
ABSTRACT
Plants growing at the edges of their range limits are expected to be particularly sensitive to changes in precipitation and temperature regimes associated with climatic change. However, non-climatic factors are increasingly recognized as important constraints to species’ range expansions. Therefore, we assessed the effects of soil provenance with respect to the alpine treeline on the germination, growth, and survival of Engelmann spruce (Picea engelmannii) seedlings. Seedlings were grown under controlled conditions in a growth chamber and greenhouse for ninety days in soils collected from four treeline ecotones in the Canadian Rocky Mountains. By controlling seed source and climatic conditions, and eliminating competition and predation, we attribute differences in seedling viability to soil properties that differed across elevation zones and individual treeline sites. Overall, alpine soils originating from beyond the species’ current elevational range were least amenable to growth, and there was some indication of reduced germination and survival in high-elevation soils. Forest soils, which were coarser and more nutrient rich, hosted seedlings with greater above- and below-ground biomass. Thus, the physical and chemical characteristics of alpine soils in our study region may constrain future treeline expansion, underscoring the importance of incorporating soil properties when considering species’ distributions under climate change.
Introduction
The expansion of a species’ distribution into new habitat can serve as an indicator of climate change if its establishment is enhanced by changes in air temperature or precipitation. As such, the elevational extent of alpine treelines has long been viewed as a sensitive ecological indicator of climate change (see Smith et al. Citation2009). Increasingly, however, the mediating effects of non-climatic constraints on species distributions are being recognized (Ameztegui et al. Citation2016; Brown and Vellend Citation2014; Butler et al. Citation2007; Cairns et al. Citation2007; Kupfer and Cairns Citation1996; Macias-Fauria and Johnson Citation2013; Tingstad et al. Citation2015), suggesting that changes in treeline position may not necessarily be as responsive to climate change as previously believed. Indeed, there has been significant variability in the global response of treelines to environmental change throughout the past century (Harsch et al. Citation2009).
Throughout western Canada, paleoecological studies have been used to identify relationships between climate and tree establishment (Beaudoin Citation1989; Brown Citation2013; Kearney Citation1982; Laroque, Lewis, and Smith Citation2001; Luckman and Kavanagh Citation2000; Pisaric et al. Citation2003; Roush Citation2009). Much of this research has provided evidence of significant upslope advance and meadow infilling by trees throughout the twentieth century as limitations imposed by temperature and precipitation have diminished. However, modeling studies have revealed modest prospects for continued treeline advance throughout the next century. Using an ecosystem-based climate envelope modeling approach, Hamann and Wang (Citation2006) found that the extent of montane climate regions suitable for conifer species will likely undergo significant decline in British Columbia during the next 100 years. Further, Gray and Hamann (Citation2012) determined that tree species growing in western Canada already lag behind their climate niches by up to 130 km latitude and 60 m in elevation. Examining the effects of geological constraints on tree recruitment in southwestern Alberta, Macias-Fauria and Johnson (Citation2013) and Westbrook (Citation2014) found that further upslope advance in their study region will be significantly limited by unsuitable topographic conditions. Altogether, these studies suggest that the regional climate is unlikely to favor unrestricted treeline advance, and that lags in the response to climate change could be due to the impact of non-climatic effects on tree establishment and survival.
Although there has been much interest in the distributional changes of high-elevation forests in recent years, species distribution models are most often developed by identifying how environmental variability influences mature plants, with little consideration to its effect on reproduction and seedling establishment. Juvenile plants tend to be more sensitive to environmental extremes and to occupy a subset of conditions experienced by mature plants (Dobrowski et al. Citation2015). Establishment bottlenecks, such as adequate seed dispersal and abundance and suitable microhabitat for establishment, have long been recognized as constraints for the establishment of invasive (nonnative) colonizers (Simberloff Citation2009), and the importance of these processes for native range expansions under climate change is now also being appreciated (Poschlod et al. Citation2013; Rosbakh, Poschlod, and Anten Citation2015). Understanding the constraints imposed by biotic and abiotic processes, as well as their interactions, is essential for predicting where future seedling establishment will occur (Elliott Citation2011; Smith et al. Citation2009).
Experimental field, growth chamber, and greenhouse studies are particularly useful approaches for disentangling the many processes that affect plant success, especially during the early life stages of trees. For example, reciprocal soil transplant experiments have been used to examine the importance of microclimate versus soil characteristics across alpine treeline ecotones (Germino et al. Citation2006) and the effects of “home” versus “away” soil provenances on seedling germination (Brown and Vellend Citation2014; Urli et al. Citation2016). Greenhouse and growth-chamber studies have also been used to identify the influence of soil properties on tree growth while controlling for the effects of climate and interspecific competition (Chapin et al. Citation1994; Urli et al. Citation2016) and to examine changes in relationships between seedling growth and soil biota with respect to elevation (Wagg et al. Citation2011). With the ability to control for factors such as climate, variability in seed quality, predation, and competition, it is possible to attribute differences in seedling success directly to the variables of interest.
Here, we focus on soil properties with respect to elevation as possible constraints to seedling germination, growth, and survival (hereafter seedling viability). Edaphic properties play an important role in determining local to regional plant distributions, but are often missing from predictions of future species ranges (Lafleur et al. Citation2010). To assess the effects of soil properties on seedling viability, we conducted growth-chamber and greenhouse studies in which Engelmann spruce (Picea engelmannii) seeds were planted in soils collected across four treeline ecotones in western Canada. Engelmann spruce was selected for the analysis because it grows in moderate to high abundance within each of the four study locations. We hypothesized that seedling success is influenced by differences in physical and chemical soil properties that vary across treeline ecotones. With the view that future establishment may be constrained by non-climatic processes, our expectation was that forest soils, collected from areas where trees are already established, would be more amenable to seedling success than soils from the transition or alpine zones. In the case that transition and/or alpine soils host less successful tree seedlings, high-elevation soil properties would be viewed as limiting to future upslope treeline advance. Given that treeline expansion has the potential to modify habitat conditions (Greenwood and Jump Citation2014; Theurillat and Guisan Citation2001) and to alter ecological processes at various scales (Beniston Citation2003; Cairns and Malanson Citation1998), identifying locally defined limitations to tree seedling regeneration is an important objective when considering where alpine treeline advance will occur in the future (Rosbakh, Poschlod, and Anten Citation2015).
