Abstract
Magnosaurus nethercombensis from the Lower Bajocian (Middle Jurassic) of Dorset, UK is a valid species, possessing a single autapomorphy: the presence of anteroposteriorly elongate foramina, inclined anterodorsally and located ventrally on the lateral surface of the dentary. It is the oldest known definite tetanuran dinosaur and shows two tetanuran features: a reduced ischial peduncle of the ilium and the presence of a marked femoral extensor groove. Other records of putative early tetanurans are reviewed: ‘Zanclodon cambrensis’ from the Rhaetian of Wales; remains from the Norian–Hettangian of Switzerland; Shuvosaurus and Protoavis from the Norian of Texas; Eshanosaurus from the Hettangian of China; theropod remains originally included in the syntype series of Scelidosaurus from the Hettangian–Sinemurian of England; a fragmentary skeleton from the Sinemurian of Italy; Cryolophosaurus from the Sinemurian–Pliensbachian of Antarctica; the partial skeleton of a small theropod from the Toarcian of Morocco; and the lost syntype material of ‘Streptospondylus cuvieri’ from the Toarcian of Whitby. None of these records can be confidently considered to be the earliest tetanuran record. An early Middle Jurassic age for the earliest-known tetanuran is more consistent with a restricted content of Ceratosauria, comprising Ceratosaurus, Elaphrosaurus, and abelisauroids, than with a wider content including coelophysoids, due to reduction in the length of phylogenetic ghost lineages.
Introduction
Tetanurans are a group of theropod dinosaurs that includes birds and a wide variety of ecological types represented by extinct taxa. Tetanurans are primarily carnivores, but include herbivores such as the therizinosaurs, toothless ‘ostrich mimic’ ornithomimids and bizarre, short-faced, crested oviraptorosaurs. From their origins in the Middle Jurassic or late Early Jurassic, tetanurans were the most abundant and conspicuous terrestrial predators of the Mesozoic. The oldest definitive tetanuran is Magnosaurus nethercombensis from the Lower Bajocian of Dorset, England. The holotype specimen of Magnosaurus nethercombensis (OUMNH J.12143) was briefly described and partly figured by von CitationHuene (1923; 1926a, Figs 41–43; 1926b, –) as a new species of Megalosaurus. It was subsequently recognised as a distinct genus (von CitationHuene 1932). Although CitationWaldman (1974, pl. 44) briefly redescribed the material as Megalosaurus nethercombensis, subsequent reviews have upheld the validity of Magnosaurus (e.g. CitationMolnar et al. 1990; CitationRauhut 2003; CitationHoltz et al. 2004). Unfortunately, many important anatomical features, particularly of the pelvis and hindlimbs, were overlooked by von CitationHuene (1926a, b) and CitationWaldman (1974). Consequently, details of several systematically informative features are not described in the published literature and few attempts have been made to establish the affinities of the taxon, although CitationRauhut (2003) considered the megalosaurid Eustreptospondylus as a junior synonym of Magnosaurus. In order to understand the diversification of Tetanurae, the anatomy of the earliest representatives of the clade must be well understood. The description of Magnosaurus given herein is an important step in an ongoing review of the British Middle Jurassic theropod fossil record (CitationDay & Barrett 2004; CitationBenson 2008a, Citationb, Citationc, Citation2009, Citation2010; CitationBenson et al. 2008; CitationSadleir et al. 2008).
The present contribution aims to provide a compre- hensive description and comparisons of the holotype and only specimen of Magnosaurus (OUMNH J.12143) that will be of enduring use to theropod systematics and phylogenetics. It also establishes the taxonomic validity of Magnosaurus nethercombensis and highlights differences between this taxon and closely-related, anatomically similar taxa such as Dubreuillosaurus and Eustreptospondylus. Finally, although it is considered here that Magnosaurusis the oldest tetanuran currently known, other authors have suggested that older records are known (e.g. CitationTaquet 1985; CitationCarrano & Sampson 2004). These records are reviewed. To understand the evolutionary dynamics of tetanuran origins and diversification it is vital to determine the age of their oldest currently-known representative. This datum also bears on the date of origin of Ceratosauria, the sister group of Tetanurae. This date has been pivotal in recent discussions on the likely taxonomic content of Ceratosauria, as morphological phylogenetic studies have yet to arrive at a consensus on this issue (CitationRauhut 2003; Allain et al. 2007; Carrano & Sampson 2008).
Institutional Abbreviations
BMNH: Natural History Museum, London, UK; GSM: Geological Survey Museum, Keyworth, UK; MACN: Museo Argentino de Ciencas Naturalles, Buenos Aires, Argentina; MNHN: Muséum National d’Histoire Naturelle, Paris, France; OUMNH: Oxford University Museum of Natural History, Oxford, UK; PVL: Instituto de Miguel Lillo, Tucumán, Argentina.
Systematic palaeontology
Dinosauria Owen, 1842
Theropoda CitationMarsh, 1881
Tetanurae CitationGauthier, 1986
Megalosauroidea CitationHuxley, 1869
Megalosauridae CitationHuxley, 1869
Genus Magnosaurus von CitationHuene, 1932
Type and only species
Magnosaurus nethercombensis (von CitationHuene, 1923).
Magnosaurus nethercombensis (von CitationHuene, 1923)
(–)
1923 ‘Megalosaurus’ nethercombensis von Huene: 450.
1926a Megalosaurus (subgen. b) nethercombensis von Huene; von Huene: 72, Figs 41–43.
1926b Megalosaurus nethercombensis von Huene; von Huene: 476, –.
1932 Magnosaurus nethercombensis (von Huene); von Huene: 218, 220 (new combination).
Holotype
OUMNH J.12143, a fragmentary skeleton comprising: right (OUMNH J.12143/1b) and left (OUMNH J.12143/1a) dentaries; a dorsal vertebra (OUMNH J.12143/9b; formerly OUMNH J.12143/5; OUMNH unpublished collection records) and its counterpart (OUMNH J.12143/9a); a caudal vertebra (OUMNH J.12143/8); impressions of ribs (OUMNH J.12143/13-14); the ischial peduncle of the right ilium (OUMNH J.12143/10); the proximal end (OUMNH J.12143.12) and shaft (OUMNH J.12143/3) of the right pubis; the proximal end of the left pubis (OUMNH J.12143/11); right femur (OUMNH J.12143/6); left femur (OUMNH J.12143/7); left tibia (OUMNH J.12143/2), right tibia (OUMNH J.12143/3-4); and indeterminate fragments (OUMNH J.12143/15-16, OUMNH uncatalogued).
Previous diagnosis
‘A small megalosaurid. Dentary with curved dorsal and ventral margins. Twelve or thirteen teeth, pillar-like in lateral view, recurved occlusally, both carinae serrated. Slender tibia, transverse diameter of head two-thirds of longitudinal diameter; cnemial crest projects anteriorly, small crista lateralis, astragalus with well-developed ascending process. Pubis rod-like, proximal median lamella’ (CitationWaldman 1974, p. 333; based on von CitationHuene 1926b and CitationDelair 1959).
Emended diagnosis
Small-bodied megalosaurid theropod that differs from all other geographically proximate, Middle Jurassic theropods (; and see below). Magnosaurus nethercombensis possesses a single potential autapomorphy: the presence of anteroposteriorly elongate foramina located ventrally on the lateral surface of the dentary. These foramina are inclined anterodorsally and continue anterodorsally as long grooves (). This feature is only interpreted as an autapomorphy hesitantly as the level of variation exhibited by nutrient foramina in theropod dentaries is not understood and may be relatively high. These foramina are only known from the right dentary as the ventral part of the lateral surface of the right dentary is damaged.
Figure 1 Megalosaurid dentaries. A-C, right dentary of Magnosaurus nethercombensis OUMNH J.12143/1b in medial (A, B) and dorsal (C) views; D, E, right dentary of Megalosaurus bucklandii OUMNH J.13505 in dorsal (D) and medial (E) views; F, right dentary of Duriavenator hesperis BMNH R332 in medial view; G, right dentary of Eustreptospondylus oxoniensis OUMNH J.13556 in medial view; H, left dentary of Dubreuillosaurus valesdunensis in medial view (reversed). In line drawing (B) light grey tone indicates tooth and dark grey tone indicates broken bone. Abbreviations: 3rd, third dentary alveolus; idp, interdental plate; mef, Meckelian fossa; meg, Meckelian groove; mfr, Meckelian foramen. Scale bars equal 100 mm. Parts of image are modified from CitationBenson et al. (2008) (D, E) and CitationBenson (2008a) (F).
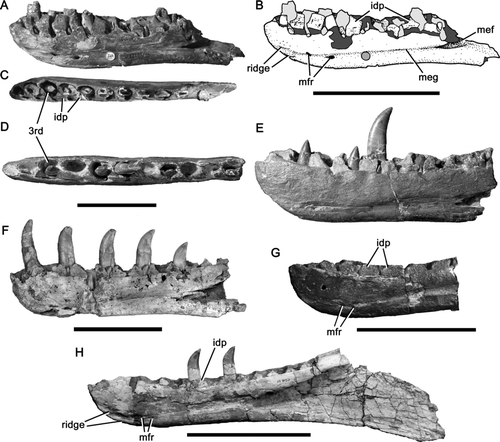
Figure 2 Dentaries of Magnosaurus nethercombensis in lateral view. A, B, right dentary OUMNH J.12143/1b with magnified (x3) portions showing elongate foramina (A); C, left dentary OUMNH J.12143/1a. Abbreviations: for, foramen. Scale bar equals 100 mm.

Occurrence
Inferior Oolite Formation of Nethercombe, Dorset, UK. According to CitationWaldman (1974) the material was collected from the Stephanoceras humphriesianum ammonite Zone and Subzone (Early Bajocian; CitationCallomon 2003; CitationGradstein et al. 2004).
