Abstract
The Early Cretaceous Jehol Biota of China includes seven named lizard taxa: Yabeinosaurus tenuis and Dalinghosaurus longidigitus; the gliding Xianglong; more fragmentary remains referred to the genera Pachygenys and Mimobecklesisaurus; and the undiagnostic juvenile skeletons Jeholosaurus and Liaoningolacerta. Here we describe new lizard remains from the Yixian Formation at Liutiaogou, Ningcheng County, Inner Mongolia. The eight specimens vary in size and ontogenetic development, but appear to pertain to a single species. Several specimens show unparalleled preservation of soft tissue impressions including scalation and pigmentation, claw sheaths, sternal and extrastapedial cartilages, as well as rarely preserved skeletal elements like orbitosphenoids and postcloacal bones. The combination of characters precludes attribution to known taxa both within and outside China, and the new lizard is named Liushusaurus acanthocaudata gen. et sp. nov. Cladistic analysis supports the placement of Liushusaurus within the Scincogekkonomorpha, as the sister group of Scleroglossa.
Introduction
Deposits of the Jehol Group of northeastern China have yielded numerous lizard remains, most of which can be attributed to one of two taxa: Yabeinosaurus tenuis CitationEndo & Shikama, 1942 (CitationJi et al. 2001; CitationEvans et al. 2005) and Dalinghosaurus longidigitus CitationJi, 1998 (CitationJi & Ji 2004; CitationEvans & Wang 2005; CitationEvans et al. 2007). Three other genera have been named on the basis of juvenile specimens: Jeholacerta CitationJi & Ren, 1999, Liaoningolacerta CitationJi, 2005 and Xianglong CitationLi et al., 2007. Although its skull is poorly preserved, Xianglong can be distinguished on the basis of the elongate gliding ribs. Unfortunately, the same is not true for either Jeholacerta or Liaoningolacerta. The type and only specimen of the first is immature and consists of a skin impression with a few problematically interpreted skull impressions. The diagnosis lacks any derived features to distinguish Jeholacerta from known taxa or to form the basis of a referral of new material. Liaoningolacerta is almost equally problematic. The type and only specimen appears to preserve some skeletal remains, but its extreme immaturity renders the existing interpretation of cranial morphology problematic, and the diagnosis is a mixture of ontogenetic observations (e.g. unfused astragalus and calcaneum) and generalised lizard characters (moderate expansion of scapula), making attribution of adult specimens to the same genus impossible. Both Jeholacerta and Liaoningolacerta must be regarded as nomina dubia. Two further lizards have been named on the basis of fragmentary remains from deposits that may be contemporaneous with the Jehol Biota (CitationWang & Evans 2006a, Citation2006b; Wang & Li 2008). These are Mimobecklesisaurus CitationLi, 1985, based on a partial skull and osteoderms from Gansu, and Pachygenys CitationGao & Cheng, 1999 from Shandong (lower jaw specimens).
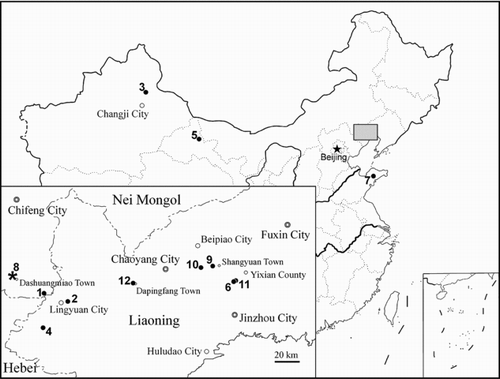
Here we describe new lizard material from the Yixian Formation at Liutiaogou, Nei Mongol (Inner Mongolia) (). There are eight individuals, ranging in age from near-hatchling to mature. Allowing for ontogenetic variation, body shape, limb proportions, scalation and skull morphology suggest they represent a single new taxon.
Figure 1 Localities (in alphabetical order) of Jurassic and Early Cretaceous lizards in China. * denotes locality of Liushusaurus acanthocaudata (8, Liutiaogou). An enlargement of the grey area is shown in the lower left corner. Jurassic localities: 1. Daohugou, Ningcheng County, Nei Mongol (Inner Mongolia): Squamata gen. et sp. indet. 1 (CitationEvans & Wang 2007) & 2 (CitationEvans & Wang 2009), ?Late Jurassic (YW); 2. Gezidong (Ketzutung), Lingyuan City, Liaoning: ‘Yabeinosaurus’ youngi (Hoffstetter 1964; regarded as questionable by Wang & Li 2008), ?Middle Jurassic; 3. Wucaiwan, Changji City, Xinjiang: Squamata gen. et sp. nov. (YW, work in progress), Middle/Late Jurassic. Localities of the Lower Cretaceous Yixian Formation: 4. Dawangzhangzi, Lingyuan City, Liaoning: Dalinghosaurus longidigitus, Yabeinosaurus tenuis (CitationEvans & Wang 2005; CitationEvans et al. 2005); 5. Hongliugedajing, Subei County, Gansu: Mimobecklesisaurus gansuensis (CitationLi 1985); 6. Jingangshan, Yixian County, Liaoning: Yabeinosaurus tenuis (CitationJi et al. 2001; CitationEvans et al. 2005); 7. Laiyang City, Shandong: Pachygenys thlastesa (CitationGao & Cheng 1999), Doushan Formation (lateral equivalent of Yixian Formation, personal communication with Wang X. L.); 8. Liutiaogou, Ningcheng County, Nei Mongol (Inner Mongolia): Liushusaurus acanthocaudata (this paper); 9. Lujiatun, Beipiao City, Liaoning: Dalinghosaurus longidigitus (CitationEvans et al. 2007); 10. Sihetun, Beipiao City, Liaoning: Dalinghosaurus longidigitus, Yabeinosaurus tenuis (CitationEvans & Wang 2005; CitationEvans et al. 2005); 11. Zaocishan (Tsaotzushan), Yixian County, Liaoning: Yabeinosaurus tenuis (specimen lost, CitationEndo & Shikama 1942; Wang & Li 2008). Locality of the Lower Cretaceous Jiufotang Formation: 12. Dapingfang, Chaoyang City, Liaoning: Yabeinosaurus tenuis (CitationEvans et al. 2005).
Geological background
All of the material described here comes from a locality close to Liutiaogou Village, Dashuangmiao Town, Ningcheng County, Chifeng City, Nei Mongol (Inner Mongolia) (). The beds are referred to the Yixian Formation, Jehol Group, Lower Cretaceous (CitationWang et al. 2000). This locality has also yielded new gomphaeschnid dragonflies (B. L. CitationZhang et al. 2008) and chresmodid insects (X. W. CitationZhang et al. 2008), exceptionally well-preserved material of a new lamprey (CitationChang et al. 2006), a juvenile pterosaur (CitationLü 2009), as yet undescribed bird and mammal material, and more typical elements of the Yixian biota including the fish Lycoptera and acipenseriforms (Peipiaosteus and Protopsephurus). Precise correlation of the horizon with those in Liaoning has yet to be made, but it is thought to be equivalent to either the Jianshangou or Dawangzhangzi bed (c. 124–122 Ma) of the Yixian Formation (X. L.Wang, pers. comm., July 2006).
Systematic palaeontology
Squamata CitationOppel, 1811
Genus Liushusaurus gen. nov.
Type species. Liushusaurus acanthocaudata gen. et sp. nov.
Diagnosis. As for type species.
Derivation of name. Liushu, Chinese, willow tree (derived from “Liutiao”, willow leaf, of Liutiaogou, meaning “willow leaf valley” where the fossils were excavated); sauros, Greek, reptile.
Occurrence. As for type species.
Liushusaurus acanthocaudata sp. nov.
