
Abstract
The sauropod dinosaur Alamosaurus sanjuanensis Gilmore, 1922 has been known from Maastrichtian deposits of south-western North America for nearly a century. Alamosaurus is the youngest sauropod taxon known in North America. Originally described from an isolated scapula and ischium from New Mexico, more of Alamosaurus was revealed by an incomplete skeleton from southern Utah. Additional referred specimens from western Texas provided the first few examples of cervical, dorsal and sacral vertebrae known for the taxon, but these came from relatively small and immature individuals. Here we describe an articulated series of cervical vertebrae of a large, mature titanosaur from Big Bend National Park, Texas, and provide evidence that the specimen can be referred to A. sanjuanensis. The specimen represents the first articulated cervical vertebral series described for the taxon, which clarifies aspects of cervical vertebral anatomy and provides at least one new diagnostic character for Alamosaurus. Many previous cladistic analyses found Alamosaurus to be a saltasaurid titanosaur, sometimes closely related to the Asian taxon Opisthocoelicaudia skarzynskii. We present cladistic analyses incorporating new data from this and other specimens from Big Bend National Park. The first places Alamosaurus as a lithostrotian titanosaur outside Saltasauridae. The second analysis, with greater focus on South American titanosaurs, finds Alamosaurus allied to Lognkosauria, a clade of South American titanosaurs notable for giant size and exceptionally robust necks. This relationship may be more congruent with the fossil record than hypotheses of phylogeny that would require saltasaurid titanosaurs to inhabit northern Laramidia, Beringia and Central Asia through the late Campanian and Maastrichtian while leaving no record of their presence.
Introduction
The titanosaur sauropod dinosaur Alamosaurus sanjuanensis Gilmore, Citation1922 has long held a special place in our understanding of North American sauropod palaeobiogeography and palaeoecological reconstructions (Gilmore Citation1921, Citation1922, Citation1938, Citation1946; Lucas & Hunt Citation1989; Coulson Citation1998; Lehman & Coulson Citation2002). The first specimens of Alamosaurus were isolated elements that only hinted at the relationships of this Maastrichtian North American taxon (Gilmore Citation1921, Citation1922). A more complete specimen described in 1946, and additional specimens collected from the far western reaches of Texas, Utah and New Mexico from the 1970s to the present, have filled in much information about the skeletal morphology of the taxon (Gilmore Citation1946; Lucas & Hunt Citation1989; Lehman & Coulson Citation2002; D'Emic et al. Citation2011; Carrano & D'Emic Citation2015). Yet even these specimens left many gaps in our understanding of the osteology of Alamosaurus, particularly with regard to cervical vertebral anatomy of large, skeletally mature individuals.
The articulated series of sauropod cervical vertebrae described here was discovered in 1997 during a joint Perot Museum of Nature and Science–University of Texas at Dallas (UTD) field project focused on latest Cretaceous strata in Big Bend National Park (Fiorillo Citation1998; Myers & Fiorillo Citation2009). While crews were excavating a locality that yielded remains of immature sauropods in the north-eastern section of the park, Dana Biasatti, then a student at UTD, discovered a nearby site that produced the articulated cervical vertebral series in this report. Members of the Perot Museum of Nature and Science and the University of Texas at Dallas worked with administrative personnel at Big Bend National Park to excavate the specimen (BIBE 45854) and transport it to Dallas, Texas.
Material and methods
Institutional abbreviations
BIBE: Big Bend National Park, Texas, USA; BYU: Brigham Young University, Provo, Utah, USA; DMNH: Perot Museum of Nature and Science, Dallas, Texas, USA; TMM: University of Texas at Austin, Vertebrate Paleontology Laboratory, Austin, Texas, USA; USNM: Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA.
Geological setting
The site from which BIBE 45854 was collected is in the north-eastern part of Big Bend National Park, in an area of low-relief badlands called Tornillo Flat () (Maxwell et al. Citation1967; Maxwell Citation1968; Cooper et al. Citation2011). Exact geographical co-ordinates are on file at the Perot Museum of Nature and Science, in Dallas, Texas, USA. When the specimen was found it was believed that the majority of the terrestrial Upper Cretaceous sediments exposed in the immediate area consisted of the Javelina Formation (Maastrichtian), with minor exposures of the Aguja Formation (late Campanian) and Black Peaks Formation (Maastrichtian–Paleocene) in the vicinity (Maxwell et al. 1967; Fiorillo Citation1998). A tuff bed in the middle of the Javelina Formation near Grapevine Hills several kilometres to the west of the fossil site was radioisotopically dated to 69.0 ± 0.9 Ma, the early late Maastrichtian (Lehman et al. Citation2006; Gradstein et al. Citation2012). However, recent geological studies and more detailed mapping within Big Bend National Park concluded that many of the rocks exposed in the Tornillo Flat area previously identified as Javelina Formation are instead referable to the overlying Black Peaks Formation (Cooper et al. Citation2011; Turner et al. Citation2011). A visit to the site in 2012 confirmed that BIBE 45854 is from the lower part of the Black Peaks Formation (Maxwell et al. Citation1967; Cooper et al. Citation2011; Turner et al. Citation2011). The Black Peaks Formation is a terrestrial unit comprised of fluvial facies, including channel, overbank floodplain, and freshwater pond and lake sediments. The site that produced BIBE 45854 is in an area of badlands with limited topographical relief, intervals of Quaternary sediment cover, and some local faulting. As of this writing, the exact stratigraphical position of BIBE 45854 within the lower Black Peaks Formation has not yet been determined.
Figure 1. Location of Big Bend National Park in Texas, USA, shown by solid grey fill. White star within expanded outline of Big Bend National Park marks the approximate location of the BIBE 45854 excavation site within Tornillo Flat.
Other remains from ontogenetically immature titanosaurs were collected from another site approximately 0.5 km to the south of BIBE 45854. As will be discussed below, elements from the site preserve features cited as autapomorphies of Alamosaurus sanjuanensis (D'Emic et al. Citation2011). We therefore refer the titanosaur material from this site to Alamosaurus. These specimens will be the subjects of future work, and with the exception of those elements pertinent to establishing the affinities of BIBE 45854, the rest of the material is not discussed here. The juvenile specimens are also housed in the collections of the Perot Museum of Nature and Science, in Dallas, Texas.
Excavation, preparation and exhibition
Excavation of BIBE 45854 was performed under National Park Service Permit BIBE-2001-SCI-0026, by personnel from the Perot Museum of Nature and Science (at that time the Dallas Museum of Natural History) with volunteers and partners from The University of Texas at Dallas. Removal of the large blocks containing each vertebra was complicated by the designated wilderness status of the immediate area. As a result, the blocks were individually airlifted from the site by a helicopter, and then driven by truck to Dallas, Texas, for preparation and curation.
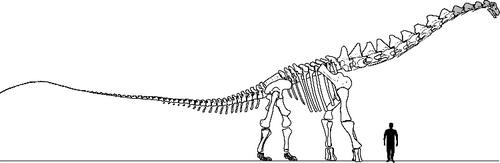
It was decided in 2009 that the articulated vertebral series would be exhibited in the Perot Museum of Nature and Science building to be built near downtown Dallas, Texas. In addition, the specimens were laser digitized, as were the holotype and referred material of Alamosaurus sanjuanensis housed at the Smithsonian Institution (USNM 10485, USNM 10487, USNM 15560), and a partial skeleton in the collections of The University of Texas at Austin (TMM 45145-1). The digital scans of all three specimens were then used to create the first full-body reconstruction of the skeleton of Alamosaurus sanjuanensis for exhibition (). The reconstructed skeleton measures approximately 26 m in length, and 5 m tall to the dorsal tip of the neural spine of the first dorsal vertebra.
Figure 2. Diagrammatic skeletal drawing of Alamosaurus sanjuanensis based on composite skeleton reconstruction exhibited at the Perot Museum of Nature and Science, in Dallas, Texas. Reconstruction based on BIBE 45854, TMM 41541-1, USNM 15560 and the distal caudal series of a not-yet-named South American titanosaur. Human silhouette represents a 1.8 m tall individual. Grey-filled elements indicate parts of reconstruction based upon estimates of anatomy or modifications of other taxa.
The fossils were encased in a concretionary layer of sandy carbonate material, with small pockets of fine sand and clay that showed fine sedimentary laminae. Naturally occurring breaks through the specimens dictated the piece-by-piece disassembly, preparation and reassembly of each vertebra, but also facilitated access to many of the deep fossae and delicate laminae preserved in the specimens. Almost all preparation was done with pneumatic ‘airscribes’. The great size and weight of the vertebrae dictated the use of strong, slow-setting epoxy resin as an adhesive during reassembly of the bones. No internal metal pins, rods or bolts were used to re-connect pieces or strengthen joined components.
Numerous photographs were taken during the time-sensitive preparation process in order to document and record anatomical features not normally visible in assembled vertebrae. Measurements of individual vertebrae were taken during the preparation process, but because of project time constraints not all the vertebrae were subjected to full sets of measurements and photographs prior to exhibition. Preparation, laser scanning and digitization of all the elements were completed by late 2011. The bones were then shipped to a third party to be mounted on external steel armatures, and posed in the articulated position in which they were found. The vertebrae were finally placed on public exhibit in the new Perot Museum of Nature and Science building in 2012.
Anatomical terminology
We use the classic ‘Romerian’ (Wilson Citation2006) anatomical terminology commonly employed in discussion and description by palaeontologists for much of the twentieth and early twenty-first centuries. There has been a recent rise in the use of terminology derived from the Nomina Anatomica Avium (Baumel et al. Citation1993) as applied to dinosaur anatomy, but we agree with arguments by Wilson (Citation2006, Citation2012) that: (1) in many cases the push for wholesale standardization of terms results in cumbersome and potentially confusing terminology; (2) it creates nomenclatural inconsistency for negligible benefit when long-standing and utilitarian terminology is replaced by novel terms; (3) it costs more in lost communication and understanding between current and prior works than any positive gain extracted from adherence to a new, rigid standard; and (4) such a push for terminological replacement is essentially a solution for a non-existent problem. Also, works that have revolutionized the descriptive standards for sauropod vertebral anatomy (Wilson Citation1999, Citation2012; Wilson et al. Citation2011) have retained the classic palaeontological terminology that has served the science well. We also use the descriptive terminology and abbreviations for vertebral laminae and fossae established by Wilson (Citation1999, Citation2012) and Wilson et al. (Citation2011), substituting capitalized abbreviations for the names of structures after their first mention in the text.
Taxonomic definitions
The taxonomy applied to groups of titanosaurian sauropods has a convoluted history, with the past two decades seeing a proliferation of new names, re-definitions proposed for previously erected and defined names, and arguments to abandon other names (Salgado et al. Citation1997; Sereno Citation1998; Wilson & Sereno Citation1998; Wilson & Upchurch Citation2003; Upchurch et al. Citation2004). Even the definition of the name Titanosauria has varied since the name was first erected by Salgado et al. (Citation1997). Originally established with a node-based definition (= Andesaurus delgadoi + Titanosauridae and all descendants of their most recent common ancestor; Salgado et al. Citation1997), some subsequent works adopted a stem-based definition for the name and anchored the definition on other, more specific taxa (= all titanosauriforms more closely related to Saltasaurus loricatus than to Euhelopus zdanskyi; Sereno Citation1998; Wilson & Sereno Citation1998); then it was used again as a node-based name (= Andesaurus delgadoi + Saltasaurus loricatus and all descendants of their most recent common ancestor, Wilson & Upchurch Citation2003). The discovery of new titanosauriform taxa in recent years has also produced new hypotheses of titanosauriform phylogeny (Curry Rogers Citation2005; Calvo et al. Citation2007a,Citationb; González Riga et al. Citation2009; D'Emic Citation2012; Mannion et al. Citation2013; Carballido & Sander Citation2014), which have sometimes resulted in taxonomic redundancy stemming from the shuffling of taxa used to anchor taxonomic definitions. For the purpose of this paper, we will follow the principle of priority whenever possible, adopting the first definition of a clade or lineage with an ancestry-based stem or node definition. A list of taxon names and definitions used in this work is given in
Table 1. Definitions of clade and lineage names mentioned in this work. Names are listed in alphabetical order. Two entries are provided for Saltasaurinae, reflecting definitions of the name as first given by Salgado et al. (Citation1997), but acknowledging the wider usage of Sereno (Citation1998).
Systematic palaeontology
Dinosauria Owen, Citation1842, sensu Padian & May 1993
Sauropoda Marsh Citation1878, sensu Wilson & Sereno Citation1998
Titanosauriformes Salgado et al., Citation1997
Titanosauria Bonaparte & Coria, Citation1993, sensu Wilson & Upchurch Citation2003
Alamosaurus sanjuanensis Gilmore, Citation1922
(–)
Revised diagnosis
Alamosaurus sanjuanensis was given an emended diagnosis by Lehman & Coulson (Citation2002), based mostly upon characters preserved in elements of TMM 43621-1, a relatively immature individual from Big Bend National Park, Texas, USA. Many of the features cited by them are now known to have a wider distribution among titanosaurs (Upchurch Citation1998; Wilson Citation2002; Upchurch et al. Citation2004; Curry Rogers Citation2005; González Riga & Ortiz David Citation2014). The case was recently made again that Alamosaurus sanjuanensis is a diagnosable taxon (D'Emic et al. Citation2011) based on features preserved in the holotype scapula (USNM 10486) and referred specimen (USNM 15560), including: the presence of two low processes on the ventral margin of the scapular blade distal to the glenoid fossa versus a single or no process in almost all other titanosauriforms; an asymmetrically concave scapular blade with thicker ventral margin and thinner dorsal margin; dorsal edge of scapular blade nearly straight and ventral edge expanded distally; a circumferential depression limited to the ventral half of the anterior condyle of the biconvex first caudal centrum; and multiple large foramina piercing the lateral surface of the centrum of the first caudal vertebra. Several additional titanosaur specimens collected within Big Bend National Park preserve additional characters that may also be diagnostic for the taxon if they can be referred to Alamosaurus (Wick & Lehman Citation2014; Fronimos & Lehman Citation2014). As explained below, BIBE 45854 can be referred to A. sanjuanensis, and we offer a new diagnostic vertebral character for the taxon: posterior cervical vertebrae with the prezygapophyseal centrodiapophyseal fossae (PRCDF) divided into dorsal and ventral sub-fossae by a lamina that intersects the anterior centrodiapophyseal lamina (ACDL) at nearly a right angle and continues anteriorly to join the base of the prezygapophysis.
