrites 2–4 together, with anterior process arched anteriorly; ventrites 3–4 slightly shorter and fifth ventrite about as long as second ventrite; surface of all ventrites densely microreticulate, with sparse shallow small punctures and very fine sparse posteriorly recumbent pale yellow setae mostly at sides. Penis () slender, elongate and gradually narrowing apically in ventral view and increasing ventral curvature in lateral view before gonopore; apical end beyond gonopore oriented nearly at straight angle with basal part of penis and enlarged ventrally at base, elongate, transversally concave dorsally and widened medially, lanceolate, with distal end as transverse projection with concave distal border and slightly acute at angles.
Females
Spermatheca () with large cornu, moderately curved and enlarged preapically before blunt apex, connected nearly at straight angle through narrower tubular part to nodulus, slightly longer and much thicker than cornu, inflated medially in same orientation as cornu, with weakly indicated ramus prebasally, opposite to cornu, and premedian insertion of spermathecal duct in opposite side.
Diagnosis
D. hirsutus sp. nov. is characterized by tibiae with dark median rings, elongate basitarsomeres and smooth teguments of pronotum and elytra, most similar to D. comans sp. nov. from which it can be separated by differences in colour of dorsal pubescence (whitish in D. hirsutus and golden in D. comans) and the shape of the distal end of penis. This species is sympatric with D. pubescens sp. nov., but they can be easily distinguished by the presence of some setae oriented anteriorly or laterally at sides of pronotum near anterior angles in D. hirsutus and all setae oriented posteriorly in the case of D. pubescens; the anterior border of hypomera recurved outwardly in the latter, regular in D. hirsutus; and the lack of dark spots on the tibiae of D. pubescens.
Derivatio nominis
The word hirsutus (m.) is a Latin adjective that is a variation and shares root with another adjective, hirtus (m.), meaning hairy.
Distribution
This species is one of the few collected in one of the few remnants of dry tropical forest in the lowlands near the SW coast of Grande Terre (). Even though we only know D. hirsutus sp. nov. from a single locality, the range of the species is expectedly larger at least in this central area of the island, despite the dramatic fragmentation of its habitat, since there are no obvious barriers for dispersal.
urn:lsid:zoobank.org:act:601060D1-6355-4DCD-9410-F05FCEF61F43
Holotype: male [JGZC-NC464] (), Nouvelle-Calédonie, Prov. Sud, ca. Farino, gate to sawing mill, −21.62416 165.70829, 290 m, 11.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Holotype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (JGZC).
Paratypes: (1) 1 male [JGZC-NC111] and 1 female [JGZC-NC463], idem, Paratype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (2) 2 males [JGZC-NC668, JGZC-NC669] and 2 females [JGZC-NC670, JGZC-NC671], Farino, refuge, −21.64877 165.78077, 261 m, 27-28.iii.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Paratype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (3) 3 males [JGZC-NC628, JGZC-NC629, JGZC-NC630] and 1 female [JGZC-NC631], ca. Farino, sawing mill, −21.62402 165.70852, 290 m, 11.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Paratype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (4) 1 male [JGZC-NC557] and 3 females [JGZC-NC125, JGZC-NC559 (), JGZC-NC560], ca. Farino, sawing mill, −21.61288 165.70209, 384 m, 11.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Paratype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); (5) 1 female [JGZC-NC556], ca. Farino, sawing mill, −21.61288 165.70209, 384 m, 11.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Paratype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (MNHW); (6) 1 female [JGZC-NC558], ca. Farino, sawing mill, −21.61288 165.70209, 384 m, 11.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg., Paratype Dematotrichus hirtus sp. nov. J. Gómez-Zurita det. [red label] (MNHN).
Other material examined: (1) 1 female, Farino, 22.iii.2005, Eduard Vives leg. (JGZC); (2) 1 male and 1 female, Col d'Amieu, La Foa, 23.iii.2005, Eduard Vives leg. (JGZC); (3) 1 female [JGZC-NC107], Col d'Amieu, 21°34.922'S 165°46.324'E, 630 m, 11.iii.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg. (JGZC); (4) 2 males [one with: JGZC-4846] and 1 female [JGZC-4863], Col d'Amieu, top of hill, ad lucem, 21°37'S 165°49'E, 450 m, 9.ii.2004, leg. M. Wanat (MNHW); (5) 1 male [JGZC-4845] and 1 female [JGZC-4862], 1 km W of Col d'Amieu, 21°37'S 165°49'E, 400 m, 10.ii.2004, leg. M. Wanat (MNHW); (6) 2 males [JGZC-NC134, JGZC-NC591], ca. Bourail, Pomea Mts, −21.58127 165.42017, 71 m, 12-13.iv.2008, J. Gómez-Zurita, J.A. Jurado & A. Cardoso leg. (JGZC).
Body elongate, elliptical, constricted behind pronotum, depressed dorsally. Head, scutellum and base of epipleura black; pronotum, hypomera and most of elytra dark brown with slight bronzy reflection; most of ventral surfaces and parts of elytra including humeri, lateral and apical declivities dark chestnut brown; labrum, antennae, legs, fifth abdominal ventrite and pygidium ochre brown, with base of antennomeres 2–4, most of antennomeres 5–7, broad apical end of femora, narrow base of tibiae and large median spot on tibiae deep brown. Length of holotype = 5.8 mm (range of species: 5.2–7.4 mm); width of holotype = 2.6 mm (range of species: 2.4–3.7 mm).