(–)
Diagnosis. A medium-sized (SPL = 66 mm in holotype) Early Cretaceous lizard. Differs from the Yixian lizards Yabeinosaurus (e.g. CitationEvans et al. 2005) and Dalinghosaurus (CitationJi 1998; CitationJi & Ji 2004; CitationEvans & Wang 2005) in having relatively longer forelimbs; differs from Dalinghosaurus in having more than six cervical vertebrae; differs from Yabeinosaurus in reaching skeletal maturity at a smaller size, in having expanded, hook-like (perforate) clavicles, and in having a proportionally shorter presacral region; differs from Xianglong (CitationLi et al. 2007) in lacking elongated gliding ribs; differs from the Japanese Early Cretaceous Kuwajimalla (CitationEvans & Manabe 2008) in lacking multicuspid teeth; and differs from both Yabeinosaurus and the Japanese Early Cretaceous Sakurasaurus (CitationEvans & Manabe 2009) in lacking co-ossification of the postorbital and postfrontal and restriction of the upper temporal fenestra. Resembles Yabeinosaurus (CitationEvans et al. 2005) and differs from Dalinghosaurus longidigitus (CitationEvans & Wang 2005) and Late Jurassic German Bavarisaurus (CitationEvans 1994a) in having proportionally shorter feet; differs from Mimobecklesisaurus (Early Cretaceous, China, CitationLi 1985) and other Jurassic-Cretaceous paramacellodids (CitationHoffstetter 1967; CitationEvans & Chure 1998) in lacking a body covering of rectangular osteoderms; differs from Jeholacerta (CitationJi & Ren 1999), Yabeinosaurus, and from the Early Cretaceous Lebanese Baabdasaurus (CitationArnold et al. 2002) in the elongate, spiky scales on the tail; differs from the Italian Early Cretaceous Chometokadmon (CitationEvans et al. 2006) in having a much shorter parietal and a relatively shorter presacral body length; resembles the Late Jurassic German Ardeosaurus (CitationMateer 1982; CitationEstes 1983) and differs from the Jurassic–Cretaceous Eichstaettisaurus (CitationEvans et al. 1999, Citation2004) and the Early Cretaceous Iberian Meyasaurus (CitationEvans & Barbadillo 1997) in having a slightly restricted upper temporal fenestra, loss of parietal foramen, and fewer teeth in the maxilla; resembles the Late Cretaceous Mongolian ‘mongolochamopines’ (CitationAlifanov 2000) in having paired frontals, loss/reduction of the parietal foramen, and a short parietal, but differs in having proportionally shorter frontals and therefore a shorter, more rounded antorbital skull; differs from Meyasaurus, Dicrodon and Bicuspidon (CitationNydam & Cifelli 2002a, mid-Cretaceous, USA), Atokosaurus (CitationNydam & Cifelli 2002b, mid-Cretaceous, USA), and the ‘mongolochamopines’ Mongolochamops and Altanteius (CitationAlifanov 2000, Upper Cretaceous, Mongolia) in having conical rather than cuspidate teeth; differs from the Early Cretaceous Spanish Hoyalacerta (CitationEvans & Barbadillo 1999) in having fewer, larger teeth and proportionally longer limbs; differs from the Early Cretaceous Mexican Huehuecuetzpalli (CitationReynoso 1998) in the retention of a large free postfrontal, the absence of a parietal foramen, paired rather than fused frontals, and the absence of an elongated premaxilla; differs from Bavarisaurus (CitationEvans 1994a) and the Spanish Early Cretaceous Scandensia (CitationEvans & Barbadillo 1998) in having well-developed vertebral condyles, and from the latter in having a cruciform rather than rhomboid interclavicle and in rib morphology (narrow vs expanded ribs); differs from the Late Jurassic German Ardeosaurus (CitationMateer 1982), British Early Cretaceous Parasaurillus (CitationEvans & Searle 2002), and Dalinghosaurus (CitationEvans & Wang 2005) in lacking obvious sculpture on the skull bones, even in mature specimens; differs from the Jurassic–Cretaceous Parviraptor (CitationEvans 1994b) in having a short single parietal, rather than paired elongate bones, and in lacking a parietal foramen; and differs from the Early Cretaceous Dorsetisaurus (CitationHoffstetter 1967) in lacking well-developed sculpture and in tooth morphology. Comparison with Pachygenys (CitationGao & Cheng 1999) is difficult as the latter is represented only by two, albeit very distinctive, lower jaws with eight teeth clustered in the anterior part of the dentary. The lower jaw dentition is not clearly visible in any of the Liushusaurus specimens, but the presence of 13 maxillary teeth distributed evenly along the bone renders it unlikely that they occluded with a lower jaw dentition like that of Pachygenys. Comparison with other taxa based on dentary specimens is similarly problematic, but Liushusaurus differs from the British Early Cretaceous Pseudosaurillus (CitationHoffstetter 1967; CitationEvans & Searle 2002) in having larger teeth (very small in relation to jaw height in Pseudosaurillus) and from the Jurassic-Cretaceous Euramerican Saurillodon (CitationSeiffert 1973) in jaw and tooth proportions (short robust dentary, larger recurved teeth in Saurillodon).
Derivation of name. From: acanthus, Latin, spine; cauda, Latin, tail, in reference to the appearance of the tail scalation.
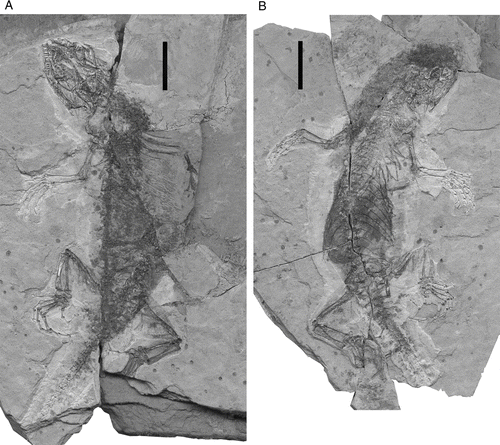
Holotype. IVPP V15587A/B (vertebrate collection number of Institute of Vertebrate Paleontology and Paleoanthropology (IVPP), Beijing), part and counterpart of a young adult specimen, snout-pelvic length c. 66 mm ().
Figure 2 Holotype specimen (IVPP V15587) of Liushusaurus acanthocaudata gen. et sp. nov., from the Liutiaogou locality, Inner Mongolia. A, part and B, counterpart. Scale bars = 10 mm.
Paratype. IVPP V14715A/B (partial skull and postcranium, ), showing well-preserved scale pattern, notably the tail scalation that forms the basis of the specific name.
Figure 3 Additional specimens of Liushusaurus acanthocaudata gen. et sp. nov. from the type locality. A, IVPP V15011; B, IVPP V14715; C, IVPP V15508; D, IVPP V14716; E, IVPP V14716, detail of pelvis; F, IVPP V14746, detail of vertebrae. All scale bars = 10 mm, except E = 5 mm.

Other material. IVPP V14716 (incomplete postcranial skeleton with skin traces and post-autotomy tail replacement, ); IVPP V14746A/B (part and counterpart of incomplete postcranial skeleton, ); IVPP V15507 A/B (isolated hind limb with fine scalation, ); IVPP V15508 A/B (complete skeleton of very young, hatchling or near hatchling, animal, ); IVPP V15011 (adult individual, ); IVPP V15586A/B (skeleton and fine skin impressions from subadult individual, with post-autotomy tail replacement, ). All specimens are from the type locality.
Figure 4 Liushusaurus acanthocaudata gen. et sp. nov., IVPP V15586, main block. A, skull; B, whole skeleton. Scale bars 1 mm and 10 mm respectively.
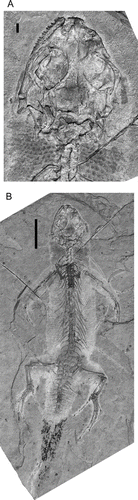
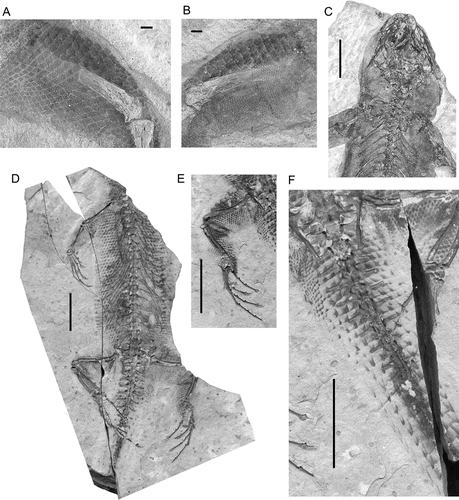
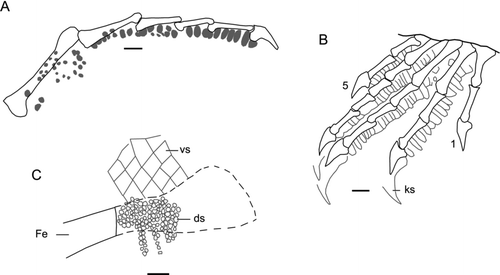
Figure 9 Liushusaurus acanthocaudata gen. et sp. nov., scalation. A and B, part and counterpart of IVPP V15507, an isolated hind limb showing rhomboid ventral scales (A) and rhomboid scales above the femur and fine dorsal scales below it (B); C, IVPP V15508B showing dorsal scalation (small granular scales); D–F, IVPP V14715, showing ventral scalation; D, counterpart block (IVPP V14715B); E, hind limb from main block (IVPP V14715A); F, tail on IVPP V14715A, showing elongate spiny scales. Scale bars: A, B = 1 mm; C = 5 mm; D-F = 10 mm.
