Abstract
The Upper Cretaceous Cantwell Formation in Denali National Park and Preserve (DENA), Alaska, contains an unparalleled fossil avian biodiversity. The Cantwell Formation, thousands of metres thick, was deposited near its current latitude and is exposed throughout much of DENA and elsewhere in the central Alaska Range. The Formation comprises a lower, dominantly fluvial sedimentary unit and an upper, mostly volcanic unit. Sedimentation of the lower unit was mainly in alluvial fan, braided and meandering stream, and lacustrine environments, with possible marginal-marine influence at times. Pollen data suggest that these sedimentary rocks are late Campanian or early Maastrichtian in age; thus the Cantwell Formation is correlative with other well-known dinosaur localities in Alaska. Bird tracks are preserved in multiple locations along a 40-km transect in DENA in fluvial and lacustrine deposits. Some bird tracks are found in association with dinosaur tracks and others are found on beds interbedded with dinosaur track-bearing layers. The approximate body sizes of the birds based on tracks show a range from sparrow- to heron-sized birds (∼25–30% larger than the modern Sandhill Crane Grus canadensis). The Cantwell Formation contains footprints assigned to several ichnotaxa found in either Asia or North America based on such morphological criteria as the presence or absence of a hallux, print size and shape, and angle of divarication: Aquatilavipes swiboldae, Ignotornis mcconnelli, Magnoavipes denaliensis sp. nov., Gruipeda vegrandiunus sp. nov. and Uhangrichnus chuni. The presence of a mixed Asian and North American ichnofauna suggests that at least some birds used Alaska as a bridge between Asia and North America. This diverse assemblage of avian traces, combined with the known fossil bone record and invertebrate trace fossil record, demonstrates that the northern Late Cretaceous polar region contained significant biodiversity.
Introduction
The study of fossil birds is thriving (e.g. Hou et al. Citation1995, Citation1996; CitationChiappe & Dyke 2002; CitationChiappe & Witmer 2002; CitationClarke 2004; CitationClarke et al. 2005; CitationFountaine et al. 2005; CitationChiappe & Dyke 2006; CitationLamanna et al. 2006), and it is clear that modern bird lineages extend into the Cretaceous (CitationClarke et al. 2005). The study of fossil bird tracks is also thriving (e.g. Lockley et al. Citation1992, Citation2003, Citation2006, Citation2009; CitationAzuma et al. 2002; CitationCoria et al. 2002; CitationLockley & Rainforth 2002; CitationKim et al. 2006; CitationZhang et al. 2006; CitationDe Valais & Merchon 2008; CitationAnfinson et al. 2009; CitationFalkingham et al. 2009), providing additional information on biodiversity as well as the opportunity for direct insight into ancient avian behaviour.
This report describes one of the most diverse records of fossil bird tracks found anywhere in the world. A variety of small- to large-sized tracks from the Upper Cretaceous Cantwell Formation in Denali National Park and Preserve (DENA), Alaska, are described and ichnotaxonomically assigned, and traces of feeding behaviour associated with the tracks are discussed.
Significant effort has been spent examining the vertebrate fossil record of northernmost Alaska (CitationBrouwers et al. 1987; CitationParrish et al. 1987; CitationNelms 1989; CitationClemens & Nelms 1993; CitationClemens 1994; CitationGangloff 1998; Fiorillo & Gangloff Citation2000, Citation2001; CitationGangloff et al. 2005; Fiorillo Citation2004, Citation2006, Citation2008a, Citation2008b). Upper Cretaceous rocks of this region contain the densest concentrations of fossil vertebrates of any high latitude location in the northern or southern hemisphere (CitationRich et al. 2002). Included within the fauna is a single avian taxon, Hersperornis (CitationBryant 1983). Late Cretaceous dinosaur remains also found in south-western Alaska demonstrate a widespread and rich continental polar ecosystem (CitationFiorillo & Parrish 2004).
Modern high latitude continental ecosystems have been extensively studied as a means of understanding the interconnectedness of biological, physical and human components within the Arctic and their implications for biodiversity and climate change studies. Alaska holds additional importance because it is at a geographical crossroad connecting two major landmasses, Asia and North America. This connection, referred to as Beringia or the Bering Land Bridge, is a route for faunal exchange extending back to the Cretaceous (CitationPaul 1988; CitationRussell 1993; CitationSereno 2000; CitationFiorillo 2008b). The new record of fossil bird tracks reported here within the boundaries of Beringia adds to our knowledge of biodiversity at this important time and place.
Geological background
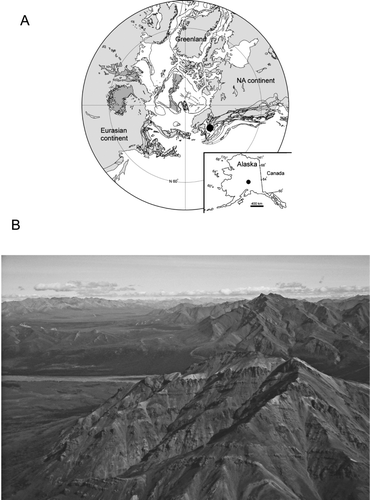
The Cantwell Formation is exposed throughout much of the central Alaska Range, which includes DENA (). This rock unit is thousands of metres thick and comprises an upper, dominantly volcanic unit and a lower, dominantly fluvial sedimentary unit (CitationKirschner 1994; CitationRidgway et al. 1997). The southern margin of Alaska was near its current latitude during the Cretaceous (CitationHillhouse & Coe 1994), as was the basin in which the Cantwell Formation was deposited (CitationRidgway et al. 1997; ).
Figure 1 A, Location of Denali National Park, Alaska, USA during the Cretaceous, shown by black dot. Inset shows current geographical location of study area. B, view to east of Cantwell Formation from Sable Mountain area.
The rocks of the lower Cantwell Formation include conglomerates, organic-rich and fossiliferous sandstones, siltstones, mudstones and coals (CitationRidgway et al. 1997; CitationFiorillo et al. 2009). They are well indurated. Finer grained units are typically organic rich and coarser units are laterally discontinuous, contained within thicker fine-grained sequences (CitationFiorillo et al. 2009). Sedimentation was dominated by alluvial fan, braided and meandering streams, and lacustrine depositional systems (CitationRidgway et al. 1997). Pollen analysis of the lower Cantwell Formation shows that these rocks were deposited during the late Campanian or early Maastrichtian (CitationRidgway et al. 1997).
Avian ichnotaxonomy
The morphological variation observed within modern tracks of known trackmakers can cross taxonomic groups (CitationRezendes 1999; CitationElbroch & Marks 2001). For fossil tracks the morphology of individual footprints can vary through the sediment layers beneath them (CitationManning 2004), within clearly defined trackways (CitationFarlow et al. 2006), and the track morphology of the same trackmaker can vary across media (CitationFiorillo 2005). Despite these caveats, however, careful observations of modern tracks allow for the correct identification and behaviour of the trackmaker. A large number of field guides are now available to assist modern trackers and hunters to identify a great variety of trackmakers and their behaviours (CitationMurie 1954; Halfpenny Citation1986, Citation1999, Citation2007; CitationForrest 1988; CitationRezendes 1999; CitationElbroch & Marks 2001; CitationElbroch 2003; CitationBrown et al. 2003). These guides attest that variation amongst tracks and trackways, if carefully considered, does have significance in understanding biodiversity.
The presence of webbing impressions is characteristic of some types of fossil bird tracks (CitationErickson 1967; CitationKim et al. 2006), but the absence of webbing makes identification of avian tracks more complicated. CitationLockley et al. (1992) defined fossil bird tracks as having: (1) similarity to modern bird tracks; (2) small size; (3) indistinct toe pad impressions; (4) wide divarication angles (110–120° or more) between digits II and IV; (5) a posteriorly directed hallux impression; (6) slender claw impressions; (7) curvature of the claw impressions of digits II and IV away from digit III; and (8) slender digit impressions. CitationDe Valais & Melchor (2008) reviewed these and additional characters for the identification of bird tracks in the fossil record. Of relevance to this paper is their reiteration of characters such as similarity to modern bird tracks and slender digit impressions.