Description
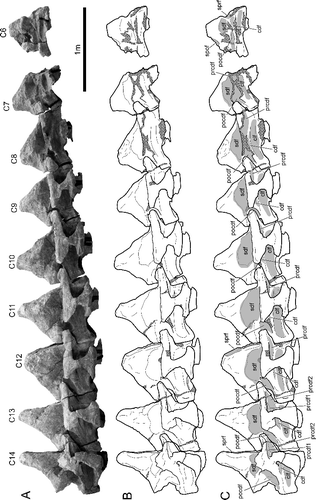
The specimen described here (BIBE 45854) consists of nine articulated cervical vertebrae (). The individual vertebrae of BIBE 45854 were given sequential letter designations during excavation, the anterior-most vertebra being designated ‘B’ and the most posterior in the articulated series being ‘J’. The alphabetical convention was consistently used to identify individual vertebrae in field notes, preparation notes, photographic records and other files related to the specimens. The cervical vertebral count varies among titanosaurs. Rapetosaurus krausei from Madagascar was reported to have 17 cervical vertebrae, while the Argentinian giant Futalognkosaurus dukei has 14 cervical vertebrae, and the Brazilian titanosaur ‘series A’ has only 13 cervical vertebrae (Powell Citation2003; Calvo et al. Citation2007a, Citationb; Curry Rogers Citation2009). We tentatively assume a cervical vertebral count of 14 in Alamosaurus, based on the complete articulated cervical series of Futalognkosaurus dukei. We also identify the most posterior preserved vertebra in BIBE 45854 as the last cervical, vertebra 14, based on its anteroposteriorly short centrum, tall but anteroposteriorly narrow neural spine, and abbreviated cervical ribs (A, B). The most anterior specimen in the preserved series (vertebra B) is therefore identified as the sixth cervical vertebra. We acknowledge that additional specimens may show these assumptions to be incorrect. In order to facilitate references between the vertebrae and their mention in preparation records and field notes, at first mention in the text we identify each vertebra by both its number in the series (cervical 6, cervical 7, etc.) and by the letter designation it carried from excavation through preparation (vertebra B, C, etc.).
Figure 3. BIBE 45854, articulated series of nine mid and posterior cervical vertebrae of a large, osteologically mature Alamosaurus sanjuanensis. Series is estimated to represent the sixth to fourteenth cervical vertebrae. A, composite photo-mosaic of the cervical series in right lateral view; identification of each vertebra indicated by C6 to C14, respectively. B, line drawing based on the photo-mosaic in A. C, line drawing in B with labels shown and vertebral fossae indicated by solid grey fill; cross-hatching represents broken bone surfaces and reconstructive material. Abbreviations: C, cervical vertebra; cdf, centrodiapophyseal fossa; clf, centrum lateral fossa; pocdf, postzygapophyseal centrodiapophyseal fossa; prcdf, prezygapophyseal centrodiapophyseal fossa; prcdf1, dorsal prezygapophyseal centrodiapophyseal fossa; prcdf2, ventral prezygapophyseal centrodiapophyseal fossa; sdf, spinodiapophyseal fossa; spof, spinopostzygapophyseal fossa; sprf, spinoprezygapophyseal fossa.
The cervical vertebrae of BIBE 45854 all share certain characteristics, and selected dimensions of the vertebrae are given in . The neurocentral sutures between the centra and the neural arches are all closed and fused, an indication of relatively advanced ontogenetic stage in dinosaurs and other archosaurs (Brochu Citation1996; Ikejiri Citation2003, Citation2004). This is in marked contrast to the cervical vertebrae of TMM 43621-1, a previously described specimen referred to Alamosaurus in which the cervical centra and neural arches were separated and disarticulated, an indication of relative immaturity (Lehman & Coulson Citation2002). In addition, the cervical ribs of BIBE 45854, where preserved, are fused to their respective vertebrae, another sign of relatively advanced developmental stage. The internal structure of all the vertebrae was extensively exposed during the preparation process. Nearly every element has extensive somphospondylus internal structure, including the centra, neural arches and spines, diapophyseal laminae, and zygapophyses (A). This is consistent with previous work examining the internal bone structure in Alamosaurus (Woodward & Lehman Citation2009). Only parts of the cervical ribs, mainly the rib shafts, are not somphospondylus.
Table 2. Selected measurements from BIBE 45854, Alamosaurus sanjuanensis. Abbreviations: VERT, vertebra number/field ID letter; CL, centrum length from anterior condyle to most posterior edge of ventral surface; CH, centrum height measured as maximum median dorsoventral height across outer rim of posterior cotyle; CW, centrum width measured as maximum mediolateral width across outer rim of posterior cotyle; NAH, neural arch height from ventral floor of neural canal anterior aperture to dorsal tip of neural spine; NSH, neural spine height from dorsal surface of intraprezygapophyseal lamina to dorsal tip of neural spine; NSW, neural spine width measured across maximum mediolateral expanse of neural spine; PPL, prezygapophysis to postzygapophysis length from extreme distal tips of each on same side; VHC, vertebra height measured from ventral surface of posterior edge to dorsal tip of neural spine; VHR, vertebra height from most ventral surface of a cervical rib to the dorsal tip of neural spine; VMW, vertebra maximum width across widest point of diapophyses or cervical ribs. – = no data; * = estimated measurement; = > value is minimum dimension. All dimensions given in millimetres.
Figure 4. BIBE 45854, Alamosaurus sanjuanensis. A, cervical vertebra 10 in dorsal view during preparation, with neural spine removed. B, interpretive line drawing of image in A. Somphospondylus internal structure is visible in broken neural spine laminae in A. Solid grey fill in B indicates broken bone surfaces. Dashed lines indicate approximate borders of anterior condyle and prezygapophyses not attached to centrum at this stage of preparation. Abbreviations: ant, anterior; crib, cervical rib; podl, postzygodiapophyseal lamina; ppr, posterior process of posterior centrodiapophyseal lamina; sdf, spinodiapophyseal fossa; spof, spinopostzygapophyseal fossa; sprf, spinoprezygapophyseal fossa; spol, spinopostzygapophyseal lamina; sprl, spinoprezygapophyseal lamina; tprl, intraprezygapophyseal lamina. Divisions on metre scale bar are 10 cm.
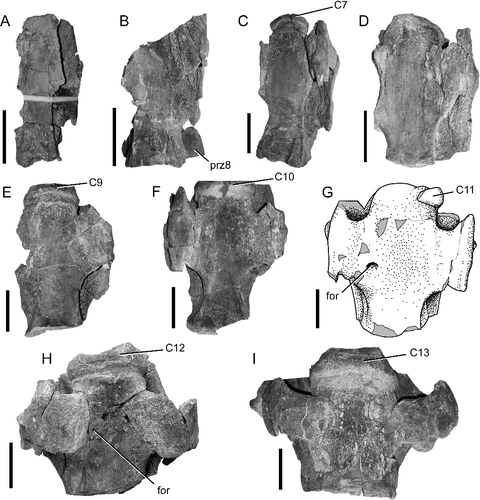
The centra are strongly opisthocoelous with articular surfaces wider than tall, and have large anterior condyles that protrude far into the deep posterior concavity of the preceding vertebral centrum. The transition from ventral to lateral surfaces of each centrum is demarcated by ventrolateral ridges posterior to the parapophyses. The parapophyses project ventrolaterally from the centrum, but not to the degree seen in diplodocids such as Apatosaurus (Upchurch et al. Citation2004). The ventral surface of each centrum is concave anteroposteriorly and transversely. The transverse concavity is most pronounced between the parapophyses. The ventral surfaces on all the vertebrae lack ventral keels and defined fossae (), in contrast to the fossae and keels seen in the posterior cervical vertebrae of Overosaurus paradasorum (Coria et al. Citation2013). The ventral surfaces of the centra are marked by numerous small foramina, possibly mirroring the somphospondylus internal structure of the vertebrae. However, there is a large, round foramen medial to the right parapophysis-cervical rib junction of cervical vertebra 12 (vertebra H), and a smaller, less invasive foramen in almost the same place on the thirteenth cervical vertebra (vertebra I) (G, H). The ventral surface of the fourteenth cervical vertebra (vertebra J) shows multiple smaller foramina adjacent to the parapophysis-cervical rib junctions, as well as near the posterior margin of the anterior condyle (I). These foramina appear to lead to the network of small internal cavities that fill the internal volume of the centra.
Figure 5. BIBE 45854, Alamosaurus sanjuanensis, cervical vertebrae in ventral view. Photos taken at various preparation stages for each vertebra. A, cervical vertebra 6. B, cervical vertebra 7 and prezygapophysis of cervical vertebra 8. C, cervical vertebra 8 and posterior rim of centrum from cervical vertebra 7; angle of photo taken in ventral slightly left lateral view. D, cervical vertebra 9. E, cervical vertebra 10 and posterior edge of centrum of cervical vertebra 9. F, cervical vertebra 11 and posterior end of cervical vertebra 10. G, line drawing of cervical vertebra 12, derived from sketch made during preparation of specimen. H, cervical vertebra 13 and posterior end of centrum of cervical vertebra 12. I, cervical vertebra 14 and posterior end of centrum of cervical vertebra 13. Abbreviations: C, part of cervical centrum; for, foramen; prz8, prezygapophysis of cervical vertebra 8. All scale bars = 20 cm.
The centra increase in anteroposterior length from the sixth cervical (vertebra B) to the eleventh (vertebra G) and twelfth cervical vertebrae, which are nearly the same length (). The exact lengths of the latter two vertebral centra are uncertain because the anterior condyle of each is articulated with the preceding centrum, and measurements can only be estimated to within one or two centimetres. The centra of cervical vertebrae 13 and 14 are successively shorter anteroposteriorly than the twelfth centrum, approximately 90% and 79% as long, respectively. The cervical centra gradually increase in maximum mediolateral width (measured just anterior to the parapophyses) from anterior to posterior in the series. The proportional changes in the cervical series are reflected in the average elongation index (aEI; Chure et al. Citation2010) of each centrum ().
Table 3. Selected measurements from BIBE 45854 (Alamosaurus sanjuanensis) reflecting vertebral proportions, including aEI. Abbreviations: VERT, vertebra number/field ID letter; CL, centrum length from anterior condyle to most posterior edge of ventral surface; CLnc, centrum length not counting anterior condyle; CH, centrum height measured as maximum median dorsoventral height across outer rim of posterior cotyle; CW, centrum width measured as maximum mediolateral width across outer rim of posterior cotyle; L/H, ratio of full centrum length including anterior condyle divided by centrum height; aEI, average elongation index, which is centrum length not including the anterior condyle, divided by centrum height. – = no data; * = estimated measurement; > = value is minimum dimension. All dimensions given in millimetres.
The lateral surfaces of each centrum are excavated by a broad fossa on each side, but there are no well-defined pneumatic sub-fossae or additional laminae dividing the larger lateral excavations. In this regard they resemble the cervical centra of TMM 43621-1, the immature individual referred to Alamosaurus from Big Bend National Park (Lehman & Coulson Citation2002). The parapophyses are anteroposteriorly elongate, measuring half or more the length of the centrum in the most posterior three to four vertebrae in the series. The parapophyses are not excavated dorsally by pneumatic fossae. Unlike the other parapophyses in the series, the anterior ends of those of the fourteenth cervical vertebra are sharply upturned, nearly forming a right angle in cross section through the base of each parapophysis.
Well-developed centrodiapophyseal laminae bound deep fossae in the neural arches ventral to the transverse processes. The ACDL and posterior centrodiapophyseal laminae (PCDL) are nearly equal in size and robustness in the more anterior vertebrae. They define the dorsal margins of small, shallow centrodiapophyseal fossa (CDF) in the seventh to ninth cervical vertebrae (). The PCDL are much larger than the ACDL in the more posterior vertebrae, attaining maximum thickness in cervical vertebrae 12 and 13 (). The ACDL are also reduced in length and thickness in the more posterior vertebrae. The ventral part of the right ACDL divides to form two smaller laminae to contact the centrum in cervical vertebra 14, but it cannot be determined at this time if its counterpart on the left side also exhibits this division. The CDF in cervical vertebrae 12 to 14 are much deeper and open more ventrolaterally than those of the anterior vertebrae in the series.