Methods and site description
Study site description
Soil samples were collected from four treelines located along a northwest–southeast transect covering approximately 2° of latitude in the Front and Main Ranges of the Canadian Rocky Mountains (, ; ). The study areas were located at Wilcox Pass (Jasper National Park, Alberta), Goodsir Pass (Kootenay National Park, British Columbia), Fortress Mountain (privately owned land near Spray Valley Provincial Park, Alberta), and Highwood Pass (Peter Lougheed Provincial Park, Alberta). The site at Fortress Mountain is located on property that was previously operated as a ski resort; however, our study area has seen little direct impact from development.
Table 1. Study site locations and characteristics of treeline environment. Lists of nonarboreal vegetation are not exhaustive but are representative of dominant species at each site. Nondominant or rarely encountered tree species are denoted by *. (ES: Engelmann spruce; LPP: Lodgepole pine; SF: subalpine fir; SL: subalpine larch; WP: Whitebark pine)
Figure 1. Site photos (a) and map (b) of the four study areas in western Canada. Soil samples were collected from the forest, transition, and alpine zones of each treeline ecotone
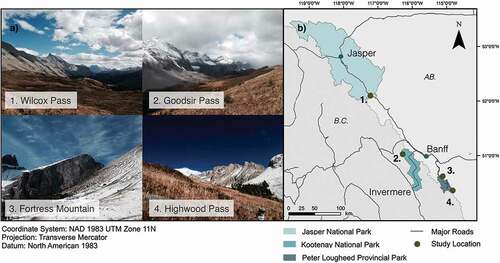
Soils at the study areas are a mix of brunisols, regosols, and podzols. Soils at Wilcox Pass, Fortress Mountain, and Highwood Pass are underlain by calcareous colluvial and morrainal material (Archibald, Klappstein, and Corns Citation1996; Holland and Coen Citation1982), whereas those from Goodsir Pass are derived from non-calcareous morrainal and fluviolacustrine material (Achuff et al. Citation1984). The four treeline environments host similar tree species but differ in the composition of non-arboreal vegetation ().
Long-term, local weather data are not available for the study areas because of their remote locations; however, temperature and precipitation normals derived from interpolated climate data (ClimateWNA; Wang et al. Citation2012) indicate that the study areas experience similar January and July mean temperatures (−11°C and 9°C, respectively). The sites differ somewhat in their precipitation regimes. Interpolated mean annual precipitation is nearly 500 mm higher at Goodsir Pass compared to the other field locations (mean annual precipitation of all sites = 1,140 mm). It is possible that the interpolated data may underestimate precipitation at Fortress Mountain, which is locally known to be an area of significant snowfall. All sites have continental climates with long, cold winters and short summers.
Soil collection
Soil samples were collected in September 2015 from the forest, transition, and alpine zones of the four study areas (twelve soil provenances in total). The forest zones were defined within the subalpine forests as having continuous tree canopy cover and tree densities of >50 trees/ha. For diffuse treelines, the transition zone had tree densities of 10–50 trees/ha, whereas the transition zone of abrupt treelines was located at an upslope distance of 15 × the height (m) of the abrupt tree edge. The alpine zone had low tree densities (<1 tree/ha) for diffuse treelines, or were located at an upslope distance of 30 × the height (m) of the abrupt tree edge (Brown et al. Citation2013). Thus, elevation increased from forest to transition to alpine zones, and although the actual elevations of the zones differed between sites, these criteria ensure that samples were collected at the same positions relative to the treeline between study sites.
Within each elevation zone, the top 10 cm of soil was collected from a minimum of five locations that were >5 m apart (volume collected, ca. 8 l). Engelmann spruce seeds are unlikely to interact directly with soil deeper than 10 cm during germination and initial growth because they fall directly on the soil surface and are not persistent in the seedbank (Johnson and Fryer Citation1996). Soil samples were stored in sealed plastic bags and transported to the University of Guelph, Ontario, where they were stored at 4°C until the beginning of the experiments. The bags of soil from each provenance were mixed to make a uniform soil treatment for each of the twelve site/elevation zone combinations.
Seedling germination and growth
Wild-sourced Engelmann spruce seeds collected from the Engelmann spruce-subalpine fir zone of a field location in southeastern British Columbia (51.02°N, 116.82°W) were provided for the study by the B.C. Tree Seed Centre (South Surrey, BC). Seed adaptations to habitat conditions have been observed in conifer species (Pickles et al. Citation2015), and using seeds collected from a single location allows us to control for the effects of such adaptations while isolating the influence of soil characteristics. Prior to planting, the seeds were stratified in a moistened all-purpose seedling starting mix for three weeks at 4°C to mimic the dormancy breaking effects of the cold and wet overwinter conditions experienced by Engelmann spruce in their natural environments (Bonner and Karrfalt Citation2008). After stratification, a hydrogen peroxide treatment was used to assess the viability of the stratified seeds following standard methods described by Leadem et al. (Citation1997).