Many recent descriptions of modern bird tracks (e.g. CitationRezendes 1999; CitationElbroch & Marks 2001; CitationBrown et al. 2003), however, have shown significant variance in tracks made by birds that extends beyond the original characteristics defined for ancient bird tracks (CitationLockley et al. 1992). Such variation renders criteria (1) and (2) of limited use for the purpose of identifying fossil bird tracks, though similarity to modern bird tracks is a qualitative character. Examination of some modern bird tracks, such as those made by the wild turkey (Meleagris gallopavo; see CitationElbroch & Marks 2001, p. 136) and particularly the bald eagle (Haliaeetus leucocephalus; see CitationElbroch & Marks 2001, p. 99), show very distinct toe pad impressions, thus the presence or absence of toe pads is not a valid means of identifying bird tracks in the fossil record. Similarly, examination of the divarication angles of some egret tracks (e.g. Reddish Egret Egretta rufescens and Great Egret Ardea alba; see CitationElbroch & Marks 2001, pp. 95 and 97, respectively) and Bald Eagle tracks (Haliaeetus leucocephalus; see CitationElbroch & Marks 2001, p. 99) shows divarication angles significantly narrower than 110°. In addition, bird tracks such as those of the Piping Plovers (Charadrius melodus) and Sanderlings (Calidris alba) do not show digit I impressions (CitationElbroch & Marks 2001, pp. 102 and 106, respectively). Finally, many bird tracks do not exhibit claw marks (CitationRezendes 1999; CitationElbroch & Marks 2001). Thus comparison of the criteria for identifying fossil bird tracks to well-illustrated varieties of modern bird tracks demonstrates that care needs to be taken when identifying avian versus non-avian footprints. Identification of fossil bird tracks is, therefore, based primarily on the presence of slender digit impressions and the somewhat subjective birdlike morphology of a track.
Tracks typically belong to one of five categories for extant birds (CitationRezendes 1999; CitationElbroch & Marks 2001). Anisodactyl tracks have a digit arrangement in which three toes (digits II–IV) point forward and one toe (digit I) points backward. Incumbent anisodactyl tracks have digits II–IV pointing forward and a greatly reduced or non-existent impression left by digit I, meaning this digit is greatly reduced or non-existent. Palmate tracks have full webbing between digits II–IV and the impression left by digit I is greatly reduced or non-existent; these resemble incumbent anisodactyl tracks. Palmate tracks also differ from incumbent anisodactyl tracks in that digits II and IV are somewhat curved medially.
Table 1 Summary of track data used in this report. Generic names only are provided in table; genus and species names can be found in the text. Average divarication angle provided in parentheses.
CitationRezendes (1999) and Elbroch & Marks (2001, pp. 20–22) recognized that morphological variation within modern tracks of known trackmakers caused by variability in sediment texture, consistency and moisture can produce tracks that belong to more than one morphological group. We have observed the same phenomena with the DENA bird tracks, even within the same trackway. The interaction of Cretaceous bird feet with a locally variable sediment surface produced similar variability in the morphology of the DENA bird tracks, that must be identified before ichnotaxonomic assessments are made.
Methods
Several hundred avian footprints have been identified from four areas in DENA located along a 40 km east–west transect. The exact localities of sites are on file with DENA and at the Museum of Nature and Science (DMNH), Dallas, Texas, but the general locations within DENA are, from east to west, Fang Mountain, Double Mountain, Tattler Creek and Cabin Peak. Most specimens were recorded either by direct collection or by silicone moulding of tracks which are still in the field. Epoxy casts of the specimens were made from these moulds. These casts are now housed at the DMNH.
Morphological characteristics were used to describe the DENA tracks, organized according to the bird track categories defined by CitationElbroch & Marks (2001). Length and width were measured for each track and an average size was determined for tracks that comprised a trackway. Divarication angles were measured between toes II–III, III–IV and II–IV; an average divarication angle between toes II–IV was determined for tracks that comprised a single trackway (). Where available, stride (the distance between successive left or right tracks), trackway width or straddle (the distance perpendicular to the median line of the trackway made by tracks), and the angle of tracks with the median line, or pace angulation, were also measured. The DENA tracks were compared to descriptions and illustrations of previously described bird tracks (e.g. CitationMehl 1931; CitationEllenberger 1972; CitationCurrie 1981; CitationLockley et al. 1992; CitationYang et al. 1995; CitationLee 1997) to determine ichnotaxonomic affinities.
Three-dimensional data acquisition was accomplished using a NextEngine™ HD Desktop 3D scanner and ScanStudio™ HD PRO software (www.nextengine.com) with a resolution of 0.127 mm. Multiple scan angles were required to capture all internal track surfaces and minimize occluded surfaces. Scans were aligned within ScanStudio™ to merge them into a single three-dimensional model. Contour lines were generated by the ScanStudio™ automated spline extractor function, which slices through the three-dimensional model in a set of parallel planes at intervals of 0.25 to 1.27 mm. The final 3D model was exported in Open Inventor™ (www.sgi.com) format that includes mesh, material, and texture files (*.obj, *.mtl, *.jpg). LightWave 3D® version 9.3 software (www.newtek.com/lightwave/) was used to render the three-dimensional object into common media file formats.
Institutional abbreviation
DMNH: Museum of Nature and Science, Dallas, Texas, USA.
Avian tracks
Anisodactyl tracks
Ignotornis mcconnelli CitationMehl, 1931; emended CitationKim et al., 2006
()
Figure 2 Ignotornis mcconnelli from Double Mountain (DENA 13342) with additional tracks attributable to Aquatilavipes swiboldae. The presence of a very small hallux impression on some of these other tracks is suggestive that these two ichnotaxa may be very similar.

Diagnosis
Moderate-sized, four-toed tracks with divarication angles between digits II and IV equal to or greater than 90°. Digit I is directed posteriorly. CitationKim et al. (2006) emended this diagnosis to include the presence of webbing impressions. CitationLockley et al. (2009) noted that the webbing impressions are more pronounced between digits III and IV than between digits II and III.
Locality
Double Mountain, Denali National Park, Alaska.
Description
Long, slender digits. Individual tracks consist of impressions of digits I–IV with the hallux of each track well pronounced; metatarsal pads usually obscured as a result of tracks being made in deep mud (). Tracks tend to be nearly as wide as they are long, averaging ∼7.1 cm (6.4–7.6 cm) long and ∼7.6 cm (6.9–8.1 cm) wide. Length of digit I variable but approaching length of digit II. Divarication angles range from 103° to 174° for this track morphotype. Digit I nearly in line with digit III.
Remarks
These fossil bird tracks, like some modern bird tracks, show an anisodactyl track morphology of digit arrangement where three toes point forward and one toe points backward (sensu CitationElbroch and Marks 2001). Their characteristics match those that define Ignotornis CitationMehl, 1931, with the exception of that the Cantwell tracks have a divarication angle up to 174°. The range of divarication angles for Ignotornis (CitationMehl 1931; CitationLockley et al. 2009) is up to 140° in the Dakota Sandstone. CitationMehl (1931) also mentioned that the depth of the Dakota tracks reached 10 mm into the medium; the Cantwell tracks only penetrate as much as 5 mm. For Triassic theropod tracks of similar morphology, CitationGatesy et al. (1999) showed convergence of digits II and IV with depth. We attribute this additional range in divarication angle to the differences in the medium, leading to penetration of the track into the subsurface.
These tracks, in a very fine-grained mud substrate, were likely made by a bird approximately 34–37 cm long based on comparisons of foot size with extant bird specimens in the DMNH ornithological collections. The Late Cretaceous bird responsible for these DENA tracks was about the length of a medium-sized wading bird, such as a modern Willet (Catoptrophorus semipalmatus).