Frontoclypeus flat, except deflexed apical half of clypeus; interocular distance twice as wide as transverse diameter of eyes; surface finely microreticulate on frons, glossy with punctures of heterogeneous size, rather uniformly distributed, with intervals generally as big or bigger than larger punctures, and smaller punctures with long fine recumbent pale yellow setae; clypeus narrowly connected to frons basally, much shorter than wide at apex, with wide arched anterior emargination, surface densely microreticulate, with punctation mostly of smaller punctures, denser than on frons, with intervals generally smaller than punctures, glabrous. Scape about twice as long as wide; pedicel slightly shorter than half scape, enlarged apically, nearly as wide as long; antennomeres 3–5 1.6×, 1.9×, and 2.3× as long as pedicel; antennomeres 6–8 more markedly enlarged apically, 2.5×, 2.7× and slightly less than 2.7× as long as pedicel; antennomeres 9–11 thinner, 2.5×, 2.4× and slightly less than 2.4× as long as pedicel. Surface of pronotum very finely, superficially microreticulate, glossy, with dense small punctures, smaller than small punctures on frons, as big or nearly as big as intervals, sparser at sides, and abundant long, fine recumbent pale yellow setae with different orientations. Surface of hypomera finely microreticulate, with relatively large punctures except near lateral border. Prosternal process broad basally, nearly 3/4 as broad as transverse diameter of procoxae, with thickly margined sides, slightly widened posteriorly before large transverse apex, wider than transverse diameter of procoxae; surface slightly uneven and rugose. Scutellum as long as wide at base, sides shortly parallel at base and regularly arched towards blunt apex; surface with sparse fine punctures and tiny posteriorly recumbent translucent setae. Elytra nearly 0.7× as long as body; surface very finely microreticulate, shiny, with sparse shallow punctures, larger than pronotal punctation, most clear at sides and base of elytra, 2–3× smaller than intervals, and numerous relatively evenly distributed tiny setigerous punctures with recumbent fine, long pale yellow setae; intervals weakly convex, tending to irregular longitudinal series laterally and apically on elytra. Epipleura finely shagreened and densely covered by posteriorly recumbent setae slightly shorter and finer than on elytra. First abdominal ventrite longer than metaventrite, with round wide anterior intermetacoxal process, finely shagreened, with sparse fine punctures and relatively short, nearly appressed fine pale yellow setae; abdominal ventrites 2–4 combined as long as first ventrite, with similar texture and pubescence as first ventrite; fifth abdominal ventrite as long as third, with wide apical emargination and elliptical median fovea close to apical border. Penis () elongate, nearly straight in basal 3/5, narrowing apically in ventral and lateral views, regularly curved ventrally at middle making apical end perpendicular to base, enlarged ventrally after base of curvature and before level of gonopore; apical third weakly widened laterally after gonopore, lanceolate, transversally concave dorsally and acute, shortly mucronate at apex; gonopore small, elliptical, transverse, covered at base by short, transverse dorsal flap.
Females
Spermatheca () small, compact, with cornu tubular, slightly curved, tapering to blunt apex, regularly curved at base connecting with nearly spherical nodulus, shorter than cornu; spermathecal duct membranous, with relatively wide attachment to nodulus opposite to base of cornu; ramus indistinguishable, with spermathecal gland attached to nodulus prebasally, opposite to orientation of cornu.
Diagnosis
This species and D. comatulus sp. nov. are the only known so far with darkened apex of femora in the genus, which makes them readily recognizable. These two species can be told apart because D. hirtus sp. nov. has sparser and finer dorsal setae including the head, so that there is no stark contrast between the vestiture of head and the rest of the body, as happens in D. comatulus.
Derivatio nominis
The Latin adjective hirtus (m.) means hairy, which in this case also refers to the conspicuous dorsal pubescence of this species.
Note
Two specimens from a coastal locality near Bourail show some degree of genetic divergence relative to the samples from other more interior provenances (NC134 and NC591, sister to D. comatulus in ). In a previous study, these genetic differences supported recognizing them as a different phylogenetic species (#51 in Papadopoulou et al., Citation2013). While the possibility of cryptic species is not discarded, they are conservatively treated here as conspecific with the specimens from interior localities identified as D. hirtus sp. nov., since there are no morphological differences that can be appreciated at present among these specimens. D. hirtus is the only species showing as paraphyletic in the mtDNA tree of Dematotrichus gen. nov., which does not challenge the idea of the species as an individual evolutionary lineage, but simply reflects processes that leave the signature of paraphyly in gene evolution (Brower et al., Citation1996; Funk & Omland, Citation2003; Shaw, Citation1998). MtDNA paraphyly of New Caledonian species of Eumolpinae is rather exceptional, and most species represented by more than one individual in previous studies show as monophyletic. Species mtDNA paraphyly was reported for Taophila subsericea Heller, Citation1916 (Gómez-Zurita & Cardoso, Citation2014), but the issue was resolved understanding that the grade actually represented several originally unrecognized species, each in turn monophyletic for mtDNA (Platania & Gómez-Zurita, Citation2022). Only the monophyly of the actual T. subsericea was challenged, by inclusion of one specimen of T. dapportoi Platania & Gómez-Zurita, Citation2022, but this situation was satisfactorily explained based on phylogeographic considerations assuming interspecific hybridization and mtDNA introgression from T. subsericea into T. dapportoi. The work of Papadopoulou et al. (Citation2013) hinted at mtDNA paraphyly of Dematochroma difficilis (Heller, Citation1916) [species 60], but it is very likely that this issue will be solved similarly as in the case of T. subsericea, recognizing the involvement of several species in the mtDNA grade. So far, there has been a single report of genuine species mtDNA paraphyly for New Caledonian Eumolpinae, affecting Thasycles variegatus Gómez-Zurita, 2022, and it shared many similarities with the case of D. hirtus sp. nov., possibly representing the same general process. Specifically, both D. hirtus and T. variegatus, species with relatively large ranges in the central region of Grande Terre, show in the mtDNA trees as unresolved assortments of haplotypes, compatible with star trees. In both cases, from this basal polytomy stems a divergent branch corresponding to a monophyletic entity that represents a different closely related species, which in both cases are allopatric to their putative sister: D. comatulus sp. nov., and T. tenuis Gómez-Zurita, 2022 (in Gómez-Zurita & Pàmies-Harder, Citation2022), respectively. This mtDNA phylogenetic structure, also considering the recency of events that it depicts and the spatial distribution of the species involved, is compatible with standard processes and outcomes of speciation (Coyne & Orr, Citation2004). In particular, and assuming that mtDNA is not under selection in this case, the patterns observed are hypothesized as the result of founder events and subsequent allopatric speciation from panmictic or demographically expanding stocks (star genealogies of widespread D. hirtus and T. variegatus), with associated incomplete lineage sorting of haplotypes in the species interpreted as parentals in this case, with the signals of population bottleneck and drift in the daughter species (Coyne & Orr, Citation2004).
Distribution
Dematotrichus hirtus sp. nov. is one of the species with the largest ranges of all studied species, including samples in southern locations of the Massif Central, from La Foa to the Boghen river valley, reaching low elevations in the dry tropical forest biome of New Caledonia, south of the small coastal range west of Bourail ().
Dematotrichus hispidus (Jolivet et al., Citation2013) comb. nov.