Trial 1
The first experiment was conducted in a growth chamber at the University of Guelph Phytotron facility from January to March 2016. Seeds were sorted from the stratification mix and sown individually into cells of plastic potting inserts (cell size: 4 cm × 4 cm × 5.5 cm) filled with soil from each of the provenances (twenty seeded cells/soil provenance). Mixing and repotting the field-collected soils inevitably alters the soil structure from that observed in natural field conditions; however, it is a practical means for isolating the effects of soil properties on seedling viability from other confounding field conditions. Trays of sown cells were positioned in the growth chamber and randomly rearranged once per week. The sown cells were initially covered with plastic humidity domes to reduce the likelihood of the soil drying out between waterings. Seedlings were watered with reverse osmosis water approximately every two days to ensure the soil stayed moist, and were gradually hardened off the humidity domes after two weeks. Aside from the Engelmann spruce seedlings, any vegetation that germinated in the trays was removed to eliminate competition for resources, and germination and mortality were monitored for ninety days from the start of the experiment. The growth chamber was programmed to 16 h light at 20°C and 8 h dark at 15°C, with a relative humidity (RH) of 65 percent.
Trial 2
The experiment was repeated from May to July 2016 using fifty seeds per soil provenance to determine if the trends observed in Trial 1 were reproducible. Because of the space constraints of using a greater number of replicates, the second trial took place in a greenhouse at the Phytotron facility. The environmental conditions within the greenhouse were more variable than in the growth chamber, with average daytime temperatures of 21–27°C, average nighttime temperatures of 17–22°C, and RH of 8–93 percent (mean = 37%). Temperatures were consistently below 30°C in the greenhouse except on days 10 and 11 of the experiment, when maximum daily air temperatures reached 35°C and 31°C, respectively. Aside from the greater environmental variability and higher number of replicates, the methods used in the greenhouse were identical to those of the growth-chamber study.
Following each trial, various physical attributes of the surviving seedlings were measured. Seedling heights were measured after clipping the shoots at the soil surface, and roots were separated from the soil and gently rinsed in a beaker of water. The roots and shoots were dried at 110°C for 48 h, after which their dry weight was measured to the nearest milligram and the shoot:root dry weight ratio was calculated.
Soil analyses
The soil samples were analyzed for various physical and chemical properties that we expected may cause differences in seedling viability. Broadly, the measured soil properties pertained to factors that indicate either the abundance or accessibility of soil water and nutrients. Soil organic content is an indicator of the physical structure and size of the potential nutrient pool and is thus related to overall soil productivity. Soils high in organic content tend to have better aeration, moisture-holding capacity, and greater inputs of plant material that enhance nutrient availability. Soil organic content was measured using a loss-on-ignition technique following methods adapted from Heiri, Lotter, and Lemcke (Citation2001). Weighed soil samples (ca. 2 gwet) were placed in dried crucibles and heated to 110°C in a muffle furnace for 24 h to dry the samples. The dry sediment weights were recorded, and the samples were returned to the furnace for 4 h at 550°C to burn organic matter. They were then cooled and weighed, and the percent of soil organic content was calculated ((1 − (weight550°C / weight110°C)) × 100). Means were calculated for four replicates of each soil provenance.
Soil particle size distributions were determined from samples that were heated for 4 h at 550°C to remove organic content (ca. 300 gdry). The ashed soil was split into four size classes (pebble >4.75 mm; granule 2–4.75 mm, sand 0.075–2 mm, and silt/clay <0.075 mm) by passing the soil through nested sieves using a sediment shaker for ten minutes and determining the resultant weights. The procedure was repeated on three samples from each provenance, and the mean of the measurements was taken as representative. Particle size distribution is indicative of a soil’s water-holding capacity (soils with smaller particles hold more water) and the ease with which plant roots can infiltrate the soil to access water and nutrients (plant roots grow more easily in soils with greater pore space).
Various metrics related to soil fertility were also measured because of their potential to influence seedling growth and survival. Phosphate concentrations (ppm) were determined using the ascorbic acid reduction method (Code 3653-SC) on prepared soil extracts and a LaMotte SMART 3 colorimeter (LaMotte Company). Soil samples were also sent to the Forest Soils Laboratory at the University of Guelph for additional chemical analyses: pH, electrical conductivity, total nitrogen, and soil nitrogen availability were quantified following methods described in Carter and Gregorich (Citation2006). Soils with greater nutrient concentrations and higher pH and electrical conductivity are generally viewed as more favorable for plant growth.
Statistical analyses
Separate generalized linear mixed-effects models (GLMMs) with binomial distributions and logit link functions were used to examine the effects of trial and elevation zone on the proportions of seedling germination, survival, and mortality. Trial, elevation zone, and their interactions were fixed effects, and site and elevation zone nested within site were the random components of the models. The trial by elevation zone interaction was not included in the seedling survival model because of failure to achieve model convergence. GLMMs were assessed using the glmer function of the lme4 package (Bates et al. Citation2015) in R (R Core Team Citation2015).
As a measure of survivorship, the proportions of seedlings living on each day of the experiment were calculated by dividing the difference between cumulative germination and mortality by the total number of seeds planted per elevation zone for the respective trial.
Separate linear mixed effects (LME) models were used to determine the fixed effects of trial, elevation zone, and their interactions on seedling height, root mass, and shoot:root mass. Seedling height is positively associated with potential photosynthetic capacity and overall seedling vigor (e.g., for Picea glauca; Benomar et al. Citation2015), whereas root biomass is an indicator of belowground carbon investment and reflects a seedling’s ability to access soil water and nutrients. Shoot:root mass provides information about overall seedling health, with smaller values reflecting structural growth restrictions caused by nutrient limitations (primarily N, P, K; Ericsson Citation1995; Ericsson, Rytter, and Vapaavuori Citation1996). In the LME models, random effects were site and elevation zone nested within site. After checking for normality in the distribution of residual versus fitted values of each model, root mass was square-root transformed, shoot:root was log transformed, and the models were re-run. Mixed effects models were assessed using the lme function of nlme (Pinheiro et al. 2016) in R. ANOVA tables of GLMM and LME models are provided in .