CitationMehl (1931) mentioned the questionable presence of a short web connecting the second, third and fourth digits, and possibly also the first digit. Contrary to CitationMehl (1931), in their review of the type material for this ichnogenus CitationLockley et al. (2009) saw no evidence of webbing between digits I and II. CitationFalkingham et al. (2009) suggested that the webbed tracks may be preservational rather than morphological features, a perspective not accepted by CitationLockley et al. (2009) or ourselves in the case of the Cantwell tracks.
The webbing impression discussed by these authors is also present in the tracks described here. Elbroch and Marks (2001, p. 22) illustrated similar features in the tracks of Spotted Sandpipers (Actitus macularia), where they have walked in deeper mud. These features are not present, however, in the tracks of birds that walked over shallower mud. Thus we interpret the Ignotornis tracks from Double Mountain as having been made in a medium that was soft and similar to the deeper mud illustrated for tracks made by Spotted Sandpipers (CitationElbroch & Marks 2001).
Incumbent anisodactyl tracks
Aquatilavipes swiboldae CitationCurrie, 1981
()
Figure 3 Aquatilavipes swiboldae from Double Mountain (DENA 20939).

Figure 4 Magnoavipes denaliensis from Tattler Creek. Original tracks remain in outcrop. Specimen moulded and cast (DMNH 2008-08-22). Contour interval = 1.27 mm.

Diagnosis
Three slender toe impressions, with individual pes impressions wider than long. No hallux (Digit I) impression. Divarication angle between digits II and IV greater than 100°. Digit III is the longest and digit IV is longer than digit II. Individual tracks are generally less than 4.5 cm long with a width approximately 26% greater than the length.
Localities
Fang Mountain, Double Mountain, Denali National Park, Alaska.
Description
Tracks are shown in . Length and width measurements were taken for more than 30 tracks. Individual tracks are ∼4.9 cm long (4.3–5.5 cm) and ∼6.7 cm wide (6.2–7.3 cm) length:width ratio being 0.72. Digit III ∼35% longer than digits II and IV. Average divarication angle is 142°, with a range of 103–174° ().
Remarks
Like modern incumbent anisodactyl tracks (sensu CitationElbroch and Marks 2001), these fossil bird footprints have three toes that point forward (digits II–IV) and a non-existent impression left by digit I. The morphology of these fossil tracks compares well with the ichnospecies Aquatilavipes swiboldae CitationCurrie, 1981. However, the DENA tracks differ from A. swiboldae in being ∼25% larger. CitationCurrie (1981) reported a range in length for A. swiboldae from the Lower Cretaceous Gething Formation of British Columbia of ∼20–44 mm. This range exceeds the difference between the length of the holotype for the ichnotaxon and the lengths observed in the tracks described here. We, therefore, attribute the tracks from the Cantwell Formation to A. swiboldae.
Magnoavipes CitationLee, 1997
(Figs –)
Figure 5 Magnoavipes denaliensis, Close up of Fig. 4, showing toe pads.
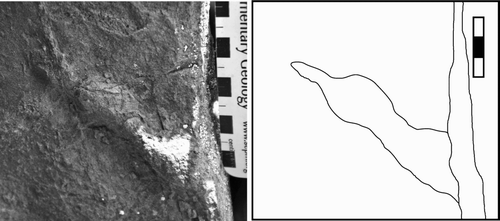
Figure 6 Magnoavipes denaliensis. Close up of resin cast DMNH 2008-08-22 digitally scanned and contoured to show toe pads. The tracks represented in this figure correspond to the tracks in Fig. 4. Longitudinal section of one digit impression showing the elevation changes along the digit that correspond to toe pad impressions.

Figure 7 Magnoavipes denaliensis. Resin cast (DMNH 2008-08-22) of tracks shown in Figs. 4 and 5, digitally scanned and contoured to show toe pads. Oval encompasses casts of tracks shown in Figs. 4 and 6. Rounded rectangle encompasses area of cast shown in Fig. 5; note the presence of prominent toe pads. Contour interval = 1.27 mm.

Figure 8 Magnoavipes isp. (DENA 20950).

Remarks
CitationLee (1997) described Magnoavipes as the tracks of a large bird from the Upper Cretaceous (Cenomanian) Woodbine Formation of Texas. The Woodbine Formation tracks are approximately the same size as the tracks found in the Cantwell Formation but the latter have a broader divarication angle. CitationLockley & Rainforth (2002) and CitationLockley et al. (2001) suggested that Magnoavipes may not be a valid avian ichnotaxon because of the divarication angle <110° exhibited on morphologically similar tracks found elsewhere in North America. The range of divarication angles among modern birds, however, is commonly <110° (CitationElbroch & Marks 2001).
Furthermore, CitationLockley et al. (2001) illustrated several examples of tracks attributed to the ichnotaxon M. caneeri as part of their discussion regarding the questionable attribution of Magnoavipes to Aves. Many of their line drawings have more in common with examples of the tridactyl undertracks illustrated by CitationManning (2004) than with true surface tracks. In addition, other tracks attributed to M. caneeri clearly have far more robust toes and much narrower divarication angles than the Cantwell tracks. Comparison of our tracks (Figs –) to the line drawings provided in these other papers discussing Magnoavipes yields little insight. We therefore accept the interpretation of this ichnotaxon being avian in origin based on the original discussion of divarication angles by CitationLee (1997) in comparison with modern birds.
Magnoavipes denaliensis isp. nov.
(Figs –)
Etymology
From the Koyukon Athabaskan word ‘Denali’, meaning ‘high one’.
Diagnosis
Long, slender birdlike toe impressions (Figs –). Very large footprint, as long as wide. Toe pads prominent. Sharp claw impressions on all digits.
Type
DMNH 2008–08-22 which is a resin cast of tracks still in the field.
Locality
Tattler Creek, Denali National Park, Alaska.
Description
The divarication angle range and length-width measurements of these DENA tracks place these specimens within the ichnogenus Magnoavipes CitationLee, 1997. Tracks exhibit a range of divarication angles from 97 to 116°. Average track length is 20 cm (19.5–20.5 cm) and average width is also 20 cm (19.5–20.5 cm). Digits II and IV are ∼15 cm and 13 cm, respectively.
Remarks
Whereas Magnoavipes lowei CitationLee, 1997 is of similar size, examination of the type specimen of this ichnospecies (DMNH 918) shows that toe pads are lacking. In contrast, the DENA tracks have pronounced, robust toe pads (Figs , ). Tracks attributed to Magnoavipes caneeri have far more robust toes and much narrower divarication angles (CitationLockley et al. 2001) than M. denaliensis.
The length–width measurements of these tracks are large compared to other fossil bird footprints and it might at first seem problematical that there are no body fossils of birds large enough to have made these tracks. However, the measurements are only 25–30% larger than tracks potentially made by Sandhill Cranes (Grus canadensis). Corresponding length–width measurements from two extant specimens in the holdings at the Museum of Nature and Science provided an average length of the foot of almost 13 cm and an average width of 15 cm. Extant birds such as the Cassowary (Casuarius) indicates birds can achieve even larger sizes. We, therefore, postulate that these tracks reasonably fall within the potential size range of avian tracemakers.
CitationLee (1997) interpreted the tracks of Magnoavipes lowei as being those of a crane-like bird. Although many factors (e.g. preservation, kinematics, trackmaker) may result in the presence or absence of toe pads, the pronounced toe pads in M. denaliensis, and lack thereof in M. lowei, may indicate differences in behaviour between the two types of animal. For example, as Elbroch & Marks (2001, p. 16) pointed out, examination of the tracks of a Great Blue Heron (Ardea herodias) and Bald Eagle (Haliaeetus leucocephalus) reveal differences in overall footprint morphology but similarity in length and width measurements. The pronounced toe pads of Bald Eagle tracks are indicative of the grasping use of the feet of this bird for holding prey, as compared to the slender, less padded toes of the Great Blue Heron that walks through mud flats.