()
Jolivet et al. (Citation2013) described a medium-sized (5 mm) male Eumolpinae from Mont Panié densely pubescent on dorsum, which was originally placed in the genus Montrouzierella Jolivet et al., Citation2007a, as M. hispida Jolivet et al., Citation2013. This species was not available for analysis here, but there are a number of characteristics in the description, apart from the uniformly pubescent body, that allow recognizing it as a member of Dematotrichus gen. nov., and specifically allied to D. capillaris sp. nov. These traits include the shape of pronotum, weakly constricted basally and markedly so at apex, with anterior angles not projecting laterally, and also vestiture of pronotum, sparsely pubescent on disc, the lack of dark spots on tibiae and the regular ventral curvature of penis, without the characteristic stepped profile of all the species of Dematotrichus, except for D. capillaris. The species is thus transferred to the genus Dematotrichus as D. hispidus (Jolivet, Verma & Mille) comb. nov. This species is currently known from a small isolated coastal range of the larger Massif of Panié (), south of the Ouaïème river and known as the Roches d'Ouaïème (Jolivet et al., Citation2013).
Dematotrichus horridus sp. nov.
()
urn:lsid:zoobank.org:act:78D83419-6BDC-4C83-B0A1-9B3BA94DD516
Holotype: female [JGZC-4855] (), New Caledonia, Upper Koghi Mts, vic. of Pic Malaoui track jct., −22.18038 166.51313, 600–700 m, 3.xii.2008, leg. M. Wanat, Holotype Dematotrichus horridus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Body elongate elliptical, constricted at base of prothorax, markedly depressed on dorsum. Head dark brown, with broad black median line on vertex, frons and base of clypeus; pronotum, hypomera and scutellum dark chestnut brown, with darker irregular patches at sides of disc of pronotum; elytra brown with small irregular blackish patches and black in humeral angles, including base of epipleura; ventral surfaces chestnut brown; labrum, maxillary palpi, antennae and legs pale brown, with dark median spot externally on all tibiae. Length: 5.9 mm; width: 2.7 mm.
Frontoclypeus with frons relatively flat, with interocular distance 1.8× as wide as transverse diameter of eyes, and clypeus subtriangular, shorter than wide at apex, flat at base and deflexed apically, with shallow apical emargination; surface finely microreticulate, with abundant small punctures, generally smaller than intervals, and slightly bigger in transition from frons to clypeus and near eyes, most bearing long pale yellow setae, recumbent and dishevelled on frons, appressed and anterior on clypeus apically. Scape about 2.5× longer than wide; pedicel longer than wide, wider medially, about 0.6× as long as scape; antennomeres 3–5 1.3×, 1.5× and 1.6× longer than pedicel; sixth antennomere nearly twice as long as pedicel and seventh antennomere longest, 2.1× longer than pedicel; antennomeres 8–11 subequal, slightly more than twice as long as pedicel. Surface of pronotum smooth, glossy, with fine microreticulation near anterior border and dense small punctures, generally as big as intervals, absent in small humps at sides of disc and near anterior border, with many punctures bearing long strongly recumbent pale yellow seta mostly with posterior orientation or radial around small unpunctured humps. Anterior border of hypomera short, slightly bent outwards, and hypomera smooth, glossy, with abundant relatively large punctures. Prosternal process relatively wide at base, wider than half diameter of procoxae, gradually widening posteriorly and enlarged laterally behind procoxae, wider than transverse diameter of procoxae. Scutellum as long as wide at base, arched apically, with scattered small punctures and short fine posteriorly recumbent translucent setae. Elytra nearly 0.7× as long as body; surface smooth, glossy with weak humps at sides and in apical declivity, with relatively dense small punctures, smaller than pronotal punctures and generally smaller than intervals, most of them with posteriorly recumbent, fine long pale yellow seta. Epipleura with abundant fine, posteriorly recumbent pale yellow setae. First abdominal ventrite about 1.5× as long as metaventrite, with anterior process arched, surface finely microreticulate, matt, with uniform small punctures and fine long recumbent pale yellow setae; abdominal ventrites 2–5 combined slightly longer than first ventrite, with similar texture and fine pubescence. Spermatheca () small with large cornu, slightly longer than wide, broadly blunt at apex, slightly narrowed basally, connecting with nodulus at weakly acute angle; nodulus as thick but slightly longer than cornu, nearly straight, weakly enlarged and round basally; ramus indicated by slight enlarged attachment of spermathecal gland, prebasal and opposite to cornu; spermathecal duct fine, attached near basal end of nodulus oriented to cornu.
Males
Unknown.
Diagnosis
This species belongs to the small group of species combining concolour femora, dark rings on tibiae, elongate basitarsomeres, and smooth and shiny dorsal teguments of pronotum and elytra. The other species with this combination of traits (D. comans sp. nov. and D. hirsutus sp. nov.) have denser and more uniform punctation on head and pronotum, stronger and denser punctation on elytra, also denser elytral pubescence, and contrary to D. horridus sp. nov., they do not have small irregular dark patches on elytra devoid of pubescence. The spermathecae of D. horridus and D. hirsutus are similar but the position of the attachment of the spermathecal duct, prebasal in D. horridus and interstitial in D. hirsutus, helps distinguishing them.
Derivatio nominis
The species name, horridus (m.), is a Latin adjective standing, among other meanings, for bristly or covered with short, stiff hairs.
Distribution
The single known specimen of this species was collected in a densely forested area at relatively high elevation of Mount Koghi, north of Nouméa ().
Dematotrichus pilosus (Jolivet et al., Citation2007b) comb. nov.
Species no. 56 in Papadopoulou et al. (Citation2013)
(, 7)
Material examined
JGZC: (1) 1 female [JGZC-NC025], Farino (M. Barbou), −21.63286 165.7068, 9.ix.2004, collecté sur divers arbres do forêt, par battage, S. Cazères leg., Dematochroma pilosa Det. P. H. Jolivet; (2) 1 female [JGZC-NC028], Farino 20 (Barbou L.), −21.61454 165.70097, 390 m, 16.xi.2004, collecté sur F. macrocarpa, S. Cazères leg., Dematochroma pilosa Det. P. H. Jolivet.