Occurrence. Liutiaogou Village, Dashuangmiao Town, Ningcheng County, Chifeng City, Nei Mongol (Inner Mongolia) (); Lower Cretaceous, Jehol Group, Yixian Formation.
Remarks. Two juvenile lizard skeletons (CitationEvans & Wang 2007, Citation2009) have recently been described from the Daohugou fossil bed of Inner Mongolia and neighbouring areas, associated with a diverse caudate fauna (CitationWang 2004; CitationWang & Rose 2005; Wang & Evans 2006b). The bed is probably of Upper Jurassic age based on composite lithological and palaeontological data, although J2, J3, and K1 ages have all been proposed and there is currently no consensus. Liushusaurus differs from the first Daohugou lizard (CitationEvans & Wang 2007) in having proportionally longer forelimbs (relative to the hind limbs), shorter hind limbs (relative to snout-pelvis length, SPL) and a shorter tibia (relative to the femur). This is also true for the second Daohugou lizard (CitationEvans & Wang 2009), which also has a markedly longer femur, tibia and foot (to SPL) (). Liushusaurus further differs from the second Daohugou lizard in having broader body shape and shorter preorbital region, in contrast to the slender body and relatively longer preorbital region of the latter.
Table 1 Comparative limb proportions in two specimens of Liushusaurus acanthocaudata gen. et sp. nov. (V15587, V15508) and two Daohugou lizards (V14386, V13747). DHG, Daohugou lizard; Fe, femur; FL, forelimb length; Ft, foot; Hd, head; HL, hind limb length; Hu, humerus; SPL, snout-pelvic length; Ti, tibia. SPL in mm; others in % proportions.
Description
Skull
The skull of Liushusaurus is complete only on IVPP V15586 and V15587 (), but IVPP V14715, V15011 and the juvenile IVPP V15508 provide supplementary information (, ). However, much of the skeleton is preserved in impression and the description that follows is based primarily on high definition peels. This limits the level of fine detail available, but larger specimens suggest the presence of weak sculpture on the frontals and parietals.
Figure 5 Liushusaurus acanthocaudata gen. et sp. nov., skull morphology drawn from silicon peels of A, IVPP V15587A and B, IVPP V15586A. The grey areas in (A) (basicranium and hyoid elements) have been superimposed from the counterpart block. Scale bars = 1 mm. Abbreviations: An, angular; Ar, articular; Br, braincase; C, coronoid; D, dentary; E, epipterygoid; Eb, epibranchial; Ec, ectopterygoid; F, frontal; Hy, hyoid; J, jugal; L, lacrimal; LJ, lower jaw; Mx, maxilla; N, nasal; Op, opisthotic; P, parietal; Pfr, postfrontal; Pmx, premaxilla; Po, postorbital; Prf, prefrontal; Pt, pterygoid; Q, quadrate; Sp, sphenoid; Sq, squamosal; St, supratemporal; Su, surangular.
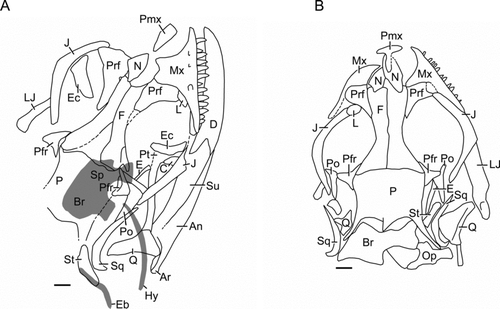
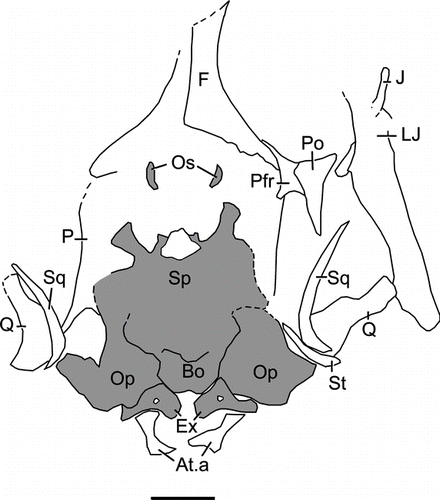
Figure 6 Liushusaurus acanthocaudata gen. et sp. nov. Skull of IVPP V15508, juvenile, with dorsal and ventral impressions superimposed. Braincase in grey. Scale bar = 1 mm. Abbreviations as in and: At.a, atlantal arches; Bo, basioccipital; Ex, exoccipital; Op, opisthotic; Os, orbitosphenoid.
The nasals are paired and short, making the skull short and rather rounded in the antorbital region. The prefrontal is large. The frontals are parallel-sided for the anterior two-thirds of their length and then expand at the fronto-parietal suture. That suture appears straight in young adults like IVPP V15586, but in larger specimens (e.g. IVPP V15011) is more irregular. The parietal is unpaired and lacks a parietal foramen. Its shape varies between immature (e.g. IVPP V15586, short and broad, with emarginations on either side of the midline) and mature (e.g. IVPP V15011, V15587, longer, more U-shaped posterior margin). The postparietal processes lengthen with maturity and are long and divergent in IVPP V15587. The pit for the processus ascendens of the braincase lies forward of posterior parietal margin. The premaxilla is not well-preserved in any specimen but IVPP V15586 () shows that it is single, broad in its alveolar portion, and has a narrow nasal process. The maxilla is rather short (∼13 teeth) with a tall anterior facial process and a short posterior process that is excluded from the orbit by the prefrontal and the jugal. The latter biramous bone reaches the anterior margin of the orbit, forms its ventral rim, and then ascends to form most of the postorbital margin, restricting the entry of the postorbital. Between the jugal and the prefrontal in both IVPP V15586 and V15587 () there is a small fragment of bone that is probably a lacrimal. The postfrontal and postorbital are separate, with the postfrontal spanning the frontoparietal junction. The postorbital has a small orbital component and a longer posterior extension. This is excluded from the margin of the lower temporal fenestra by a contact between the jugal and the squamosal. The latter is slender and classically ‘hockey-stick shaped’ (CitationRobinson 1967), with a curved posteroventral tip that met the quadrate. Behind the squamosal in IVPP V15587 is a shorter, blade-like bone that we interpret as the supratemporal. The quadrate has a well-developed lateral conch and its tympanic crest is notched dorsally for a ventral squamosal peg. In IVPP V15587, the lateral conch is broken away revealing the medial pterygoid wing with a distinct pterygoid lappet. Ventrally the bone tapers to a narrow mandibular condyle. The epipterygoid is columnar.
No specimen has the skull preserved in palatal view, so nothing is known of this region except that the slender quadrate rami of the pterygoids contact the quadrates.
The braincase is not well preserved in any specimen. In IVPP V15586, the braincase lies behind the parietal and has partially disarticulated on the right side, with the opisthotic being slightly displaced. Rather short paroccipital processes meet the suspensorium bilaterally, and in IVPP V15011, the tips of elongate prootic alary processes extend forward to about half the length of the parietal body. The sphenoid (parasphenoid+basisphenoid), preserved in impression in IVPP V15587 () and in the juvenile IVPP V15508 (), is wide with divergent basipterygoid processes, and is roughly equal in length to the basioccipital. IVPP V15508 also preserves small, comma-shaped orbitosphenoids in situ (, Os). These delicate bones, supporting the sides of the cartilage-membrane braincase, are rarely preserved in fossils.
In the lower jaw, the dentary is short and strongly bifurcated posteriorly, accommodating a large surangular. The sutures between the surangular, angular and articular are not clear on the casts, but there is certainly a separate angular and the articular has a strong retroarticular process. The hyoid apparatus is represented by a slender element that is probably a first ceratobranchial and, in the region of the supratemporal (IVPP V15587, ), a second cartilaginous element that could be an epibranchial.