Remarks
A partial theropod skeleton from Nethercombe in the collection of James Parker in Oxford was reported and referred to the new species Megalosaurus nethercombensis by von CitationHuene (1923). He distinguished it from Megalosaurus bucklandii on the basis of the teeth being thicker in their mesial half and the pubis being rod-like distally. He later made this the type species of the new genus Magnosaurus von CitationHuene, 1932. However, neither of the differences noted by von CitationHuene (1923) are supported by the present study: the pubis of Magnosaurus is only rod-like because its distal end, which may have been expanded and similar to that of Megalosaurus, is not preserved; the most complete adult tooth of Magnosaurus (OUMNH J.121431a; ) is not thick mesially and it is likely that von Huene's (1923) perception of the tooth profile was influenced by the fact that most of the teeth preserve only the base, which is thicker mesially than is the crown. Nonetheless, M. nethercombensis is clearly distinct from Megalosaurus on the basis of features of the lectotype dentary of M. bucklandii listed in the emended diagnosis and discussion here, and recent authors have not followed Waldman's (1974) suggestion that M. nethercombensis falls within the genus Megalosaurus (e.g. CitationRauhut 2003; CitationHoltz et al. 2004).
The majority of the features cited in Waldman's (1974, p. 333) diagnosis are general descriptions of features that are present in the majority of theropods. Only the shape of the dentary and the number of teeth have a particularly restricted distribution. The dentary has a similar shape to that of other small (in these cases represented by juvenile specimens) megalosauroids such as Dubreuillosaurus (CitationAllain 2002) and Eustreptospondylus (CitationSadleir et al. 2008), and more basal theropods such as Dilophosaurus (CitationWelles 1984). Similar numbers of teeth are present in Dubreuillosaurus and Eustreptospondylus as well as other taxa. CitationRauhut (2003) suggested that Eustreptospondylus was a subjective junior synonym of Magnosaurus on the basis of characters shared by the two taxa that appeared to be exclusive among Middle Jurassic theropods. He formulated the following diagnosis for Magnosaurus (CitationRauhut 2003, p. 20): ‘Lateral nutrient foramina placed in a shallow longitudinal groove with a subrectangular cross-section on posterior part of dentary; squamosal with lateral flange on the rim of the infratemporal fenestra’. However, the squamosal is not preserved in OUMNH J.12143, and the well-defined groove () cited by CitationRauhut (2003), and other shared features of the dentaries of Eustreptospondylus and Magnosaurus, are also present in the middle Bathonian taxon Dubreuillosaurus from Normandy (CitationAllain 2002; CitationSadleir et al. 2008). This suggests that the groove is more widely distributed among megalosauroids and cannot be used to support an exclusive clade of Eustreptospondylus and Magnosaurus. CitationSadleir et al. (2008) also noted that the interdental plates of Magnosaurus are taller than broad, whereas those of Eustreptospondylus are broader than tall. On this basis they reinstated the validity of Eustreptospondylus.
CitationVon Huene (1932) tentatively referred the holotype tooth of ‘Megalosaurus lydekkeri’ (BMNH 41352) to Magnosaurus to form the new combination Magnosaurus lydekkeri. However, this specimen is an isolated tooth lacking diagnostic features. ‘M. lydekkeri’ is generally considered to be a nomen dubium (CitationCarrano & Sampson 2004; CitationHoltz et al. 2004), and its referral to Magnosaurus is unjustified.
Description
Dentary
The dentary is expanded dorsoventrally towards its anterior end and has a slightly upturned, rounded anterior termination. In dorsal view the outline of the lateral wall of the dentary is convex, swollen laterally around an enlarged third dentary tooth, whereas the outline of the medial wall is straight. Eleven alveoli are preserved in the right dentary (OUMNH J.12143/1b; ) and fourteen are estimated to have originally been present by comparison with the dentaries of Eustreptospondylus and Dubreuillosaurus, which are of similar size and general morphology. The interdental plates are damaged, but formed individual polygonal plates medial to the alveoli. They have smooth medial surfaces, are moderately high relative to their basal width compared to other theropods (), and are relatively higher than those of Dubreuillosaurus (CitationAllain 2002) and Eustreptospondylus (CitationSadleir et al. 2008).
The Meckelian groove is shallow but distinct, and weakly inclined anteroventrally. Two Meckelian foramina are present in the vicinity of the anterior termination of the Meckelian groove. They are arranged so that the more anterior of the two is located almost directly anterior to the more posterior (). The spacing between the Meckelian foramina is wide relative to that in Eustreptospondylus (CitationSadleir et al. 2008; OUMNH J.13558) and Dubreuillosaurus (; CitationAllain 2002; MNHN 1998-13).
The symphysial region of the medial surface of the dentary lacks a distinct facet for articulation with the opposite element. A broad horizontal groove, upturned at its anterior end, is present in this position and is bounded dorsally and ventrally by short, rounded ridges. The same morphology is present in the megalosaurid megalosauroids Dubreuillosaurus (; CitationAllain 2002), Eustreptospondylus (CitationSadleir et al. 2008; OUMNH J.13558), and Duriavenator hesperis (CitationBenson 2008a, ), but is absent in the spinosaurid Baryonyx (CitationCharig & Milner 1997; BMNH R9951).
Numerous foramina (12 in OUMNH J.12143/1b), most arranged randomly, are situated anteriorly on the lateral surface of the dentary. The most posterior three of these are arranged into a horizontal row. Additional foramina are located more posteriorly in a well-defined horizontal groove at about two-thirds the height of the lateral surface of the dentary. Three such foramina are preserved in the right dentary. They are anteroposteriorly elongate and slit-like, and open dorsolaterally.
Anteroposteriorly elongate foramina are present on the lateral surface of the dentary adjacent to its ventral margin at the level of the seventh and tenth alveoli (). They superficially resemble grooves due to their elongate morphology. However, unlike grooves they penetrate the bone surface. They are oriented anterodorsally and a shallow channel extends anterodorsally from the foramen at the level of the seventh alveolus (). In other theropods such as Baryonyx (BMNH R9951) and Dubreuillosaurus (MNHN 1998–13), a row of foramina parallel to the ventral margin of the bone is present at this level on the lateral surface but in none of these taxa are the openings so elongated, and this morphology is considered as a potential autapomorphy of Magnosaurus. Although the foramina in Dubreuillosaurus appear to be suboval and slightly elongated in the figures of CitationAllain (2002, Fig. 15), they are substantially shorter than those of Magnosaurus and do not extend anterodorsally as grooves. Furthermore, the long axes of the foramina of Dubreuillosaurus are horizontal, whereas those of Magnosaurus are oriented anterodorsally. A third elongate foramen located at the level of the fifth alveolus in Magnosaurus is oriented horizontally ().
Broken adult teeth are present in the second, fourth, and ninth alveoli of the right dentary and the fourth, fifth, and seventh alveoli of the left dentary. Emerging teeth are present in the first, third, and fifth alveoli of the right dentary. Broken tooth bases are present in the sixth, seventh, eighth and tenth alveoli of the right dentary and the first, second, third, sixth, eighth, ninth, and tenth alveoli of the left dentary. Replacement teeth are visible between the interdental plates medial to the second, third, fourth, fifth, sixth, and eighth alveoli of the right dentary and the second, fourth, sixth, and eighth alveoli of the left dentary. The carinae are arranged almost symmetrically placed; the posterior carina is slightly offset towards the labial side of the tooth and the anterior carina is slightly offset toward the lingual side of the tooth. The denticles are chisel-shaped and not inclined apically. Enamel wrinkles and interdenticular sulci (sensu CitationSmith 2007) are both absent. The anterior carina extends less than one-third of the way towards the base of the tooth crown. Based on the distal carinae of the fifth tooth of the left dentary and the ninth tooth of the right dentary there are 28–30 denticles per centimetre. Measurements were taken just apically of midlength of the carina. Other sections of the carinae are inaccessible but the serration density of the mesial carina is estimated as being approximately equal to that of the distal carina.
Vertebrae
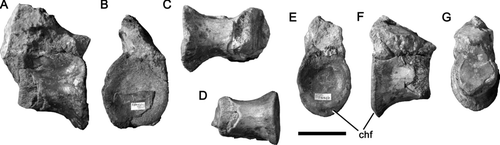
Two vertebrae are preserved (measurements in ). OUMNH J.12143/9 is a partial dorsal vertebra that preserves the centrum and base of the neural arch. The posterior end of the vertebra is abraded (). OUMNH J.12143/8 is a caudal vertebra and preserves the centrum and base of the neural arch (). The rim of the anterior articular surface is abraded. The ventral part of the posterior surface is bevelled for articulation with a chevron. The centra and neural arches are co-ossified, although the neurocentral sutures are still visible. In the similarly-sized vertebrae of Eustreptospondylus the neurocentral sutures are totally unfused (CitationSadleir et al. 2008), this probably indicates that the holotype of Magnosaurus was closer to ontogenetic maturity and had a much smaller adult size. Although CitationIrmis (2007) observed considerable variation in the timing of neurocentral suture closure among archosaurs, it is likely that Magnosaurus and Eustreptospondylus shared similar patterns due to their close systematic relationship (both are megalosaurids). In both vertebrae of Magnosaurus, the articular surfaces are gently concave and higher than they are wide, broad pleurocentral depressions are present on the lateral surfaces of the centra ventral to the closed neurocentral suture, although pleurocoels are absent. A weak longitudinal ridge is present on the ventral surfaces of both centra.
Table 1 Selected measurements (in millimetres) of Magnosaurus nethercombensis vertebrae. Abbreviations: anterior dv, dorsoventral height of anterior surface of centrum; anterior ml, mediolateral width of anterior surface of centrum; posterior dv, dorsoventral height of posterior surface of centrum; posterior ml, mediolateral width of posterior surface of centrum; i, measurement incomplete due to damage; e, measurement estimated from incomplete specimen.