The original description of D. pilosus reported the species from Farino (type locality), in the Central Massif, but also from Mount Humboldt (Jolivet et al., Citation2007b). Subsequent works by the same authors reported it from Aoupinié, Col d'Amieu and Poya (Jolivet et al., Citation2009), from Mandjélia (Jolivet et al., Citation2010), and from a couple of localities in Mont Panié (Jolivet et al., Citation2013). These data combined are compatible with a species with a large distribution range, in most of Grande Terre and with a broad elevation range as well. However, based on the large diversity of phenotypically similar species reported in this work, it is quite possible that most, if not all of the reports from localities other than those in the Central Massif actually belong to different species, some of them described here. Another problem associated with this taxon is that, since its description was written unaware of the high species diversity of phenotypically similar species, four of them coexisting in the same general central region around the localities of Farino and Sarraméa, the information provided is rather generic and superficial without including useful diagnostic traits that could allow recognizing the species in most cases. In particular, this description applies well to every other species revealed here as present in the region of Farino (). This further complicates the issue of establishing which one of these species is the one that was effectively described in Jolivet et al. (Citation2007b). Our study includes data from one female specimen (JGZC-NC025) seemingly obtained exactly in the same collection event as the type of D. pilosus, which appears as closely related genetically to another female specimen from the same area (JGZC-NC028) and collected on the same plants as the type series (Jolivet et al., Citation2007b). The phylogeny allows recognizing the divergence of these conspecific specimens (species #56 in Papadopoulou et al., Citation2013) from other species in the same region. It is safe to assume that these specimens represent the original species, Dematochroma pilosa, and despite them being females, thus with differences with the male holotype owing to sexual secondary traits (e.g., the sculpture of teguments and body proportions) that may complicate their reliable identification, they also share some features present in the type which reinforce the idea that they are indeed conspecific. These include the generally elongate shape (ratio length/width ∼ 2.2), elytral pubescence tending to clump on elytral protuberances apically on elytra, or femora uniformly brown, without darkened apex (). The original description of D. pilosus included genitalic characters, which might have been crucial for the unambiguous recognition of the species (figs 10 and 11 in Jolivet et al., Citation2007b). However, these represent additional taxonomic problems. In the case of the male genitalia, this is drawn in lateral view, showing attributes common to most species in the genus, like the sharp ventral curvature of the apical portion, the basal ventral enlargement in the point of maximum curvature or the tapering distal end; unfortunately, a dorsal detail of the apical end of penis is missing, which would be the most informative for species recognition in this group. In the case of the spermatheca, drawn with a characteristic bulbous nodulus and short cornu (fig. 11 in Jolivet et al., Citation2007b), it is completely different from the spermathecae of the specimens identified in this work as the females of D. pilosus and also their closest relatives, and suspiciously similar to the spermatheca of D. hirtus sp. nov. (). These two species are sympatric, very similar, although with some noticeable differences (e.g., darkened apex of femora in D. hirtus), and it is likely that both were mixed in the original series as paratypes, which were reported as collected in the "same area" as the holotype (Jolivet et al., Citation2007b). If this interpretation is correct, the correct description of the spermatheca of D. pilosus is as follows (): spermatheca small, hook-shaped, with long, thick cornu, enlarged medially and tapering to blunt apex, connected basally in regular curve with nodulus, narrower and shorter than cornu, weakly concave towards cornu, with blunt conical basal end; slight enlargement before basal narrowing of nodulus and opposite to cornu identifiable as ramus, with attachment of spermathecal gland; spermathecal duct narrowly attached prebasally and opposite to cornu.
Fig. 7. Dorsal (a) and lateral (b) views of the male holotype of Dematotrichus pilosus (Jolivet, Verma & Mille) and accompanying labels (c).

urn:lsid:zoobank.org:act:2CA192B8-0AE1-4121-B439-A6534DAD5A0C
Holotype: male [JGZC-NC796] (), Nouvelle-Calédonie, Prov. Sud, Poya, Mépouiri, Station Beaupré (M. Merzdorf), 21°25.453'S 165°08.807'E, 4 m, 31.vii-5.ix.2007, Malaise trap, St. rech. fruit. Pocquereux staff, Holotype Dematotrichus pubescens sp. nov. J. Gómez-Zurita det. [red label] (JGZC).
Paratypes: (1) 3 males [JGZC-NC798, JGZC-NC838, JGZC-NC839], idem, Paratype Dematotrichus pubescens sp. nov. [red label] (JGZC); (2) 1 male [JGZC-NC797], idem, Paratype Dematotrichus pubescens sp. nov. [red label] (MNHW).
Body slender, elongate elliptical, constricted at base of pronotum, depressed dorsally. Head, pronotum, scutellum, hypomera, base of epipleura, and narrow base of tibiae dark brown; labrum, elytra, most of epipleura, antennae except scape, and ventral surfaces dark chestnut brown; scape, legs and pygidium ochre brown. Length of holotype = 5.1 mm (length range of paratypes: 5.1–5.4 mm); width of holotype = 2.1 mm (width range of paratypes: 2.1–2.3 mm).
Frontoclypeus nearly flat on frons, and clypeus weakly convex basally and deflexed apically; interocular space 1.9× transverse diameter of eyes; surface microreticulate on temples and apically on clypeus, but most of frons and base of clypeus smooth and glossy, with dense moderate punctures as big as intervals, tighter in narrow transition area between frons and clypeus, and abundant long very fine translucent setae. Scape about 1.8× as long as wide; pedicel short, about as long as transverse diameter of scape, slightly longer than wide; antennomeres 3–6 1.4×, 1.5×, 1.9× and 1.9× as long as pedicel; antennomeres 7 and 11 longest, about 2.3× as long as pedicel; antennomeres 8–9 slightly shorter, subequal, some 2.2× as long as pedicel, and antennomere 10 slightly shorter than ninth; antennomeres 3–10 enlarged apically but more markedly in antennomeres 6–8. Surface of pronotum smooth, glossy, with dense, ill-defined punctures as large as punctures on frons, larger than intervals, densely and finely vermiculate, mainly on disc, and abundant long fine nearly translucent pale yellow setae, posteriorly recumbent except near posterior border, bent anteriorly. Anterior border of hypomera short, slightly convex, conspicuously bent externally; surface of hypomera smooth, glossy, with abundant relatively large punctures. Prosternal process narrow at base, about 0.3× as wide as transverse diameter of procoxae, slightly widening posteriorly, markedly enlarged transversely at posterior end, with posterior border narrower than transverse diameter of procoxae; surface slightly rugose. Scutellum nearly as wide at base as long at middle, with sides parallel and regularly round apex; surface with few tiny, imperceptible punctures and tiny fine, translucent setae. Elytra nearly 0.7× as long as body length; surface smooth, glossy, with dense punctures, larger than punctures on pronotum, and intervals slightly wider than punctures, convex, more prominently in lateral and apical declivities, and abundant, uniform clothing of long, posteriorly recumbent, fine nearly translucent pale yellow setae. Epipleura smooth with abundant posteriorly recumbent short fine translucent setae. First abdominal ventrite nearly 1.3× as long as metaventrite, with relatively long anterior process, slightly less than half as long as ventrite, about as long as wide basally, arched anteriorly; ventrites 2–4 combined slightly longer than first ventrite, and fifth ventrite about as long as fourth, widely and shallowly emarginate at apex, with deep elliptic fovea near posterior border medially; surface of ventrites finely shagreened, matt, with sparse fine punctation and fine posteriorly recumbent pale yellow setae, sparser medially. Penis () long, slender, moderately curved ventrally and gradually narrowing apically in basal 2/3, enlarged ventrally at basal level with gonopore with characteristic stepped profile in lateral view; apical third lanceolate, transversally concave dorsally, widening from gonopore and gradually narrowing apically, with tip prolonged as relatively large transverse tooth with angles slightly protruding laterally, acute, and shallowly biconvex in apical border; gonopore small, elliptical, transverse, covered basally by short dorsal flap expanded apically.