Axial skeleton
There were 26 presacral vertebrae of which the first seven or eight are cervicals, but it is not clear which rib is the first to attach to the sternum. The vertebrae are all procoelous, with well-developed rounded condyles that lack any trace of waisting at the contact with the centrum (contra varanoids) (). The neural spines are low, the zygapophyses are broad and rather strongly angled (especially in the dorsal region) and there are well-developed zygosphenoidal surfaces. All rib attachments are single. The small atlantal arches are typically disarticulated (e.g. ) and the atlantal intercentrum lies between the axis and the braincase. Cervical vertebrae are slightly shorter than the dorsals, with mid-ventral keels and keeled hypapophyses between at least the first four vertebrae in the series (e.g. IVPP V15586). The two sacral ribs are of roughly equal size and make contact distally. The second sacral ribs lack any trace of a posterior process or flange. The anterior five to nine caudal vertebrae bear long, laterally directed transverse processes, followed by another four with shorter processes. The first caudal is shorter than those following and its transverse processes are deeply grooved. In IVPP V14715 the caudal fracture plane occurs on or about caudal eight, but posterior caudals are poorly preserved. Two specimens (IVPP V14716, V15586) have truncated tails with traces of the unossified regrowth (, ). A haemal arch is visible on some tail vertebrae of IVPP V14715, lying in an intercentral, or posterior central, position.
The ribs are slender and single-headed. The first rib is on the fourth cervical vertebra as shown in IVPP V15586 (). This and the following two ribs are short and thick, with expanded ends for muscles attaching the pectoral girdle to the axial skeleton; rib four (cervical vertebra seven) is longer, more slender, and strongly curved, as are the next few in the series. All bear traces of the terminal costal cartilages but it is not clear which of these is the first to reach the sternum. The remaining presacral vertebrae all bear slender free ribs, although the last six are markedly shorter than those preceding them. Rib curvature decreases through the body. In the two larger specimens (IVPP V14716 and V14746), in particular, there are several pairs of long thoracoabdominal ribs that show little curvature and seem to span a body that was either unusually wide or rather flattened, suggesting an adult shape like that of the living iguanian Sauromalus.
Pectoral girdle and forelimb
The pectoral girdle is divided between the part and counterpart blocks in the holotype specimen, and the ventral elements are obscured by the vertebral column. A cruciform interclavicle is preserved in IVPP V15587A, with a short anterior process and a longer posterior stem (). Clavicles are not well preserved on any specimen, but the main block of V14715 has a right clavicle that is almost complete and has a hooked medial head (interpreted as a perforate clavicle without a complete posteromedial rim) and a short tapering dorsal process. The left clavicle lies adjacent to it, under the first right cervical rib, but only the hook-like medial part is visible (). In the young IVPP V15508, the clavicles are sigmoid and unexpanded medially. The scapulocoracoids are preserved on several specimens (). The component parts are fully co-ossified in IVPP V14715 and V15587, but not in IVPP V15586, indicating immaturity. The scapula part is relatively short with a small dorsal expansion but no scapular fenestra; the coracoid part is larger. Scapulocoracoid and anterior (primary) coracoid fenestrae are present, and there is a small posterior (secondary) coracoid notch. A shallow suprascapular cartilage is preserved above the scapula in IVPP V14715 (), its vertebral border showing clear traces of the notching seen in many living taxa. The cartilaginous sternum (presternum of CitationRussell & Bauer 2008) is preserved in IVPP V14715 and V15587 () and appears to have been a wide rhomboid, like that of many living taxa. The posterolateral margins angle towards the midline, with three direct attachment points for ribs and, on either side of the midline, attachment of a bifurcate mesosternal element (sensu CitationRussell & Bauer 2008) to which two further costal cartilages attach (). All of the cartilage traces are thin; none shows any evidence of mineralization. It is not possible to say whether or not sternal fontanelles were present.
The humeri are preserved as clear impressions in IVPP V14715 (). The joint surfaces are fully formed, with ossified and attached epiphyses. The proximal end of the bone is relatively narrow and lacks a strong deltopectoral crest. The shaft is long (humerus 79% of femoral length) and rather slender. Distally, the bone expands asymmetrically, the ectepicondyle remaining small (with an enclosed ectepicondylar foramen) and the entepicondyle flaring out into a large flexor muscle expansion. Both radial and ulnar condyles are well rounded. The radius and ulna of IVPP V14715 are in articulation, again with complete joint surfaces and a short wide ossified olecranon (ulna, ). Like the humerus, they are comparatively long (forearm roughly 85% of crural length). Distally the ulna is expanded into a hemispherical condyle for articulation with the carpus whereas the radius develops a distinct styloid process (). The carpus is preserved on several specimens but with different degrees of mineralization. It is well-preserved in the holotype (). There are nine elements, including a distal series of five bones, either distal carpals 1–5 (CitationFabrezi et al. 2007; CitationRussell & Bauer 2008) or the medial centrale and distal carpals 2–5 (CitationRomer 1956; CitationMaisano 2001). Three more proximal carpals are interpreted as ulnare, radiale and centrale (= lateral centrale of some authors, e.g. CitationRomer 1956), and there is a large pisiform. The size of this bone, in conjunction with the expansion of the entepicondylar region, suggests the flexor carpi ulnaris muscle may have been strong. A small element associated with the ulnare is probably an intermedium (IVPP V15587, ).
The manus is 63% of pedal length, with metacarpal (Mc) three significantly longer than Mc4, and a phalangeal formula of 2:3:4:5:3. Mc1 and Mc5 are somewhat more robust than Mc2–4. Although the third digit has one less phalanx than the fourth, it is of roughly the same overall length. This is partly due to the longer third metacarpal but also because the second, third and fourth phalanx of the fourth digit are short (). The fifth digit is also only slightly longer than digit one. Overall, therefore, although the hand is quite long, the digits have a rather symmetrical arrangement. The penultimate phalanges are longer than those preceding them, and the unguals are long and curved, with a pronounced flexor tubercle. Keratin claw sheaths are visible on both V14715 and V14716, and where fully exposed, they are sharp and at least twice as long as the underlying ungual phalanx ().
Figure 7 Liushusaurus acanthocaudata gen. et sp. nov., details of the postcranial skeleton. A, IVPP V14715B, humerus, radius and ulna, flexor surface; B, IVPP V14715A, partial humerus, radius and ulna, extensor surface; C, IVPP V15587A, manus, drawn from peel; D, IVPP V14715B, pectoral girdle, showing cartilaginous regions in grey; E, IVPP V14715A, detail of left clavicle, drawn from peel; F, IVPP V14716, pelvis with postcloacal bone in grey; G, IVPP V15587B, pectoral girdle, with cartilaginous sternum and sternal rib attachments in grey. All scale bars = 1 mm, except F = 5 mm. Abbreviations: c, centrale; ef, ectepicondylar foramen; en, entepicondyle; Fe, femur; H, humerus; i, intermedium; Il, ilium; Int, interclavicle; Is, ischium; me, mesosternum; ol, olecranon process; pc, possible postcloacal bone; pi, pisiform; Pu, pubis; R, radius; ra, radiale; rs, radial styloid process; Sa, sacrum; ScC, scapulocoracoid; SSc, suprascapula; Ste, sternum; U, ulna; ul, ulnare; V, vertebra; 1,5, digit numbers.
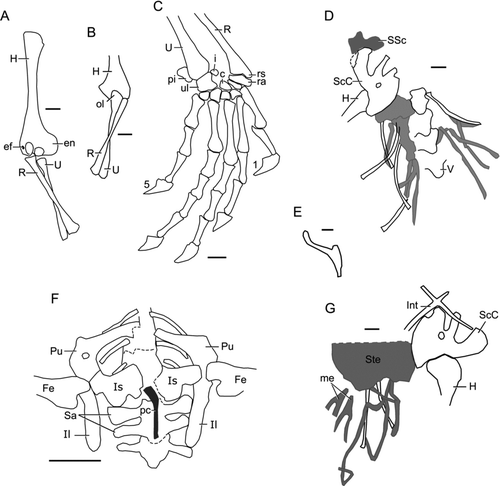
Figure 10 Liushusaurus acanthocaudata gen. et sp. nov., scalation. A, IVPP V14716, pes, fourth digit, drawn from peel, subdigital scales in grey; B, IVPP V14716, manus with outlines of subdigital scales and keratin claw sheathes, drawn from peel; C, IVPP V15507B, detail of femoral region showing size difference between small dorsal and large ventral scales. All scale bars = 1 mm. Abbreviations: ds, dorsal scales; Fe, femur; ks, keratin claw sheath; vs, ventral scales; 1, 5, digit numbers.