The presence of toe pads in the fossil tracks may be a function of taphonomic processes rather than anatomy. For example, Brown et al. (2003, p. 74) illustrated variation in the tracks of Capercaillie (Tetrao urogallus), presumably related to variation in substrate, such as grain size or water content. The range in morphology of the toes illustrated for a Capercaillie is encompassed by both M. lowei and M. denaliensis. While we cannot measure the water content in the substrates that these taxa walked across, the lithology of the rock containing the Cantwell tracks, a fine-grained sandstone, is very similar to the lithology of the rock containing the type specimen in Texas. This suggests that these differences in morphology are probably real.
Magnoavipes isp.
()
Description
Long, slender, birdlike toe impressions () with a divarication angle of 110°. Track length is ∼19.5 cm and width ∼22.5 cm. Digit III is the longest, digit IV is ∼17 cm and digit II is ∼15 cm. Claw impressions not well defined. The track is a raised impression of a right foot, the result of relief of the track caused by erosion.
Remarks
This track, which is isolated on a block (DENA 20950), is preserved in a fine-grained grey sandstone. Though from a different locality, this lithology is similar to that containing the tracks of M. denaliensis. The absence of toe pads on the track suggests that there are two morphotypes of this ichnogenus preserved within the Cantwell Formation.
Gruipeda CitationPanin & Avram, 1962; emend. CitationDe Valais & Melchor, 2008
Gruipeda vegrandiunus isp. nov.
()
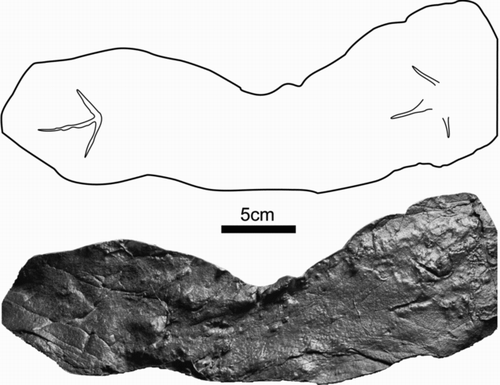
Figure 9 Resin cast of Gruipeda vegrandiunus (DMNH 18352) from Cabin Peak, DENA.
Type
DMNH 18352 which is a resin cast made from the original still in the field.
Etymology
Vegrandiunus from a Latin combination for small one, vegrandis unus.
Diagnosis
Small tracks that are ∼3.5 cm long and are ∼4 cm wide (). The length:width ratio for tracks is ∼0.88. Divarication angles consistently 108°–109°. Pad impressions are indistinct. Full stride measurements are unavailable. Half stride distance is 22 cm. The orientation of digit III effectively parallels median line.
Table 2 Comparison of average track sizes between Gruipeda vegrandiunus and Gruipeda moabensis. Averages for G. moabensis are from data provided by CitationLockley et al. (1992). Measurements in centimetres. N = 2 for G. vegrandiunus, and N = 4 for G. moabensis.
Figure 10 Uhangrichnus chuni from Cabin Peak. Original track remains in outcrop. Specimen moulded and cast (DMNH 2008-08-06). The curvature between digits II and III, and digits III and IV are interpreted as webbing, a characteristic defining this ichnogenus.

Locality
Cabin Peak, Denali National Park, Alaska.
Description
These are very small tracks (; ). The divarication angle is determined by extending the imprints of digits II and IV until their traces intersect to form a bisecting angle. Pad impressions and claw impressions are indistinct. The tracks are very small compared to other fossil bird tracks. Traces of webbing are present on one track ().
Remarks
CitationDe Valais and Melchor (2008) recently synonymized the ichnogenus Trisauropodiscus (CitationEllenberger 1972; CitationLockley et al. 1992) with Gruipeda based on a marked similarity between the two ichnotaxa. The authors do note in their revision that some ichnospecies of Trisauropodiscus such as T. superaviforma may warrant a different ichnogeneric designation.
The main differences between the Cantwell Formation tracks and those previously described for Gruipeda are that these tracks are smaller and much younger in geologic age. These tracks, therefore, are distinguished only at the ichnospecies level.
The size range of this track falls within the overall size range of tracks described for this ichnogenus (CitationEllenberger 1972; CitationLockley et al. 1992) but the Gruipeda vegrandiunus tracks are much smaller than the other North American form G. moabensis (CitationLockley et al. 1992; ). Whereas some of the tracks described by CitationEllenberger (1972) included an impression of digit I, not all tracks attributed to this ichnogenus exhibited that particular impression. Similarly, the larger North American ichnospecies T. moabensis (CitationLockley et al., 1992) lacks the hallux impression, as do the tracks described here.
Also, there is significant variation in the divarication angles of tracks attributed to this ichnotaxon. The Moroccan forms of this ichnotaxon, which are under revision (Belvedere, pers. comm.), have divarication angles of 90°, whereas the North American forms have divarication angles of 110°. The divarication angles of the tracks from the Cantwell Formation are very close to the average angle for those tracks described from the Navajo Sandstone of North America (CitationLockley et al. 1992) but clearly significantly younger in geologic age.
The orientation of the tracks to the median line of a trackway in G. vegrandiunus is virtually parallel. Tracks for most ichnospecies of Gruipeda also show a similar pattern (CitationLockley et al. 1992). The most notable exception is the larger tracks of G. moabensis, which shows tracks divergent from the median line. The trackways illustrated by CitationDe Valais and Melchor (2008) for Gruipeda dominguensis are ambiguous with respect to the angle of the tracks to the median line of the trackway, but some tracks suggest convergence towards the median line.
Though previous records for this ichnotaxon are from the Jurassic, CitationLockley et al. (1992) point out that the tracks of Gruipeda are virtually indistinguishable from tracks made by post-Jurassic inferred water birds. This new ichnospecies establishes the presence of the ichnogenus Gruipeda in the Late Cretaceous of North America.
Palmate tracks
Uhangrichnus CitationYang et al., 1995
(Figs –)
Figure 11 Uhangrichnus chuni. Resin cast (DMNH 2008-08-06) digitally scanned and contoured. Contour interval = 1.27 mm.
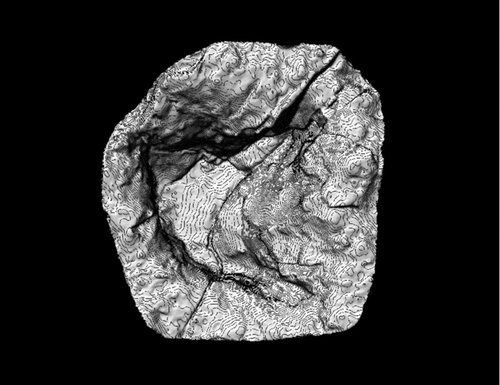
Figure 12 Uhangrichnus isp. Large track attributable to this ichnogenus still in outcrop.

Diagnosis
As for the type.
Uhangrichnus chuni CitationYang et al., 1995
Uhangrichnus isp. CitationYang et al., 1995
Diagnosis
Tridactyl tracks showing webbing between digits. Digits II and IV curve medially.
Locality
Cabin Peak, Denali National Park, Alaska.
Description
Uhangrichnus tracks are defined as tridactyl, showing clear webbing between digits (CitationYang et al. 1995). and show one example of a relatively small track attributable to this ichnotaxon, whereas shows a single larger track found at the site. The hallux is absent in all examples. Digits II and IV in all examples show slight medial curvature. Divarication angles range from 135° to 173° for the two smaller tracks, whereas a divarication angle of 116° is present in the larger track.
Remarks
These tracks (sensu CitationElbroch & Marks 2001) resemble incumbent anisodactyl tracks, in that the impression left by digit I is greatly reduced or non-existent. They differ from regular incumbent anisodactyl tracks in that digits II and IV are somewhat curved medially. Modern birds that exhibit foot structure that produce this type of track have webbed feet (CitationElbroch & Marks 2001).
Two examples of these tracks have been found at the Cabin Peak locality, a smaller set of tracks and one much larger track. Though the smaller tracks are somewhat larger in width and length () than the range reported by CitationYang et al. (1995), as well as having a broader total divarication angle than is defined by CitationYang et al. (1995), these tracks are attributed to the ichnotaxon Uhangrichnus chuni. Given that the larger track is well beyond the size range of described variations for the ichnogenus and only one track was found, we refer this single track to Uhangrichnus isp.