Females
Unknown.
Diagnosis
D. pubescens sp. nov. is atypical in the genus presenting nearly concolour tibiae, when most species have conspicuous dark median spots. The only species lacking this trait are the markedly divergent D. capillaris sp. nov. and D. hispidus (Jolivet, Verma & Mille), but these can be easily separated from D. pubescens and every other Dematotrichus by the lack of lateral projections in anterior angles of pronotum, the sparser pubescence on disc of pronotum, and the lack of the characteristic stepped lateral profile of penis. D. pubescens is sympatric with D. hirsutus sp. nov., and apart from the difference in colouration of tibiae, they can be distinguished by the presence of some setae oriented anteriorly or laterally at sides of pronotum near anterior angles in D. hirsutus and all setae oriented posteriorly in the case of D. pubescens.
Derivatio nominis
The name pūbēscēns is the present participle of the verb pūbēscō (ind. pres., 1st, sing.), which, among other things, means to be covered or to be clothed, thus the species name in this case means clothed with hairs, hairy.
Distribution
This species is found together with D. hirsutus sp. nov. in a relatively protected, if tiny (< 70 ha) remnant of dry tropical forest near the south coast, not far from the mouths of the Poya and Moindah Rivers (). This habitat is heavily fragmented along the southwestern dry regions of Grande Terre, and the species may be present in at least nearby patches of this type of forest.
Dematotrichus setosus sp. nov.
()
urn:lsid:zoobank.org:act:A8D57F39-C504-4650-AA12-C20D4FC2EAAC
Holotype: female [JGZC-4856] (), New Caledonia, Col des Roussettes, rainforest nr. refuge, −21.4074 165.5250, 530 m, 2.xii.2010, leg. M. Wanat & R. Ruta, Holotype Dematotrichus setosus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Body elongate elliptical, constricted at base of pronotum, depressed on dorsum. Head dark brown, with blackish frons and bronzy clypeus; pronotum dark brown, with slight purplish reflections and blackish spots at sides of disc; scutellum black with slight bronzy reflection; elytra dark brown with diffuse darker areas around small humps mostly laterally and in apical declivity, humeral angles and base of epipleura; ventral surfaces, including hypomera brown with darkened areas particularly medially on basal abdominal ventrites; labrum, maxillary palpi, antennae, most of legs pale brown with dark brown narrow base of tibiae and relatively broad median rings on protibiae and slightly before middle in meso- and metatibiae. Length: 6.1 mm; width: 2.7 mm.
Frontoclypeus broadly continuous between weak, small antennal calli only indicated anteriorly by fine suture; interocular distance about 1.8× as wide as transverse diameter of eyes; frons flat, finely microreticulate, with moderate punctures generally smaller than interspaces anteriorly and long whitish recumbent setae, radial at end of finely impressed median suture on frons; clypeus broad and flat at base, deflexed in anterior half with moderately emarginate anterior border, microreticulate, with heterogeneous relatively small punctures at base and finer punctures apically, and fine, short translucent setae. Scape about 2.5× longer than wide; pedicel clavate, slightly longer than wide; antennomeres 2–6 progressively longer, with antennomeres 5–6 subequal, nearly 1.7× as long as pedicel; seventh antennomere longest, twice as long as pedicel; antennomeres 8–10 subequal, slightly over 1.8× longer than pedicel, progressively narrower; last antennomere about as long as seventh. Surface of pronotum microreticulate with relatively dense small punctures, smaller than head punctures and with intervals generally larger than diameter of punctures, and sparser larger punctures, similar to punctures on frons, near lateral borders; many punctures with long recumbent pale yellow setae, mostly oriented posteriorly except near posterior border, oriented anteriorly. Surface of hypomera finely microreticulate, with sparse punctures except near inner and outer borders, as large as frontal punctures. Base of prosternal process half as wide as transverse diameter of procoxae, gradually widened behind middle and strongly expanded laterally behind procoxae, wider than transverse diameter of procoxae; surface finely granulose, unpunctured. Scutellum slightly longer than wide at base, regularly round at apex, with few tiny punctures on disc and posteriorly appressed short fine translucent setae. Elytra about 0.7× as long as body; surface finely microreticulate, almost microgranulose, matt, slightly uneven, with rather uniform shallow punctures, as big as punctures at sides of pronotum, and intervals at least 3× larger than diameter of punctures, and numerous tiny setigerous punctures on intervals bearing relatively long, posteriorly recumbent pale yellow setae, tending to form lose tufts at sides and in posterior third of elytra, alternating with glabrous, darker areas; generally shorter setae also present on disc of elytra, along suture and periscutellar area. Epipleura finely microreticulate, unpunctured, with short recumbent setae aligned in single row in posterior half. First abdominal ventrite 1.5× longer than metaventrite and nearly as long (> 0.9×) as ventrites 2–5 combined, with anterior process broadly arched and with thick margins, finely microreticulate, matt, with sparse small, imperceptible punctures and posteriorly recumbent short fine translucent setae; ventrites 2–4 progressively shorter, with similar texture and pubescence as first, but fourth ventrite with premarginal transverse area of denser pubescence at middle; fifth ventrite as long as second, slightly emarginate apically, granulose in apical half, with deep round fovea near apex at middle. Spermatheca () small, comma-shaped, with thick short, weakly curved and apically blunt cornu, slightly narrowed at base and connected at nearly right angle with narrower and shorter, straight nodulus, round basally; ramus visible as prebasal enlargement of nodulus, opposite to cornu, with large attachment of spermathecal gland; spermathecal duct attached prebasally, opposite to ramus, thick, shortly cylindrical at base, elbowed and strongly narrowed to membranous, thin duct.