Pelvic girdle and hind limb
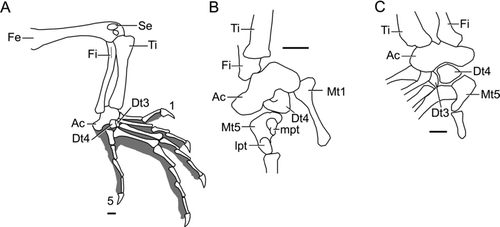
Like the scapulocoracoid, the pelvic elements can be separate, sutured or fused depending on the size/age of the individual. The ilium is long and slender, with an apparently horizontal dorsal edge, a short preacetabular process and a prepubic process. The pubis and ischium are best preserved on the counterpart block of IVPP V14715, on IVPP V14716 (, ), and IVPP V14746. They frame a large thyroid fenestra. The pubis is relatively elongate with a slender distal portion that is oriented anteromedially and ends with a narrow, moderately elongated but anteromedially directed symphysial process. The obturator foramen is proximal in position, as is the small, slightly incurved lateral pubic process. The ischia are flask-like, with a distinct posterior angle (posterior edges damaged in the specimen shown, ) and short symphysial process. Altogether, the puboischiadic plate is only half as long as it is wide. It lies anterior to the level of the first sacral rib due to the posterior position of the sacroiliac attachment, well behind the level of the acetabulum.
IVPP V14716 and V14746 both preserve a long, asymmetric, rod-like element, expanded anteriorly and tapering posteriorly (). This underlies the sacral region, but is quite distinct from the pubis and ischium and is most reasonably interpreted as a postcloacal or hemipenial bone, on the assumption that only one of the pair has been preserved in each case.
The mature femur is long, robust and sigmoid. At its distal end there may be one or more sesamoid bones (). The tibia and fibula are shorter, with the tibia the stouter of the two, although even in the large specimens (e.g. IVPP V. 14716, ) there is only a slight development of a cnemial crest. The distal end is not clearly preserved in any specimen but it appears to be notched in IVPP V14715 and V14746. The fibula meets the side of the lateral femoral condyle and bears a weak proximal crest in the largest specimens (e.g. IVPP V14746). Distally it has a convex epiphysis. Both elements meet the astragalocalcaneum in a relatively broad articulation. The three-dimensional structure of the tibio-talar joint is difficult to reconstruct, but it is at least incipiently sellar. The astragalus and calcaneum are fully co-ossified in the adult but, like the pectoral girdle and pelvis, progress from fully separated, to sutured, to fully co-ossified in individuals of different ontogenetic age. The adult bone is relatively short and wide with a distinct rounded calcaneal tuber (e.g. IVPP V14715, V14716, V14746, ). Distally it meets a large fourth distal tarsal (Dt4) in a complex articulation whereby Dt4 has a tongue-like process that fits under the edge of the astragalocalcaneum but the latter bone, in turn, seems to fit into a notch on the proximal surface of Dt4. There is also a smaller distal tarsal three. The metatarsals (Mt) increase in size from Mt1 to Mt4, although Mt4 is only slightly longer than Mt3. Mt5 is short and hooked, with equally pronounced plantar tubercles separated only by a narrow groove for the long flexor tendon. Proximally, Mt5 meets Dt4 but it is uncertain whether or not it contacted the astragalocalcaneum in life. The pedal phalangeal formula is usually 2:3:4:5:4 (2:3:4:5:3 in IVPP V15011). In IVPP V14715, the pedal digits are like those of the hand in being relatively slender; in IVPP V14716 they are more robust and therefore appear shorter. The unguals are long and curved, with a distinct flexor tubercle. Where preserved and exposed, the keratin claws are more than twice the length of the underlying ungual phalanges.
Figure 8 Liushusaurus acanthocaudata gen. et sp. nov., details of hind limb and foot. A, IVPP V14716 foot, drawn from peel, grey area shows extent of pedal soft tissues; B, IVPP V14715B, ankle region; C, IVPP V15587B, ankle region, drawn from peel,. All scale bars = 1 mm. Abbreviations: Ac, astragalocalcaneum; Dt3,4, distal tarsals; Fe, femur; Fi, fibula; lpt, lateral plantar tubercle; mpt, medial plantar tubercle; Mt1,5, metatarsals; Se, sesamoid; Ti, tibia; 1,5, digit numbers.
Soft tissue preservation and scalation
IVPP V14715, V14716, V15508, V15011, and V15587 show clear body outlines. Most specimens represent a rather stocky lizard with short sturdy legs, but IVPP V14716 is remarkably broad, both in the body and in the proximal parts of the legs and tail. Its body outline extends beyond the wide rib cage and the legs and tail appears thick. None of the body or tail impressions, either dorsally or ventrally, shows any trace of osteoderms.
IVPP V14715 and IVPP V15587 are notable in preserving much of the cartilaginous parts of the skeleton (as described in the sections above) and also for the exquisite preservation of the body scales, although scalation is preserved to different degrees in most specimens. Depending on the original position of interment, either the dorsal or the ventral body scales are better preserved. These dorsal and ventral scales differ in size and in shape. The dorsal scales are visible only on the edges of the trunk in IVPP V14715 but the ventral scales are also imprinted onto the counterpart block, obscuring the dorsal layer (). IVPP V14716 and V15508 () preserves this dorsal layer and shows that the dorsal scales are much smaller than those of the venter. On the venter, the scales are rhomboidal and overlapping. These continue onto the neck but the scales seem to become smaller on the head, at least on the side of the head that is exposed in the main part of IVPP V14715. Each of the ventral scales has a light base and a dark free edge (), but the amount of colour along the free edge varies from a thin strip to almost complete coverage. Whether this is representative of the original pigment pattern is uncertain. The darkest scales are on the middle of the belly, with the remaining scales grading in colour from medial to lateral, although the darkly pigmented edge always faces towards the midline. There are 43–48 scale rows (c. 4 per segment) between the front and hind limbs (allowing for the difficulty of locating the axillary position precisely), and roughly 28–30 scales across the body. The large rhomboid ventral scales continue onto the underside of the limbs, but on the dorsal surface the scales are again smaller and rounded, the diameter decreasing dorsally and distally. IVPP V15507 is the part and counterpart of an incomplete leg, attributed to the new species only on scalation pattern (, ). It is significant only in the exquisite preservation of the dorsal and ventral scalation, with a stark contrast between the large rhomboid ventral scales and the tiny granular dorsal scales (>0.5 mm diameter).
In the foot, the first four metatarsals are bound together in a single sheath of skin, but the fifth is divergent. On the plantar surfaces of the manus and pes, some of the scale impressions become much deeper and this is particularly marked on the undersides of the digits (). These deep depressions reflect the presence of wide tuberculous plantar scales, a common feature in modern lizards (CitationArnold et al. 2002). The penultimate phalanges bear a ventral median row of 5–6 of these wide scales (), each tubercle separated from the next by a thin band of skin. This structure is particularly clear in V14716.
On the ventral side of the tail (), the anterior scales are like those of the body but further distally, they elongate and taper, so that the tail seems to have been covered in a sheath of longitudinal spines. From the base of the tail to the level of the fifth caudal vertebrae, however, the lateral flanking scales and the dorsal scales are smaller and rounded, like those on the dorsum of the legs. Further distally the flanking scales take the form of the ventral scales and become spiky. The more dorsal scales are visible in IVPP V14716 and on IVPP V14746. They seem to be smaller and have a more annular arrangement, with a slight size alternation between rows (e.g. IVPP V14746).
The head scales are small with no development of the enlarged, plate-like scales seen in many scincomorph and anguimorph lizards. Unusually, IVPP V14715 preserves the ear region. The area immediately in front of the quadrate is covered by small scales; these resume behind the jaw, but there is an uncovered area immediately behind the quadrate that marks the position of the tympanic membrane.