Figure 3 Vertebrae of Magnosaurus nethercombensis. A-C, dorsal vertebra OUMNH J.12143/9 in right lateral (A), anterior (B), and ventral (C) views; D-F, proximal caudal vertebra OUMNH J.12143/8 in ventral (D), posterior (E), right lateral (F), and anterior (G) views. Abbreviations: chf, chevron facet. Scale bar equals 50 mm.
Ilium
An iliac fragment including the ischial peduncle (OUMNH J.12143/10) preserves little informative detail (), although it is clear that the supracetabular shelf was not continuous with the lateral wall of the brevis shelf as it is in some non-tetanuran theropods (CitationTykoski & Rowe 2004). The distal end of the ischial peduncle is broken and it cannot be determined whether it bore a flat articular surface as in most non-coelurosaurian tetanurans (e.g. Sinraptor, CitationCurrie & Zhao 1994) or terminated in a tapering edge as in coelurosaurs such as Stokesosaurus (CitationBenson 2008b). Although the pubic peduncle is not preserved, comparison of the size of the ischial peduncle with the proximal end of the pubis suggests that the ischial peduncle was reduced relative to the pubic peduncle as in other tetanurans (; CitationSereno et al. 1994).
Figure 4 Pelvic bones of Magnosaurus nethercombensis. A, B, partial right ilium OUMNH J.12143/10 in lateral (A) and medial (B) views; C, D, proximal portion of left pubis OUMNH J.12143/11 in lateral (C) and medial (D) views; E-G, shaft of right pubis OUMNH J.12143/3 in medial (E), anterior (F), and posterior (G) views. Abbreviations: ipr, ischial process; isp, ischial peduncle; pua, pubic apron. Scale bar equals 100 mm.

Pubis
Fragments of the proximal ends of both pubes (left: OUMNH J.12143/12; right: OUMNH J.12143/11) and the shaft of the right pubis (OUMNH J.12143/3) are preserved. The proximal ends are relatively uninformative as the obturator flange is not preserved and the condition of the obturator foramen cannot be determined (). The pubic shaft is straight in lateral view and has a relatively narrow pubic apron (). It is therefore similar to that of Megalosaurus (CitationBenson 2010).
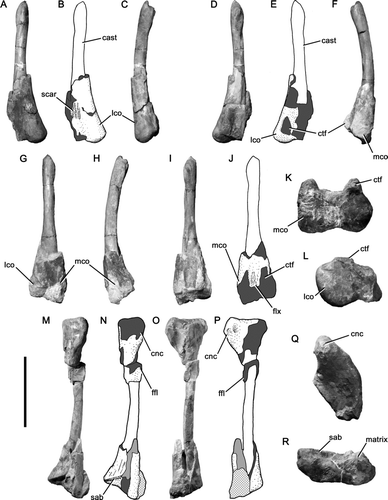
Femur
The distal ends of both femora (OUMNH J.12143/6-7) are preserved. They are abraded and their shafts are preserved as internal casts (). The casts of the shafts are gently curved posterolaterally, which may represent the original curvature of the femoral shaft. A shallow extensor groove is present distally on the anterior surface of the femur. The anterior surface of the lateral condyle is mediolaterally convex and that of the medial condyle is not preserved. A series of longitudinal striations form an oval scar on the medial side of the anterior surface of the shaft approximately 50 mm in proximodistal length and situated around 60 mm from the distal end of the medial condyle (), similar to that in other megalosauroids such as Megalosaurus (CitationBenson 2010) and Piatnitzkysaurus (PVL 4073; considered a megalosauroid (spinosauroid) by CitationHoltz et al. 2004; CitationBenson 2008a, Citationb), but unlike in most non-tetanuran theropods such as Ceratosaurus (CitationMadsen & Welles 2000) and neotetanurans such as Allosaurus (CitationMadsen 1976), in which a large oval depression that extends to the distal end of the femur is present in this location. It is notable that in specimens of Allosaurus from the Cleveland-Lloyd Dinosaur Quarry the morphology of the depression is consistent across all ontogenetic stages represented (UMNH VP specimens). It is thus unlikely that the morphologies discussed above result from differences in absolute size or age. The medial distal crest is developed as a low, rounded bar forming the anterior boundary of a shallow fossa on the medial surface of the medial condyle. Further details of the morphology of the medial condyle cannot be determined due to poor preservation. The lateral condyle has a subcircular outline in distal view and is convex. The crista tibiofibularis is poorly preserved in both femora but extends as far distally as the lateral condyle. The floor of the flexor groove between the crista tibiofibularis and the posterior extension of the medial condyle is flat and bears weak longitudinal striations. A transverse ridge connecting the crista tibiofibularis and posterior extension of the medial condyle, as is present in some more basal theropods including coelophysoids (e.g. CitationTykoski & Rowe 2004), is absent.
Figure 5 Hindlimb bones of Magnosaurus nethercombensis. A-F and L, left femur OUMNH J.12143/7 in anterior (A, B), lateral (C), posterior (D, E), medial (F), and distal (L) views; G-K, right femur OUMNH J.12143/6 in anterior (G), medial (H), posterior (I-J), and distal (K) views; M-R, left tibia OUMNH J.12143/2 in anterior (M, N), lateral (O, P), proximal (Q), and distal (R) views. In line drawings (B, E, J, N, P) crossed hatching indicates matrix, light grey tone indicates plaster, and dark grey tone indicates broken bone. Abbreviations: cnc, cnemial crest; ctf, crista tibiofibularis; ffl, fibular flange; flx, flexor groove; lco, lateral condyle; mco, medial condyle; sab, suprastragalar buttress. Scale bar equals 200 mm.
Tibia
Both tibiae are preserved in a similar style to the femora, whereby the shaft is preserved as an internal cast (). The left tibia is 490 mm long and preserves abraded portions of both proximal and distal ends (OUMNH J.12143/2), whereas the right tibia preserves only the fragmentary proximal end (OUMNH J.12143/4). The shaft of the tibia curves gently anteriorly and is slightly sigmoid in posterior view; the distal portion curves medially and the proximal portion curves laterally.
The medial condyle is slightly less broad transversely (40 mm) than is the lateral condyle (37 mm; OUMNH J.12143/4) and rises slightly higher proximally. The condyles are separated by a deep posterior cleft, as is usual for theropods. The cnemial crest is robust and the outline of this structure in lateral view is rounded. It rises higher proximally than both the condyles and is separated from the lateral condyle by a gentle incisura tibialis. A low, anteroproximally inclined ridge is present on the lateral surface of the crest. The tibia gradually expands both medially and laterally towards its distal end, which is broad (116 mm) relative to its anteroposterior thickness (52 mm) and has a triangular outline in distal view. A gentle depression on the distal surface of the bone would have articulated with a corresponding eminence on the astragalus. The astragalar facet is well-defined and 54 mm high proximodistally, indicating an ascending process of the astragalus approximately 11% of the length of the tibia. The suprastragalar buttress is a well-defined, step-like, proximolaterally oriented ridge, similar to those of most other basal tetanurans (e.g. CitationMadsen 1976).
Discussion
The validity of Magnosaurus nethercombensis
Magnosaurus nethercombensis has previously been considered as a junior synonym of Megalosaurus (von CitationHuene 1923; CitationWaldman 1974) or a senior synonym of Eustreptospondylus (CitationRauhut 2003) (see above). Only a single potential autapomorphy is identified herein. Therefore, M. nethercombensis is compared with other geographically proximate Middle Jurassic theropods here to support its validity as a distinct taxon.
Among those taxa for which the dentary is known, Magnosaurus differs from Duriavenator hesperis (Lower Bajocian, Dorset, United Kingdom) and Megalosaurus bucklandii (Lower-Middle Bathonian, Oxfordshire and Gloucestershire, United Kingdom) in possessing a dorsoventrally lower dentary with a dorsoventrally expanded anterior end (; CitationBenson 2008a; CitationBenson et al. 2008). It differs further from D. hesperis in possessing a weaker Meckelian groove (CitationBenson 2008a) and from M. bucklandii in possessing an expanded, subcircular third dentary alveolus and a well-defined groove for nutrient foramina on the lateral surface of the dentary (; CitationBenson et al. 2008). M. nethercombensis differs from Proceratosaurus bradleyi as the anterior dentary teeth are not reduced in size, the lateral surfaces of the teeth are not fluted, and only 14 teeth are present in the dentary, whereas in P. bradleyi at least 17 are present (CitationWoodward 1910).
The outline of the dentary of M. nethercombensis in lateral view resembles those of Dubreuillosaurus valesdunensis (middle Bathonian, France) and Eustreptospondylus oxoniensis (Callovian, Oxfordshire, United Kingdom), as they are dorsoventrally low with a rounded, expanded anterior end (). However, D. valesdunensis possesses very low interdental plates that appear triangular in medial view (; CitationAllain 2002) and the femur of D. valesdunensis lacks an extensor groove (CitationAllain 2005), unlike that of M. nethercombensis. The interdental plates of the dentary of E. oxoniensis are also lower dorsoventrally than those of M. nethercombensis (; CitationSadleir et al. 2008) and in both D. valesdunensis and E. oxoniensis the Meckelian foramina are closer together than those of M. nethercombensis ().
Magnosaurus nethercombensis differs from Poekilopleuron bucklandii (Middle Bathonian, France), for which the dentary is not known, as the medial malleolus of the tibia is more prominent in P. bucklandii and forms a distinct ‘shoulder’ in the outline in anterior view (Eudes-Deslongchamps 1838; CitationAllain & Chure 2002), whereas that of M. nethercombensis is less prominent (). Other overlapping elements are not known or the anatomy is too poorly known to enable comparison.
There is also little overlap between material of Afrovenator abakensis (CitationSereno et al. 1994; ?Middle Jurassic-Lower Cretaceous, Niger; CitationRauhut & López-Arbarello 2009) and M. nethercombensis. However, A. abakensis seems to lack the ventral ridge that is present in the dorsal vertebra of M. nethercombensis.