Feeding traces
Several dozen near-circular to suboval depressions are associated with the Ignotornis and Aquatilavipes tracks at Double Mountain (). These depressions are ∼3.6 mm and ∼4.3 mm along their respective semi-minor and semi-major axes of symmetry. Many surface expressions of these depressions contain very thin folds, ∼0.1–0.5 mm wide and up to one-third to one-half as long as the opening is wide, that radiate outward. Others are puncture-like, short, sharply tapering conical features. In some cases the oval features are closely spaced (2–5 mm), nearly touching the closest areas being flattened; or they appear connected and form a short dumbbell-shaped figure-of-eight. Although no cross-sections are available for study of the depressions, they appear to be shallow (<5 mm), as they do not penetrate the bedset (∼1 cm) in which they are contained. In some cases the depressions are infilled with slightly coarser matrix.
Figure 13 A, DENA 20915 showing association of bird tracks and bill-probe marks. Small ripple marks indicate the presence of shallow water during the formation of this bedding plane. B, Close up photo of bill-probe marks.

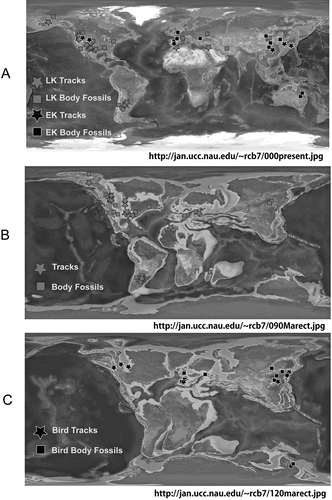
Figure 14 Map showing distribution of known Cretaceous bird localities. Base maps from Dr Ron Blakey, Northern Arizona University, http://jan.ucc.nau.edu/~rcb7. Data from CitationLockley et al. (1992). A, Cretaceous bird body fossils and tracks; B, Late Cretaceous bird body fossils and tracks; C, Early Cretaceous bird body fossils and tracks.
The morphology of the depressions, their association with bird tracks, and comparison with modern traces suggests that they are shallow punctures produced by the narrow bill of a bird. Such structures produced by extant birds while exploring the sediment with their beaks for invertebrates are referred to as probe marks (e.g. CitationLane & Davies 1987; CitationElbroch & Marks 2001). A closed bird bill used for probing the sediment probably produced the open near-circular and strongly tapering conical traces. A slightly opened bird bill used for probing the sediment probably produced the closely spaced, slightly flattened depressions and the figure-of-eight impressions. The associated depressions preserve the individual shapes of the upper and lower bill of the trace-making bird. In her re-examination of two large slabs of bird tracks (KS001 and KS064 of Lim et al. 2000) from the Lower Haman Formation of South Korea, CitationFalk (2009) interpreted similar features to those discussed here as bird bill probing traces associated with web-footed bird tracks. The web-footed bird tracks were originally recognized and described by Lim et al. (2000).
Structures interpreted as bird feeding traces differ from paired opening of U-shaped burrows (i.e. Arenicolites) and closely spaced individual burrows (e.g. Skolithos) in that these trace fossils have well-defined, sharp circular openings similar to like traces in other Mesozoic and Cenozoic fluvial-lacustrine deposits (e.g. Hasiotis Citation2002, Citation2004). These burrow morphologies are penetrative from several centimetres for U-shaped burrows to over 0.5 m for narrow diameter (≤3 mm) vertical burrows.
The DENA features compare well with probe marks produced by modern members of the Charadriiformes (e.g. American Woodcock, Scolopax minor, CitationElbroch & Marks 2001). Charadriiformes have been recognized from the Cretaceous (CitationOlson & Parris 1987) and long bills for probing sediment are largely restricted to the Charadriiformes. The sizes of these depressions compare well with the bill sizes of extant birds of this group, with similar foot sizes to the tracks preserved in the Cantwell Formation. Given the general uniqueness of the bill morphology of Charadriiformes, we suggest that these traces are consistent with the trackmaker being a member of this modern avian group.
Birds probe the sediment today in search of a variety of invertebrates (CitationGill 2007), though their diet can include items other than invertebrates. The Sanderling (Calidris alba), for example, consumes seeds, plant buds, algae and mosses in addition to a variety of invertebrates (CitationRichards 1988). Invertebrates often leave evidence of their presence in the form of burrows. Definitive subhorizontal tubular invertebrate trace fossils are found commonly in the section containing the Ignotornis and Aquatilavipes footprints, confirming the existence of an invertebrate food source for the birds that made these tracks.
CitationErickson (1967), in his study of Eocene bird tracks, used the term dabble pattern to describe a nearly continuous series of shallow depressions that wove around a trackway of webbed footprints. He described the marks as “a series of asymmetrically connected S-shaped curves along the route of progression”. He inferred that the Eocene web-footed bird moved its bill in a side-to-side motion, feeding along the way, much as modern members of the Anseriiformes (ducks and geese) feed.
Comparison of Erickson's figured tracks with the ones described here shows two significant points of dissimilarity. Firstly, Erickson's trackway dabble marks are clearly connected as part of a progression, whereas the circular traces described here are isolated. And secondly, the Cantwell Formation traces puncture through the bedding plane surface, as evidenced by their being infilled with coarser sandy matrix; there is no indication that the traces figured by Erickson do likewise.
The Cantwell Formation feeding traces appear similar to those illustrated by CitationGenise et al. (2009, fig. 11), associated with Lower Jurassic birdlike tracks described from the Santo Domingo Formation in Argentina. Detailed descriptions and photographs, however, are lacking from their report and cannot be used with any confidence to clearly differentiate bird probing traces from paired burrow openings or closely spaced single burrow openings—ancient or extant.
These Cantwell Formation traces, therefore, constitute a second style of avian feeding behaviour from the fossil record, the first being that defined by CitationErickson (1967). Accepting the DENA traces as evidence of probe-style feeding behaviour in Cretaceous members of the Charadriiformes shows that this specialized feeding behaviour evolved early in this taxonomic group.
Discussion
Field research in the lower Cantwell Formation in DENA has produced seven avian ichnotaxa that represent three different foot morphotypes: (1) Ignotornis mcconnelli and Aquatilavipes swiboldae (anisodactyl); (2) Magnoavipes denaliensis, Magnoavipes isp. and Gruipeda vegrandiunus, (Incumbent anisodactyl); and (3) Uhangrichnus chuni and Uhangrichnus isp. (Palmate). Associated with the anisodactyl tracks are numerous feeding traces comprising thin, circular impressions produced by shallow penetration into the sediment by the bills of birds. The track and feeding trace fossils demonstrate that DENA has one of the most diverse ichnological records for birds in the world. Equally important is the fact that this great diversity occurs in a high-latitude, polar continental ecosystem.
These tracks do not compare well with footprints that might have been made by the only previously known Cretaceous bird, Hesperornis from Alaska (CitationBryant 1983). The clear arrangement of the toes in these DENA tracks differs from the foot structure of Hesperornis (CitationMarsh 1880) which would have had a totipalmate foot morphology in which all of the toes were webbed (sensu CitationElbroch & Marks 2001).
shows the distribution of fossil birds from the Cretaceous and the global distribution of Cretaceous bird tracks. A report of an isolated bone attributable to Hesperornis has been found elsewhere in Upper Cretaceous rocks of Alaska (e.g. CitationBryant 1983).