Males
Unknown.
Diagnosis
See the diagnosis of D. capillosus sp. nov. for the characteristics of the group where D. setosus sp. nov. belongs and how to differentiate these species. D. setosus has relatively uniform primary and secondary punctation on elytra (unlike D. villosus sp. nov.), microreticulate, matt scutellum (unlike D. crinitus sp. nov.), fine sparse punctation on disc (unlike D. pilosus (Jolivet, Verma & Mille)), and strong, relatively uniform punctures on hypomera (unlike D. capillosus).
Derivatio nominis
The species name is a Latin adjective (m.) that means bristly, covered with setae, in reference to the hairy dorsum of the species.
Distribution
The single specimen currently known for this species was collected in rainforest in the mid elevation area of Col des Roussettes, in the central mountain chain of Grande Terre ().
urn:lsid:zoobank.org:act:91895179-FB1F-43BF-822C-27721DD7B9B7
Holotype: male [JGZC-4853] (), New Caledonia, Farino, Parc des Grandes Fougères, Camp de la Houe, −21.61176 165.75406, 400 m, 13.xi.2008, leg. M. Wanat, Holotype Dematotrichus villosus sp. nov. J. Gómez-Zurita det. [red label] (MNHW).
Paratypes: female [JGZC-NC850] (), Nouvelle-Calédonie, Col d'Amieu, 21°34.922'S 165°46.324'E, 630 m, 24.viii-20.ix.2007, Malaise trap, St. rech. fruit. Pocquereux staff, Paratype Dematotrichus villosus sp. nov. J. Gómez-Zurita det. [red label] (JGZC); female, New Caledonia (S), Col d’Amieu, 6.5–7.0 km to gate, −21.5868 165.7738, 450 m, 14.xi.2008, M. Wanat leg. (MNHW).
Body elongate elliptical, slender, constricted at base of pronotum, depressed dorsally. Head, pronotum, scutellum, hypomera, most of thoracic ventrites, trochanters, base of epipleura and most of abdominal ventrites 1–3 dark brown, with slight purplish hue mostly on head, pronotum and scutellum; elytra slightly paler with some darker areas with similar purplish hue, mainly along suture; labrum, antennae, most of legs, abdominal ventrites 4–5 and pygidium brown, with base of most antennomeres, narrow base of tibiae and small median spot on tibiae dark brown. Length of holotype = 6.4 mm (length of paratypes = 7.2 mm); width of holotype = 2.7 mm (width of paratypes = 3.2 mm).
Frontoclypeus feebly convex on frons with weakly impressed median suture, slightly depressed between supraantennal calli, with interocular distance about twice as wide as transverse diameter of eyes and clypeus deflexed in apical 2/3; clypeus shorter than wide at apex, widely but not deeply emarginate apically; surface tightly and markedly microreticulate, alutaceous, with dense small punctures as big as intervals, larger and tighter around supraanntennal calli and sparse dishevelled long golden setae. Scape nearly half as wide as long; pedicel enlarged preapically, longer than wide; antennomeres 3–7 1.5×, 1.7×, 2.1×, 2.2× and 2.3× longer than pedicel, respectively; antennomeres 8–10 subequal, about as long as fifth; 11th antennomere slightly shorter than 10th. Surface of pronotum very densely microreticulate, matt, with dense fine punctures on disc as big as small punctures on frons, with intervals as big or slightly larger than punctures, and sparser, larger punctures at sides, with abundant recumbent, long golden setae, mainly oriented posteriorly except near posterior border and sides posteriorly, with anterior or lateral orientation, and scattered setae on disc with different orientations. Surface of hypomera finely alutaceous, with abundant moderate punctures except near lateral margin. Prosternal process narrow at base, half as wide as diameter of procoxae, with sides gradually widening posteriorly and strongly expanded laterally behind; surface of process uneven, microsculptured. Scutellum slightly longer than wide at base, with sides nearly parallel and arched apex; surface with several small punctures. Elytra nearly 0.7× as long as body; surface densely microreticulate as on pronotum, matt, with abundant primary punctures slightly larger than punctures on pronotum separated by large intervals, and more numerous secondary, smaller punctures tending to cluster around primary punctures, and long posteriorly recumbent golden setae relatively dense at lateral and apical declivities and sparser on disc. Epipleura finely microreticulate, covered with abundant short recumbent pale yellow setae. First abdominal ventrite longer than metaventrite, markedly transverse posteriorly, with anterior process subtrapezoidal and thickly margined; surface finely shagreened, with abundant small punctures and long posteriorly recumbent pale yellow setae; ventrites 2–4 combined as long as first ventrite, each individually of similar length, strongly transverse, finely shagreened, with fine punctures and posteriorly recumbent fine pale yellow setae; fifth ventrite as long as fourth, widely emarginate apically, with deep median fovea close to apical border. Penis () long, slender, with long tubular base, gradually narrowing apically and weakly curved ventrally in basal half, regularly curved apically before gonopore with distal end of penis nearly perpendicular to base in lateral view; apex elongate and lanceolate after gonopore, transversally concave dorsally, and markedly enlarged ventrally relative to basal tubular part of penis; tip of penis with subrectangular, transverse median tooth, slightly acute at angles; gonopore small, basal to lanceolate apex, covered basally by small subquadrate dorsal flap.
Females
Spermatheca () small, with cornu thicker at base, gradually tapering to slightly curved blunt apex; nodulus tubular, straight, as wide as base of cornu, connected to cornu at right angle; spermathecal duct connected prebasally to nodulus oriented towards cornu, nearly as thick as nodulus basally and gradually tapering to narrower duct curved towards nodulus; ramus short, annular, prebasal to nodulus and opposite to cornu.
Diagnosis
The characteristics of elytral punctation of D. villosus sp. nov., presenting secondary punctures tending to cluster around primary punctures on disc on a tight microgranulose background, are unique and should allow recognizing this species at once. Among the species in its group (ringed tibiae, elongate basitarsomeres and microreticulated dorsum), seems to be the only one with relatively uniform dense punctation on pronotum, and it also has sparser, shorter setae on disc of elytra.