Systematic affinities of Magnosaurus
Magnosaurus nethercombensis was recovered as a megalosaurid by CitationBenson (2008c, Citationd, Citation2010), but its position within Megalosauridae is unstable. Character optimisation onto the strict reduced consensus tree of CitationBenson (2008c, d, 2010) reveals that Magnosaurus possesses two tetanuran features: a substantially reduced ischial peduncle of the ilium and the presence of an extensor groove on the anterior surface of the femur distally. Although CitationCarrano (2007) noted that the ischial peduncle of the ilium of the abelisauroid Majungasaurus was reduced relative to the pubic peduncle, this slight reduction is much less than that seen in tetanurans (CitationRauhut 2003; CitationHoltz et al. 2004).
Reanalysis of the dataset of CitationBenson (2008c, Citationd, Citation2010), excluding all taxa eliminated from the strict reduced consensus other than Magnosaurus, results in the recovery of eight trees of length 586 steps. When Magnosaurus is constrained to fall outside of Megalosauridae, it forms the sister taxon of Megalosauridae + Spinosauridae in the strict consensus of the resulting eight trees of length 588 steps. When Magnosaurus is constrained to fall outside of Megalosauroidea then it forms the sister taxon of Dilophosaurus, a non-tetanuran theropod, in the strict consensus of the resulting 16 trees of length 590 steps. This relationship is supported by shared features, such as the enlarged third alveolus of the dentary. The rugose scar located medially on the anterior surface of the distal femur of Magnosaurus is a megalosauroid synapomorphy that is also found in Dilophosaurus (UCMP 37302) and also supports their grouping when Magnosaurus is excluded from Megalosauroidea.
Within Megalosauroidea, the enlarged third dentary alveolus indicates referral to the clade Megalosauridae + Spinosauridae. Magnosaurus possesses a single unambiguous synapomorphy of Megalosauridae, a well-defined groove on the lateral surface of the dentary. Unfortunately, the presence of other megalosaurid features, such as a paradental groove that is wide anteriorly and narrow posteriorly (CitationBenson 2008a; CitationBenson et al. 2008), cannot be determined due to poor preservation. CitationRauhut (2003) stated that in his analysis the scores for OUMNH J.12143 (the holotype of M. nethercombensis) were identical to those of OUMNH J.13558 (the holotype of Eustreptospondylus) and effectively excluded OUMNH J.12143 by safe taxonomic reduction (CitationWilkinson 1995) when he synonymised Eustreptospondylus with Magnosaurus. Magnosaurus (sensu CitationRauhut 2003) was recovered as the most basal megalosauroid (spinosauroid) (CitationRauhut 2003). New anatomical data presented here supports Rauhut's (2003) implied megalosauroid affinities for M. nethercombensis.
Body size of Magnosaurus
Due to fragmentary preservation, the body size of Magnosaurus is estimated by comparison with other taxa. The tibia OUMNH J.12143/2 is 490 mm long. This is comparable with those of Piatnitzkysaurus, which are 515 mm (MACN CH 895; left tibia, possibly representing a subadult individual, O. W. M. Rauhut pers. comm., 2009) and 490 mm (PVL 4073; right tibia) long. PVL 4073 has a right femur 543 mm long and metatarsal III measuring 282 mm. Most basal tetanurans have similar hindlimb proportions. This indicates a fully extended femur–metatarsus length of around 1315 mm in Magnosaurus, suggesting that the hip was held just over one metre off the ground. Piatntizkysaurus (PVL 4073) has a minimum femoral circumference of 240 mm. This gives a body mass estimate of 504 kg using the equations of CitationAnderson et al. (1985). This method tends to underestimate body mass relative to three-dimensional modelling techniques such as that of CitationHenderson (1999). It is probable that Magnosaurus had a similar body mass to Piatnitzkysaurus but this cannot be confirmed due to poor preservation of the shafts of weight-bearing limb bones. Both taxa are small compared to many other Jurassic non-coelurosaurian tetanurans including Allosaurus, Megalosaurus, Sinraptor, and Torvosaurus, all of which weighed more than a ton (CitationMadsen 1976; CitationBritt 1991; CitationCurrie & Zhao 1994; CitationBenson 2010). However, most of these taxa are from the Late Jurassic, and Middle Jurassic theropods such as Condorraptor, Gasosaurus, Piatnitzkysaurus, and Poekilopleuron (CitationEudes-Deslongchamps 1838) are comparable in size to Magnosaurus. Nonetheless, larger Middle Jurassic theropods such as Megalosaurus and Monolophosaurus (CitationZhao & Currie 1994) are known and it is possible that some other taxa are represented by subadult specimens. The Middle Jurassic megalosaurids Dubreuillosaurus and Eustreptospondylus are represented by the remains of individuals with similar body size to Magnosaurus. However, both of these taxa are represented by juvenile specimens showing unfused neurocentral and cranial sutures, suggesting that they grew to larger sizes than Magnosaurus (CitationAllain 2002, 2005; CitationSadleir et al. 2008).
The earliest fossil records of tetanurans
Rhaetian, Wales, UK
CitationNewton (1898, pl. 10) reported the natural mould of a left dentary (BMNH R2912) from the Rhaetic Beds (Rhaetian) near Bridgend, Glamorganshire, Wales, as a new species, Zanclodon cambrensis. CitationWaldman (1974) considered that this specimen bore considerable resemblance to ‘Megalosaurus’ hesperis (now Duriavenator hesperis), and CitationMolnar (1990) referred the specimen to ?Megalosaurus based on three putative synapomorphies of the dentary. However CitationRauhut & Hungerbühler (2000) noted that these characters had a more general distribution. They also noted that BMNH R2912 lacked autapomorphies and diagnostic features of any particular theropod clade and that ‘Z. cambrensis’ is therefore a nomen dubium representing indeterminate theropod remains. Nonetheless, CitationGalton (1998, 2005) used the combination ?M. cambrensis, and CitationWeishampel et al. (2004) listed the taxon as Tetanurae incertae sedis. Rauhut & Hungerbüler (2000) and Naish & Martill (2007, p. 500) noted that the interdental plates of BMNH R2912 were low, as in theropods outside of the clade comprising Ceratosauria and Tetanurae and that the specimen probably represented a ‘coelophysoid or near-coelophysoid’. That viewpoint is followed here. I also note that the specimen, which was figured by CitationGalton (2005, Figs 10–11) and Naish & Martill (2007, ), possesses only a single Meckelian foramen, unlike basal tetanurans, which possess two (e.g. CitationMadsen 1976; CitationBenson et al. 2008).
Norian–Hettangian, Kanton Schaffhausen, Switzerland
Weishampel et al. (2004, p. 524) listed remains of ?Tetanurae indet. from an unnamed unit (?late Norian–Hettangian; CitationClemens 1980) in Kanton Schaffhausen based on data in CitationPeyer (1944) and CitationTartarinov (1985). However, neither of the latter authors reported theropod remains.
Norian, Texas, USA
CitationChatterjee (1991) erected the taxon Protoavis for many bones and fragments from the Tecovas and Bull Canyon Formations of Garza County, Texas and suggested that this material belonged to the earliest-known bird (CitationChatterjee 1991, Citation1999). However, much of the material is reconstructed and most authors believe that the material represents multiple individuals from different taxa (summarised by CitationNesbitt et al. 2007). CitationNesbitt et al. (2007) reviewed this material and suggested that the cervical vertebrae belonged to a drepanosaurid. They also noted that although other elements such as the astragalus and calcaneum may belong to theropods, the anatomy was more consistent with coelophysoid or otherwise non-tetanuran, non-neoceratosaurian affinities. The braincase of Protoavis is the only element that shows avian features (CitationWitmer 2001; CitationNesbitt et al. 2007). However CitationNesbitt & Norell (2006) showed high levels of ‘dinosaur-like’ convergent evolution among non-dinosaurian archosaurs such as Shuvosaurus. On the basis of these observations CitationNesbitt et al. (2007)recommended a thorough redescription of the ‘Protoavis’ material before any definitive statement on the avian affinities of the material could be made. In particular they considered it unlikely that the origins of birds, and therefore coelurosaurs and other tetanurans could have occurred in the Late Triassic and remain otherwise undetected in the fossil record.
A second putative coelurosaur from the Norian of Texas, Shuvosaurus, was reported by CitationChatterjee (1993) based on a single skull and possibly associated postcranial material (CitationRauhut 1997). CitationChatterjee (1993) assigned Shuvosaurus to Ornithomimosauria and this assignment was questioned by numerous subsequent authors (summarised by CitationNesbitt et al. 2007). Decisively, CitationNesbitt & Norell (2006) and CitationNesbitt (2007) described specimens of Effigia, a non-dinosaurian archosaur from the Late Triassic of New Mexico and demonstrated that Shuvosaurus was related to this taxon and not to ornithomimosaurs.
Hettangian, Yunnan, China
CitationXu et al. (2001) erected the new genus and species Eshanosaurus deguchiianus for a therizinosauroid dentary from the Lower Lufeng Formation (Hettangian) of Eshan County, Yunnan. Although some subsequent authors have suggested that this dentary represents a prosauropod (CitationKirkland & Wolfe 2001; CitationRauhut 2003), the detailed morphology of the specimen was not considered to be consistent with sauropodomorph affinities by CitationBarrett (2009), and the present author concurs that it is likely to belong to a therizinosauroid. If Eshanosaurus does represent a therizinosauroid from the Lower Jurassic then, under most recent hypotheses of theropod relationships, phylogenetic ghost lineages imply the presence of representatives of other major coelurosaurian clades including Ornithomimosauria, Paraves and Oviraptorosauria during this epoch. Representatives of these clades are not currently known until the Upper Jurassic (von CitationMeyer 1861; CitationXu et al. 2006) and must have remained undetected for around 40 million years if they originated at the start of the Jurassic period. Numerous Jurassic theropod fossils are known (CitationWeishampel et al. 2004), and these all conform to a more conservative conception of the timing of origin of superfamily-level clades. A recent review of therizinosauroids (CitationClark et al. 2004) considered the systematic position of Eshanosaurus as tentative and the age of the Lower Lufeng Formation as poorly constrained. CitationBarrett (2009) suggested that further work was required to confirm the Lower Jurassic age of Eshanosaurus. On this basis, Eshanosaurus should not be regarded as the earliest tetanuran until better stratigraphic data are available.