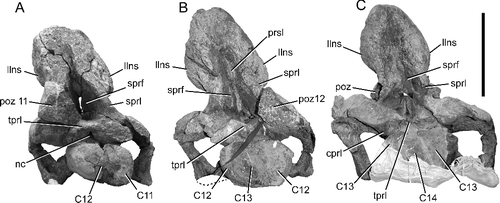
The prezygapophyses project anteriorly far beyond the condyle of each cervical vertebra, to the extent that the prezygapophyses closely approach the postzygodiapophyseal laminae (PODL) of the preceding vertebrae (, ). In this regard, the prezygapophyses resemble the condition in many titanosaurs such as Malawisaurus dixeyi, Trigonosaurus pricei and Dreadnoughtus schrani (Campos et al. Citation2005; Gomani Citation2005; Lacovara et al. Citation2014). This is in marked contrast to the condition in the saltasaurines Saltasaurus and Neuquensaurus, in which the prezygapophyses are directed anterolaterally and stop far short of the anterior end of the condyle (Powell Citation2003; Salgado et al. Citation2005; Zurriaguz & Powell Citation2015). The centroprezygapophyseal laminae (CPRL) are dorsoventrally deep and extend nearly to the anterior tip of the prezygapophyses. The prezygodiapophyseal laminae (PRDL) also originate near the anterior tip of the prezygapophyses. They are directed mostly ventrally in the mid-cervical series, and become more horizontally oriented in the posterior cervical vertebrae, especially in vertebrae thirteen and fourteen (, ). The intraprezygapophyseal laminae (TPRL) are robust throughout the series, each making a thick dorsal roof to the anterior aperture of the neural canal (). The TPRL is mediolaterally broad and horizontally oriented in cervical vertebrae 6 to 12 (A), but is angled on the midline to form a broad ‘V’ in cervical vertebrae 13 and 14 (B, C).
Figure 6. BIBE 45854, Alamosaurus sanjuanensis, cervical vertebrae 6 to 14 in dorsal view. A, cervical vertebrae 14, 13 and posterior part of 12. B, anterior part of cervical vertebra 13, cervical vertebra 12 and posterior part of cervical vertebra 11. C, anterior part of cervical vertebra 12, cervical vertebra 11 and posterior part of cervical vertebra 10. D, anterior part of cervical vertebra 10, cervical vertebra 9 and posterior half of cervical vertebra 8. E, anterior part of cervical vertebra 9, cervical vertebra 8 and posterior part of cervical vertebra 7. F, anterior half of cervical vertebra 8 and cervical vertebra 7. G, anterior part of incomplete cervical vertebra 7 and cervical vertebra 6. Solid grey fill in G indicates broken bone surface. All scale bars = 50 cm.

Figure 7. BIBE 45854, Alamosaurus sanjuanensis posterior cervical vertebrae in anterior view. A, cervical vertebra 12. B, cervical vertebra 13; security strap lashing vertebra to shipping pallet visible in photo. C, cervical vertebra 14; temporary plaster transportation base obscures ventral edges of vertebra. Photos taken during late stages of specimen preparation. Abbreviations: C, cervical vertebra, identifies parts of adjacent vertebrae in images; cprl, centroprezygapophyseal lamina; llns, lateral lamina of neural spine; nc, neural canal; poz, postzygapophysis; poz 11, postzygapophysis of cervical 11; poz 12, postzygapophysis of cervical 12; prsl, prespinal lamina; sprf, spinoprezygapophyseal fossa; sprl, spinoprezygapophyseal lamina; tprl, intraprezygapophyseal lamina. Scale bar = 50 cm.
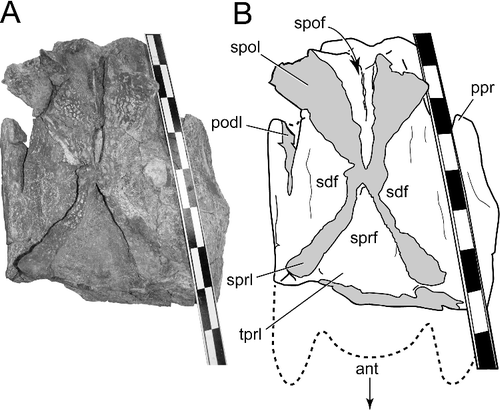
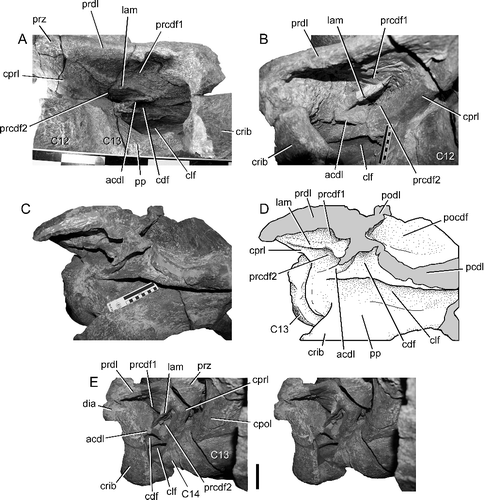
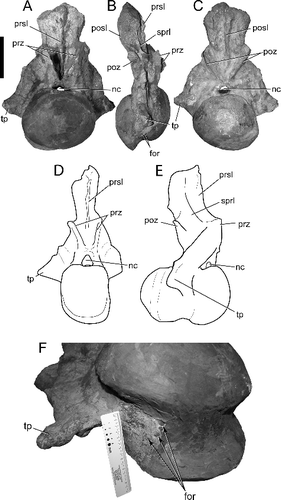
The dorsoventrally deep CPRL, pronounced PRDL and ACDL enclose large, anteroventrally facing PRCDF on each vertebra. On the thirteenth and fourteenth cervical vertebrae, a delicate, thin lamina divides each PRCDF into dorsal and ventral sub-fossae (, ). The same laminae and divisions of the PRCDF are preserved on both sides of each of these two vertebrae, evidence against the condition being the result of pneumatic asymmetry or some other individual aberration. The unusual lamina arises from the wall of the fossa, tapers away anteriorly onto the lateral surface of the prezygapophyseal process and posteriorly intersects the mid-point of the ACDL at nearly a right angle (). This division of the PRCDF of the posterior-most cervical vertebrae appears to be unique among titanosaurs, and is recognized here as an autapomorphy for Alamosaurus sanjuanensis.
Figure 8. BIBE 45854, Alamosaurus sanjuanensis. Close-up views of autapomorphic lamina dividing the prezygapophyseal centrodiapophyseal fossa into dorsal and ventral sub-fossae. A, cervical vertebra 13, left lamina and fossae in anterolateral view. B, cervical vertebra 13, right lamina and fossae in anterolateral view. C, cervical vertebra 14, centrum and arch laminae and fossae in lateral view during preparation of specimen. D, line drawing of view in C. E, cervical vertebra 14, stereophotopairs of right prezygapophyseal centrodiapophyseal fossa and dividing lamina in anterolateral view. Solid grey fill indicates broken bone surface. Abbreviations: acdl, anterior centrodiapophyseal lamina; C, cervical vertebra; cdf, centrodiapophyseal fossa; clf, centrum lateral fossa; cpol, centropostzygapophyseal lamina; cprl, centroprezygapophyseal lamina; crib, cervical rib; dia, diapophysis; lam, lamina dividing the prezygapophyseal centrodiapophyseal fossa; pcdl, posterior centrodiapophyseal lamina; pocdf, postzygapophyseal centrodiapophyseal fossa; podl, postzygodiapophyseal lamina; pp, parapophysis; prcdf1, dorsal prezygapophyseal centrodiapophyseal fossa; prcdf2, ventral prezygapophyseal centrodiapophyseal fossa; prdl, prezygodiapophyseal lamina; prz, prezygapophysis. Scale bar in A shows 10 cm divisions; scale bars in B, C and E = 10 cm.
The postzygapophyses are short on all the cervical vertebrae, and do not extend as far as the posterior rim of their respective centra (). In this regard BIBE 45854 again stands in marked contrast to the saltasaurines Saltasaurus and Neuquensaurus, taxa in which the cervical postzygapophyses project far posterior beyond their respective centra. The cervical postzygapophyses also reach to or beyond the posterior rim of the centra in other titanosaurs, such as Isisaurus colberti, Rapetosaurus krausei and Overosaurus paradasorum (Jain & Bandyopadhyay Citation1997; Curry Rogers Citation2009; Coria et al. Citation2013). The presence of short postzygapophyses in BIBE 45854 more closely resembles the condition in Dreadnoughtus schrani, some posterior cervical vertebrae of Malawisaurus dixeyi, Trigonosaurus pricei, brachiosaurids, Sauroposeidon proteles and TMM 43621-1 (Wedel et al. Citation2000a, Citationb; Lehman & Coulson Citation2002; Campos et al. Citation2005; Gomani Citation2005; Lacovara et al. Citation2014). The postzygapophyses are dorsoventrally deep. A thick epipophysis on each postzygapophysis blends smoothly into the spinopostzygapophyseal lamina (SPOL), making the epipophysis difficult to discern (). Each vertebra bears short but robust postzygodiapophyseal laminae (PODL), and dorsoventrally tall centropostzygapophyseal laminae (CPOL) (). Together with the strongly developed PCDL, these three laminae bound very deep postzygodiapophyseal centrodiapophyseal fossae (POCDF) on all the preserved vertebrae (). The anterior-ward depths of the fossae are best exposed on the right side of cervical vertebrae seven and eight, from which the cervical ribs and zygodiapophyseal laminae have been broken away (). The intrapostzygapophyseal laminae (TPOL) dip from each side to the midline, forming a shallow V-shape in all the preserved cervical vertebrae.
The proximal part of each transverse process projects laterally, then the distal end angles ventrally to meet the cervical rib (, ). The transverse processes are all aligned at approximately the same level across the vertebral series (A, B). The breadth of the cervical transverse processes increases posteriorly along the cervical vertebrae, resulting in broad diamond-shaped profiles to the zygodiapophyseal tables (Wilson et al. Citation2011) of the posterior vertebrae in dorsal and ventral views (, ). There is a posterior projection or diapophyseal process at the distal downturn of each transverse process in cervical vertebrae 6 to 12. These posterior processes protrude well beyond the edge of the PODL, closely approaching the prezygapophyses of the following vertebrae.
The massive neural spines on the cervical vertebra collectively constitute perhaps the most visually distinctive feature of the cervical vertebral series. The neural spines are tall and triangular in lateral view (). They are anteroposteriorly broad in cervical vertebrae 6 to 12, increasing in anteroposterior length to vertebra 12 then rapidly decreasing in anteroposterior dimension in the thirteenth and fourteenth cervical vertebrae. The neural spines also increase in height posteriorly down the series, as measured from the dorsal surface of the TPRL to the dorsal tip of the spine. The exception to this pattern is cervical 12, which has damage to the dorsal end of the neural spine, making it appear shorter than that of vertebra 11. The neural spines angle slightly posterodorsally in vertebrae 6 to 9, becoming more vertically oriented down the series to the tenth vertebra. In contrast, the neural spines of vertebrae 14 to 11 angle slightly anterodorsally, becoming more vertical from posterior to anterior in the series. The neural spine of cervical vertebra 10 is almost symmetrically triangular and its axis is dorsally directed. In this regard, cervical vertebra 10 is reminiscent of the anticlinal vertebra present in the thoracic vertebral column of some mammalian taxa.
The proportions of the tall, triangular neural spines and arches of BIBE 45854 are very different from the same elements of TMM 43621-1, an immature titanosaur referred to Alamosaurus from Big Bend National Park (Lehman & Coulson Citation2002). The neural spines and arches of TMM 43621-1 are proportionally low, resembling the condition in titanosaurs such as Malawisaurus and Trigonosaurus (Lehman & Coulson Citation2002; Campos et al. Citation2005; Gomani Citation2005). The differences in cervical neural arch and neural spine proportions between BIBE 45854 and TMM 43621-1 may be explained in part by taphonomic factors. As reported, TMM 43621-1 was preserved in fine-grained mudstone, and most of the pieces of the specimen suffered post-burial compaction and distortion (Lehman & Coulson Citation2002). It is possible, if not likely, that the dorsoventrally low proportions of the cervical neural arches and spines in TMM 43621-1 are the result of post-burial deformation of the specimens.
Thick spinoprezygapophyseal laminae (SPRL) rise from the dorsal edge of the prezygapophyses and angle posterodorsally to meet the neural spine, with the exception of the laminae of vertebrae 13 and 14, which are directed dorsally. The laminae terminate on the anterior face of the spine approximately two-thirds to three-quarters of the distance between the TPRL and the dorsal tip of the spine. The SPRL of vertebrae 8 to 12 bow anteriorly, giving those neural spines an anteriorly convex profile in lateral view (). Between the laminae is a deep spinoprezygopophyseal fossa (SPRF) on the anterior surface of each neural spine (). The fossae are deepest at the base of the spine and rapidly shallow dorsally. There are low prespinal laminae (PRSL) within the SPRF of some vertebrae (B), but the depth of the fossae, narrow space and preservational factors made it difficult to fully expose and ascertain the presence of PRSL in all vertebrae of BIBE 45854.
The spinopostzygapophyseal laminae (SPOL) are even larger than the SPRL, originating from the dorsal surface of the postzygapophyses and epipophyses. The laminae converge at a dorsally high point on the neural spine, forming the lateral boundaries of very broad and exceptionally deep spinopostzygapophyseal fossae (SPOF). In lateral view the margins of the SPOL vary from being posteriorly concave in cervical vertebrae 7, 9, 13 and 14, to a more sinuous border with distinct posterior convexities as seen in cervical vertebrae 8, 10, 11 and 12 ().