Derivatio nominis
The specific epithet for this species, villōsus, is an adjective (m.) derived from the Latin noun villus (m.), which can be translated as shaggy hair in English, thus meaning hairy or shaggy.
Distribution
The three known specimens of D. villosus sp. nov. were collected in nearby localities in the Central Massif, within the reserve of Col d'Amieu ().
Identification key of species of Dematotrichus
Anterior angles of pronotum inconspicuous in dorsal view and posterior angles weakly protruding; colour of legs relatively uniform; ventral curvature of penis regular, without stepped profile basal to gonopore.2
-. Anterior angles of pronotum protruding laterally, clearly visible in dorsal view, and posterior angles of pronotum prominent, as lateral expansions of posterior border of pronotum; legs bicolour, usually with dark median spot on all tibiae (except in D. pubescens sp. nov.).3
Body shape stockier, with elytra about 0.6× as long as body; surface of elytra rather uniform and punctures fine and shallow.D. capillaris sp. nov.
-. Body shape more slender, with elytra about 0.65× as long as body; surface of elytra uneven, slightly corrugated and punctation conspicuous.D. hispidus (Jolivet, Verma & Mille)
Tibiae concolour or slightly darkened basally.D. pubescens sp. nov.
-. All tibiae with dark median spot (perhaps weakly developed in some individuals of D. hirsutus sp. nov.).4
Apex of femora largely dark; first metatarsomere shorter than tarsomeres 2–3 combined.5
-. Apex of femora concolour; first metatarsomere slender, as long or longer than tarsomeres 2–3 combined.6
Dorsum densely pubescent, with head comparatively appearing nearly glabrous.D. comatulus sp. nov.
-. Dorsum more sparsely pubescent, with head similarly clothed as pronotum.D. hirtus sp. nov.
Teguments of pronotum and elytra with conspicuous microreticulation.7
-. Teguments of pronotum and particularly of elytra smooth and glossy.11
Elytra with sparse small primary punctures and smaller secondary punctures tending to cluster around primary punctures.D. villosus sp. nov.
-. Elytra without or with sparse irregular secondary punctation.8
Microreticulation on pronotum and elytra fine and superficial, producing a silkier shine to dorsal surfaces; scutellum microreticulate but glossy.D. crinitus sp. nov.
-. Microreticulation on pronotum and elytra strong, dense, producing a gritty shine to dorsal surfaces; scutellum finely microreticulated, but not glossy.9
Tiny, superficial punctures on disc of pronotum, generally smaller than intervals.10
-. Small but bigger and deeper punctures on disc of pronotum, about as big as intervals.D. pilosus (Jolivet, Verma & Mille)
Hypomera with scattered deep punctures throughout; dark spots in pro- and metatibiae shorter, spanning some 3–4 setae.D. setosus sp. nov.
-. Hypomera with sparse shallow punctures in basal half; dark spot in pro- and metatibiae longer, spanning some 5–6 setae.D. capillosus sp. nov.
Punctures on pronotum dense, but intervals on average as big as punctures; elytral surface on disc relatively even, with punctures about as big as punctures on pronotum.D. horridus sp. nov.
-. Punctures on pronotum very dense, larger than intervals; elytral surface on disc uneven, lumpy, with punctures confused between convex intervals generally larger than punctures on pronotum.12
Dorsal setae golden to yellow and denser on elytra.D. comans sp. nov.
-. Dorsal setae pale yellow to whitish and sparser on elytra.D. hirsutus sp. nov.
Associate Editor: Dr Dimitar Dimitrov
Supplemental Material
Download PDF (503.8 KB)Acknowledgements
Fieldwork was greatly facilitated by Dr Christian Mille (Institute Agronomique néo-Calédonien, La Foa, New Caledonia) and his help and friendship are always valued. Important samples that have been recognized as new species were obtained from Dr Marek Wanat (MHNW, Wroclaw, Poland), and his support is once again warmly thanked for allowing me to study the New Caledonian Eumolpinae of his magnificent collection. Anabela Cardoso (IBB, CSIC, Barcelona) and Maria Pàmies helped producing DNA sequence data used in this study. Antoine Mantilleri and Maxime Le Cesne (MNHN, Paris, France) kindly helped in providing high-quality pictures of the generic type, Dematochroma pilosa Jolivet, Verma & Mille, giving me a solid basis to make all the relevant decisions in this revision. Dr D. Dimitrov and two anonymous reviewers made very useful comments to prepare the final version of the manuscript and their kind, constructive and encouraging manners are especially acknowledged.
Supplemental Material
Supplemental material for this article may be accessed here: https://doi.org/10.1080/14772000.2022.2084471.
Additional information
Funding
References
- Baly, J. S. (1864). Descriptions of New Genera and Species of Phytophaga. J. S. Baly, Stationer's Hall.
- Brower, A. V. Z., DeSalle, R., & Vogler, A. P. (1996). Gene trees, species trees, and Systematics: a cladistic perspective. Annual Review of Ecology and Systematics, 27(1), 423–450. https://doi.org/10.1146/annurev.ecolsys.27.1.423
- Brown, R. W. (1954). Composition of Scientific Words. A manual of methods and a lexicon of materials for the practice of logotechnics (pp. 882). George W. King Printing Co.
- Chapuis, F. (1874). Genera des Coléoptères ou exposé méthodique et critique de tous les genres proposés jusqu’ici dans cet ordre d’insectes. Tome X. Famille des Phytophages. Librairie Encyclopédique de Roret.
- Chevrolat, A. (1836). Chalcoplacis. In P. F. M. A. Dejean (Ed.), Catalogue des Coléoptères de la collection de M. le Comte Dejean (pp. 409). Livraison 5. Méquignon-Marvis.
- Coyne, J. A., & Orr, H. A. (2004). Speciation. Sinauer Associates, Sunderland MA.