Hettangian–Sinemurian, Dorset, UK
CitationNewman (1968) identified three specimens from the Blue Lias (Hettangian–early Sinemurian; CitationGetty 1980) of Charmouth, Dorset, UK, as theropod (‘megalosaurid’). These specimens were originally included in the syntype series of Scelidosaurus: a distal femur with articulated proximal tibia and fibula (BMNH 39496; ), a partial small left femur (GSM 109560; ; CitationOwen 1861: pl. 1, –), and an ungual phalanx (GSM 109561). CitationNaish & Martill (2007) stated that this material belonged to a tetanuran. CitationCarrano & Sampson (2004) suggested that BMNH 39496 represented one of the earliest tetanurans, but neither CitationCarrano & Sampson (2004) nor CitationNaish & Martill (2007) provided observations supporting these affiliations and none of the specimens shows unequivocal anatomical evidence for a tetanuran affinity.
Figure 6 Theropod hindlimb bones from the Blue Lias of Charmouth, Dorset. A-F and L right femur BMNH 39496 in anterior (A, B), medial (C), posterior (D, E), lateral (F), and distal (L) views; G-K and M, right tibia BMNH 39496 in anterior (G), medial (H), posterior (I), lateral (J, K), and proximal (M) views; N-S, left femur GSM 109560 in medial (N, O), anterior (P, Q), posterior (R), and lateral (S) views. In line drawings (B, E, K, O, Q) dark grey tone indicates broken bone and light grey tone indicates reconstructed areas. Abbreviations: ail, anterior intermuscular line; cnc, cnemial crest; ctf, crista tibiofibularis; ffl, fibular flange; for, foramen; ft, fourth trochanter; ict, incisura tibialis; lco, lateral condyle; lt, lesser trochanter; mco, medial condyle; mdc, medial distal crest; trs, trochanteric shelf. Scale bars equal 100 mm.
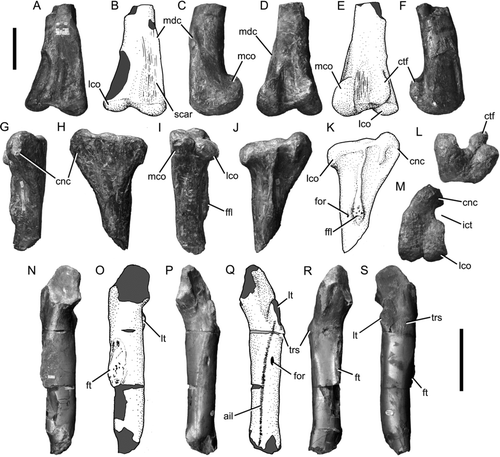
BMNH 39496 was partly figured by CitationNewman (1968, pl. 7, , pl. 8, , , , 8, 10, 13) but is figured again here with line drawings for clarity (). The fibula cannot currently be located. The femur of BMNH 39496 lacks an extensor groove, the anterior surface of the distal end is therefore approximately flat as in Dubreuillosaurus (CitationAllain 2005) and non-tetanuran theropods (e.g. CitationMadsen & Welles 2000; CitationTykoski & Rowe 2004). The medial distal crest is not as strongly developed as those of taxa that possess a prominent medial distal crest such as coelophysoids, neoceratosaurs (e.g. Ceratosaurus; CitationMadsen & Welles 2000) and neotetanurans (e.g. Neovenator; CitationBrusatte et al. 2008), but is more prominent than the low bar present in taxa such as Dilophosaurus (CitationWelles 1984) and megalosauroids (e.g. Magnosaurus and Megalosaurus; CitationBenson 2010). A proximodistally elongate, oval depression is present on the anterior surface of the femur adjacent to the crest, as in other taxa possessing a prominent medial distal crest such as Allosaurus (CitationMadsen 1976). The crista tibiofibularis and the posterior extension of the medial condyle are prominent, tuberous structures. A groove connects the lateral margin of the base of the crista tibiofibularis to the center of the distal surface of the femur as in basal theropods such as Dilophosaurus (UCMP 37302) and Megalosaurus (CitationBenson 2010), but unlike neotetanurans such as Allosaurus (CitationMadsen 1976; CitationBenson 2010) and Sinraptor (CitationCurrie & Zhao 1994).
The cnemial crest of the tibia of BMNH 39496 is prominent and rises dorsally above the level of the proximal surface of the tibia. As in most theropods, the crest is separated from the lateral condyle by a deep concavity in the outline of the bone in proximal view, the incisura tibialis (CitationRauhut 2003). The condyles are subequal in width and are squared off posteriorly in proximal view. The fibular flange is expanded anteroposteriorly so that it has a suboval outline in lateral view as in the tetanurans Megalosaurus (CitationBenson 2010), Piatnitzkysaurus (PVL 4073), and Sinraptor (CitationCurrie & Zhao 1994). However, it extends to the proximal end of the tibia as a prominent ridge as in non-tetanuran theropods (CitationRauhut 2003).
When included in the phylogenetic data matrix of CitationBenson (2008c, Citationd, Citation2010) (Appendix), analyzed using the search strategy outlined by CitationBenson (2010), BMNH 39496 is recovered as the sister taxon of ‘Syntarsus’ kayentakatae in the strict consensus of the resulting eight trees of length 587 steps. However, this data set was constructed to resolve basal tetanuran relationships and the affinities of BMNH 39496 among non-tetanurans should be considered unresolved pending inclusion among a broader taxon sample. Nonetheless, if BMNH 39496 showed decisively tetanuran morphology it would be expected to fall within the well-sampled tetanuran clade. Future discoveries of more complete specimens may demonstrate that BMNH 39496 is the earliest tetanuran. However, the combination of anatomical features known presently does not unequivocally support either a tetanuran or non-tetanuran affinity.
The left femur (GSM 10960) from Charmouth is abraded proximally and distally () and is much less well-preserved than indicated by the heavily reconstructed specimen figured by CitationNewman (1968, pl. 7, ). The head is directed anteromedially, although only the base is preserved so the exact orientation cannot be determined. The lesser trochanter is abraded and its original morphology cannot be determined. A robust buttress extends distomedially from the base of the lesser trochanter. The apex of this buttress is scored by transverse striations representing the anterior intermuscular line (). A well developed and rugose, mound-like trochanteric shelf protrudes from the lateral surface of the femur adjacent to the base of the lesser trochanter, from there a short ridge extends distally along the lateral margin of the posterior surface. The fourth trochanter is a low flange. A rugose, proximodistally oval muscle scar occupies its medial surface. GSM 10960 does not preserve any features characteristic of a particular clade within Theropoda. The ungual phalanx from Charmouth (GSM 10961) has been reduced to a stump by abrasion. Therefore, both specimens represent theropods of indeterminate affinities.
Sinemurian, Regione Lombardia, Italy
CitationDal Sasso (2003) briefly described fragmentary theropod remains from the Saltrio Formation (Sinemurian) near Saltrio. Weishampel et al. (2004, p. 533) listed these as an undescribed tetanuran. However, CitationDal Sasso (2003) suggested that the specimen represented a transitional form between Ceratosauria and Tetanurae and only possibly a basal tetanuran. He listed three putatively tetanuran features of the specimen: the proximal articular facet of manual phalanx III-1 is a single depression, rather than two depressions in non-tetanurans; a low degree of supination of the manual phalanges; and the presence of fused clavicles (a furcula). However, the first of these features was also described in Dilophosaurus by CitationWelles (1984), and contrary to Dal Sasso's (2003) observation, the distal articular surface of metacarpal III bears a double condyle in tetanurans such as Allosaurus (CitationMadsen 1976, pl. 44). Additionally, a true furcula has now been described in several non-tetanuran theropods (CitationTykoski et al. 2002; CitationRauhut 2003; CitationCarrano et al. 2005; CitationRhinehart et al. 2007). Osteological correlates associated with the ‘low degree of supination of the manual phalanges’ of tetanurans described by CitationDal Sasso (2003, p. 61) were not proposed and confirmation of this condition as a tetanuran synapomorphy awaits more detailed description and comparisons. On the basis of these observations, the systematic position of the Italian specimen should be considered uncertain pending new data.
Sinemurian–Pliensbachian, Antarctica
Cryolopho- saurus ellioti from the Hanson Formation (Sinemurian– Pliensbachian) of Antarctica was originally described as an allosauroid tetanuran (CitationHammer & Hickerson 1994). It was subsequently recovered as an allosauroid by the phylogenetic analysis of CitationSereno et al. (1996). However, a comprehensive description of the taxon, including a reassessment of basal theropod relationships revealed that the taxon was a theropod outside of the clade comprising Ceratosauria and Tetanurae (CitationSmith et al. 2007). Although Carrano & Sampson (2008, p. 214) described Cryolophosaurus as ‘the oldest confirmed tetanuran’, supporting data was not provided. It is also probable that this comment was committed before publication of CitationSmith et al. (2007).
Toarcian, Morocco
CitationJenny et al. (1980) reported a partial theropod skeleton from the Toarcian of Morocco. CitationTaquet (1985) referred to this as a coelurosaur and CitationAllain (2002) stated that it was a basal tetanuran, both for unspecified reasons. CitationTaquet (1985) figured parts of the hindlimb. From these figures the taxon appears to be a non-tetanuran theropod. The specimen does not possess any tetanuran synapomorphies and shows a low, block-like ascending process of the astragalus. This is unlike the condition in coelurosaurs where the process forms a tall, thin sheet that extensively overlaps the anterior surface of the distal tibia (CitationWelles & Long 1974). It is also unlike the condition in basal tetanurans in which the ascending process is taller and anteroposteriorly thinner than in non-tetanurans (CitationWelles & Long 1974).