The base of each neural spine is excavated laterally by a deep spinodiapophyseal fossa (SDF) (C). The fossae closely approach each on the midline of the neural spine base, as illustrated in . The cervical neural spines of most titanosaur cervical vertebrae have some degree of an SDF, but it is usually much shallower than those present in BIBE 45854. Deep SDF similar to those in BIBE 45854 are also present in Futalognkosaurus dukei and Mendozasaurus neguyelap (Calvo et al. Citation2007b; González Riga Citation2005).
The neural spines are laterally expanded dorsal to the SDF, beginning at approximately the eighth and ninth cervical vertebrae. The lateral expansion of the neural spine is especially pronounced in the posterior vertebrae, to the degree that the neural spines dorsal to the SDF are wider than the centrum (). The lateral expansions of the neural spines are not derived from either the SPRL or SPOL, such as is the case in Bonitasaura salgadoi and Ligabuesaurus leanzai (Bonaparte et al. Citation2006; Gallina Citation2011). Instead, the lateral expansion is independent of the spinal laminae. Similar lateral expansion of the posterior cervical neural spines is also present in Futalognkosaurus dukei, Mendozasaurus neguyelap, Puertasaurus reuilli and, to a lesser degree, Quetecsaurus rusconii (González Riga Citation2003, Citation2005; Novas et al. Citation2005; Calvo et al. Citation2007b; González Riga & Ortiz Citation2014).
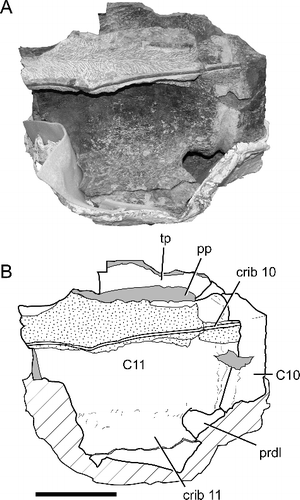
The cervical ribs are fully fused to their respective diapophyses and parapophyses in all the cervical vertebrae of BIBE 45854, forming a strong bony loop (= ansa costotransversaria; Baumel et al. Citation1993; Wedel & Sanders Citation2002) (, ). The ribs of cervical vertebrae 8 to 13 have long anterior processes, although the processes become shorter the further posteriorly in the series. The cervical rib of vertebra 14 does not have an anterior process. The anterior ends of the ribs on vertebrae 6 and 7 are broken away, although we suspect they also bore anterior processes. The medial surfaces of the ribs between the tuberculum and capitulum have complex textures and small foramina such as those visible on many surfaces of the vertebrae; however, there are no clearly defined pneumatic foramina. The cervical ribs become rapidly narrower posteriorly, tapering to thin, ovoid broken ends. During preparation, sections of long, thin (approximately 1.5 cm to 2.5 cm diameter), coarsely fibrous, overlapping cervical rib shafts were found paralleling some of the vertebral centra. One of these thin rib shafts was traced across three separate vertebra-bearing blocks, starting from the body of the right tenth cervical rib and continuing posteriorly at least as far as cervical vertebra 12. A long section of this rib shaft ventral to cervical vertebra 11 is shown in , prior to its removal during preparation of the vertebra. This section of the elongate cervical rib shaft is preserved in the Perot Museum collections. Cervical rib 14 does not appear to have a posteriorly directed rib shaft (, ).
Figure 9. BIBE 45854, Alamosaurus sanjuanensis A, cervical vertebra 11 in ventral view at an incomplete stage of preparation; segment of thin, right cervical rib 10 visible embedded in sediment ventral to centrum. B, interpretive line drawing of view in A; solid grey fill indicates broken bone surface; stippling indicates rock; diagonal cross-hatching indicates temporary support jacket constructed during preparation. Abbreviations: C10, posterior rom of centrum of cervical vertebra 10; C11, cervical vertebra 11; crib10, part of shaft of right cervical rib 10; crib 11, main body of left cervical rib 11; pp, parapophysis; prdl, prezygodiapophyseal lamina; tp, transverse process. Scale bar = 20 cm.
Discussion
Can BIBE 45854 be referred to Alamosaurus?
The holotype of Alamosaurus sanjuanensis consists of a single scapula, and an ischium collected nearby was designated the paratype (Gilmore 1922). As a result, there has been some difficulty in assigning subsequent sauropod elements collected from contemporaneous sediments in south-western North America. Gilmore (Citation1946) was certain of the affinities of the partial skeleton described from the North Horn Formation of Utah (USNM 15560), assigning it to Alamosaurus with little reservation based on similarities between the scapulae and ischia. Many subsequent works have followed in that vein, considering almost all Maastrichtian sauropod remains from south-western North America to be Alamosaurus (Lucas & Hunt Citation1989; Sullivan & Lucas Citation2000; Lehman & Coulson Citation2002; Woodward & Lehman Citation2009; Carrano & D'Emic Citation2015). Others have cast doubt on whether the limited holotype and paratype specimens possessed any diagnostic autapomorphies, and have hesitated to assign some Maastrichtian sauropod specimens in the region to the taxon (Lucas & Sullivan Citation2000; Sullivan & Lucas Citation2000; Fronimos & Lehman Citation2014; Wick & Lehman Citation2014). However, the case was recently made (D'Emic et al. Citation2011) that the holotype of Alamosaurus bears diagnosable characters and that these characters link the holotype to the specimen from the North Horn Formation of Utah described by Gilmore (Citation1946).
BIBE 45854 does not share elements with the holotype, paratype or North Horn specimens. The obvious question is, can the specimen be assigned to Alamosaurus sanjuanensis? This question is especially pertinent because there are differences in cervical vertebral proportions and other details between BIBE 45854 and an immature individual from Big Bend National Park previously identified as Alamosaurus (Lehman & Coulson Citation2002). Luckily, evidence preserved in other specimens from Big Bend National Park establishes a series of links that allows referral of BIBE 45854 to A. sanjuanensis. These specimens include a partial skeleton of a large individual (TMM 41541-1) found several kilometres west of BIBE 45854 in the uppermost part of the Javelina Formation, and disarticulated remains from small, immature titanosaur individuals at a site only 500 m away and at approximately the same horizon as BIBE 45854 within the Black Peaks Formation.
As part of the project to create a reconstruction of a complete Alamosaurus skeleton for display at the Perot Museum of Nature and Science, an agreement was made between the Museum and the Vertebrate Paleontology Laboratory at the University of Texas at Austin to use a partial skeleton of another large titanosaur from a different locality in Big Bend National Park. This skeleton (TMM 41541-1) was found by Wann Langston, Jr. in 1972, and collected under his supervision in 1973. When found, the specimen preserved two incomplete posterior cervical vertebrae, an articulated and nearly complete series of dorsal vertebrae with associated dorsal ribs, a sacrum, the first caudal vertebra, parts of both ilia, ischia and pubes, a humerus, a femur and some unidentifiable bone pieces. There are no duplicated elements in the specimen, and based on field notes and maps this specimen almost certainly represents much of a single individual. Most of the bones remained in field jackets until 2011, when arrangements for their preparation were made. The elements of TMM 41541-1 were prepped to varying degrees as necessary for laser scanning, digital 3D modelling, and replication by Research Casting International of Trenton, Ontario, Canada. The specimen was returned to The University of Texas at Austin in the summer of 2014.
TMM 41541-1 shares several similarities with the paratype (USNM 10487) and North Horn Formation (USNM 15560) specimens of Alamosaurus sanjuanensis. The first caudal vertebra of TMM 41541-1 has a bi-convex centrum (A–C), as does USNM 15560 (D, E) (Gilmore Citation1946). A biconvex first caudal centrum is also described in a few other titanosaurs, including Pellegrinisaurus powelli, Baurutitan britoi, Dreadnoughtus schrani and Neuquensaurus australis, although the biconvex vertebra of the latter was recently re-identified as a seventh sacral vertebra (Salgado Citation1996; Kellner et al. Citation2005; D'Emic & Wilson Citation2011; Lacovara et al. Citation2014). The first caudal centrum in TMM 41541-1 is anteroposteriorly short, such that the centrum might have been only as long as tall (B). In this regard it resembles the first caudal of Pellegrinisaurus more than the more elongate first caudal of USNM 15560 (E) (Gilmore Citation1946). The ventral sections of the first caudal's articular condyles of TMM 41541-1 are reconstructed in plaster. As a result, it cannot be determined whether there is a circumferential depression of the ventral rim of the centrum, a feature cited as part of a uniquely diagnostic character set shared by other specimens referred to A. sanjuanensis by D'Emic et al. (Citation2011). The transverse processes on the first caudal of TMM 41541-1 are not backswept, but rather are oriented in almost a transverse plane through the vertebra (B). This also differs from the first caudal of USNM 15560 (E). The distal end of the neural spine is missing in TMM 41541-1, but it still has pronounced PRSL and postspinal laminae (POSL), and broadens distally as in USNM 15560 (A–D).
Figure 10. Alamosaurus sanjuanensis, first caudal vertebra. A–C, TMM 41541-1 first caudal vertebra in A, anterior, B, right lateral, and C, posterior views. D, E, USNM 15560, line drawings of first caudal vertebra in D, anterior, and E, right lateral views. F, TMM 41541-1, close-up of first caudal centrum in right lateroventral and slightly anterior view; arrows point to at least three foramina ventral to transverse process. D and E drawn from Gilmore (Citation1946). Abbreviations: for, foramen; nc, neural canal; ns, neural spine; posl, postspinal lamina; prsl, prespinal lamina; poz, postzygapophysis; prz, prezygapophysis; sprl, spinoprezygapophyseal lamina; tp, transverse process. A–C, scale bar = 20 cm.
Wilson (Citation2002) identified another autapomorphy of Alamosaurus sanjuanensis, anterior and middle caudal vertebrae with several foramina opening at the base of the transverse process. These foramina are present in USNM 15560 and in another biconvex first caudal vertebra from the North Horn Formation (BYU 11392), which was used in part to justify referral of the latter to Alamosaurus sanjuanensis (D'Emic et al. Citation2011). The right side of the first caudal centrum of TMM 41541-1 is pierced by at least three foramina ventral to the transverse process (B, F). The presence of this feature in TMM 41541-1 is evidence that this individual can also be referred to Alamosaurus sanjuanensis.
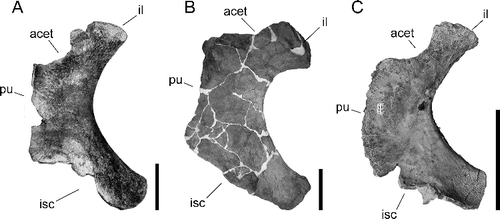
The ischium in TMM 41541-1 closely resembles the paratype ischium for Alamosaurus sanjuanensis, USNM 10487 (A, B). The short and broad ischial shaft, deeply concave posterior margin, and long pubic and ischial contacts are all similarities shared between the two specimens. The iliac peduncle is shorter in TMM 41541-1, but it appears this may be the result of post-depositional deformation. The same morphology is also shared with the articulated ischia of USNM 15560 (Gilmore 1922, Citation1946), as well as with an ischium pertaining to a small, immature individual (TMM 43621-1) from another site in Big Bend National Park (Lehman & Coulson Citation2002). No autapomorphies of Alamosaurus have been identified in the ischium, but the morphological similarity of the ischia of these specimens supports their assignment to the same taxon.
Figure 11. Right ischia of Alamosaurus sanjuanensis in medial view. A, USNM 10487; B, TMM 41541-1; C, BIBE 45909. Image of USNM 10487 taken from Gilmore (Citation1922). Abbreviations: acet, acetabular border; il, iliac peduncle; isc, ischial contact edge; pu, pubic contact edge. Scale bars = 20 cm.
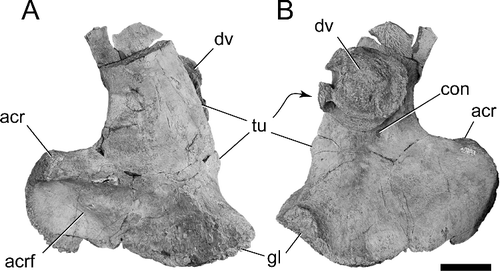
BIBE 45854 was discovered during efforts to collect fossils of relatively small, immature titanosaurs from a locality 500 m to the south, at approximately the same stratigraphical level in the Black Peaks Formation. Among the numerous elements recovered on the nearby site was a left scapula, BIBE 45958, which is missing only the distal half of the scapular blade and part of the contact for the coracoid (). The scapula matches up well with the general morphology of the holotype of Alamosaurus (USNM 10486) and the scapula of the North Horn Formation skeleton, USNM 15560 (Gilmore 1922, Citation1946). Importantly, it also preserves the two scapular autapomorphies of Alamosaurus sanjuanensis proposed by D'Emic et al. (Citation2011). These include the presence of two low tuberosities on the ventral edge of the base of scapular blade, and an asymmetrical concavity on the medial surface of the base of the scapular blade, resulting in a blade that is thicker ventrally than dorsally. Based on the presence of both characters in BIBE 45958, the specimen is referred to A. sanjuanensis. We also consider the other titanosaur bones from the site to also be remains of Alamosaurus.
Figure 12. BIBE 45958, Alamosaurus sanjuanensis. Incomplete left scapula and dorsal vertebral centrum in A, lateral, and B, medial views. Dorsal vertebral centrum is attached to medial surface of scapula. Abbreviations: acr, acromion process; acrf, acromion fossa; con, asymmetrical medial concavity; dv, dorsal vertebral centrum; gl, glenoid fossa; tu, tuberosity. Scale bar = 10 cm.