Beringia, a land bridge connection centred on Alaska and connecting North America and Asia, was present intermittently throughout the Cenozoic and probably explains the exchanges of a broader suite of terrestrial organisms between these two modern land masses (CitationFlerow 1967; Hopkins Citation1967, Citation1996; CitationColinvaux 1980; CitationCwynar & Ritchie 1980; Guthrie Citation1982, Citation2001; CitationHopkins et al. 1982; Repenning Citation1987, Citation1990; CitationRepenning et al. 1987; CitationWest 1996; CitationAbbott & Brochman 2003; CitationO’Neill 2004). The concept of Beringia has been extended back into the Cretaceous (CitationPaul 1988; CitationRussell 1993; CitationSereno 2000; CitationFiorillo 2008b), supported by the striking similarity in dinosaurian faunas from Asia and North America at very broad taxonomic levels. For example, within the Tyrannosauridae in the latest Cretaceous, Tyrannosaurus is present in North America, while Tarbosaurus is present in Asia. Similarly the Asian small theropod Velociraptor and the North American theropod Saurornitholestes show a great deal of morphological similarity (CitationPaul 1988).
The fossil bird tracks described here offer further insight into the ecological significance of the Beringia land bridge. Irving (Citation1960, Citation1972) showed that the migratory origins of modern Alaskan nesting birds include both Asia and North America. There is little surprise then that the Cantwell Formation records an assemblage of bird tracks previously described in Asia, North America, or both. Indeed other non-Alaskan examples exist of avian ichnotaxa that have been found in both Asia and North America (CitationLockley & Rainforth 2002; CitationKim et al. 2006; Anfinson et al. 2009). This overlap of Asian and North American ichnotaxa is evidence that the avian fauna of Alaska likely shared traits of both continents.
Conclusions
The avian trace fossil record within the Upper Cretaceous lower Cantwell Formation in the central part of the Alaska Range, Alaska, is extraordinarily diverse and represents one of the most diverse record for birds found anywhere in the world. This rock unit, thousands of metres thick in DENA, was deposited near its current latitude, making these tracks the most polar fossil bird tracks known to date. The record consists of several hundred bird tracks and several dozen bill probe marks resulting from feeding activity.
Bird tracks are preserved in multiple locations along a 40-km transect in DENA in fluvial and lacustrine deposits; they are not preserved as a single-track assemblage from one bedding plane. Some bird tracks are found in association with dinosaur tracks and others are found on beds interbedded with dinosaur-bearing layers. The approximate body sizes of the birds based on tracks ranges from sparrow-sized to birds ∼25–30% larger than a modern Great Blue Heron (Ardea herodias).
The presence of a mixed Asian and North American ichnofauna suggests that at least some birds used Alaska as a bridge between Asia and North America. This diverse assemblage of avian traces, combined with the known fossil bone record and invertebrate trace-fossil record, demonstrates that the northern Late Cretaceous polar region contained significant biodiversity.
Acknowledgements
We thank the National Park Service Alaska System Support Office and Denali National Park for their logistical and financial support for this project, particularly Russell Kucinski, Linda Stromquist and Phil Brease. We thank Erik Brandlen, Susi Tomsich and Kevin Hobbs for assistance in the field, Dr Ronald Tykoski and Karen Morton for assisting with the preparation of study materials, Dr Gareth Dyke for opinions on the traces described, and Drs Kenneth Ridgway and Jeff Trop for sharing their experiences of the Cantwell Formation. Thanks also go to Thomas Adams and Mike Polcyn and the Visualization Lab in the Huffington Department of Earth Sciences at Southern Methodist University, for 3D scanning and rendering of the tracks used in the figures for this paper, as well as taking the time to write up the specific methodology for inclusion here. Dr Lawrence Lawver and Lisa Gahagan and the PLATES Project University of Texas Institute of Geophysics kindly provided the base map for , Dr Ron Blakey similarly provided the base maps for Fig. 14 and Andrew Epps undertook photographic work for Figs. and . We also thank Drs Yuong-Nam Lee and Matteo Belvedere for their respective insightful reviews of an earlier version of this manuscript, and Drs Gareth Dyke and Larry Martin for their invitation to participate in this symposium. ARF gratefully acknowledges the Museum of Nature and Science, American Airlines and Whole Earth Provision Company for support in the field. This work is dedicated to the memory of Phil Brease.
Related Research Data
References
- Abbott , R. J. and Brochmann , C. 2003 . History and evolution of the arctic flora: in the footsteps of Eric Hultén . Molecular Ecology , 12 : 299 – 313 .
- Anfinson , O. A. , Lockley , M. G. , Kim , S. H. , Kim , K. S. and Kim , J. Y. 2009 . First report of the small bird track Koreanaornis from the Cretaceous of North America: implication for avian ichnotaxonomy and paleoecology . Cretaceous Research , 30 : 885 – 894 .
- Azuma , Y. , Arakawa , Y. , Tomida , Y. and Currie , P. J. 2002 . Early Cretaceous bird tracks from the Tetori Group, Fukui Prefecture, Japan . Memoir of the Fukui Prefectural Dinosaur Museum , 1 : 1 – 6 .
- Brouwers , E. M. , Clemens , W. A. , Spicer , R. A. , Ager , T. A. , Carter , L. D. and Sliter , W. V. 1987 . Dinosaurs on the North Slope, Alaska: High latitude, latest Cretaceous environments . Science , 237 : 1608 – 1610 .
- Brown , R. , Ferguson , J. , Lawrence , M. and Lee , D. 2003 . Tracks & signs of the birds of Britain & Europe , 2nd edition , 333 London : Christopher Helm .
- Bryant , L. J. 1983 . Hesperornis in Alaska . PaleoBios , 40 : 1 – 8 .
- Chiappe , L. M. and Dyke , G. J. 2002 . The Mesozoic radiation of birds . Annual Review of Ecology and Systematics , 33 : 91 – 124 .
- Chiappe , L. M. and Dyke , G. J. 2006 . The early evolutionary history of birds . Journal of the Paleontological Society of Korea , 22 : 133 – 151 .
- Chiappe , L. M. and Witmer , L. M. 2002 . Mesozoic Birds: above the heads of dinosaurs , 532 Berkeley : University of California Press .
- Clarke , J. A. 2004 . Morphology, phylogenetic taxonomy, and systematics of Ichthyornis and Apatornis (Avialae: Ornithurae) . Bulletin of the American Museum of Natural History , 286 : 1 – 179 .
- Clarke , J. A. , Tambussi , C. P. , Noriega , J. I. , Erickson , G. M. and Ketcham , R. A. 2005 . Definitive fossil evidence for the extant avian radiation in the Cretaceous . Nature , 433 : 305 – 308 .
- Clemens , W. A. 1994 . “ Continental vertebrates from the Late Cretaceous of the North Slope, Alaska ” . In 1992 Proceedings International Conference on Arctic Margins Edited by: Thurston , D. K. and Fujita , K. 395 – 398 . Outer Continental Shelf Study, Mineral Management Service 94–0040
- Clemens , W. A. and Nelms , L. G. 1993 . Paleoecological implications of Alaskan terrestrial vertebrate fauna in latest Cretaceous time at high paleolatitudes . Geology , 21 : 503 – 506 .
- Colinvaux , P. A. 1980 . Vegetation of the Bering Land Bridge revisited . Quarterly Review of Archaeology , 1 : 2 – 15 .
- Coria , R. A. , Currie , P. J. , Eberth , D. and Garrido , A. 2002 . Bird footprints from the Anacleto Formation (Late Cretaceous) in Neuquen Province, Argentina . Ameghiniana , 39 : 1 – 11 .
- Currie , P. J. 1981 . Bird footprints from the Gething Formation (Aptian, Lower Cretaceous) of northeastern British Columbia, Canada . Journal of Vertebrate Paleontology , 1 : 257 – 264 .
- Cwynar , L. C. and Ritchie , J. C. 1980 . Arctic steppe-tundra: a Yukon perspective . Science , 208 : 1375 – 1377 .
- De Valais , S. and Melchor , R. N. 2008 . Ichnotaxonomy of bird-like footprints: an example from the Late Triassic-Early Jurassic of northwest Argentina . Journal of Vertebrate Paleontology , 28 : 145 – 159 .
- Elbroch , M. 2003 . Mammal tracks and sign , 779 Stackpole Books : Mechanicsburg .
- Elbroch , M. and Marks , E. 2001 . Bird Tracks and Sign , 456 Mechanicsburg : Stackpole Books .