Toarcian, Whitby, UK
CitationOwen (1842) described a posterior dorsal vertebra from the Lias Shales (Toarcian; CitationWalker 1964) of Whitby, in the collection of Mr Ripley, as one of the syntypes of Streptospondylus cuvieri. The specimen is currently considered lost and was never figured. CitationOwen (1842) described anterior and posterior centrodiapophyseal laminae, and pleurocentral depressions. CitationOwen (1842) cited the opisthocoelous centrum, constricted at midlength and approximately as long as high, as diagnostic features enabling a referral to Streptospondylus. However, all of these are now known to be more generally present in basal tetanurans (e.g. CitationMadsen 1976). It is possible that this specimen represents the earliest known basal tetanuran; however, it may also belong to a small sauropod and since the specimen is lost its affinities cannot be confirmed.
Tetanuran origins and diversification
Although Magnosaurus is the earliest definitive tetanuran, it is a megalosaurid megalosauroid and therefore is not the most basal tetanuran taxon. CitationRauhut (2003) and CitationSmith et al. (2007) recovered Piatnitzkysaurus or a clade of Piatnitzkysaurus and Condorraptor as the most basal tetanurans, branching off outside the clade containing Megalosauroidea, Allosauroidea and Coelurosauria. CitationHoltz et al. (2004) recovered ‘Szechuanosaurus’ zigongensis and Xuanhanosaurus as the most basal tetanurans, forming successively more proximate outgroups to the major tetanuran clades. CitationBenson (2008c, Citationd, Citation2010) recovered most of these possible basalmost tetanurans within Megalosauroidea. Within Megalosauroidea, Megalosauridae was resolved as the sister taxon of Spinosauridae. A clade comprising Condorraptor, Piatnitzkysaurus, Marshosaurus, and Xuanhanosaurus and a clade comprising Chuandongocoelurus and Monolophosaurus were recovered as successively more proximate outgroups to Megalosauridae + Spinosauridae within Megalosauroidea. Under any of these hypotheses of basal tetanuran relationships, several clades of basal tetanurans other than Megalosauridae must have been present by the origin of Magnosaurus.
Relationships between Allosauroidea, Coelurosauria and Megalosauroidea (Spinosauroidea) remain uncertain. Although most authors recover a monophyletic Neotetanurae, comprising Allosauroidea and Coelurosauria, as the sister taxon of Megalosauroidea (CitationHoltz et al. 2004; CitationYates 2006; CitationSmith et al. 2007), CitationRauhut (2003) recovered a monophyletic Carnosauria comprising Allosauroidea and Megalosauroidea to the exclusion of Coelurosauria and branch support values for both arrangements are generally low. The earliest coelurosaur, Proceratosaurus, a tyrannosauroid (CitationRauhut & Milner 2008; Li et al. 2009), is from the Bathonian of the UK (CitationWoodward 1910). Shidaisaurus, from the poorly constrained Upper Lufeng Formation (?early Middle Jurassic) was compared closely with Sinraptor and Yangchuanosaurus by CitationWu et al. (2009) and may be the earliest allosauroid. This provides evidence that representatives of all three major tetanuran clades were present by the first half of the Middle Jurassic.
These observations suggest that basal tetanuran diversification was well underway in the early Middle Jurassic. The globally poor Middle and late Early Jurassic record of theropod dinosaurs (e.g. CitationWeishampel et al. 2004) is an impediment to understanding this early diversification.
‘Ceratosaur’ paraphyly
Two major hypotheses of basal theropod relationships have been proposed. CitationGauthier (1986) performed the first cladistic analysis to include a representative sample of theropod dinosaurs. He included a single supraspecific taxon for the sister taxon of Tetanurae including Late Triassic–Early Jurassic coelophysoids such as Coelophysis and Late Jurassic neoceratosaurs such as Ceratosaurus. The proposed monophyly of this taxon, for which he used CitationMarsh's (1884) name Ceratosauria, was based primarily on the presence of a shelf-like trochanteric shelf in some specimens. Some subsequent cladistic studies corroborated Gauthier's (1986) conception of Ceratosauria (CitationRowe 1989; CitationRussell & Dong 1993). However, more recently a debate has emerged over the monophyly of Ceratosauria sensu CitationGauthier (1986). Although some analyses support this notion (CitationPérez-Moreno et al. 1993; CitationHoltz 1994, Citation2000; CitationSereno 1999; CitationTykoski & Rowe 2004; CitationAllain et al. 2007), others recover a paraphyletic ‘ceratosaur’ lineage subtending two or three smaller clades leading up to Tetanurae, only the most proximate of which can be termed Ceratosauria (CitationForster 1999; CitationCarrano et al. 2002; CitationRauhut 2003; Sereno et al. 2004; CitationYates 2006; CitationSmith et al. 2007). This issue is not resolved and the anatomical evidence supporting these opposing hypotheses has yet to be reviewed comprehensively. The timing of tetanuran origins has become a key component of this debate (CitationCarrano & Sampson 1999, 2008; CitationRauhut 2003). If Ceratosauria, the sister taxon of Tetanurae, is a large, inclusive clade, then the earliest tetanurans must have been present alongside the earliest coelophysoids in the Carnian–Norian (Late Triassic, c. 220 Mya). On the other hand, if Ceratosauria is a small clade comprising Kimmeridgian–Tithonian (Late Jurassic) taxa such as Ceratosaurus and Elaphrosaurus, and abelisauroids from the Cretaceous and possibly the Middle Jurassic ‘Ozraptor’ (CitationRauhut 2005; but see CitationCarrano & Sampson 2008) or the late Early Jurassic Berberosaurus (CitationAllain et al. 2007; c. 190–175 Mya), then the implied ghost lineage of Tetanurae is shorter, indicating a younger origin for the clade. To date, no phylogenetic analysis has recovered any undisputed taxon within Tetanurae earlier than the Middle Jurassic. Furthermore, a review of putative early tetanuran records (above) demonstrates that Magnosaurus (Early Bajocian, Middle Jurassic; c. 175 Mya) is the oldest known tetanuran. This provides stratigraphic data more consistent with a pectinate ‘ceratosauria’ (sensu CitationGauthier 1986) and restricted content of true Ceratosauria.
Conclusions
Magnosaurus nethercombensis is a valid taxon that is anatomically distinct. In particular it shows differences from all other European Middle Jurassic theropods and a single potential autapomorphy. M. nethercombensis is a megalosaurid megalosauroid (Dinosauria, Theropoda). It is Lower Bajocian in age and therefore the oldest-known definitive tetanuran. All purported records of older tetanurans are shown to be unreliable at present. A Middle Jurassic first appearance of Tetanurae is consistent with a paraphyletic ‘ceratosaur’ lineage.
Appendix
Character scorings for BMNH 39496 based on the character list of Benson (2008c, d).
?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ??01100101 001??????? ???
Acknowledgements
I particularly thank P. Jeffrey and H. Philip Powell (OUMNH) for access to and curation of specimens. I also thank Sandra Chapman (BMNH), Louise Neep (GSM), Alejandro Kramarz (MACN), Philippe Taquet (MNHN), and Jaime Powell (PVL). Thanks go to P. M. Barrett and D. B. Norman for supervision and encouragement. N. D. Smith and O. W. M. Rauhut provided constructive comments on an earlier version of the manuscript. This research was supported by NERC studentship NER/S/A/2005/13488 and comparative study was aided by travel grants from the Palaeontographical Society, SYNTHESYS and The Jurassic Foundation.
Related Research Data
References
- Allain , R. 2002 . Discovery of a megalosaur (Dinosauria, Theropoda) in the middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae . Journal of Vertebrate Paleontology , 22 : 548 – 563 .
- Allain , R. 2005 . The postcranial anatomy of the megalosaur Dubreuillosaurus valesdunensis (Dinosauria Theropoda) from the Middle Jurassic of Normandy, France . Journal of Vertebrate Palaeontology , 25 : 850 – 858 .
- Allain , R. and Chure , D. J. 2002 . Poekilopleuron bucklandii, the theropod dinosaur from the Middle Jurassic (Bathonian) of Normandy . Palaeontology , 45 : 1107 – 1121 .
- Allain , R. , Tykoski , R. , Aquesbi , N. , Jalil , N. -E. , Monbaron , M. , Russell , D. and Taquet , P. 2007 . An abelisauroid (Dinosauria: Theropoda) from the Early Jurassic of the High Atlas Mountains, Morocco, and the radiation of ceratosaurs . Journal of Vertebrate Paleontology , 27 : 610 – 624 .
- Anderson , J. F. , Hall-Martin , A. and Russell , D. A. 1985 . Long-bone circumference and weight in mammals, birds and dinosaurs . Journal of Zoology , 207 : 53 – 61 .
- Barrett , P. M. 2009 . The affinities of the enigmatic dinosaur Eshanosaurus deguchiianus from the Early Jurassic of Yunnan Province, People's Republic of China . Palaeontology , 52 : 681 – 688 .
- Benson , R. B. J. 2008a . A redescription of ‘Megalosaurus’ hesperis (Dinosauria, Theropoda) from the Inferior Oolite (Bajocian, Middle Jurassic) of Dorset, United Kingdom . Zootaxa , 1931 : 57 – 67 .
- Benson , R. B. J. 2008b . New information on Stokesosaurus, a tyrannosauroid (Dinosauria: Theropoda) from North America and the United Kingdom . Journal of Vertebrate Paleontology , 28 : 732 – 750 .
- Benson , R. B. J. 2008c . “ Reassessment of Middle Jurassic theropod relationships with a focus on the UK record ” . In 6th meeting of the European Association of Vertebrate Palaeontologists, 30th June-5th July, Spišska Nová Ves, Slovak Republic , Edited by: Krempaská , Z. 15 – 16 . Spišska Nová Ves, , Slovakia : The Museum of Spiš .