Among the other elements collected with BIBE 45958 were a pair of well-preserved ischia, BIBE 45909 (right) and BIBE 45911 (left), that may have come from the same individual. The ischia are slightly smaller than the same element in TMM 43621-1. The ischia have the same short, broad, ischial shaft, long ischial and pubic contacts, and deeply concave posterior margin seen in other Alamosaurus ischia (C). BIBE 45909 and 45911 differ slightly from the other ischia in having a more slender iliac peduncle, and an anteriorly convex pubic contact. The distal ends of the ischial shafts are relatively squared off (C), a feature in common with TMM 41541-1 (B), but differing from the more rounded off shaft in USNM 10487 (A). Overall, the differences between all these ischia can be explained by factors relating to size, ontogenetic stage, individual variation and preservation variables between specimens.
TMM 41541-1 includes parts of at least two and perhaps as many as four incomplete posterior cervical vertebrae. Comparisons can be made between two cervical vertebrae of TMM 41541-1 that have been partially prepared and the corresponding elements of BIBE 45854. One of the Austin cervical vertebrae consists mostly of just a centrum, which remains partially jacketed and upside down, exposing its ventral surface, parts of the cervical ribs, and ventral surfaces of the zygodiapophyseal table. The centrum has the same massive proportions as the centrum of cervical 13 in BIBE 45854, and the posterior width across the centrum is approximately 10–15% larger than the same element in BIBE 45854. The Austin specimen also has large, laterally overhanging PRDL that produce a nearly diamond-shaped profile to the vertebra in dorsal and ventral views, resembling the thirteenth cervical vertebra of BIBE 45854 (H, A).
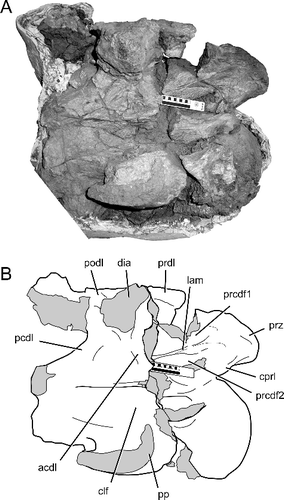
The other partially prepared cervical vertebra of TMM 41541-1 currently has only the right side partially prepared (). It has a short centrum similar in proportions to the fourteenth cervical vertebra of BIBE 45854. The right cervical rib is missing, and the parapophysis and diapophysis are broken through their bases. The broken cross section of the parapophysis shows it is anteroposteriorly short with a sharply upturned anterior end. This also matches the morphology of the parapophysis in the fourteenth cervical vertebra of BIBE 45854. The lateral surface of the centrum and the diapophyseal fossae ventral to the zygodiapophyseal table are widely exposed and visible. There is a thin lamina that arises perpendicularly from the ACDL and extends anteriorly to the base of the prezygapophysis (). The lamina divides the PRCDF into dorsal and ventral sub-fossae, just as in the thirteenth and fourteenth cervical vertebrae of BIBE 45854 (A, B). The presence of this otherwise unique vertebral character in both BIBE 45854 and TMM 41541-1 demonstrates that both specimens represent individuals of the same taxon. Given the links that tie TMM 41541-1 to Alamosaurus sanjuanensis, the case is made that BIBE 45854 is also an example of Alamosaurus sanjuanensis. BIBE 45854 is the first articulated series of cervical vertebrae described for this Maastrichtian North American titanosaur.
Figure 13. TMM 41541-1, Alamosaurus sanjuanensis. A, incomplete posterior cervical vertebra in right lateral view. B, interpretive line drawing of image in A; solid grey fill indicates broken bone surfaces. Abbreviations: acdl, anterior centrodiapophyseal lamina; clf, centrum lateral fossa; cprl, centroprezygapophyseal lamina; dia, diapophysis; lam, lamina dividing the prezygapophyseal centrodiapophyseal fossa; pcdl, posterior centrodiapophyseal lamina; podl, postzygodiapophyseal lamina; pp, parapophysis; prcdf1, dorsal prezygapophyseal centrodiapophyseal fossa; prcdf2, ventral prezygapophyseal centrodiapophyseal fossa; prdl, prezygodiapophyseal lamina; prz, prezygapophysis. Solid line in scale bar = 10 cm.
Phylogenetic analysis
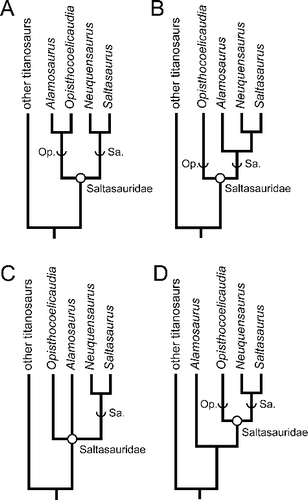
Alamosaurus sanjuanensis has been known for nearly a century, and relatively complete specimens have been known for more than half that time. Yet the phylogenetic position of the taxon has been surprisingly uncertain. A number of phylogenetic analyses have found Alamosaurus sanjuanensis to be a member of Saltasauridae (sensu Sereno Citation1998), a derived clade of mostly South American titanosaurs (Salgado et al. Citation1997; Sereno Citation1998; Wilson Citation2002; Curry Rogers Citation2005; González Riga & Ortiz David Citation2014). Some of these analyses have in turn produced hypotheses showing Alamosaurus in a sister-taxon relationship with the unusual Mongolian taxon Opisthocoelicaudia skarzynskii Borsuk-Białynicka Citation1977, or on the saltasaurid stem lineage Opisthocoelicaudiinae (A) (Sereno Citation1998; Wilson Citation2002, Citation2005; Wilson & Upchurch Citation2003; Curry Rogers Citation2005; Calvo et al. Citation2007a, Citationb; González Riga et al. Citation2009; Zaher et al. Citation2011; González Riga & Ortiz David Citation2014). Other analyses have instead placed Alamosaurus as a saltasaurine (B) (Salgado et al. Citation1997; Calvo & González Riga Citation2003; Mannion et al. Citation2013; Saegusa & Ikeda Citation2014), or in an unresolved basal polytomy within Saltasauridae (C) (Lacovara et al. Citation2014). Yet other studies have even found Alamosaurus to be a lithostrotian titanosaur outside the clade Saltasauridae (D) (Upchurch Citation1998; Upchurch et al. Citation2004; Carballido et al. Citation2011; Carballido & Sander Citation2014; Poropat et al. Citation2015). Other anatomical studies have concluded that Alamosaurus bears greater similarity to South American titanosaurs than to Opisthocoelicaudia (Lehman & Coulson Citation2002; Fronimos & Lehman Citation2014).
Figure 14. Hypotheses for the phylogenetic position of Alamosaurus sanjuanensis in previous cladistic analyses. A, Alamosaurus as the sister taxon to Opisthocoelicaudia, or on the stem lineage Opisthocoelicaudiinae (Sereno Citation1998; Curry Rogers Citation2005; Wilson Citation2002; Calvo et al. Citation2007a; González Riga et al. Citation2009; Zaher et al. Citation2011; González Riga & Ortiz David Citation2014); B, Alamosaurus as a member of stem lineage Saltasaurinae (Salgado et al. Citation1997; Calvo & González Riga Citation2003; González Riga Citation2003; Mannion et al. Citation2013; Saegusa & Ikeda Citation2014); C, Alamosaurus as a saltasaurid in a basal polytomy with Saltasaurinae and Opisthocoelicaudia (Lacovara et al. Citation2014); D, Alamosaurus as a lithostrotian titanosaur outside Saltasauridae (Upchurch Citation1998; Upchurch et al. Citation2004; Carballido et al. 2011; Carballido & Sander Citation2014; Poropat et al. Citation2015). Abbreviations: Op. = Opisthocoelicaudiinae; Sa. = Saltasaurinae.
Previous cladistic analyses of titanosaur relationships that included Alamosaurus sanjuanensis could only evaluate cervical vertebral anatomy based on a few disarticulated elements of an immature individual, TMM 43621-1 (Lehman & Coulson Citation2002). There are morphological differences between the cervical vertebrae of TMM 43621-1 and BIBE 45854, especially with regard to the relative size of the neural spines and other proportions of the neural arches and centra. The differences might be attributable to the different ontogenetic stages represented by the two individuals. It is also possible that the smaller specimens were subjected to post-burial dorsoventral deformation. BIBE 45854 and TMM 41541-1 provide the first chance to assess vertebral characters of osteologically mature individuals of Alamosaurus and contribute the data from them into cladistics analyses. In addition, a number of new titanosaurs have been described in recent years, especially from South America. The increase in taxonomic sampling has increased our awareness of the great diversity of titanosauriform sauropods and has resulted in changes in our hypotheses of titanosaur systematics.
We tested the phylogenetic position of Alamosaurus sanjuanensis in light of new morphological data preserved in BIBE 45854, TMM 41541-1, additional specimens in the collections of the Perot Museum and the Vertebrate Paleontology Laboratory of the University of Texas at Austin, and recent literature (Carrano & D'Emic Citation2015). We ran two analyses with the updated information using two previously published data sets, those of Lacovara et al. (Citation2014) and González Riga & Ortiz David (Citation2014). Both of these studies found Alamosaurus to be a member of Saltasauridae, and the latter supported a sister-taxon relationship between Alamosaurus and Opisthocoelicaudia. The purpose of running two analyses was to account for the potential impacts of markedly different taxon and character sampling in light of the new morphological data available for Alamosaurus.
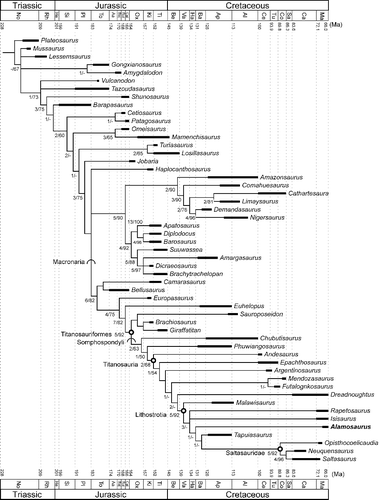
Our first analysis was based on the published data set of Lacovara et al. (Citation2014), which was in turn an elaboration of the cladistic analysis conducted by Carballido & Sander (Citation2014). The analysis of Lacovara et al. (Citation2014) included 53 taxa from across a broad array of Sauropoda and sauropodomorphs basal to Sauropoda, evaluating them for 341 characters. Antetonitrus was deleted from our analysis because it proved to be a highly labile taxon that served to greatly inflate the number of equally most parsimonious trees generated by the analysis. Twenty-three characters were re-evaluated and scored differently for Alamosaurus based on information preserved in the new specimens described here. We revised a small number of characters for Opisthocoelicaudia, and re-wrote one character to reflect variation in the superficially similar but not homologous lateral expansions of the posterior cervical vertebral neural spines in some taxa. A list of characters with their new character state scores is given in the Supplemental Material, as is the taxon-character list and NEXUS file used for this analysis.
A heuristic parsimony analysis was conducted using PAUP* (Swofford Citation2002), which produced 40 equally most parsimonious trees, each with a length of 958 evolutionary steps, a consistency index (CI) of 0.4238, and a retention index (RI) of 0.7072. The strict consensus of the 40 equally most parsimonious trees is shown in . Our strict consensus tree is generally similar to that depicted by Lacovara et al. (Citation2014), and we restrict our discussion to the macronarian branch of the resulting hypotheses. The macronarian branch of our strict consensus differs from that of Lacovara et al. (Citation2014) in only two ways. Epachthosaurus and Argentinosaurus branch from successively more derived nodes within Titanosauria in our results, in contrast to Lacovara et al. (Citation2014) in which these taxa were in a polytomy with the lineage leading to Lognkosauria, Dreadnoughtus and Lithostrotia. The other notable difference is that Alamosaurus is not a member of Saltasauridae, contrary to the findings of Lacovara et al. (Citation2014). Instead, we find Alamosaurus to be the sister taxon to a clade comprised Tapuiasaurus and Saltasauridae. This position for Alamosaurus is generally consistent with some previous works (D) (Upchurch Citation1998; Upchurch et al. Citation2004; Carballido et al. Citation2011; Carballido & Sander Citation2014; Poropat et al. Citation2015).
Figure 15. Results of first phylogenetic analysis based on data set of Lacovara et al. (Citation2014). Strict consensus of 40 equally most parsimonious trees (length = 958 evolutionary steps; consistency index = 0.4238, retention index = 0.7072) set upon a geological timescale (Gradstein et al. Citation2012). Thickened branch sections indicate approximate temporal range for each terminal taxon. Temporal ranges of taxa were taken from multiple sources (Borsuk-Bialynicka Citation1977; Upchurch et al. Citation2004; Weishampel et al. Citation2004; de Jesus Faria et al. Citation2015). Open circles at nodes indicate a node-defined clade name. Arcs on branches indicate a stem-defined lineage name. See for name definitions. Numbers at nodes indicate bootstrap and decay index (Bremer value), formatted as decay index/bootstrap value. A dash (-) indicates a decay index of 0 and/or bootstrap value < 50%.