Differences in growth across the elevation zones of individual sites were also examined. LME models revealed that trial identity and/or its interaction with elevation had significant effects on seedling growth, making it inappropriate to pool the raw data from the two experiments. Height, root mass, and shoot:root mass from each trial were thus transformed to z scores using the grand mean of the respective trial prior to pooling the data. The normality of the resultant data was assessed using Shapiro-Wilk’s tests with a significance cutoff of p ≤ .05 (results not shown). Differences in pooled and site-level seedling heights between forest, transition, and alpine zones, which were normally distributed, were assessed using parametric Student’s t-tests that accounted for unequal variances between sample populations. Differences in site-level root mass and shoot:root mass were assessed using Wilcoxon rank sum tests because of non-normality in the z-score transformed data.
We were unable to achieve model convergence when measurements of the various soil properties were incorporated into the GLMM and LME models, likely because of the large number of explanatory variables relative to the sample size. The measured soil properties were therefore examined using principal components analysis (PCA) to infer the cause of the variance observed in the measured seedling variables. The PCA was developed using the prcomp function in the stats package in R. To explore possible relationships between topographic positioning and soil properties, the scores of PCs 1 and 2 were correlated (Spearman’s rank) to aspect, latitude, and elevation zone (treated as ordinal data).
Results
Seedling viability
Pretrial germination tests indicated that the average viability of Engelmann spruce seeds was 90 percent before Trials 1 and 2. Germination was consistently higher in Trial 1 (growth chamber) than in Trial 2 (greenhouse; , ). The proportion of germinated seedlings decreased with elevation in Trial 1, whereas germination was similar in the forest and alpine zones and lowest in the transition zone in Trial 2 (, ). The GLMM for seedling germination supports these observed trends; germination was significantly lower in alpine soils and in Trial 2 (zalpine = −2.33, palpine = .02; ztrial2 = −5.70, ptrial2 = <.01; ) compared to forest soils and Trial 1. There was a significant interaction between Trial 2 and the alpine zone (ztrial2×alpine = 2.53, ptrial2×alpine = .01), indicating that the difference in the proportion of germination between forest and alpine soils was smaller in Trial 2 than in Trial 1.
Table 2. Results of Engelmann spruce germination, mortality, and growth. Trials 1 and 2 are separated by commas (WP: Wilcox Pass; GP: Goodsir Pass; FM: Fortress Mountain; HP: Highwood Pass)
Table 3. Results of generalized linear mixed effects (germination, survival, and mortality) and linear mixed effects (height, root mass, and shoot:root) models of the effects of trial and soil elevation on measures of seedling viability. Effects are expressed as relative to the forest zone and to Trial 1 (baseline conditions). Probabilities in bold indicate p ≤ .05
Figure 2. Average measures (± standard error) of seedling viability and growth across elevation zones and pooled by site. The data are divided by Trials 1 (![]()
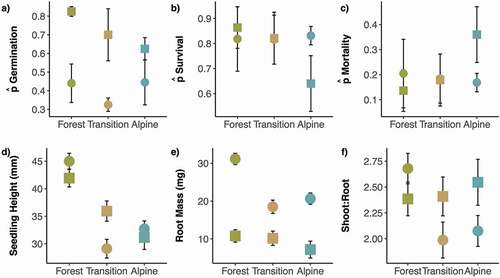
Seedling survival (the number of seedlings that survived the experiment out of those that germinated) did not differ significantly between trials or elevation zones (ztrial2 = 0.79, ptrial2 > .05; ztransition = −0.14, ptransition > .05; zalpine = −0.96, palpine > .05; interaction terms not tested), although seedling survival decreased from forest to transition to alpine soil zones in Trial 1 (, ). In Trial 1, the number of seedlings that germinated and survived the experiments () was highest in the forest, followed by the transition and alpine zones. In Trial 2, survivorship was highest in the alpine zone and lowest in the transition zone. The combined effects of germination and mortality on seedling survival can be seen in , where the proportions of living seedlings differed between trials and elevations in a similar fashion to survival of germinated seedlings in .
Figure 3. Proportion of seedlings living on a given day of the experiment (![]()
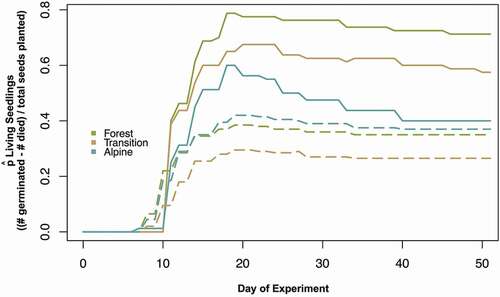
The GLMM of proportional mortality identified significant effects of alpine elevation (zalpine = 2.17, palpine = .03) and a significantly negative effect of the interaction between Trial 2 and the alpine soil source (ztrial2xalpine = 0.01; ptrial2xalpine = .01). The significant interaction term indicates that the higher mortality in alpine relative to forest soils in Trial 1 was not apparent in Trial 2 (). The steep, negative slope after day 18 and the low proportion of living seedlings in the survival curve of alpine seedlings in Trial 1 () reveals the combined effects of low overall germination followed by high mortality.
Growth metrics: Seedling height, root mass, shoot:root mass
Seedling height decreased with increasing elevation zone in both trials, and trial identity on its own did not have significant effects on shoot growth (ttrial2 = 0.60, ptrial2 > .05; , ). The effects of transition and alpine zones were both negative in the LME model, and alpine soil had a significantly reduced seedling height relative to the forest (ttransition = −1.68, ptransition = .14; talpine = −3.19, palpine = .02). The effect of the interaction between Trial 2 and transition zone soil was significant (ttrial2xtransition = −2.87, ptrial2×transition < .01), indicating that seedling growth in the transition zone soil was more reduced in Trial 2 relative to the forest zone than it was in Trial 1. When sites were examined separately, all except Wilcox Pass showed a significant elevational decrease in seedling height between the forest, transition, and alpine zones (). Differences in seedling height between transition alpine zones were variable at the site level.