Discussion
Phylogenetic position
Liushusaurus is a squamate, as demonstrated by the columnar epipterygoid, the streptostylic quadrate, reduced squamosal, procoelous vertebrae and fenestrate scapulocoracoid. As outlined in the diagnosis, it can be differentiated from other valid Yixian and Yixian equivalent lizards, and from well-preserved lizard specimens from other contemporaneous, or roughly contemporaneous, localities worldwide.
The retention of a relatively large postfrontal argues against iguanian affinity, as does the absence of a distinct mid-dorsal scale row (CitationEstes et al. 1988). A bifurcated postfrontal that clasps the fronto-parietal suture has been listed as a scleroglossan synapomorphy (CitationEstes et al. 1988, but see CitationTownsend et al. 2004; CitationVidal & Hedges 2005), as has the presence of perforate clavicles (although this is present in some iguanians, e.g. Gambelia wislizeni, Basiliscus, CitationRussell & Bauer 2008), of a cruciform interclavicle with a long anterior process, and of a notched (sellar) distal tibial epiphysis (CitationEstes et al. 1988). Alternation in the size of tail scales has been cited as an autarchoglossan trait (CitationArnold et al. 2002), as has a marked difference in dorsal and ventral scale patterns (CitationCamp 1923), although gekkotan skin patterns might be secondary (and the iguanian Sceloporus comes close to scleroglossan scalation, SEE pers. obs.).
The absence of body osteoderms precludes attribution to Scincidae, Cordylidae or Anguidae, or to the extinct Paramacellodidae. The absence of plate-like head scales differentiates Liushusaurus from Lacertidae, Gymnophthalmidae, Teiidae, Xantusiidae, Cordyliformes, or Scincidae, and many anguimorph lineages. Small regular head scales are found in iguanians, gekkotans and some anguimorphs (xenosaurs, shinisaurs or platynotans) and presumably represent the primitive squamate condition. The simple vertebral condyles preclude a varanoid relationship, whereas among recent genera, postcloacal bones are found in some gekkotans (CitationKluge 1982) and xantusiids (CitationRieppel 1976), and occasionally in Varanus (CitationSmith 1935). Liushusaurus resembles lacertiforms (sensu CitationEstes et al. 1988) generally in having a pterygoid lappet on the quadrate, a retroarticular process with a dorsal pit, and well-developed vertebral zygosphenes (CitationEstes et al. 1988), and teiioids more closely in the absence of the parietal foramen, the shape of the parietal (long posterior processes, no lateral parietal flanges, no extended squamosal/parietal contact: CitationConrad 2008; CitationEvans 2008), the absence of strong dermal rugosities or osteoderms, and the presence of a posterior coracoid emargination. However, teiioids lack an ectepicondylar foramen on the humerus and have fused frontals, although these are paired in the related boreoteiioids (sensu CitationNydam et al. 2007).
Figure 11 Tree showing hypothesis of relationship for Liushusaurus acanthocaudata gen. et sp. nov. within Squamata, based on a simplified strict consensus of three equally parsimonious trees (L = 3687) from a heuristic search using TNT (CitationGoloboff et al. 2008) and the full data matrix of CitationConrad (2008).
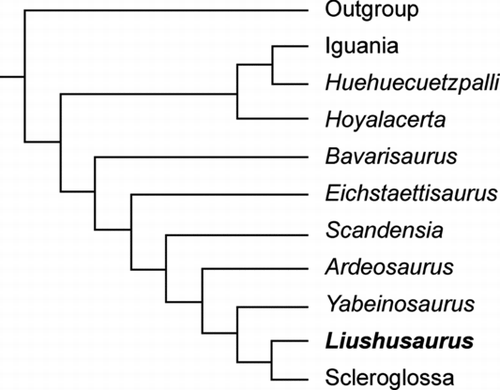
Table 2 Tibia length (mm) in eight available specimens of Liushusaurus acanthocaudata gen. et sp. nov. (in mm, all from IVPP collection).
This mosaic of scale and skeletal characters complicates systematic assignment, but is suggestive of a closer relationship to Scleroglossa than to Iguania. This was tested by phylogenetic analysis, using the data set of CitationConrad (2008) with 131 living and extinct genera representative of all major squamate clades (having limited all genera to single representatives, and limited mosasauroids to a representative sample). The matrix was run first on PAUP 4.0b10 (CitationSwofford 2002), completely unordered and then using the ordering of CitationConrad (2008), multistate taxa were treated as uncertainty, and Kuehneosauridae, Marmoretta and Rhynchocephalia were the designated outgroups. A preliminary heuristic analysis on unordered characters yielded 6 equally parsimonious trees (Tree length [L] = 2536; Consistency Index [CI] = 0.331; Rescaled Consistency Index [RC] = 0.188), the Strict Consensus of which placed Liushusaurus on the stem of Scleroglossa. We then re-ran the heuristic analysis using the ordering of CitationConrad (2008) (20 characters ordered) obtaining 3055 equally parsimonious trees (L = 2580; CI = 0.193; RC = 0.126) in which Liushusaurus was again placed as a stem scleroglossan, within the wider clade that CitationConrad (2008) refers to as Scincogekkonomorpha (effectively: Scleroglossa + those taxa closer to Scleroglossa than to Iguania). Allowing one additional step (L = 2581) collapsed the topology at the base of the tree, leaving Liushusaurus with an unresolved scincogekkonomorph placement. The matrix is too large for a full Branch and Bound analysis in PAUP, but we ran Liushusaurus in a smaller analysis (18 taxa) against representatives of living lizard clades; it again emerged in an unresolved scincogekkonomorph position. We then ran the analysis using the full Conrad data set in the programme TNT (CitationGoloboff et al. 2008), using Conrad's settings. It yielded three trees (L = 3687) which placed Liushusaurus as the sister group of Scleroglossa ().
Whether this placement as a stem scleroglossan truly reflects the phylogenetic position of Liushusaurus, or is indicative of our limited ability to differentiate between early members of major squamate clades using morphological characters, is uncertain. Placement of other Jurassic–Early Cretaceous genera like Parviraptor, Eichstaettisaurus, Ardeosaurus and Scandensia has been equally problematic.
Lifestyle
The body outline, particularly in IVPP V14716, suggests that the adult Liushusaurus was either a rather flattened lizard (like the Malagasy gecko Uroplatus or the African cordylid Platysaurus), or was broad-bodied like the North American iguanine Sauromalus. The relatively short scapula and short suprascapular cartilage are consistent with a depressed body shape, as are the long, thin, weakly curved posterior ribs. Body flattening in lizards can serve to maximise surface area exposed to the sun (e.g. the cordylid Platysaurus), aid crevice dwelling (e.g. Platysaurus), help to disguise the body outline (e.g. the Malagasy gekkotan Uroplatus), and/or permit a semi-controlled fall (e.g. the ‘Butterfly lizard’, Leiolepis) (CitationLosos et al. 1989). The long, slender tapering claws of Liushusaurus would be consistent with a climber on rough surfaces (rock or vegetation) (CitationZani 2000), as are the relatively long forelimbs, absence of marked asymmetry in the manus and pes, and well-developed tubercular plantar digital scales (CitationArnold et al. 2002). The simple conical teeth suggest a diet of insects or other small invertebrates.
Sexual dimorphism
Two specimens (IVPP V14716, IVPP V14746) bear a ventral pelvic element that we interpret as a possible post-cloacal bone, a sexually dimorphic (male) feature found in some living lizards (notably gekkotans). They are the 1st and 3rd largest specimens in the collection (the 2nd largest specimen, V15507, is preserved in dorsal view so the presence or absence of post-cloacal bones cannot be determined) (), hinting at size dimorphism as in many living taxa.
Ontogenetic variation
As for Yabeinosaurus (CitationEvans et al. 2005) and Dalinghosaurus (CitationEvans et al. 2007), Liushusaurus shows age related variation (e.g. in the co-ossification of elements, the shape of the parietal, the frontoparietal suture, the shape of the clavicle). This variation highlights the problems that arise when juvenile skeletons are used as holotypes, especially if they lack diagnostic features. The morphology and phylogenetic position of Yabeinosaurus (e.g. CitationEstes 1983) was misunderstood for several decades because the holotype chosen was a juvenile, a problem compounded by the selection of an equally juvenile specimen as a neotype (CitationJi et al. 2001).
Among the eight available specimens of Liushusaurus, only the tibia/fibula length can be measured in all specimens. It ranges from 3.62 to 11.85 mm and thus shows a considerable size variation (). As IVPP 15587 is the largest specimen that has a skull and it provides diagnostic skull information, we selected it as the holotype to minimize possible bias due to ontogenetic variation. As shown in , Liushusaurus has proportionally shorter forelimbs and longer hind limbs in small individuals, and proportionally shorter feet but longer femora and tibiae in larger ones.