- Benson , R. B. J. 2008d . A new theropod phylogeny focussing on basal tetanurans, and its implications for European ‘megalosaurs’ and Middle Jurassic dinosaur endemism . Journal of Vertebrate Paleontology , 28 : 51A
- Benson , R. B. J. 2009 . An assessment of variability in theropod dinosaur remains from the Bathonian (Middle Jurassic) of Stonesfield and New Park Quarry, UK and taxonomic implications for Megalosaurus bucklandii and Iliosuchus incognitus . Palaeontology , 52 : 857 – 877 .
- Benson , R. B. J. 2010 . A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the United Kingdom and the relationships of Middle Jurassic theropods . Zoological Journal of the Linnean Society , doi is 10.1111/j.1096-3642.2009.00569.x.
- Benson , R. B. J. , Barrett , P. M. , Powell , H. P. and Norman , D. B. 2008 . The taxonomic status of Megalosaurus bucklandii (Dinosauria, Theropoda) from the Middle Jurassic of Oxfordshire, UK . Palaeontology , 51 : 419 – 424 .
- Britt , B. B. 1991 . Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri . Brigham Young University Geology Studies , 37 : 1 – 72 .
- Brusatte , S. L. , Benson , R. B. J. and Hutt , S. 2008 . The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden Group (Barremian) of the Isle of Wight . Palaeontographical Society Monographs , 162 ( 631 ) : 1 – 75 .
- Callomon , J. H. 2003 . The Middle Jurassic of western and northern Europe: its subdivisions, geochronology and correlations . Geological Survey of Denmark and Greenland Bulletin , 1 : 61 – 73 .
- Carrano , M. T. 2007 . The appendicular skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar . Society of Vertebrate Paleontology Memoir , 8 : 163 – 179 .
- Carrano , M. T. and Sampson , S. D. 1999 . Evidence for a paraphyletic “Ceratosauria” and its stratigraphic implications for theropod dinosaur evolution . Journal of Vertebrate Paleontology , 19 : 36A
- Carrano , M. T. and Sampson , S. D. 2004 . A review of coelophysoids (Dinosauria: Theropoda) from the Early Jurassic of Europe, with comments on the late history of the Coelophysoidea . Neues Jahrbuch für Geologie und Paläontologie Monatshefte , 2004 : 537 – 558 .
- Carrano , M. T. and Sampson , S. D. 2008 . The phylogeny of Ceratosauria (Dinosauria: Theropoda) . Journal of Systematic Palaeontology , 6 : 183 – 236 .
- Carrano , M. T. , Sampson , S. D. and Forster , C. A. 2002 . The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar . Journal of Vertebrate Paleontology , 22 : 510 – 534 .
- Carrano , M. T. , Hutchinson , J. H. and Sampson , S. D. 2005 . New information on Segisaurus halli, a small theropod dinosaur from the Early Jurassic of Arizona . Journal of Vertebrate Paleontology , 25 : 835 – 849 .
- Charig , A. J. and Milner , A. C. 1997 . Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey . Bulletin of the Natural History Museum, Geology Series , 53 : 11 – 70 .
- Chatterjee , S. 1991 . Cranial anatomy and relationships of a new Triassic bird from Texas . Philosophical Transactions of the Royal Society of London, Series B , 332 : 277 – 342 .
- Chatterjee , S. 1993 . Shuvosaurus, a new theropod . National Geographic Research and Exploration , 9 : 284
- Chatterjee , S. 1999 . Protoavis and the early evolution of birds . Palaeontographica Abteilung A , 254 : 1 – 100 .
- Clark , J. M. , Maryanska , T. and Barsbold , R. 2004 . “ Therizinosauroidea ” . In The Dinosauria, , 2nd edition , Edited by: Weishampel , D. B. , Dodson , P. and Osmólska , H. 151 – 164 . Berkeley : University of California Press .
- Clemens , W. A. 1980 . Rhaeto-Liassic mammals from Switzerland and West Germany . Zitteliana , 5 : 51 – 92 .
- Currie , P. J. and Zhao , X. -J. 1994 . A new carnosaur (Dinosauria: Theropoda) from the Jurassic of Xinjiang, People's Republic of China . Canadian Journal of Earth Sciences , 30 : 2037 – 2081 . [for 1993]
- Dal Sasso , C. 2003 . Dinosaurs of Italy . Comptes Rendus Palevol , 2 : 45 – 66 .
- Day , J. J. and Barrett , P. M. 2004 . Material referred to Megalosaurus (Dinosauria: Theropoda) from the Middle Jurassic of Stonesfield, Oxfordshire, England: one taxon or two? . Proceedings of the Geologists’ Association , 115 : 359 – 366 .
- Delair , J. B. 1959 . The Mesozoic Reptiles of Dorset: Part 2 . Proceedings of the Dorset Natural History and Archaeological Society , 80 : 52 – 90 .
- Eudes-Deslongchamps , J. A. 1838 . Mémoire sur le Poekilopleuron bucklandii, grand saurian fossile, intermédiare entre les crocodiles et les lizards . Mémoire de la Société Linnéenne de Normandie , 6 : 37 – 146 .
- Forster , C. A. 1999 . Gondwanan dinosaur evolution and biogeographic analysis . Journal of African Earth Sciences , 28 : 169 – 185 .
- Galton , P. M. 1998 . Saurischian dinosaurs from the Upper Triassic of England: Camelotia (Prosauropoda, Melanorosauridae) and Avalonianus (Theropoda, ?Carnosauria) . Palaeontographica Abteilung A , 250 : 155 – 172 .
- Galton , P. M. 2005 . Bones of large dinosaurs (Prosauropoda and Stegosauria) from the Rhaetic Bone Bed (Upper Triassic) of Aust Cliff, southwest England . Revue de Paléobiologie , 24 : 51 – 74 .
- Gauthier , J. A. 1986 . “ Saurischian monophyly and the origin of birds ” . In The Origin of Birds and the Evolution of Flight , Edited by: Padian , K. Vol. 8 , 1 – 55 . Memoirs of the California Academy of Science .
- Getty , T. A. 1980 . “ Hettangian and Sinemurian correlation chart ” . In A Correlation of Jurassic Rocks in the British Isles. Part One: Introduction and Lower Jurassic , Edited by: Cope , J. C. W. , Getty , T. A. , Howarth , M. K. , Morton , N. and Torrens , H. S. Vol. 14 , 33 – 47 . London : Special Reports of the Geological Society .
- Gradstein , F. M. , Ogg , J. G. and Smith , A. G. 2004 . A Geologic Time Scale 2004 , 610 Cambridge : Cambridge University Press .
- Hammer , W. R. and Hickerson , W. J. 1994 . A crested theropod dinosaur from Antarctica . Science , 264 : 828 – 830 .
- Henderson , D. M. 1999 . Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing . Paleobiology , 25 : 88 – 106 .
- Holtz , T. R. Jr . 1994 . The phylogenetic position of the Tyrannosauridae: implications for theropod systematics . Journal of Paleontology , 68 : 1100 – 1117 .
- Holtz , T. R. Jr . 2000 . A new phylogeny of the carnivorous dinosaurs . Gaia , 15 : 6 – 61 . [for 1998]
- Holtz , T. R. Jr , Molnar , R. E. and Currie , P. J. 2004 . “ Basal Tetanurae ” . In The Dinosauria, , 2nd edition , Edited by: Weishampel , D. B. , Dodson , P. and Osmólska , H. 71 – 110 . Berkeley : University of California Press .
- Huene , F. von . 1923 . Carnivorous Saurischia in Europe since the Triassic . Geological Society of America Bulletin , 34 : 449 – 458 .
- Huene , F. von . 1926a . The carnivorous Saurischia in the Jura and Cretaceous formations, principally in Europe . Revista del Museo de La Plata , 29 : 35 – 167 .
- Huene , F. von . 1926b . On several known and unknown reptiles of the order Saurischia from England and France . Annals and Magazine of Natural History, Series 9 , 17 : 473 – 489 .
- Huene , F. von . 1932 . Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte . Monographien zur Geologie und Palaontologie, Series 1 , 4 : 1 – 361 .
- Huxley , T. H. 1869 . On the upper jaw of Megalosaurus . Quarterly Journal of the Geological Society , 25 : 311 – 314 .
- Irmis , R. B. 2007 . Axial skeleton ontogeny in the Parasuchia (Archosauria: Pseudosuchia) and its implications for ontogenetic determination in archosaurs . Journal of Vertebrate Paleontology , 27 : 350 – 361 .
- Jenny , J. , Jenny-Deshusses , C. , Le Marrec , A. and Taquet , P. 1980 . Découverte d’ossements dans le Jurassique inférieur (Toarcien) du Haut Atlas central (Maroc) . Comptes Rendus de l’Académie des Sciences , 290 : 839 – 842 .
- Kirkland , J. I. and Wolfe , D. G. 2001 . First definitive therizinosauroid (Dinosauria: Theropoda) from North America . Journal of Vertebrate Paleontology , 21 : 410 – 414 .
- Li , D. , Norell , M. A. , Gao , K. , Smith , N. D. and Makovicky , P. J. 2010 . A longirostrine tyrannosauroid from the Early Cretaceous of China . Proceedings of the Royal Society of London, Series B , 277 : 183 – 190 .
- Madsen , J. H. Jr . 1976 . Allosaurus fragilis: a revised osteology . Utah Geological and Mineralogical Survey, Bulletin , 109 : 3 – 163 .
- Madsen , J. H. Jr and Welles , S. P. 2000 . Ceratosaurus (Dinosauria, Theropoda) a revised osteology . Miscellaneous Publications of the Utah Geological Survey , 2 : 1 – 80 .
- Marsh , O. C. 1881 . Classification of the Dinosauria . American Journal of Science, Series 3 , 23 : 241 – 244 .
- Marsh , O. C. 1884 . Principal characters of American Jurassic dinosaurs. Part VIII. The Order Theropoda . American Journal of Science, Series 3 , 27 : 329 – 340 .