Figure 4. Box plots of seedling height (a), root mass (b), and shoot:root (c) by study area and elevation zone (![]()
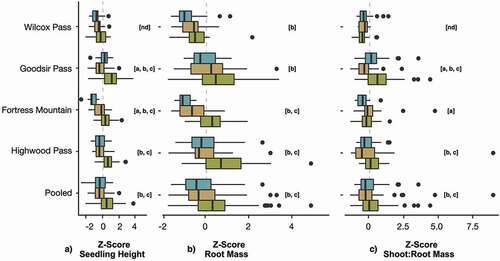
Root mass was significantly reduced in Trial 2 (ttrial2 = 12.71; ptrial2 < .01) and in alpine soils (talpine = −2.98; palpine = .02; , ). Interactions between Trial 2 and the transition zone (ttrial2×transition = −4.52, ptrial2xtransition < .01) and between Trial 2 and alpine zone (ttrial2×alpine = −2.01, ptrial2xalpine = .04) were significantly negative. This indicates that there was a greater reduction in root mass with increasing elevation zone in Trial 2 compared to Trial 1 (). At the site level, root mass tended to decrease with increasing elevation zone () and was highest and most variable in the forest zone.
Shoot:root mass did not differ between trials (ttrial2 = 0.13, ptrial2 = .90; , ) or elevation zones (ttransition = −0.71, ptransition = .51; talpine = 0.32, palpine = .71). There was a significant negative interaction between Trial 2 and the alpine zone (ttrial2×alpine = −2.09, ptrial2×alpine = .04). Overall, there was no consistent elevational trend in shoot:root mass across elevation zones within each site; however, shoot:root mass was significantly higher in the forest zones of Goodsir and Highwood Passes relative to the transition and alpine zones of the same sites ().
Soil properties
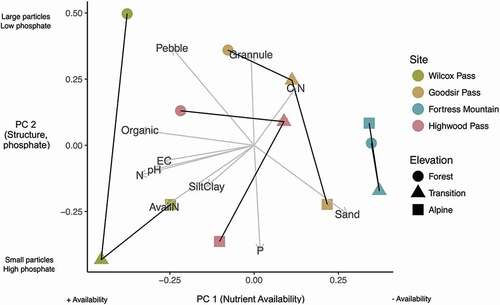
The principal components analysis revealed that trends in soil nutrient availability (Principal component (PC) 1; proportion of variance explained = 45.1%) and physical soil structure (PC 2; 18.4%) varied both within and between sites (; raw measurements of soil properties are available in ). Along PC 1, soil provenances differed primarily at the site level, with organic matter and available nitrogen content highest at Wilcox Pass and lowest at Fortress Mountain. Notably, differences in soil nutrient availability between sites did not correspond to between-site differences in seedling viability (e.g., the site with the highest nutrient availability, Wilcox Pass, tended to rank lowest in measures of seedling viability). Within sites, forest soils tended to have slightly more negative values of PC 1 (i.e., greater nutrient availability) than alpine soils. Soils from the transition zone were variable relative to the other elevation zones and showed no clustering. The results of the Spearman rank correlation revealed that PC 1 was strongly positively correlated with aspect (rs = 0.87, p < 0.01) but not latitude (rs = −0.41, p = 0.2) or elevation zone (rs = 0.12, p = 0.7). This indicates that nutrient availability was greater in soils collected from sites with cooler aspects. The distribution of soil provenances along PC 2 demonstrates that phosphate content and soil particle size varies mainly by elevation zone (rs = 0.65, p = .02). PC 2 was not correlated with aspect (rs = 0.07, p = .80) or latitude (rs = 0.06, p = .80). Alpine soils generally had more negative PC 2 values, indicating that they have a greater proportion of small particles and higher concentrations of phosphate. Conversely, forest and transition soils tended to fall higher on PC 2, indicating their greater proportions of pebbles and granules relative to alpine soils. Soils from Fortress Mountain did not follow these trends; soil particle size and nutrient content were comparable between the forest and alpine zones.
Figure 5. Results of principal component analysis demonstrating variability in soil properties explained by components 1 and 2. The cumulative variance explained by the first two components is 63.5 percent. PC 1 (proportion of variance = 45.1%) is primarily related to soil nutrient availability, and PC 2 (proportion of variance = 18.4%) is related to soil structure (particle size) and phosphate concentration
Discussion
Our objective was to determine whether differences in soils collected along treeline ecotones influence the viability of Engelmann spruce seedlings when controlling for climate, competition, and other potentially limiting factors. We expected that soils collected from forest zones, where mature trees are currently growing in high densities, would be the most favorable to seedling viability. Overall, the results of the growth chamber and greenhouse studies support this expectation as they were indicative of greater seedling growth and survival in the forest zone relative to the alpine. There was, however, some variability in the results between trials.
Seedling establishment and growth
The higher proportions of seedling germination and survival in the forest soils of Trial 1 suggest that these low-elevation soils were most suitable for Engelmann spruce establishment. This effect was not apparent in Trial 2, with little difference between germination and survival between elevation zones. It is possible, however, that the differences in establishment processes between trials was related to the variability in environmental conditions in the greenhouse (Trial 2) relative to the growth chamber (Trial 1; see discussion of trial effects further on). We therefore cautiously interpret the results of seedling germination and survival as indicative that forest soils were more favorable for establishment.
Forest-zone soils also seemed more favorable for seedling growth; seedling height and root mass were highest in the forest soils of all sites (except for seedling height from Wilcox Pass). Differences in the effect of soil provenance existed between trials (significant interaction terms in LME models); however, the tendency for lower shoot height and root growth in alpine soils relative to forest-zone soils was apparent in both LME models (significant negative effect of alpine-zone soil). The absence of significant fixed effects in the shoot:root model indicate that this metric was not strongly influenced by differences in soil properties between elevation zones. Altogether, the decline in shoot height and root mass with respect to elevation zone suggests that seedlings grown in forest soils were the most robust. For instance, their greater shoot and root biomass is indicative of higher growth potentials by way of increased surface area for photosynthesis and through a greater capacity to access water and nutrients from the soil via higher root growth. Seedling growth immediately following germination is recognized as a bottleneck for tree establishment in harsh alpine environments (Castanha et al. Citation2013), and our results suggest that seedlings can experience reduced growth in alpine soils even in the absence of climate limitations.