Conclusions
The deposits of the Yixian Formation of northeastern China continue to produce exquisite fossils of a wide range of groups. Squamates are well represented in this assemblage, with at least six diagnosable genera including the new Liushusaurus. Of these, two (Mimobecklesisaurus, Yabeinosaurus) seem to be surviving representatives of essentially Jurassic lineages and may therefore be relicts of a once wider Laurasian or Eurasian distribution. The phylogenetic analysis suggests Liushusaurus may fall into the same category. Dalinghosaurus on the other hand (as a relative of the shinisaurs, CitationEvans & Wang 2005; CitationConrad 2008) and the contemporaneous Japanese Kuwajimalla (as an early relative of the boreoteiioids, CitationEvans & Manabe 2008) probably represent squamate lineages that evolved within Asia. The same may be true of Pachygenys and the gliding Xianglong, but additional material of each is needed to determine their relationships.
Appendix
Data matrix for Liushusaurus
The principle phylogenetic analysis described in the text used the data matrix of CitationConrad (2008). Listed below are the 364 character codings for Liushusaurus. A = states 0 & 1; B = 0 & 2; C = 1 & 2.
Acknowledgements
This manuscript derives from work completed as part of a joint Anglo-Chinese project funded by the Royal Society, London, and the National Natural Science Foundation (NNSF) of China. Field work and fossil collection were supported by grants from NNSF (40121202) and Ministry of Science and Technology (2006CB806405) of China. We are grateful to Dr Xiaolin Wang (IVPP, Beijing) for additional information about the Jehol Biota and the geology of the Yixian Formation; and to two anonymous reviewers for their comments on an earlier draft. We acknowledge the sponsorship of the Willi Hennig Society in making the programme TNT (CitationGoloboff et al. 2008) available for use, and Dr Paul Upchurch (UCL) for his help in running the TNT analysis.
Related Research Data
References
- Alifanov , V. R. 2000 . Macrocephalosaurs and the early evolution of lizards of Central Asia . Trudi Paleontologicheskoyo Instituta , 272 : 1 – 126 . [In Russian]
- Arnold , E. N. , Azar , D. , Ineich , I. and Nel , A. 2002 . The oldest reptile in amber; a 120 million year old lizard from Lebanon . Journal of Zoology , 258 : 7 – 10 .
- Camp , C. L. 1923 . Classification of the lizards . Bulletin of the American Museum of Natural History , 48 : 289 – 481 .
- Chang , M. M. , Zhang , J. Y. and Miao , D. S . 2006 . A lamprey from the Cretaceous Jehol biota of China . Nature , 441 : 972 – 974 .
- Conrad , J. 2008 . Phylogeny and systematics of Squamata (Reptilia) based on morphology . Bulletin of the American Museum of Natural History , 310 : 1 – 182 .
- Endo , R. and Shikama , T. 1942 . Mesozoic reptilian fauna in the Jehol mountainland, Manchoukuo . Bulletin of the Central National Museum of Manchoukuo , 3 : 1 – 19 .
- Estes , R. 1983 . “ Sauria Terrestria, Amphisbaenia ” . In Handbuch der Paläoherpetologie, 10A , Edited by: Wellnhofer , P. 249 Stuttgart : Gustav Fischer Verlag .
- Estes , R. , de Queiroz , K. and Gauthier , J. A. 1988 . “ Phylogenetic relationships within Squamata ” . In Phylogenetic relationships of the lizard families , Edited by: Estes , R. and Pregill , G. 119 – 281 . Stanford : Stanford University Press .
- Evans , S. E. 1994a . The Solnhofen (Jurassic: Tithonian) lizard genus Bavarisaurus: new skull material and a reinterpretation . Neues Jahrbuch fur Paläontologie und Geologie, Abhandlungen , 192 : 37 – 52 .
- Evans , S. E. 1994b . A new anguimorph lizard from the Jurassic and Lower Cretaceous of England . Palaeontology , 37 : 33 – 49 .
- Evans , S. E. 2008 . “ The skull of lizards and Tuatara ” . In Biology of the Reptilia, Vol. 20. The skull of Lepidosauria , Edited by: Gans , C. , Gaunt , A. S. and Adler , K. 1 – 347 . New York : Ithaca . Society for the Study of Reptiles & Amphibians, Contributions to Herpetology, 23
- Evans , S. E. and Barbadillo , L. J. 1997 . Early Cretaceous lizards from Las Hoyas, Spain . Zoological Journal of the Linnean Society , 119 : 23 – 49 .
- Evans , S. E. and Barbadillo , L. J. 1998 . An unusual lizard (Reptilia, Squamata) from the Early Cretaceous of Las Hoyas, Spain . Zoological Journal of the Linnean Society , 124 : 235 – 266 .
- Evans , S. E. and Barbadillo , L. J. 1999 . A short-limbed lizard from the Early Cretaceous of Spain . Special Papers in Palaeontology , 60 : 73 – 85 .
- Evans , S. E. and Chure , D. 1998 . Paramacellodid lizard skulls from the Jurassic Morrison Formation at Dinosaur National Monument, Utah . Journal of Vertebrate Paleontology , 18 : 99 – 114 .
- Evans , S. E. , Lacasa-Ruiz , A. and Erill Rey , J. 1999 . A lizard from the Early Cretaceous (Berriasian–Hauterivian) of Montsec . Neues Jahrbuch fur Paläontologie und Geologie, Abhandlungen , 215 : 1 – 15 .
- Evans , S. E. and Manabe , M. 2008 . A herbivorous lizard from the Early Cretaceous of Japan . Palaeontology , 51 : 487 – 498 .
- Evans , S. E. and Manabe , M. 2009 . The Early Cretaceous lizards of Eastern Asia: new material of Sakurasaurus from Japan . Special Papers in Palaeontology , 81 : 1 – 17 .
- Evans , S. E. , Raia , P. and Barbera , C. 2004 . New lizards and sphenodontians from the Early Cretaceous of Italy . Acta Palaeontologica Polonica , 49 : 393 – 408 .
- Evans , S. E. , Raia , P. and Barbera , C. 2006 . A revision of the Early Cretaceous lizard Chometokadmon from Italy . Cretaceous Research , 27 : 673 – 683 .
- Evans , S. E. and Searle , B. 2002 . Lepidosaurian reptiles from the Purbeck Limestone Group of Dorset, southern England . Special Papers in Palaeontology , 68 : 145 – 159 .
- Evans , S. E. and Wang , Y. 2005 . Dalinghosaurus, a lizard from the Early Cretaceous Jehol Biota of Northeast China . Acta Palaeontologica Polonica , 50 : 725 – 742 .
- Evans , S. E. and Wang , Y. 2007 . A juvenile lizard specimen with well-preserved skin impressions from the Upper Jurassic/Lower Cretaceous of Daohugou, Inner Mongolia, China . Naturwissenschaften , 94 : 431 – 439 .
- Evans , S. E. and Wang , Y. 2009 . A long-limbed lizard from the Upper Jurassic/Lower Cretaceous of Daohugou, Ningcheng, Nei Mongol, China . Vertebrata PalAsiatica , 47 : 21 – 34 .
- Evans , S. E. , Wang , Y. and Jones , M. E. H. 2007 . An aggregation of lizard skeletons from the Lower Cretaceous of China . Senckenbergiana Lethaea , 87 : 109 – 118 .
- Evans , S. E. , Wang , Y. and Li , C. 2005 . The Early Cretaceous lizard Yabeinosaurus from China: resolving an enigma . Journal of Systematic Palaeontology , 3 : 319 – 335 .
- Fabrezi , M. , Abdala , V. and Martinez Oliver , M. I. 2007 . Developmental basis of limb homology in lizards . The Anatomical Record , 290 : 900 – 912 .
- Gao , K. Q. and Cheng , Z. W. 1999 . A new lizard from the Lower Cretaceous of Shandong, China . Journal of Vertebrate Paleontology , 19 : 456 – 465 .
- Goloboff , P. , Farris , J. and Nixon , K. 2008 . TNT: a free program for phylogenetic analysis . Cladistics , 24 : 774 – 786 .