- Meyer , H. von . 1861 . Archaeopteryx lithographica (Vogel-Feder) und Pterodactylus von Solnhofen . Neues Jarhbuch für Mineralogie, Geologie und Paläontologie , 1861 : 678 – 679 .
- Molnar , R. E. 1990 . “ Problematic Theropoda: “carnosaurs” . In The Dinosauria , Edited by: Weishampel , D. B. , Dodson , P. and Osmólska , H. 306 – 317 . Berkeley : University of California Press .
- Molnar , R. E. , Kurzanov , S. M. and Zhiming , D. 1990 . “ Carnosauria ” . In The Dinosauria , Edited by: Weishampel , D. B. , Dodson , P. and Osmólska , H. 169 – 209 . Berkeley : University of California Press .
- Naish , D. and Martill , D. M. 2007 . Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia . Journal of the Geological Society , 164 : 493 – 510 .
- Nesbitt , S. J. 2007 . The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa . Bulletin of the American Museum of Natural History , 302 : 1 – 84 .
- Nesbitt , S. J. and Norell , M. 2006 . Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda) . Proceedings of the Royal Society of London, Series B , 273 : 1045 – 1048 .
- Nesbitt , S. J. , Irmis , R. B. and Parker , W. G. 2007 . A critical re-evaluation of the Late Triassic dinosaur taxa of North America . Journal of Systematic Palaeontology , 5 : 209 – 243 .
- Newman , B. H. 1968 . The Jurassic dinosaur Scelidosaurus harrisoni Owen . Palaeontology , 11 : 40 – 43 .
- Newton , E. T. 1898 . On a megalosaurid jaw from the Rhaetic Beds near Bridgend (Glamorganshire) . Quarterly Journal of the Geological Society , 55 : 89 – 96 .
- Owen , R. 1842 . Report on British fossil reptiles. Part II . Reports of the British Association for the Advancement of Science , 1841 : 60 – 204 .
- Owen , R. 1861 . Monograph of the fossil Reptilia of the Liassic formations. Part I. A monograph of the fossil dinosaur (Scelidosaurus harrisonii Owen) of the Lower Lias . Palaeontolographical Society Monographs , 13 : 1 – 14 .
- Pérez-Moreno , B. P. , Sanz , J. L. , Sudre , J. and Sigé , B. 1993 . A theropod dinosaur from the Lower Cretaceous of southern France . Revue de Paléobiologie , 7 : 173 – 188 .
- Peyer , B. 1944 . Über Wirbeltierfunde aus dem Rhät von Hallau (Kt. Schaffhausen) . Eclogae Geologicae Helvetiae , 36 : 260 – 263 .
- Rauhut , O. W. M. 1997 . “ Zur Schädelanatomie von Shuvosaurus inexpectatus (Dinosauria; Theropoda) ” . In Treffen der deutschsprachigen Palaeoherpetologen , Edited by: Sachs , S. R. , Rauhut , O. W. M. and Weigert , A. 17 – 21 . Düsseldorf : Terra Nostra .
- Rauhut , O. W. M. 2003 . The interrelationships and evolution of basal theropod dinosaurs . Special Papers in Palaeontology , 69 : 1 – 213 .
- Rauhut , O. W. M. 2005 . Post-cranial remains of ‘coelurosaurs’ (Dinosauria, Theropoda) from the Late Jurassic of Tanzania . Geological Magazine , 142 : 97 – 107 .
- Rauhut , O. W. M. and Hungerbühler , A. 2000 . A review of European Triassic theropods . Gaia , 15 : 75 – 88 . [for 1998]
- Rauhut , O. W.M. and López-Arbarello , A. 2009 . Considerations on the age of the Tiouaren Formation (Iullemmeden Basin, Niger, Africa): Implications for Gondwanan Mesozoic terrestrial vertebrate faunas . Palaeogeography, Palaeoclimatology, Palaeoecology , 271 : 259 – 267 .
- Rauhut , O. W. M. and Milner , A. C. 2008 . Cranial anatomy and systematic position of the Middle Jurassic theropod dinosaur Proceratosaurus from England . Journal of Vertebrate Paleontology , 28 : 130A
- Rhinehart , L. F. , Lucas , S. G. and Hunt , A. P. 2007 . Furculae in the Late Triassic theropod dinosaur Coelophysis bauri . Paläontologische Zeitschrift , 81 : 174 – 180 .
- Rowe , T. 1989 . A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta Formation of Arizona . Journal of Vertebrate Paleontology , 9 : 125 – 136 .
- Russell , D. A. and Dong , Z.-M. 1993 . The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People's Republic of China . Canadian Journal of Earth Sciences , 30 : 2107 – 2127 .
- Sadleir , R. W. , Barrett , P. M. and Powell , H. P. 2008 . The anatomy and systematics of Eustreptospondylus oxoniensis, a theropod dinosaur from the Middle Jurassic of Oxfordshire, England . Palaeontographical Society Monographs , 160 : 1 – 82 . [for 2006]
- Sereno , P. C. , Wilson , J. A. and Conrad , J. L. 2004 . New dinosaurs link southern landmasses in the mid-Cretaceous . Proceedings of the Royal Society B , 271 : 1325 – 1330 .
- Sereno , P. C. 1999 . The evolution of dinosaurs . Science , 284 : 2137 – 2147 .
- Sereno , P. C. , Wilson , J. A. , Larsson , H. C. E. , Dutheil , D. E. and Sues , H.-D. 1994 . Early Cretaceous dinosaurs from the Sahara . Science , 266 : 451 – 476 .
- Sereno , P. C. , Dutheil , D. B. , Iarochene , M. , Larsson , H. C. E. , Lyon , G. H. , Magwene , P. M. , Sidor , C. A. , Varrichio , D. J. and Wilson , J. A. 1996 . Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation . Science , 272 : 986 – 991 .
- Smith , J. B. 2007 . Dental morphology and variation in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar . Society of Vertebrate Paleontology Memoir , 8 : 103 – 126 .
- Smith , N. D. , Makovicky , P. , Hammer , W. R. and Currie , P. J. 2007 . Osteology of Cryolophosaurus ellioti from the Early Jurassic of Antarctica and implications for early theropod evolution . Zoological Journal of the Linnean Society , 151 : 377 – 421 .
- Tartarinov , L. P. 1985 . New data on a collection of tetrapod teeth from the Rhaetian of Hallau (Switzerland) . Paleontological Journal , 1985 : 138 – 140 .
- Taquet , P. 1985 . “ Two new Jurassic specimens of coelurosaurs (Dinosauria) ” . In The beginnings of birds , Edited by: Hecht , M. K. , Ostrom , J. H. , Viohl , G. and Wellnhofer , P. 229 – 232 . Eichstätt : Freunde des Jura-Museums .
- Tykoski , R. S. and Rowe , T. 2004 . “ Ceratosauria ” . In The Dinosauria, , 2nd edition , Edited by: Weishampel , D. B. , Dodson , P. and Osmólska , H. 47 – 70 . Berkeley : University of California Press .
- Tykoski , R. S. , Forster , C. A. , Rowe , T. , Sampson , S. D. and Darlington , M. 2002 . A furcula in the coelophysoid theropod Syntarsus . Journal of Vertebrate Paleontology , 22 : 728 – 733 .
- Waldman , M. 1974 . Megalosaurids from the Bajocian (Middle Jurassic) of Dorset . Palaeontology , 17 : 325 – 340 .
- Walker , A. D. 1964 . Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs . Philosophical Transactions of the Royal Society, Series B , 244 : 53 – 134 .
- Weishampel , D. B. , Barrett , P. M. , Coria , R. A. , Le Loeuff , J. , Xu , X. , Zhao , X.-J. , Sahni , A. , Gomani , E. M. P. and Noto , C. R. 2004 . “ Dinosaur distribution ” . In The Dinosauria, , 2nd edition , Edited by: Weishampel , D. B. , Dodson , P. and Osmólska , H. 517 – 606 . Berkeley : University of California Press .
- Welles , S. P. 1984 . Dilophosaurus wetherilli (Dinosauria, Theropoda). Osteology and comparisons . Palaeontographica Abteilung A , 185 : 85 – 180 .
- Welles , S. P. and Long , R. A. 1974 . The tarsus of theropod dinosaurs . Annals of the South African Museum , 64 : 191 – 218 .
- Wilkinson , M. 1995 . Coping with abundant missing entries in phylogenetic inference using parsimony . Systematic Biology , 44 : 501 – 514 .
- Witmer , L. M. 2001 . “ The role of Protoavis in the debate on avian origins ” . In New Perspectives on the Origin and Early Evolution of Birds , Edited by: Gauthier , J. and Gall , L. F. 537 – 548 . New Haven : Peabody Museum of Natural History .
- Woodward , A. S. 1910 . On a skull of Megalosaurus from the Great Oolite of Minchinhampton (Gloucestershire) . Quarterly Journal of the Geological Society , 66 : 111 – 115 .
- Wu , X. –C. , Currie , P. J. , Dong , Z. , Pan , S. and Wang , T. 2009 . A new theropod dinosaur from the Middle Jurassic of Lufeng, Yunnan, China . Acta Geologica Sinica , 83 : 9 – 24 .
- Xu , X. , Zhao , X.-J. and Clark , J. M. 2001 . A new therizinosaur from the Lower Jurassic lower Lufeng Formation of Yunnan, China . Journal of Vertebrate Paleontology , 21 : 477 – 483 .
- Xu , X. , Clark , J. M. , Forster , C. A. , Norell , M. A. , Erickson , G. M. , Eberth , D. A. , Jia , C. and Zhao , Q. 2006 . A basal tyrannosauroid dinosaur from the Late Jurassic of China . Nature , 439 : 715 – 718 .
- Yates , A. M. 2006 . A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods . Palaeontologica Africana , 41 : 105 – 122 .
- Zhao , X.-J. and Currie , P. J. 1994 . A large crested theropod from the Jurassic of Xinjiang, People's Republic of China . Canadian Journal of Earth Sciences , 30 : 2027 – 2036 . [for 1993]