The results of our growth chamber and greenhouse experiments are broadly similar to other experimental field and growth-chamber studies of seedling germination and survival. Reciprocal soil transplant and seed addition studies have shown that tree seedlings planted in alpine soils that have been translocated to the forest have reduced germination relative to seedlings planted in the forest soil itself (e.g., Brown and Vellend [Citation2014] for Acer saccharum). In addition, it has been suggested that enhanced survival of Engelmann spruce seedlings in forest soils can be caused by preconditioning of the soil by mature trees. Germino et al. (Citation2006) found that the colonization of seedling root tips by soil fungi was higher in forest relative to treeline soils, and that most seedlings that survived from the emergent to juvenile life stage had their root tips colonized. Reduced fungal colonization at high elevations is believed to be caused by low densities of mature trees that support the fungal community (Germino et al. Citation2006; Hasselquist et al. Citation2005). Finally, lower seedling growth in high-elevation soils has also been observed for interior spruce (Picea glauca x engelmannii) and lodgepole pine (Pinus contorta) seedlings. In a growth-chamber study, Wagg et al. (Citation2011) found that the relative growth rates of seedlings were lower in soils collected from a high-elevation field site in British Columbia, Canada, compared to growth in low-elevation soils from the same site. Unlike their study, which also found that shoot:root mass increased with elevation (Wagg et al. Citation2011), we found limited evidence that elevation zone was related to this growth ratio, and we found that root mass decreased, rather than increased, with elevation.
Trial effects
A confounding factor in our study is the effect of trial on measures of seedling viability. Overall, differences between trials appeared to have greater effects on seedling germination and survival (and thus mortality) than on seedling growth. This was likely caused by the consequences of extreme temperatures during a critical period of seedling germination and survival in the greenhouse trial. In both Trials 1 and 2, seedling germination peaked around days 10–12, during which time air temperatures reached more than 30°C in Trial 2. Observations of germinants during this period were that many seeds that were undergoing germination (visible at the soil surface) around day 10 of Trial 2 did not emerge, and many emerged germinants died during and immediately after the event. Additionally, germinating seeds and emerged seedlings of select trays of forest soils appeared to be disproportionately affected by the hot temperatures in Trial 2. This reduced the number of germinating and surviving seedlings in forest-zone soils overall (). We cautiously conclude that when extreme climatic events are minimized, as in Trial 1, forest-zone soils are more favorable for establishment. In other words, the disproportionate impact of the hot conditions on forest soils in Trial 2 make the measures of germination and survival in Trial 1 more reflective of soil properties. We cannot identify the cause of differences in the effects of temperature extremes between soil provenances without further study. Our findings suggest that temperature extremes, which are expected to become increasingly common with continued climate warming (IPCC Citation2014), can outweigh the effects of soil properties during periods critical for seedling germination and survival. However, climate projections indicate that springtime temperatures in our study area are unlikely to reach the intensity of those in the greenhouse trial during the next century (mean daily temperatures >30°C; Wang et al. Citation2012).
Effects of soil properties on seedling viability
The results of the principal components analysis suggest that soil nutrient availability and particle size distribution are important determinants of seedling viability across the four studied treeline ecotones. We did not directly correlate measures of seedling viability to PCA scores because of the clustered nature of the data; however, some trends emerge when soil properties and viability are compared more generally. In our analysis, we found that soil nutrient availability was related to slope aspect and differed most significantly between sites. Cooler slopes tended to have lower values of PC 1, indicating greater nutrient availability. The tendency for cool aspects to have greater nutrient availability is counter to the generally accepted idea that warmer soils enable greater microbial activity and thus greater nutrient availability (Holtmeier Citation2009). However, these cooler aspects could also experience more abundant or longer periods of insulating snow cover, which can prevent soil from freezing and allow decomposition to occur for longer time periods (Holtmeier Citation2009). Between-site differences were likely a result of the greater similarity in parent material and climate conditions within sites than between them. No clear relationship emerged between nutrient content and elevation zone; however, forest soils tended to have lower PC 1 values (greater nutrient availability) than their alpine counterparts from the same site.
Many studies that have analyzed relationships between soil nutrient content and elevation have found high-elevation soils to be less fertile than soils from lower elevations (Huber et al. Citation2007; Loomis et al. Citation2006; Wagg et al. Citation2011), although the relationship is far from universal (Brown and Vellend Citation2014; Germino et al. Citation2006; Liptzin, Sanford, and Seastedt Citation2012; Liptzin and Seastedt Citation2009). Variability in soil fertility across elevation zones can be caused by several factors, such as patterns of snow accumulation, microtopography, and nutrient deposition (Holtmeier Citation2009; Seastedt et al. Citation2004). The lower fertility of our studied alpine soils relative to forest soils from the same sites could be caused by greater temperature extremes, cooler and wetter conditions, and reduced inputs of organic matter in the alpine compared to the lower elevation zones. In addition to lower nutrient content in general, alpine soils also tended to have slightly lower pH and electrical conductivity than forest soils. This would have made the fewer nutrients available less accessible to seedlings and thus could have contributed to reduced seedling viability in alpine soils. Although some studies have found that soil nutrient availability constrains the growth of mature trees at the treeline (Sullivan et al. Citation2015), we hesitate to suggest that seedlings in any soil provenance experienced critical levels of nutrient limitation. Differences in seedling viability metrics between sites did not correspond to between-site differences in nutrient availability.