- Hoffstetter , R. 1964 . Les Sauria du Jurassique supérieur et specialement les Gekkota de Bavière et de Mandchourie . Senckenbergianna biologica , 45 : 281 – 324 .
- Hoffstetter , R. 1967 . Coup d’oeil sur les Sauriens (Lacertiliens) des couches de Purbeck (Jurassique supérieur d’Angleterre) . Problemes Actuels de Paléontologie (Evolution des Vertebrés) Centre National de la Recherche Scientifique , 163 : 349 – 371 .
- Ji , S. A. 1998 . “ A new long-tailed lizard from the Upper Jurassic of Liaoning, China ” . In Collected Works of International Symposium on Geological Science, Peking University, Beijing, China. , 496 – 505 . Beijing, , China : Seismological Press . Department of Geology, Peking University (ed.)
- Ji , S. A. 2005 . A new Early Cretaceous lizard with well-preserved scale impressions from western Liaoning, China . Progress in Natural Science , 15 : 162 – 168 .
- Ji , S. A. and Ji , Q. 2004 . Postcranial anatomy of the Mesozoic Dalinghosaurus (Squamata): evidence from a new specimen of Western Liaoning . Acta Geologica Sinica , 78 : 897 – 906 .
- Ji , S. A. and Ren , D. 1999 . First record of lizard skin fossil from China with description of a new genus (Lacertilia, Scincomorpha) . Acta Zootaxonomica Sinica , 24 : 114 – 120 .
- Ji , S. A. , Lu , L. W. and Bo , H. C. 2001 . New material of Yabeinosaurus tenuis (Lacertilia) . Land and Resources , 2001 : 41 – 43 . [In Chinese]
- Kluge , A. G. 1982 . Cloacal bones and sacs as evidence of gekkonoid lizard relationships . Herpetologica , 38 : 348 – 355 .
- Li , J. L. 1985 . A new lizard from the Late Jurassic of Subei, Gansu . Vertebrata PalAsiatica , 23 : 13 – 18 . [In Chinese with English summary]
- Li , P. P. , Gao , K. Q. , Hou , L. H. and Xu , X. 2007 . A gliding lizard from the Early Cretaceous of China . Proceedings of the National Academy of Sciences , 104 : 5507 – 5509 .
- Losos , J. B. , Papenfuss , T. J. and Macey , J. R. 1989 . Correlates of sprinting, jumping and parachuting performance in the butterfly lizard, Leiolepis belliani . Journal of Zoology , 217 : 559 – 568 .
- Lü , J. C. 2009 . A baby pterodactyloid pterosaur from the Yixian Formation of Ningcheng, Inner Mongolia, China . Acta Geologica Sinica , 83 : 1 – 8 .
- Maisano , J. A. 2001 . A survey of the state of ossification in neonatal squamates . Herpetological Monographs , 15 : 135 – 157 .
- Mateer , N. 1982 . Osteology of the Jurassic lizard Ardeosaurus brevipes (Meyer) . Palaeontology , 25 : 461 – 469 .
- Nydam , R. L. and Cifelli , R. L. 2002a . A new teiid lizard from the Cedar Mountain Formation (Albian-Cenomanian boundary) of Utah . Journal of Vertebrate Paleontology , 22 : 276 – 285 .
- Nydam , R. L. and Cifelli , R. L. 2002b . Lizards from the Lower Cretaceous (Aptian–Albian) Antlers and Cloverly Formations . Journal of Vertebrate Paleontology , 22 : 286 – 298 .
- Nydam , R. L. , Eaton , J. G. and Sankey , J. 2007 . New taxa of transversely-toothed lizards (Squamata; Scincomorpha) and new information on the evolutionary history of ‘teiids’ . Journal of Paleontology , 81 : 538 – 549 .
- Oppel , M. 1811 . Die Ordnungen, Familien und Gattungen der Reptilien, als Prodrom einer Naturgeschichte derselben , 87 Munich : Joseph Lindauer .
- Reynoso , V. H. 1998 . Huehuecuetzpalli mixtecus gen. et sp. nov: a basal squamate (Reptilia) from the Early Cretaceous of Tepexi de Rodríguez, Central México . Philosophical Transactions of the Royal Society B , 353 : 477 – 500 .
- Rieppel , O. 1976 . On the presence and function of post-cloacal bones in the Lacertilia . Monitore Zoologico Italiano , 10 : 7 – 13 .
- Robinson , P. L. 1967 . The evolution of the Lacertilia . Problems Actuels de Paleontologie, Colloques Internationaux du Centre national de la Recherche Scientifique, Paris , 163 : 395 – 407 .
- Romer , A. S. 1956 . Osteology of the Reptiles , 772 Chicago : University of Chicago Press .
- Russell , A. P. and Bauer , A. M. 2008 . “ The appendicular locomotor apparatus of Sphenodon and normal-limbed squamates ” . In Biology of the Reptilia, Vol. 21. The skull and appendicular locomotor apparatus of Lepidosauria , Edited by: Gans , C. , Gaunt , A. S. and Adler , K. 1 – 465 . New York : Ithaca . Society for the Study of Reptiles & Amphibians, Contributions to Herpetology 24
- Seiffert , J. 1973 . Upper Jurassic lizards from Central Portugal . Memoria, Serviços Geológicos de Portugal , 22 : 1 – 85 .
- Smith , M. A. 1935 . The Fauna of British India. Vol. 2, Sauria , 441 London : Taylor and Francis .
- Swofford , D. L. 2002 . PAUP* (Version 4.06b10 for 32-bit Microsoft Windows) , Sunderland, Massachusetts, , USA : Sinauer Associates Incorporated, Publishers .
- Townsend , T. M. , Larson , A. , Louis , E. and Macey , J. R. 2004 . Molecular phylogenetics of Squamata: the position of snakes, amphisbaenians, and dibamids, and the root of the squamate tree . Systematic Biology , 53 : 735 – 757 .
- Vidal , N. and Hedges , S. B. 2005 . The phylogeny of squamate reptiles (lizards, snakes, and amphisbaenians) inferred from nine nuclear protein-coding genes . Comptes Rendus Biologies , 328 : 1000 – 1008 .
- Wang , X. L. , Wang , Y. Q. , Zhang , F. C. , Zhang , J. Y. , Zhou , Z. H. , Jin , F. , Hu , Y. M. , Gu , G. and Zhang , H. C. 2000 . Vertebrate biostratigraphy of the Lower Cretaceous Yixian Formation in Lingyuan, western Liaoning and its neighbouring southern Nei Mongol (Inner Mongolia), China . Vertebrata PalAsiatica , 38 : 81 – 99 . [In Chinese with English summary]
- Wang , Y. 2004 . A new Mesozoic caudate (Liaoxitriton daohugouensis sp. nov.) from Inner Mongolia, China . Chinese Science Bulletin , 49 : 858 – 860 .
- Wang , Y. and Evans , S. E. 2006a . A new short-bodied salamander from the Upper Jurassic/Lower Cretaceous of China . Acta Palaeontologica Polonica , 51 : 127 – 130 .
- Wang , Y. and Evans , S. E. 2006b . Advances in the study of fossil amphibians and squamates from China: The past fifteen years . Vertebrata PalAsiatica , 44 : 60 – 73 .
- Wang , Y. and Li , J. 2008 . “ Order Squamata ” . In The Chinese fossil reptiles and their kin , Edited by: Li , J. , Wu , X. and Zhang , F. 115 – 137 . Beijing : Science Press . 2nd edition
- Wang , Y. and Rose , C. 2005 . Jeholotriton paradoxus (Amphibia: Caudata) from the Lower Cretaceous of southeastern Inner Mongolia, China . Journal of Vertebrate Paleontology , 25 : 523 – 532 .
- Zani , P. A. 2000 . The comparative evolution of lizard claw and toe morphology and clinging performance . Journal of Evolutionary Biology , 13 : 316 – 325 .
- Zhang , B. L. , Ren , D. and Pang , H. 2008 . New dragonflies (Insecta: Odonata: Gomphaeschnidae) from the Yixian Formation in Inner Mongolia, China . Progress in Natural Science , 18 : 59 – 64 .
- Zhang , X. W. , Ren , D. , Pang , H. and Shih , C. K. 2008 . A new genus and species of Chresmodidae (Insecta: Gryllones) from Upper Jurassic–Lower Cretaceous of Yixian Formation, Inner Mongolia, China . Zootaxa , 1702 : 26 – 40 .