- Farris, J. S. (1979). The information content of the phylogenetic system. Systematic Zoology, 28(4), 483–519. https://doi.org/10.2307/2412562
- Funk, D. J., & Omland, K. E. (2003). Species-level paraphyly and polyphyly: frequency, causes, and consequences, with insights from animal mitochondrial DNA. Annual Review of Ecology. Annual Review of Ecology, Evolution, and Systematics, 34(1), 397–423. https://doi.org/10.1146/annurev.ecolsys.34.011802.132421
- Gómez-Zurita, J. (2018). Description of Kumatoeides gen. nov. (Coleoptera: Chrysomelidae, Eumolpinae) from New Caledonia. Zootaxa, 4521(1), 89–115. https://doi.org/10.11646/zootaxa.4521.1.4
- Gómez-Zurita, J., & Cardoso, A. (2014). Systematics of the New Caledonian endemic genus Taophila Heller (Coleoptera: Chrysomelidae, Eumolpinae) combining morphological, molecular and ecological data, with description of two new species. Systematic Entomology, 39(1), 111–126. https://doi.org/10.1111/syen.12038
- Gómez-Zurita, J., & Pàmies-Harder, M. (2022). Phylogenetic restitution and taxonomic revision of the New Caledonian endemic genus Thasycles Chapuis (Coleoptera: Chrysomelidae, Eumolpinae). Zoologischer Anzeiger, 297, 16–41. https://doi.org/10.1016/j.jcz.2022.01.003
- Gómez-Zurita, J., Platania, L., & Cardoso, A. (2020). A new species of the genus Tricholapita nom. nov. and stat. nov. (Coleoptera: Chrysomelidae, Eumolpinae) from New Caledonia. Zootaxa, 4857, 85–94.
- Heller, K. M. (1916). Die Käfer von Neu-Caledonien und den benachbarten Inselgruppen. In F. Sarasin & J. Roux (Eds.), Nova Caledonia, Zoologie (Vol. II, pp. 229–364). L. III. C. W. Kreidels Verlag, Wiesbaden.
- Hollister, J., Shah, T., Robitaille, A. L., Beck, M. W., Johnson, M. (2022). elevatr: access elevation data from various APIs. https://github.com/jhollist/elevatr.
- Jolivet, P., Verma, K. K., & Mille, C. (2007a). New genera and species of Eumolpinae from New Caledonia (Coleoptera, Chrysomelidae). Revue Française D'Entomologie, 29, 77–92.
- Jolivet, P., Verma, K. K., & Mille, C. (2007b). New species of Eumolpinae from the genera Dematochroma Baly, 1864 and Taophila Heller, 1916 from New Caledonia (Coleoptera, Eumolpidae). Revue Française D'Entomologie, 29, 33–47.
- Jolivet, P., Verma, K. K., & Mille, C. (2009). ]. Eumolpinae recently collected in New Caledonia and Vanuatu (Coleoptera, Chrysomelidae). Nouvelle Revue D'Entomologie (N.S.), 26, 3–17.
- Jolivet, P., Verma, K. K., & Mille, C. (2010). Eumolpinae recently collected in New Caledonia: a consideration of local geographic intraspecific variability in Bohumiljania humboldti (Coleoptera, Chrysomelidae). Revue Française D'Entomologie, 32, 141–149.
- Jolivet, P., Verma, K. K., & Mille, C. (2013). New and known leaf beetles collected in Mont Panié in 2010 (Coleoptera, Chrysomelidae, Eumolpinae). Nouvelle Revue D'Entomologie (N.S, ), 29, 141–154.
- Katoh, K., & Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution, 30, 772–780.
- Laporte, F. L. (1833). Mémoire sur les divisions du genre Colaspis. Revue Entomologique, 1, 18–25.
- Lawrence, J. F., Beutel, R. G., Leschen, R. A. B., & Slipinski, A. (2010). Glossary of morphological terms. In R. A. B. Leschen, R. G. Beutel, & J. F. Lawrence (Eds.), Handbook of Zoology. Arthropoda: Insecta. Part 38. Coleoptera, Beetles. Vol. 2.Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia Partim). Walter de Gruyter, Berlin. pp. 9–20.
- Li, W.-H. (1997). Molecular Evolution. Sinauer Associates.
- Lindroth, C. H. (1957). The principal terms used for male and female genitalia in Coleoptera. Opuscula Entomologica, 22, 241–256.
- Papadopoulou, A., Cardoso, A., & Gómez-Zurita, J. (2013). Diversity and diversification of Eumolpinae (Coleoptera: Chrysomelidae) in New Caledonia. Zoological Journal of the Linnean Society, 168(3), 473–495. https://doi.org/10.1111/zoj.12039
- Platania, L., Cardoso, A., & Gómez-Zurita, J. (2020). Diversity and evolution of new Caledonian endemic Taophila subgenus Lapita (Coleoptera: Chrysomelidae: Eumolpinae). Zoological Journal of the Linnean Society, 189(4), 1123–1154. https://doi.org/10.1093/zoolinnean/zlz119
- Platania, L., & Gómez-Zurita, J. (2022). Integrative taxonomic revision of the New Caledonian endemic genus Taophila Heller (Coleoptera: Chrysomelidae, Eumolpinae). Insect Systematics and Evolution, 53, 111–184.
- R Core Team. (2021). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
- Samuelson, G. A. (2010). Review of Taophila, a genus endemic to New Caledonia (Coleoptera: Chrysomelidae: Eumolpinae). Zootaxa, 2621(1), 45–62. https://doi.org/10.11646/zootaxa.2621.1.3
- Samuelson, G. A. (2015). Acronymolpus, a new genus of Eumolpinae, endemic to New Caledonia (Coleoptera, Chrysomelidae). ZooKeys, 547, 93–102. https://doi.org/10.3897/zookeys.547.9698
- Sharp, D. (1886). On New Zealand Coleoptera, with descriptions of new genera and species. Scientific Transactions of the Royal Dublin Society, 3, 351–452.
- Shaw, K. L. (1998). Species and the diversity of natural groups. In D. J. Howard, & S. H. Berlocher (Eds.), Endless Forms. Species and Speciation (pp. 44–56). Oxford University Press.
- Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics (Oxford, England), 22(21), 2688–2690. https://doi.org/10.1093/bioinformatics/btl446
- Wagner, T. (2007). Monolepta Chevrolat, 1837, the most speciose galerucine taxon: redescription of the type species Monolepta bioculata (Fabricius, 1781) and key to related genera. Journal of Natural History, 41(1-4), 81–100. https://doi.org/10.1080/00222930601127384
- Wickham, H. (2016). ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New York. https://ggplot2.tidyverse.org.
- Zhang, J., Kapli, P., Pavlidis, P., & Stamatakis, A. (2013). A general species delimitation method with applications to phylogenetic placements. Bioinformatics (Oxford, England), 29(22), 2869–2876.