The results of the PCA also revealed that soil particle size distribution may be important in determining seedling viability across elevation zones. Differences in soil properties with respect to elevation appeared to be most strongly related to PC 2. Forest soils, in which seedlings grew taller, invested more in belowground biomass, and were more likely to germinate and survive in Trial 1, tended to have a greater proportion of large particles and lower phosphate concentrations relative to alpine soils (higher values of PC 2).
Elevational differences in soil particle size could be caused by increased physical weathering in the alpine, an important process of soil formation at high elevations (Howell and Harris Citation1978). A possible explanation for the apparent relationship between particle size and seedling viability is that root growth is facilitated in the more porous forest soils, which would indirectly improve access to water and nutrients (Oren and Sherriff Citation1995; Perry, Oren, and Hart Citation2008). Well-draining soil also tends to favor nutrient availability because more oxygen is available for the microbial decomposition of organic material (Perry, Oren, and Hart Citation2008). Germination and survival rates should benefit from soil properties that are also conducive to seedling growth, because robust seedlings would be better able to tolerate stressful conditions. The higher phosphate concentrations observed in alpine soils was surprising but could be caused by the relatively higher nutrient inputs that often occur in alpine environments through atmospheric deposition (Seastedt et al. Citation2004). Based on the tendency of seedling viability to be higher in forest-zone relative to alpine-zone soils, it appears that the increased phosphate concentrations did not greatly benefit seedling viability, either because phosphorus was not limiting or because seedlings were unable to access it in alpine soils.
Recently, there has been increasing appreciation for the role that edaphic properties have in moderating treeline dynamics (e.g., Dawes et al. Citation2017; Lafleur et al. Citation2010; Müller et al. Citation2015, Citation2016). It is important to recall, however, that no single factor is responsible for determining species’ range limits. Outside of the controlled environment of the greenhouse and growth chamber, seedlings establishing under the natural conditions of our field sites will also experience a variety of other factors that will influence their success. For example, seed predation by animals (Cairns et al. Citation2007), differences in light availability across the treeline ecotone (Maher and Germino Citation2006), and competition and facilitation with other plant species (Callaway Citation1998) are only a few of many factors that have the potential to interact with edaphic properties and climate to determine where seedlings can survive. The moderating effects of soil and other non-climatic factors suggest that treeline position may not be as sensitive to changes in climate as once believed (Kupfer and Cairns Citation1996).
Implications for future species distributions and forest management
Taken together, the directional trend of key viability indicators supports the assertion that, in isolation from the effects of climate, competition, seed predation, and dispersal limitations, the properties of alpine soils from our study areas in western Canada reduce the viability of Engelmann spruce seedlings. This finding suggests that the continued upslope advance of this species may be limited by soil properties, even if climate conditions become more favorable. Ecological models that accurately predict future species distributions are a much-needed tool for developing climate change adaptation strategies, and incorporating information about how geomorphic properties and processes modulate a species’ response to climate change has been identified as a key component of refining predictions of their future distribution (Lafleur et al. Citation2010; Macias-Fauria and Johnson Citation2013).
In addition to considerations of the effects of future changes in alpine habitat, our findings also have implications for management activities related to the assisted migration of subalpine tree species. Concern that economically and ecologically important tree species may not be able to keep pace with climate change has led to the suggestion that assisted migration of select species to higher latitudes or elevations may be an appropriate resource-management strategy (Ste-Marie et al. Citation2011; Winder, Nelson, and Beardmore Citation2011). Whether species dispersal is assisted or occurs naturally, our results suggest that establishment beyond existing tree-species ranges may see reduced success if soil properties are unsuitable for establishment, regardless of warming temperatures that might otherwise be expected to benefit their performance. Overall, our findings support the assertions of others that including edaphic properties and factors directly affecting establishment processes improves our understanding of future species distributions under climate change scenarios (Dubuis et al. Citation2013; Lafleur et al. Citation2010; Poschlod et al. Citation2013; Rosbakh, Poschlod, and Anten Citation2015), and that more consideration of the role of edaphic properties in determining future species distributions is warranted.
Supplementary Materials
Download PDF (491.7 KB)Acknowledgments
This research was funded by an NSERC Discovery grant awarded to ZMG and an NSERC Canada Graduate Scholarship (Doctoral) awarded to ELD. The seeds for this study were kindly provided by the B.C. Tree Seed Centre, Surrey, British Columbia. We thank Parks Canada, Alberta Parks, and Fortress Mountain Ski Resort for allowing access to field sites; Sean Fox and Spencer Reitenbach for their insights into growing and caring for tree seeds and seedlings; and Christine Loziak for assistance in the field.
ELD designed and executed the study, performed the analysis, and wrote the draft of the manuscript. HAH and ZMG contributed ideas to the study design and assisted with statistical analyses. All authors contributed to the final manuscript.
Supplementary data
Supplemental data for this article can be accessed here.
Related Research Data
References
- Achuff, P. L., W. D. Holland, G. M. Coen, and K. Van Tighem. 1984. Ecological land classification of Kootenay National Park, British Columbia. Vol. I: Integrated resource description: Alberta Institute of Pedology. Edmonton, Alberta: Alberta Institute of Pedology.
- Ameztegui, A., L. Coll, L. Brotons, and J. M. Ninot. 2016. Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees. Global Ecology and Biogeography 25 (3):263–73. doi:https://doi.org/10.1111/geb.12407.
- Archibald, J. H., G. D. Klappstein, and I. G. W. Corns, 1996. Field guide to ecosites of southwestern Alberta, Special Report 8. Edmonton, Alberta: UBC Press.
- Bates, D., M. Maechler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67 (1):1–48. doi:https://doi.org/10.18637/jss.v067.i01.
- Beaudoin, A. B. 1989. The response of three treeline zone to climatic change in the Canadian Rocky Mountains. The Canadian Geographer 33 (1):83–86.