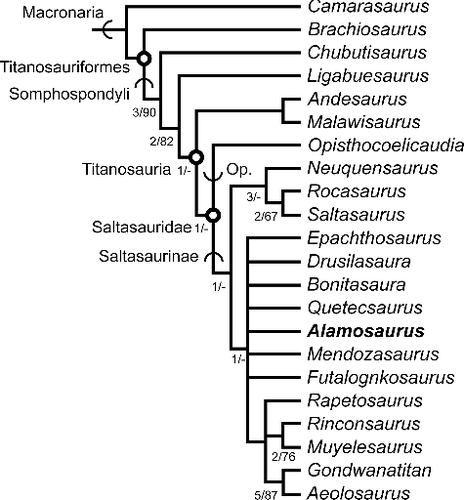
The second analysis was based on the data set of González Riga & Ortiz David (Citation2014). This data set evaluated far fewer taxa (21 ingroup taxa and Camarasaurus as the single outgroup taxon) and characters (82), but, importantly, it incorporated a number of South American titanosaurs which were not in the study by Lacovara et al. (Citation2014). These included Ligabuesaurus, Gondwanatitan, Aeolosaurus, Rinconsaurus, Muyelensaurus, Drusilasaura, Bonitasaura, Quetecsaurus and Rocasaurus. The characters evaluated in this smaller data set were also more focused on titanosaur relationships, whereas the larger Lacovara et al. (Citation2014) study was a broader, shallower investigation across many sauropodomorph clades. Alamosaurus was found to be the sister taxon to Opisthocoelicaudia within Saltasauridae by González Riga & Ortiz David (Citation2014). We coded 10 characters differently for Alamosaurus to reflect new data present in the specimens discussed here. A list of those re-coded characters is given in the Supplemental Material, as is the taxon-character list for Alamosaurus and the NEXUS file used for this analysis. After these characters were re-coded, 62 characters in the analysis were parsimony informative. All characters were treated as unordered and equally weighted. The second test was conducted as a heuristic parsimony analysis, using PAUP* (Swofford Citation2002).
The second analysis initially produced 13 equally most parsimonious trees, each with a length of 146 evolutionary steps, a CI of 0.6918 and RI of 0.7115. The strict consensus of the 13 trees () is markedly different than the findings of González Riga & Ortiz David (Citation2014). A Malawisaurus + Andesaurus clade was found to be the sister taxon to all other members of Titanosauria. Opisthocoelicaudia was placed at the next more derived node than the Andesaurus + Malawisaurus clade, a dramatically different relationship for this Mongolian taxon than in previous analyses. Alamosaurus was in a polytomous relationship with several other titanosauriforms closer to Saltasaurus. Closer examination showed that the lack of resolution in the trees was caused by the labile behaviour of the taxon Drusilasaura.
Figure 16. Results of the second analysis. Strict consensus of 14 equally most parsimonious trees (length = 146 evolutionary steps, consistency index = 0.6918, retention index = 0.7115) generated from analysis based on data set of González Riga & Ortiz David (Citation2014). Open circles at nodes indicate a node-defined clade name. Arcs on branches indicate a stem-defined lineage name. Op. = Opisthocoelicaudiinae. Numbers at nodes indicate bootstrap and decay index (Bremer value), formatted as decay index/bootstrap value. A dash (-) indicates a decay index of 0 and/or bootstrap value <50%.
The data set was analysed again, but this time changed by deleting Drusilasaura from consideration. The analysis resulted in a single, fully resolved tree with a length of 145 steps, a CI of 0.6966, and an RI of 0.7124 (). The positions of Andesaurus, Malawisaurus and Opisthocoelicaudia were the same as before. In this tree the node-clade Saltasauridae sensu Sereno (Citation1998), which is comprised of the stem-sister lineages Opisthocoelicaudiinae and Saltasaurinae, is the sister taxon to the Andesaurus + Malawisaurus clade. Saltasauridae therefore encompasses all titanosaurs in the analysis other than Andesaurus and Malawisaurus, a taxonomic expansion of the clade compared to earlier analyses. The analysis did recover a Neuquensaurus + (Rocasaurus + Saltasaurus) clade, which matches the content of the prior node-based Saltasaurinae sensu Salgado et al. (Citation1997).
Figure 17. Results of the second analysis. Single resolved tree (length = 145 evolutionary steps, consistency index = 0.6966, retention index = 0.7124) generated by the second analysis based on data matrix of González Riga & Ortiz David (Citation2014) after Drusilasaura was deleted from the analysis, and set on a geological time scale (Gradstein et al. Citation2012). Thickened branch sections indicate approximate temporal range for each terminal taxon. Temporal ranges of taxa were taken from multiple sources (Borsuk-Bialynicka Citation1977; Upchurch et al. Citation2004; Weishampel et al. Citation2004; de Jesus Faria et al. Citation2015). Open circles at nodes indicate a node-defined clade name. Arcs on branches indicate a stem-defined lineage name. See for name definitions. Lo. = Lognkosauria. Op. = Opisthocoelicaudiinae. Numbers at nodes indicate bootstrap and decay index (Bremer value), formatted as decay index/bootstrap value. A dash (-) indicates a decay index of 0 and/or bootstrap value <50%.
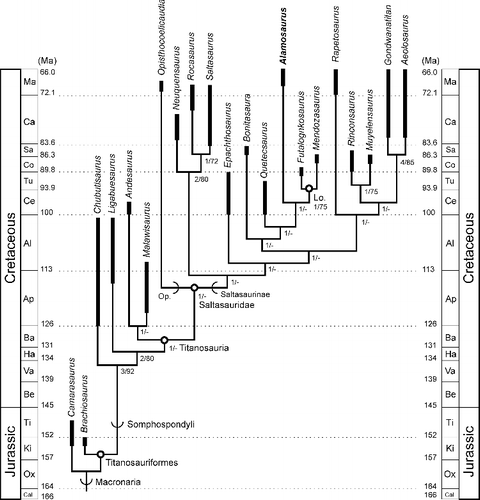
The position of Alamosaurus was an unexpected result. The analysis recovered Alamosaurus as the sister taxon to Lognkosauria, a departure from previous studies. The Alamosaurus + Lognkosauria clade is supported by three unequivocal synapomorphies: the presence of laterally expanded cervical neural spines originated by lateral laminae that reach or surpass the width of the centra; posterior cervical vertebrae with a total height versus centrum length ratio greater than 1.5; and deep and extended spinodiapophyseal fossae in the posterior cervical vertebrae. It should be noted that only the most posterior cervical vertebra in BIBE 45854 is 1.5 times taller than the length of the centrum. Under accelerated transformation (ACCTRAN) character state optimization, an equivocal character that may diagnose the clade is the presence of a sub oval-shaped coracoid. This stands in contrast to the distinctly quadrangular coracoid present in Neuquensaurus, Saltasaurus, Rinconsaurus, Opisthocoelicaudia and Quetecsaurus. However, this feature is not preserved in Mendozasaurus and Futalognkosaurus, and a rounded coracoid may instead prove to be independently diagnostic of Alamosaurus and Rapetosaurus in the results recovered here.
The next more basal node on the tree includes Quetecsaurus rusconii, and is supported by one unequivocal and one equivocal character under ACCTRAN character state optimization. The unequivocal synapomorphy is the presence of lateral laminae on the neural spines of the posterior cervical vertebrae. The lateral laminae in Quetecsaurus are not expanded to the extent seen in Alamosaurus and the lognkosaurs, but they are more pronounced than in most other titanosaurs (González Riga & Ortiz David Citation2014). The equivocal character is the condition of having sternal plates with a straight posterior border, a morphology also present in Malawisaurus (Gomani Citation2005).
The position of Opisthocoelicaudia was a result contrary to most previous cladistic studies of titanosaurs. However, support for this and other nodes in the hypothesis of phylogeny depicted in is not strong. The recovered tree was manipulated in MacClade (Madison & Madison Citation2005) by placing Alamosaurus and Opisthocoelicaudia as sister taxa on a branch just outside the Neuquensaurus + (Saltasaurus + Rocasaurus) clade. This increased tree length by only two steps. Returning Alamosaurus as the sister taxon to Lognkosauria, it required only one additional step to keep Opisthocoelicaudia as the sister taxon to the Neuquensaurus + (Saltasaurus + Rocasaurus) clade. The low support for the recovered tree is reflected in the weak Bremer indices at most nodes, as well as less than 50% bootstrap support at most nodes (). Even so, the fact that an analysis with a narrow focus on titanosaur relationships to the exclusion of non-macronarian taxa produced such a novel hypothesis for the relationships of Alamosaurus should prompt further investigations with similar taxonomic sampling and greater emphasis on details of titanosaur anatomy. It also illustrates the importance of specimens such as BIBE 45854, which bring new data into analyses and introduce new and unexpected directions for further study.
Palaeobiogeographical implications
The presence of Alamosaurus sanjuanensis in the Maastrichtian deposits of southern Laramidia has long attracted the attention of vertebrate palaeontologists. The appearance of at least one sauropod taxon in the North American record after an apparent absence of tens of millions of years marks a notable faunal event on the continent. Indeed, the informal term ‘Alamosaurus fauna’ has been used to identify the assemblage of large vertebrate taxa that inhabited the southern part of the Laramidian landmass, contributing to increasing evidence of faunal provincialism across Laramidia during Campanian and Maastrichtian time (Lehman Citation2001; Sampson & Loewen Citation2010; Sampson et al. Citation2010; Loewen et al. Citation2013; Fiorillo & Tykoski Citation2014).
A question that lingers is, from where did Alamosaurus or its immediate ancestors come? The nature of the debate was recently summarized by Fronimos & Lehman (Citation2014), who listed the various assertions of relationships and potential origins for Alamosaurus made over the preceding two decades. Was the so-called ‘sauropod hiatus’ (Lucas & Hunt Citation1989) a period when sauropods were actually absent from North America, to be ended via immigration from adjacent landmasses during the Campanian–Maastrichtian (D'Emic & Foreman Citation2012)? Or is the perceived hiatus an artefact of preservation and sampling, with sauropods inhabiting locations inland away from depositional environments during the Late Cretaceous of North America (Lehman Citation2001; Mannion & Upchurch Citation2011)? D'Emic et al. (Citation2010) held the opinion that the question of how sauropods came to appear in the Maastrichtian record of Laramidia required better understanding of the lower level phylogenetic affinities of Alamosaurus. Do the hypotheses produced by the phylogenetic analyses here offer new information pertinent to this problem?
Phylogenetic analyses that recover Alamosaurus and Opisthocoelicaudia as closely related saltasaurids, sometimes even as sister taxa, require certain events and conditions to explain the presence of Alamosaurus in southern Laramidia and Opisthocoelicaudia in central Asia during the Maastrichtian. The first is that whether saltasaurids originated in Asia and dispersed to South American or they originated in South America and made their way to central Asia, they had to have traversed and inhabited the entire latitudinal breadth of the Laramidian landmass in the process. Second, under that scenario saltasaurids must have done so without leaving evidence of their presence in Campanian and Maastrichtian sediments across the middle and higher latitude regions of Laramidia. The absence of sauropod remains in these sediments is problematic, because the richest, most-sampled dinosaur faunas on the continent are known from these deposits. Even the Beringian dispersal route between Asia and Laramidia has a growing record of body and trace fossils of dinosaurs and other terrestrial vertebrates of this age (Gangloff et al. Citation2005; Fiorillo Citation2008; Fiorillo & Adams Citation2012; Fiorillo & Tykoski Citation2012, Citation2014; Fiorillo et al Citation2014). To date, no trace of sauropods has been found in the Beringian corridor connecting the two continents.
The results from the two phylogenetic analyses conducted here indicate that either: (1) Alamosaurus is not a saltasaurid titanosaur, but rather is a lithostrotian titanosaur outside Saltasauridae; or (2) Alamosaurus is closely allied to Lognkosauria on the saltasaurine branch of a greatly expanded Saltasauridae. Our analysis based upon the data set of Lacovara et al. (Citation2014) found Alamosaurus to branch from a node more derived than Rapetosaurus and Isisaurus from the Maastrichtian of Madagascar and India, respectively, and basal to Tapuiasuarus + Saltasauridae from the Early Cretaceous to Maastrichtian of South America (). This could indicate that Alamosaurus was descended from an earlier, more widespread distribution of Gondwanan lithostrotian titanosaurs. Alamosaurus would then represent either a Maastrichtian introduction of non-saltasaurid lithostrotians from South America, or the last of a long-lived lineage of Laramidian non-saltasaurid lithostrotians that somehow immigrated to and inhabited the continent since at least earlier in the Late Cretaceous. In either case, the presence of Alamosaurus in southern Laramidia during the Maastrichtian would contribute little to answering the question of how saltasaurid titanosaurs ranged from South America to central Asia without leaving a North American record.
In contrast to the results produced by the analysis based on the data set of Lacovara et al. (Citation2014), the phylogenetic analysis based upon the data set of González Riga & Ortiz David (Citation2014) produced novel and unexpected relationships within Titanosauria, which offers a different set of possibilities to explain the distribution of saltasaurid titanosaurs. Under this hypothesis of phylogeny the split between opisthocoelicaudine and saltasaurine saltasaurids occurred relatively early in the evolution of titanosaurs, facilitating a nearly global distribution of saltasaurids via connections between landmasses prior to full continental breakups. Opisthocoelicaudia would represent an Asian member of a long-diverged branch of saltasaurids, eliminating the need for complex explanations to account for the apparent lack of a Late Cretaceous fossil record of saltasaurids in Laramidia linking closely related central Asian and South American taxa. Also in this hypothesis of phylogeny, Alamosaurus is on the branch leading to Lognkosauria, titanosaurs known from the Turonian to Coniacian of South America.