- Ellenberger , P. 1972 . Contribution a la classification des pistes de vertebras du Trias: les types du Stormberg d’Afrique du Sud (I) . Palaeovertebrata Memoire Extraordinaire , : 1 – 152 .
- Erickson , B. R. 1967 . Fossil bird tracks from Utah . Museum Observer , 5 : 1 – 7 .
- Falk , A. 2009 . Interpreting behavior from Early Cretaceous bird tracks and the morphology of bird feet and trackways , 140 University of Kansas . Department of Geology Master's thesis
- Falkingham , P. L. , Margetts , L. , Smith , I. M. and Manning , P. L. 2009 . Reinterpretation of palmate and semi-palmate (webbed) fossil tracks; insights from finite element modeling . Palaeogeography, Palaeoclimatology, Palaeoecology , 271 : 69 – 76 .
- Farlow , J. O. , Langston , W. Jr. , Deschner , E. E. , Solis , R. , Ward , W. , Kirkland , B. L. , Hovorka , S. , Reece , T. L. and Whitcraft , J. 2006 . Texas Giants: Dinosaurs of the Heritage Museum of the Texas Hill Country , 105 Calgary : Dominion Exploration and Production .
- Fiorillo , A. R. 2004 . The dinosaurs of arctic Alaska . Scientific American , 291 : 84 – 91 .
- Fiorillo , A. R. 2005 . Turtle tracks in the Judith River Formation (Upper Cretaceous) of south-central Montana . Palaeontologica Electronica , 8 1 MB http://www.uv.es/pe/2005_1/fiorillo9/fiorillo9.pdf
- Fiorillo , A. R. 2006 . Review of the dinosaur record of Alaska with comments regarding Korean dinosaurs as comparable high-latitude fossil faunas . Journal of Paleontological Society of Korea , 22 : 15 – 27 .
- Fiorillo , A. R. 2008a . On the occurrence of exceptionally large teeth of Troodon (Dinosauria:Saurischia) from the Late Cretaceous of northern Alaska . Palaios , 23 : 322 – 328 .
- Fiorillo , A. R. 2008b . “ Cretaceous dinosaurs of Alaska: Implications for the origins of Beringia ” . In The Terrane Puzzle: new perspectives on paleontology and stratigraphy from the North American Cordillera Edited by: Blodgett , R. B. and Stanley , G. 313 – 326 . Geological Society of America Special Paper 442
- Fiorillo , A. R. and Gangloff , R. A. 2000 . Theropod teeth from the Prince Creek Formation (Cretaceous) of Northern Alaska, with speculations on arctic dinosaur paleoecology . Journal of Vertebrate Paleontology , 20 : 675 – 682 .
- Fiorillo , A. R. and Gangloff , R. A. 2001 . The caribou migration model for Arctic hadrosaurs (Ornithischia: Dinosauria): a reassessment . Historical Biology , 15 : 323 – 334 .
- Fiorillo , A. R. and Parrish , J. T. 2004 . The first record of a Cretaceous dinosaur from western Alaska . Cretaceous Research , 25 : 453 – 458 .
- Fiorillo , A. R. , Hasiotis , S. T. , Kobayashi , Y. and Tomsich , C. S. 2009 . A pterosaur manus track from Denali National Park, Alaska Range, Alaska, USA . Palaios , 24 : 466 – 472 .
- Flerow , C. C. 1967 . “ On the origin of the mammalian fauna of Canada ” . In The Bering Land Bridge , Edited by: Hopkins , D. M. 271 – 280 . Stanford : Stanford University Press .
- Forrest , L. R. 1988 . Field Guide to Tracking Animals in Snow , 192 Mechanicsburg : Stackpole Books .
- Fountaine , T. M. R. , Benton , M. J. , Dyke , G. J. and Nudds , R. L. 2005 . The quality of the fossil record of Mesozoic birds . Proceedings of the Royal Society, Series B , 272 : 289 – 294 .
- Gangloff , R. A. 1998 . “ Arctic dinosaurs with emphasis on the Cretaceous record of Alaska and the Eurasian–North American connection ” . In Lower and Middle Cretaceous Terrestrial Ecosystems Edited by: Lucas , S. G. , Kirkland , J. I. and Estep , J. W. 211 – 220 . New Mexico Museum of Natural History and Science Bulletin, 14
- Gangloff , R. A. , Fiorillo , A. R. and Norton , D. W. 2005 . The first Pachycephalosaurine (Dinosauria) from the Paleo-Arctic and its paleogeographic implications . Journal of Paleontology , 79 : 997 – 1001 .
- Gatesy , S. M. , Middleton , K. M. , Jenkins , F. A. Jr. and Shubin , N. H. 1999 . Three-dimensional preservation of foot movements in Triassic theropod dinosaurs . Nature , 399 : 141 – 144 .
- Genise , J. F. , Melchor , R. N. , Archangelsky , M. , Bala , L. O. , Straneck , R. and Silvina de Valais , S. 2009 . Application of neoichnological studies to behavioural and taphonomic interpretation of fossil birdlike tracks from lacustrine settings: The Late Triassic–Early Jurassic? Santo Domingo Formation, Argentina . Palaeoecology, Palaeoclimatology, Palaeoecology , 272 : 143 – 161 .
- Gill , F. B. 2007 . Ornithology , 3rd edition , 758 New York : W. H. Freeman and Company .
- Guthrie , R. D. 1982 . “ Mammals of the mammoth steppe as paleoenvironmental indicators ” . In Paleoecology of Beringia , Edited by: Hopkins , D. M. , Matthews , J. V. Jr. , Schweger , C. E. and Young , S. B. 307 – 326 . New York : Academic Press .
- Guthrie , R. D. 2001 . Origin and causes of the mammoth steppe: a story of cloud cover, woolly mammal tooth pits, buckles, and inside-out Beringia . Quaternary Science Reviews , 20 : 549 – 574 .
- Halfpenny , J. C. 1986 . A field guide to mammal tracking in North America , 161 Boulder : Johnson Books .
- Halfpenny , J. C. 1999 . Scats and tracks of the Pacific Coast, including British Columbia , 145 Helena : Falcon Publishing .
- Halfpenny , J. C. 2007 . Scats and tracks of Alaska, including the Yukon and British Columbia , 146 Helena : Falcon Publishing, Inc. .
- Hasiotis , S. T. 2002 . Continental Trace Fossils , 132 SEPM . Short Course Notes Number 51, Tulsa
- Hasiotis , S. T. 2004 . Reconnaissance of Upper Jurassic Morrison Formation ichnofossils, Rocky Mountain region, USA: environmental, stratigraphic, and climatic significance of terrestrial and freshwater ichnocoenoses . Sedimentary Geology , 167 : 277 – 368 .
- Hillhouse , J. W. and Coe , R. S. 1994 . “ Paleomagnetic data from Alaska ” . In Geology of Alaska, The Geology of North America G-1 , Edited by: Plafker , G. and Berg , H. C. 797 – 812 . Boulder : Geological Society of America .
- Hopkins , D. M. 1967 . The Bering Land Bridge , 495 Stanford : Stanford University Press .
- Hopkins , D. M. 1996 . “ Introduction: the concept of Beringia ” . In American Beginnings: the prehistory and palaeoecology of Beringia , Edited by: West , F. H. Chicago : University of Chicago Press . Pp. xvii–xxi
- Hopkins , D. M. , Matthews , J. V. Jr. , Schweger , C. E. and Young , S. B. 1982 . “ Paleoecology of Beringia ” . 489 New York : Academic Press .
- Hou , L. , Zhou , Z. , Martin , L. D. and Feduccia , A. 1995 . A beaked bird from the Jurassic of China . Nature , 377 : 616 – 618 .
- Hou , L. , Martin , L. D. , Zhou , Z. and Feduccia , A. 1996 . Early adaptive radiation of birds: evidence from northeast China . Science , 274 : 1164 – 1167 .
- Irving , L. 1960 . Birds of Anaktuvuk Pass, Kobuk, and Old Crow: a study in arctic adaptation . United States National Museum Bulletin , 217 : 1 – 409 .