A northward dispersal to southern Laramidia by members of that South American titanosaur lineage in the late Campanian to early Maastrichtian would match the evidence preserved in the North American fossil record. This scenario is supported by the anatomical analyses of Lehman & Coulson (Citation2002) and Fronimos & Lehman (Citation2014), which found more similarities between titanosaur fossils from Big Bend National Park and South America than with Opisthocoelicaudia. There is evidence for at least intermittent land connections between southern Laramidia and northern South America during the Campanian and perhaps through to the Paleocene, which correlates to the appearance of Laurasian taxa such as marsupial mammals and hadrosaurid dinosaurs appearing in South America in the Campanian–Maastrichtian (Iturralde-Vinent Citation2006; Ortiz-Jaureguizar & Pascual Citation2011). A recent phylogenetic study found a clade comprised of the hadrosaurids Secernosaurus koerneri and an un-named taxon from the Chubut and Rio Negro provinces of Argentina to be the sister taxon of a yet-to-be-named hadrosaurid taxon from Big Bend National Park, Texas (Prieto-Marquez & Salinas Citation2010). Whatever the exact nature of the connection between Laramidia and South America at the time, it was sufficient to facilitate the exchange of large herbivorous dinosaurs between the landmasses.
Conclusions
The articulated cervical vertebral series BIBE 45854 from Big Bend National Park, Texas, can be referred to the titanosaur Alamosaurus sanjuanensis on the basis of shared morphological characters linking it to TMM 41541-1, another specimen from Big Bend National Park. TMM 41541-1 can in turn be linked to other specimens of A. sanjuanensis, including remains of immature individuals found a few hundred metres from BIBE 45854 that can be referred to Alamosaurus, USNM 15560 from the North Horn Formation of Utah, and the holotype (USNM 10486) and paratype (USNM 10487) specimens of A. sanjuanensis from the San Juan Basin of New Mexico. BIBE 45854 provides the first thorough view of the cervical vertebral osteology of a large, mature Alamosaurus sanjuanensis. Vertebral characters are heavily relied upon in sauropod systematics, and the morphological data preserved in the new specimens described here have much to contribute to our understandings of the relationships and biology of Alamosaurus. BIBE 45854 also provides the first evidence of a new autapomorphy for A. sanjuanensis, the posterior-most two cervical vertebrae with the PRCDF divided into dorsal and ventral sub-fossae by a thin lamina. This autapomorphy was subsequently verified in TMM 41541-1. The cervical vertebrae of Alamosaurus more closely resemble those of non-saltasaurid titanosaurs such as Dreadnoughtus, Trigonosaurus, Bonitasaura and Futalognkosaurus rather than the cervical vertebrae of saltasaurid titanosaurs such as Neuquensaurus and Saltasaurus.
Two cladistic phylogenetic analyses based on previously published data sets were conducted, with Alamosaurus re-scored in each to reflect new data preserved in BIBE 45854 and TMM 41541-1. The original data sets were selected because they varied greatly in terms of their taxonomic and character content. The first (Lacovara et al. Citation2014) was a broad but relatively shallow analysis across a large number of sauropod clades. The second (González Riga & Ortiz David Citation2014) lacked the number of taxa and characters present in the first data set, but was more focused on titanosaur inter-relationships. Our first analysis produced hypotheses of phylogeny that found Alamosaurus to be a lithostrotian titanosaur outside Saltasauridae, a result generally consistent with some previous studies (Upchurch Citation1998; Upchurch et al. Citation2004; Carballido et al. 2011; Carballido & Sander Citation2014; Poropat et al. 2014). The difference between our results and those of Lacovara et al. (Citation2014) reflects the impact of new data preserved in BIBE 45854, TMM 41541-1, and other specimens of Alamosaurus from Big Bend National Park.
The results of the second test produced a novel hypothesis of phylogeny with regard to Alamosaurus and greatly expanded the taxonomic content of Saltasauridae. Alamosaurus was found to be the sister taxon to Lognkosauria, a recently recognized clade of giant, thick-necked, South American titanosaurs including Futalognkosaurus dukei and Mendozasaurus neguyelap. The synapomorphies linking Alamosaurus with lognkosaurs are all based on cervical vertebral morphology, and include: the presence of laterally expanded cervical neural spines originated by lateral laminae that reach or surpass the width of the centra; posterior cervical vertebrae with a total height versus centrum length ratio greater than 1.5; and deep and extended SDF in the posterior cervical vertebrae. The results of these analyses highlight the important role of vertebral characters in cladistic analyses of sauropod phylogeny. It also serves as a reminder that hypotheses of titanosaur phylogeny may yet be susceptible to substantial revision as new taxa are discovered or data from new specimens such as BIBE 45854 and TMM 41541-1 alter old assumptions about long-known taxa.
Supplemental material
Supplemental material for this article can be accessed here: http://dx.doi.org/10.1080/14772019.2016.1183150.
Suppl_Figure_17_Colour.tif
Download TIFF Image (3.3 MB)Suppl_Figure_15_Colour.tif
Download TIFF Image (4.1 MB)Suppl_Figure_3_Colour.tif
Download TIFF Image (14.3 MB)Suppl_Figure_2_Colour.tif
Download TIFF Image (1.8 MB)Tykoski_Fiorillo_Test2.nex
Download (11.1 KB)Tykoski_Fiorillo_Test1.nex
Download (30.6 KB)Tykoski_Fiorillo_Supplementary_Online_Material.docx
Download MS Word (21.8 KB)Acknowledgements
We thank the numerous volunteers who worked on the preparation of these materials in the Paleontology Laboratory at the Perot Museum of Nature and Science. We especially recognize the efforts of Perot Museum personnel Tommy Diamond and Marcie Keller, who prepared out large sections of BIBE 45854. We also thank the staff of the University of Texas at Dallas Department of Science/Mathematics Education Department, particularly Homer Montgomery and Fred Fifer, for their cooperation and assistance in the various phases of this work that led to the successful excavation of this specimen. We thank Tim Rowe, Wann Langston Jr., Matt Brown and Chris Sagebiel for agreeing to let us use and study specimens in the collections of the Vertebrate Paleontology Laboratory of the University of Texas at Austin. We are also extremely grateful to the staff at Big Bend National Park, especially Don Corrick, for their efforts in facilitating this excavation and their continued interest in the project. Finally, we thank Michael D'Emic and Philip Mannion, whose reviews greatly improved this manuscript.
Related Research Data
References
- Baumel, J. J., King, A. S., Breazile, J. E., Evans, H. E. & Vanden Berge, J. C. 1993. Handbook of avian anatomy: Nomina Anatomica Avium. 2nd edition. Publications of the Nuttall Ornithological Club, Cambridge.
- Bonaparte, J. F. & Coria, R. 1993. Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albaniano-Cenomaniano) de la Provincia Neuquén, Argentina. Ameghiniana, 30, 271–282.
- Bonaparte, J. F., Gonzalez Riga, B. J. & Apesteguia, S. 2006. Ligabuesaurus leanzai gen. et sp. nov. (Dinosauria, Sauropoda), a new titanosaur from the Lohan Cura Formation (Aptian, Lower Cretaceous) of Neuquén, Patagonia, Argentina. Cretaceous Research, 27, 364–376.
- Borsuk-Białynicka, M. 1977. A new camarasaurid sauropod Opisthocoelicaudia skarzynskii gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontologia Polonica, 37, 1–64.
- Brochu, C. A. 1996. Closure of neurocentral sutures during crocodilian ontogeny: implications for maturity assessment in fossil archosaurs. Journal of Vertebrate Paleontology, 16, 49–62.
- Calvo, J. O. & González Riga, B. J. 2003. Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. Revista Geológica de Chile, 30, 333–353.
- Calvo, J. O., Porfiri, J. D., González Riga, B. J. & Kellner, A. W. A. 2007a. A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur. Anais da Academia Brasiliera de Ciências, 79, 529–541.
- Calvo, J. O., Porfiri, J. D., González Riga, B. J. & Kellner, A. W. A. 2007b. Anatomy of Futalognkosaurus dukei Calvo, Porfiri, González Riga, and Kellner, 2007 (Dinosauria, Titanosauridae) from the Neuquén Group (Late Cretaceous), Patagonia, Argentina. Arquivos do Museu Nacional, Rio de Janeiro, 65, 511–526.
- Campos, D. A., Kellner, A. W. A., Bertini, R. J. & Santucci, R. M. 2005. On a titanosaurid (Dinosauria, Sauropoda) vertebral column from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museu Nacional, Rio de Janeiro, 63, 565–593.
- Carballido, J. L., Rauhut, O. W. M., Pol, D. & Salgado, L. 2011. Osteology and phylogenetic relationships of Tehuelchesaurus benitezii (Dinosauria, Sauropoda) from the Upper Jurassic of Patagonia. Zoological Journal of the Linnean Society, 163, 605–662.
- Carballido, J. L. & Sander, P. M. 2014. Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria. Journal of Systematic Palaeontology, 12, 335–387.
- Carrano, M. T. & D'Emic, M. D. 2015. Osteoderms of the titanosaur sauropod dinosaur Alamosaurus sanjuanensis Gilmore, 1922. Journal of Vertebrate Paleontology, 35, e901334.
- Chure, D., Britt, B. B., Whitlock, J. A. & Wilson, J. A. 2010. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Naturwissenschaften, 97, 379–391.
- Cooper, R. W., Lehman, T. M., Stevens, J. B., Stevens, M. S. & Cooper, D. A. 2011. Geology of the McKinney Springs Quadrangle, Big Bend National Park, Texas, Sheet 2. In R. W. Cooper (ed.) Miscellaneous Map No. 50: Geologic maps of the Upper Cretaceous and Tertiary strata, Big Bend National Park, Texas. Bureau of Economic Geology, Jackson School of Geosciences, The University of Texas at Austin.
- Coria, R. A., Filippi, L. S., Chiappe, L. M. García, R. & Arcucci, A. B. 2013. Overosaurus paradasorum gen. et sp. nov., a new sauropod dinosaur (Titanosauria: Lithostrotia) from the Late Cretaceous of Neuquén, Patagonia, Argentina. Zootaxa, 3683, 357–376.
- Coulson, A. B. 1998. Sedimentology and taphonomy of a juvenile Alamosaurus site in the Javelina Formation (Upper Cretaceous), Big Bend National Park, Texas. Unpublished Masters thesis, Texas Tech University.
- Curry Rogers, K. 2005. Titanosauria. Pp. 50–103 in K. Curry Rogers & J. Wilson (eds) The sauropods: evolution and paleobiology. University of California Press, Berkeley.
- Curry Rogers, K. 2009. The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology, 29, 1046–1086.
- de Jesus Faria, C. C., González Riga, B., dos Anjos Candeiro, C. R., da Silva Marinho, T., Ortiz David, L., Mederios Simbras, F., Barboza Castahno, R., Pereira Muniz, F. & da Costa Pereira, P. V. L. G. 2015. Cretaceous sauropod diversity and taxonomic succession in South America. Journal of South American Earth Sciences, 61, 154–163.
- D'Emic, M. D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society, 166, 624–671.
- D'Emic, M. D., Wilson, J. A. & Thompson, W. 2010. The end of the sauropod hiatus in North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 297, 486–490.
- D'Emic, M. D. & Wilson, J. A. 2011. New remains attributable to the holotype of the sauropod dinosaur Neuquensaurus. Acta Palaeontologica Polonica, 56, 61–73.
- D'Emic, M. D., Wilson, J. A. & Williamson, T. E. 2011. A sauropod dinosaur pes from the latest Cretaceous of North America and the validity of Alamosaurus sanjuanensis (Sauropoda, Titanosauria). Journal of Vertebrate Paleontology, 31, 1072–1079.
- D'Emic, M. D. & Foreman, B. Z. 2012. The beginning of the sauropod hiatus in North America: insights from the Lower Cretaceous Cloverly Formation of Wyoming. Journal of Vertebrate Paleontology, 32, 883–902.
- Fiorillo, A. R. 1998. Preliminary report on a new sauropod locality in the Javelina Formation (Late Cretaceous), Big Bend National Park, Texas. Pp. 29–31 in V. Santucci and L. McClelland (eds) National Park Service Paleontological Research, Technical Report NPS/NRGRD/GRDTR-98/01. Lakewood: Geological Resource Division.
- Fiorillo, A. R. 2008. Cretaceous dinosaurs of Alaska: implications for the origins of Beringia. Geological Society of America Special Paper, 442, 313–326.
- Fiorillo, A. R. & Adams, T. L. 2012. A therizinosaur track from the Lower Cantwell Formation (Upper Cretaceous) of Denali National Park, Alaska. Palaios, 27, 395–400.
- Fiorillo, A. R. & Tykoski, R. S. 2012. A new Maastrichtian species of the centrosaurine ceratopsid Pachyrhinosaurus from the North Slope of Alaska. Acta Palaeontologica Polonica, 57, 561–573.
- Fiorillo, A. R. & Tykoski, R. S. 2014. A diminutive new tyrannosaur from the top of the world. PLoS ONE, 9, e91287.
- Fiorillo, A. R., Hasiotis, S. T. & Kobayashi, Y. 2014. Herd structure in Late Cretaceous polar dinosaurs: a remarkable new dinosaur tracksite, Denali National Park, Alaska, USA. Geology, 42, 719–722.
- Fronimos, J. A. & Lehman. T. M. 2014. New specimens of a titanosaur sauropod from the Maastrichtian of Big Bend National Park, Texas. Journal of Vertebrate Paleontology, 34, 883–899.