- Irving , L. 1972 . Arctic Life of Birds and Mammals, Including Man , 192 Berlin : Springer-Verlag .
- Kim , J. Y. , Kim , S. H. , Kim , K. S. and Lockley , M. 2006 . The oldest record of webbed bird and pterosaur tracks from South Korea (Cretaceous Haman Formation, Changseon and Sinsu Islands): more evidence of high avian diversity in East Asia . Cretaceous Research , 27 : 56 – 69 .
- Kirschner , C. E. 1994 . “ Interior basins of Alaska ” . In Geology of Alaska, The Geology of North America G-1 , Edited by: Plafker , G. and Berg , H. C. 469 – 493 . Boulder : Geological Society of America .
- Lamanna , M. C. , You , H.-L. , Harris , J. D. , Chiappe , L. M. , Ji , S.-A. , Lu , J.-C. and Ji , Q. 2006 . A partial skeleton of an enantiornithine bird from the Early Cretaceous of northwestern China . Acta Palaeontologica Polonica , 51 : 423 – 434 .
- Lane , B. A. and Davies , J. N. 1987 . Shorebirds in Australia , 692 Melbourne : Nelson Publishers .
- Lee , Y. N. 1997 . Bird and dinosaur footprints in the Woodbine Formation (Cenomanian), Texas . Cretaceous Research , 18 : 849 – 864 .
- Lim , J.-D. , Zhou , Z. , Martin , L. D. , Baek , K.-S. and Yang , S.-Y. 2000 . The oldest known tracks of web-footed birds from the Lower Cretaceous of South Korea . Naturwissenschaften , 87 : 256 – 259 .
- Lockley , M. G. and Rainforth , E. C. 2002 . “ The track record of Mesozoic birds and pterosaurs: an ichnological and paleoecological perspective ” . In Mesozoic Birds: above the heads of dinosaurs , Edited by: Chiappe , L. and Witmer , L. M. 405 – 418 . Berkeley : University of California Press .
- Lockley , M. G. , Yang , S. Y. , Matsukawa , M. , Fleming , R. and Lim , S. K. 1992 . The track record of Mesozoic birds: evidence and implications . Philosophical Transactions of the Royal Society, Series B , 336 : 113 – 134 .
- Lockley , M. G. , Wright , J. L. and Matsukawa , M. 2001 . A new look at Magnoavipes and so-called “big-bird” tracks from Dinosaur Ridge (Cretaceous, Colorado) . The Mountain Geologist , 38 : 137 – 146 .
- Lockley , M. G. , Nadon , G. and Currie , P. J. 2003 . A diverse dinosaur-bird footprint assemblage from the Lance Formation, Upper Cretaceous, eastern Wyoming: implications for ichnotaxonomy . Ichnos , 11 : 229 – 249 .
- Lockley , M. , Matsukawa , M. , Ohira , H. , Li , J. , Wright , J. , White , D. and Chen , P. 2006 . Bird tracks from Liaoning Province, China: new insights into avian evolution during the Jurassic-Cretaceous transition . Cretaceous Research , 27 : 33 – 43 .
- Lockley , M. , Chin , K. , Houck , K. , Matsukawa , M. and Kukihara , R. 2009 . New interpretations of Ignotornis, the first-reported Mesozoic avian footprints: implications for the paleoecology and behavior of an enigmatic Cretaceous bird . Cretaceous Research , 30 : 1041 – 1061 .
- Manning , P. L. 2004 . “ A new approach to the analysis and interpretation of tracks: examples from the Dinosauria ” . In The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis , Edited by: McIlroy , D. 93 – 123 . Geological Society of London . Special Publication, 228
- Marsh , O. C. 1880 . Odontornithes: a monograph on the extinct toothed birds of North America Vol. 201 , United States Geological Exploration of the 40th Parallel, Clarence King, Geologist-in-charge, 7
- Mehl , M. G. 1931 . Additions to the vertebrate record of the Dakota Sandstone . American Journal of Science, Fifth Series , 21 : 441 – 452 .
- Murie , O. J. 1954 . A Field Guide to Animal Tracks , 374 Boston : Houghton Mifflin Co. .
- Nelms , L. G. 1989 . Late Cretaceous dinosaurs from the North Slope of Alaska . Journal of Vertebrate Paleontology , 9 ( supplement to 3 ) : 34A
- Olson , S. L. and Parris , D. C. 1987 . The Cretaceous birds of New Jersey . Smithsonian Contributions to Paleobiology , 63 : 1 – 22 .
- O’Neill , D. 2004 . The last giant of Beringia: the mystery of the Bering Land Bridge , Vol. 231 , Boulder : Westview Press .
- Panin , N. and Avram , E. 1962 . Noe urme de vertebrate in Miocenul Subcarpaţilor romineşti . Studii şi Cercetări de Geologie , 7 : 455 – 484 .
- Parrish , M. J. , Parrish , J. T. , Hutchinson , J. H. and Spicer , R. A. 1987 . Late Cretaceous vertebrate fossils from the North Slope of Alaska and implications for dinosaur ecology . Palaios , 2 : 377 – 389 .
- Paul , G. S. 1988 . “ The small predatory dinosaurs of the mid-Mesozoic, the horned small theropods—Proceratosaurus and Ornitholestes—of the Morrison and Great Oolite, and the sickle claw theropods—Deinonychus, Velociraptor and Saurornithoides—of the Cloverly, Djadokhta and Judith River ” . Vol. 2 , 1 – 9 . Hunteria
- Repenning , C. A. 1987 . “ Biochronology of the microtine rodents of the United States ” . In Cenozoic Mammals of North America: geochronology and biostratigraphy , Edited by: Woodburne , M. O. 236 – 268 . Berkeley : University of California Press .
- Repenning , C. A. 1990 . Of mice and men in the Late Pliocene of North America . Arctic , 43 : 314 – 323 .
- Repenning , C. A. , Brouwers , E. M. , Carter , L. D. , Marincovich , L. Jr. and Ager , T. A. 1987 . The Beringian history of Phenacomys (Rodentia: Cricetidae) and the beginning of the modern Arctic Ocean biota . United States Geological Survey Bulletin , 1687 : 1 – 35 .
- Rezendes , P. 1999 . Tracking and the Art of Seeing: how to read animal tracks and sign , 2nd edition , 336 New York : Harper Collins Publishers .
- Rich , T. H. , Vickers-Rich , P. and Gangloff , R. A. 2002 . Polar Dinosaurs . Science , 295 : 979 – 980 .
- Richards , A. 1988 . Shorebirds: a complete guide to their behavior and migrations , 224 New York : Gallery Books .
- Ridgway , K. D. , Trop , J. M. and Sweet , A. R. 1997 . Thrust-top basin formation along a suture zone, Cantwell basin, Alaska Range: implications for the development of the Denali fault system . Geological Society of American Bulletin , 109 : 505 – 523 .
- Russell , D. A. 1993 . The role of central Asia in dinosaurian biogeography . Canadian Journal of Earth Sciences , 30 : 2002 – 2012 .
- Sereno , P. C. 2000 . “ The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia ” . In The age of dinosaurs in Russia and Mongolia , Edited by: Benton , M. J. , Shishkin , M. A. , Unwin , D. M. and Kurochkin , E. N. 480 – 516 . Cambridge : Cambridge University Press .
- West , F. H. 1996 . American Beginnings: the prehistory and paleoecology of Beringia , 576 Chicago : University of Chicago Press .
- Yang , S.-Y. , Lockley , M. G. , Greben , R. , Erickson , B. R. and Lim , S-K. 1995 . Flamingo and duck-like bird tracks from the Late Cretaceous and Early Tertiary: evidence and implications . Ichnos , 4 : 21 – 34 .
- Zhang , J. , Li , D. , Li , M. , Lockley , M. G. and Bai , Z. 2006 . Diverse dinosaur-, pterosaur-, and bird-track assemblages from Hakou Formation, Lower Cretaceous of Gansu Province, northwest China . Cretaceous Research , 27 : 44 – 55 .