- Gallina, P. A. 2011. Notes on the axial skeleton of the titanosaur Bonitasaura salgadoi (Dinosauria-Sauropoda). Anais da Academia Brasileira de Ciências, 83, 235–245.
- Gangloff, R. A., Fiorillo, A. R. & Norton, D. W. 2005. The first pachycephalosaurine from the paleo-Arctic of Alaska and its paleogeographic implications. Journal of Paleontology, 79, 997–1001.
- Gilmore, C. W. 1921. Discovery of dinosaur remains in the Upper Cretaceous of New Mexico. Science, 54, 274.
- Gilmore, C. W. 1922. A new sauropod dinosaur from the Ojo Alamo Formation of New Mexico. Smithsonian Miscellaneous Collections, 72, 1–9.
- Gilmore, C. W. 1938. Sauropod dinosaur remains in the Upper Cretaceous. Science, 87, 299–300.
- Gilmore, C. W. 1946. Reptilian fauna from the North Horn Formation of central Utah. United States Geological Survey Professional Paper, 210, 29–52.
- Gomani, E. M. 2005. Sauropod dinosaurs from the Early Cretaceous of Malawi, Africa. Palaeontologia Electronica, 8, 1–37.
- González Riga, B. J. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana, 40, 155–172.
- González Riga, B. J. 2005. Nuevos restos fósiles de Mendozasaurus neguyelap (Sauropoda: Titanosauria) del Cretácico Tardío de Mendoza, Argentina. Ameghiniana, 42, 535–538.
- González Riga, B. J., Previtera, E. & Pirrone, C. A. 2009. Malarguesaurus florenciae gen. et sp. nov., a new titanosauriform (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Cretaceous Research, 30, 135–148.
- González Riga, B. J. & Ortiz David, L. 2014. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous (Cerro Lisandro Formation) of Mendoza Province, Argentina. Ameghiniana, 51, 3–25.
- Gradstein, F. M., Ogg, J. G., Schmitz, M. D. & Ogg, G. M. (eds). 2012. The geologic time scale 2012. Elsevier, Boston.
- Ikejiri, T. 2003. Sequence of closure of neurocentral sutures in Camarasaurus (Sauropoda) and implications for phylogeny in Reptilia. Journal of Vertebrate Paleontology, 23, Supplement to No. 3, 65A.
- Ikejiri, T. 2004. Anatomy of Camarasaurus lentus (Dinosauria: Sauropoda) from the Morrison Formation (Late Jurassic), Thermopolis, central Wyoming, with determination and interpretation of ontogenetic, sexual dimorphic, and individual variation in the genus. Unpublished Masters thesis, Fort Hays State University, 311 pp.
- Iturralde-Vinent, M. A. 2006. Meso–Cenozoic Caribbean paleogeography: implications for the historical biogeography of the region. International Geology Review, 48, 791–827.
- Jain, S. J. & Bandyopadhyay, S. 1997. A new titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of central India. Journal of Vertebrate Paleontology, 17, 114–136.
- Kellner, A. W. A., Campos, D. A. & Trotta, M. N. F. 2005. Description of a titanosaurid caudal series from the Bauru Group, Late Cretaceous of Brazil. Arquivos do Museu Nacional, Rio de Janeiro, 63, 529–564.
- Lacovara, K. J., Lamanna, M. C., Ibiricu, L. M., Poole, J. C., Schroeter, E. R., Ullmann, P. V., Voegele, K. K., Boles, Z. M., Carter, A. M., Fowler, E. K., Egerton, V. M., Moyer, A. E., Coughenour, C. L., Schein, J. P., Harris, J. D., Martinez, R. D. & Novas, F. E. 2014. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Nature, Scientific Reports, 4, 6196.
- Lehman, T. M. 2001. Late Cretaceous dinosaur provinciality. Pp. 310–328 in D. H. Tanke & K. Carpenter (eds) Mesozoic vertebrate life. Indiana University Press, Bloomington.
- Lehman, T. M. & Coulson, A. B. 2002. A juvenile specimen of the sauropod dinosaur Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park, Texas. Journal of Paleontology, 76, 156–172.
- Lehman, T. M., McDowell, F. W. & Connelly, J. N. 2006. First isotopic (U-Pb) age for the Late Cretaceous Alamosaurus vertebrate fauna of west Texas, and its significance as a link between two faunal provinces. Journal of Vertebrate Paleontology, 26, 922–928.
- Loewen, M. A., Irmis, R. B., Sertich, J. J. W., Currie, P. J. & Sampson, S. D. 2013. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLoS ONE, 8, e79420.
- Lucas, S. G. & Hunt, A. P. 1989. Alamosaurus and the sauropod hiatus in the Cretaceous of the North American Western Interior. Geological Society of America Special Paper, 238, 75–85.
- Lucas, S. G. & Sullivan, R. M. 2000. The sauropod dinosaur Alamosaurus from the Upper Cretaceous of the San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science Bulletin, 17, 147–156.
- Maddison D. R. & Maddison, W. P. 2005. MacClade 4. Updated at: http://macclade.org/macclade.html.
- Mannion, P. D. & Upchurch, P. 2011. A re-evaluation of the ‘mid-Cretaceous sauropod hiatus’ and the impact of uneven sampling of the fossil record on patterns of regional dinosaur extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 299, 529–540.
- Mannion, P. D., Upchurch, P., Barnes, R. N. & Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society, 168, 98–206.
- Marsh, O. C. 1878. Principal characters of American Jurassic dinosaurs: Part I. American Journal of Science (Series 3), 16, 411–416.
- Maxwell, R. A., Lonsdale, J. T., Hazzard, R. T. & Wilson, J. A. 1967. Geology of Big Bend National Park, Brewster County, Texas. The University of Texas at Austin Bureau of Economic Geology Publication 6711, 320 pp.
- Maxwell, R. A. 1968. The Big Bend of the Rio Grande: a guide to the rocks, landscape, geologic history, and settlers of the area of Big Bend National Park. Bureau of Economic Geology, The University of Texas at Austin, Austin, 138 pp.
- Myers, T. S. & Fiorillo, A. R. 2009. Evidence for gregarious behaviour and age segregation in sauropod dinosaurs. Palaeogeography, Palaeoclimatology, Palaeoecology, 274, 196–204.
- Novas, F. E., Salgado, L., Calvo, J. & Agnolin, F. 2005. Giant titanosaur (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia. Revista del Museo Argentino de Ciencias Naturales, n.s., 7, 37–41.
- Ortiz-Jaureguizar, E. & Pascual, R. 2011. The tectonic setting of the Caribbean region and the K/T turnover of the South American land-mammal fauna. Boletín Geológico y Minero, 122, 333–344.
- Owen, R. 1842. Report on British fossil reptiles, Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science, 60–204.
- Powell, J. E. 2003. Revision of the South American titanosaurid dinosaurs: palaeontological, palaeobiogeographical, and phylogenetic aspects. Records of the Queen Victoria Museum, 111, 1–173.
- Poropat, S. F., Upchurch, P., Mannion, P. D., Hocknull, S. A., Kear, B. P., Sloan, T., Sinapius, G. H. K. & Elliott, D. A. 2015. Revision of the sauropod dinosaur Diamantinasaurus matildae Hocknull et al. 2009 from the mid-Cretaceous of Australia: implications for Gondwanan titanosauriform dispersal. Gondwana Research, 27, 995–1033.
- Prieto-Marquez, A. & Salinas, G. C. 2010. A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. Journal of Vertebrate Paleontology, 30, 813–837.
- Saegusa, H. & Ikeda, T. 2014. A new titanosauriform sauropod (Dinosauria: Saurischia) from the Lower Cretaceous of Hyogo, Japan. Zootaxa, 3848, 1–66.
- Salgado, L. 1996. Pellegrinisaurus powelli nov. gen. et sp. (Sauropoda, Titanosauridae) from the Upper Cretaceous of Lago Pellegrini, northwestern Patagonia, Argentina. Ameghiniana, 33, 355–365.
- Salgado, L., Coria, R. A. & Calvo, J. O. 1997. Evolution of the titanosaurid sauropods. I: phylogenetic analysis based on the postcranial evidence. Ameghiniana, 34, 3–32.
- Salgado, L., Apesteguía, S. & Heredia, S. E. 2005. A new specimen of Neuquensaurus australis, a Late Cretaceous saltasaurine titanosaur from north Patagonia. Journal of Vertebrate Paleontology, 25, 623–634.
- Sampson, S. D., Loewen, M. A., Farke, A. A., Roberts, E. M., Forster, C. A., Smith, J. A. & Titus, A. L. 2010. New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism. PLoS ONE, 5, e12292.
- Sampson, S. D. & Loewen, M. A. 2010. Unravelling a radiation: a review of the diversity, stratigraphic distribution, biogeography, and evolution of horned dinosaurs. (Ornithischia:Ceratopsidae). Pp. 405–427 in M. J. Ryan, B. J. Chinnery-Allgeier & D. A. Eberth (eds) New perspectives on horned dinosaurs. Indiana University Press, Bloomington.
- Sereno, P. C. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläeontologie, Abhandlungen, 210, 41–83.
- Sullivan, R. M. & Lucas, S. G. 2000. Alamosaurus (Dinosauria: Sauropoda) from the Late Campanian of New Mexico and its significance. Journal of Vertebrate Paleontology, 20, 400–403.
- Swofford, D. L. 2002. PAUP*: phylogenetic analysis using parsimony (and other methods). 4.0 Beta. Sinaur Associates, Sunderland.
- Turner, K. J., Berry, M. E., Page, W. R., Lehman, T. M., Bohannon, R. G., Scott, R. B., Miggins, D. P., Budahn, J. R., Cooper, R. W., Drenth, B. J., Anderson, E. D. & Williams, V. S. 2011. Geologic Map of Big Bend National Park, Texas. United States Geological Survey, Scientific Investigations Map 3142, 84 pp.
- Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society, 124, 43–103.
- Upchurch, P., Barrett, P. M. & Dodson, P. 2004. Sauropoda. Pp. 259–322 in D. Weishampel, P. Dodson & H. Osmolska (eds) The Dinosauria. 2nd edition. University of California Press, Berkeley.
- Wedel, M. J., Cifelli, R. L. & Sanders, R. K. 2000a. Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. Journal of Vertebrate Paleontology, 20, 109–114.
- Wedel, M. J., Cifelli, R. L. & Sanders, R. K. 2000b. Osteology, paleobiology, and relationships of the sauropod dinosaur Sauroposeidon. Acta Palaeontologica Polonica, 45, 343–388.
- Wedel, M. J. & Sanders, R. K. 2002. Osteological correlates of cervical musculature in Aves and Sauropoda (Dinosauria: Saurischia), with comments on the cervical ribs of Apatosaurus. PaleoBios, 22, 1–6.
- Weishampel, D. B., Barrett, P. M., Coria, R. A., Le Loeuff, J., Xu, X., Zhao, X., Sahni, A., Gomani, E. M. P. & Noto, C. 2004. Dinosaur distribution. Pp. 517–606 in D. Weishampel, P. Dodson & H. Osmolska (eds) The Dinosauria. 2nd edition. University of California Press, Berkeley.
- Wick, S. L. & Lehman, T. M. 2014. A complete titanosaur femur from west Texas with comments regarding hindlimb posture. Cretaceous Research, 49, 39–44.
- Wilson, J. A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology, 19, 639–653.
- Wilson, J. A. 2002. Sauropod dinosaur phylogeny: critique and cladistics analysis. Zoological Journal of the Linnean Society, 136, 217–276.
- Wilson, J. A. 2005. Overview of sauropod phylogeny and evolution. Pp. 15–49 in K. Curry Rogers & J. Wilson (eds) The sauropods. University of California Press, Berkeley.
- Wilson, J. A. 2006. Anatomical nomenclature of fossil vertebrates: standardized terms or ‘lingua franca’? Journal of Vertebrate Paleontology, 26, 511–518.
- Wilson, J. A. 2012. New vertebral laminae and patterns of serial variation in vertebral laminae of sauropod dinosaurs. Contributions from the Museum of Paleontology, University of Michigan, 32, 91–110.
- Wilson, J. A. & Sereno, P. C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5, Supplement to 18(2), 1–68.
- Wilson, J. A. & Upchurch, P. 2003. A revision of Titanosaurus Lydekker (Dinosauria – Sauropoda), the first dinosaur genus with ‘Gondwanan’ distribution. Journal of Systematic Palaentology, 1, 125–160.
- Wilson, J. A., D'Emic, M. D., Ikejiri, T., Moacdieh, E. M. & Whitlock, J. A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS ONE, 6, e17114.
- Woodward, H. N. & Lehman, T. M. 2009. Bone histology and microanatomy of Alamosaurus sanjuanensis (Sauropoda: Titanosauria) from the Maastrichtian of Big Bend National Park, Texas. Journal of Vertebrate Paleontology, 29, 807–821.
- Zaher, H., Pol, D., Carvalho, A. B., Nascimento, P. M., Riccomini, C., Larson, P., Juarez-Valieri, R., Pires-Domingues, R., da Silva, N. J. & Campos, D. A. 2011. A complete skull of an Early Cretaceous sauropod and the evolution of advanced titanosaurians. PLoS ONE, 6, e16663.
- Zurriaguz, V. & Powell, J. 2015. New contributions to the presacral osteology of Saltasaurus loricatus (Sauropoda, Titanosauria) from the Upper Cretaceous of northern Argentina. Cretaceous Research, 54, 283–300.