- Beniston, M. 2003. Climatic change in mountain regions: A review of possible impacts. Climatic Change 59:5–31. doi:https://doi.org/10.1023/A:1024458411589.
- Benomar, L., M. S. Lamhamedi, I. Villeneuve, A. Rainville, J. Beaulieu, J. Bousquet, and H. A. Margolis. 2015. Fine-scale geographic variation in photosynthetic-related traits of Picea glauca seedlings indicates local adaptation to climate. Tree Physiology 35:864–78. doi:https://doi.org/10.1093/treephys/tpv054.
- Bonner, F. T., and R. P. Karrfalt, eds. 2008. The Woody Plant Seed Manual. Agricultural Handbook No. 727. Washington, DC.: U.S. Department of Agriculture, Forest Service. 1223 p.
- Brown, C. D., J. F. Johnstone, S. D. Mamet, and A. J. Trant. 2013. Global treeline range expansion experiment field protocols. Available: www.treelineresearch.com/about/.
- Brown, C. D., and M. Vellend. 2014. Non-climatic constraints on upper elevational plant range expansion under climate change. Proceedings of the Royal Society B: Biological Sciences 281:20141779–9. doi:https://doi.org/10.1098/rspb.2014.1779.
- Brown, R. 2013. Response of alpine treeline ecotones to 20th century climate change: A comparative analysis from Kananaskis Country, 81. Guelph, ON: Alberta, Department of Geography, University of Guelph.
- Butler, D. R., G. P. Malanson, S. J. Walsh, and D. B. Fagre. 2007. Influences of geomorphology and geology on alpine treeline in the American West: More important than climatic influences? Physical Geography 28 (5):434–50. doi:https://doi.org/10.2747/0272-3646.28.5.434.
- Cairns, D. M., C. Lafon, J. Moen, and A. Young. 2007. Influences of animal activity on treeline position and pattern: Implications for treeline responses to climate change. Physical Geography 28 (5):419–33. doi:https://doi.org/10.2747/0272-3646.28.5.419.
- Cairns, D. M., and G. Malanson. 1998. Environmental variables influencing the carbon balance at the alpine treeline: A modeling approach. Journal of Vegetation Science 9 (5):679–92. doi:https://doi.org/10.2307/3237286.
- Callaway, R. M. 1998. Competition and facilitation on elevation gradients in subalpine forests of the Northern Rocky Mountains, USA. Oikos 82 (3):561–73. doi:https://doi.org/10.2307/3546376.
- Carter, M. R., and E. G. Gregorich. 2006. Soil Sampling and Methods of Analysis (2nd ed.). Boca Raton, FL: CRC Press.
- Castanha, C., M. S. Torn, M. J. Germino, B. Weibel, and L. M. Kueppers. 2013. Conifer seedling recruitment across a gradient from forest to alpine tundra: Effects of species, provenance, and site. Plant Ecology & Diversity 6 (3–4):307–18. doi:https://doi.org/10.1080/17550874.2012.716087.
- Chapin, F. S., L. R. Walker, C. L. Fastie, and L. C. Shearman. 1994. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska. Ecological Monographs 64 (2):149–75. doi:https://doi.org/10.2307/2937039.
- Dawes, M. A., P. Schleppi, S. Hattenschwiler, C. Rixen, and F. Hagedorn. 2017. Soil warming opens the nitrogen cycle at the alpine treeline. Glob Chang Biol 23 (1):421–34. doi:https://doi.org/10.1111/gcb.13365.
- Dobrowski, S. Z., A. K. Swanson, J. T. Abatzoglou, Z. A. Holden, H. D. Safford, M. K. Schwartz., and D. G. Gavin. 2015. Forest structure and species traits mediate projected recruitment decline in western US tree species. Global Ecology & Biogeography 24 (8):917–27. doi:https://doi.org/10.1111/geb.12302.
- Dubuis, A., S. Giovanettina, L. Pellissier, J. Pottier, P. Vittoz, A. Guisan, D. Rocchini, and D. Rocchini. 2013. Improving the prediction of plant species distribution and community composition by adding edaphic to topo-climatic variables. Journal of Vegetation Science 24 (4):593–606. doi:https://doi.org/10.1111/jvs.2013.24.issue-4.
- Elliott, G. P. 2011. Influences of 20th-century warming at the upper tree line contingent on local-scale interactions: Evidence from a latitudinal gradient in the Rocky Mountains, USA. Global Ecology and Biogeography 20 (1):46–57. doi:https://doi.org/10.1111/geb.2011.20.issue-1.
- Ericsson, T. 1995. Growth and shoot: Root ratio of seedlings in relation to nutrient availability. Plant and Soil 168–169:205–14. doi:https://doi.org/10.1007/BF00029330.
- Ericsson, T., L. Rytter, and E. Vapaavuori. 1996. Physiology of carbon allocation in trees. Biomass and Bioenergy 11 (2/3):115–27. doi:https://doi.org/10.1016/0961-9534(96)00032-3.
- Germino, M. J., N. J. Hasselquist, T. McGonigle, W. K. Smith, and P. P. Sheridan. 2006. Landscape- and age-based factors affecting fungal colonization of conifer seedling roots at the alpine tree line. Canadian Journal of Forest Research 36 (4):901–09. doi:https://doi.org/10.1139/x05-303.
- Gray, L. K., and A. Hamann. 2012. Tracking suitable habitat for tree populations under climate change in western North America. Climatic Change 117 (1–2):289–303. doi:https://doi.org/10.1007/s10584-012-0548-8.
- Greenwood, S., and A. S. Jump. 2014. Consequences of treeline shifts for the diversity and function of high altitude ecosystems. Arctic, Antarctic, and Alpine Research 46:829–40. doi:https://doi.org/10.1657/1938-4246-46.4.829.
- Hamann, A., and T. Wang. 2006. Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 87 (11):2773–86. doi:https://doi.org/10.1890/0012-9658(2006)87[2773:PEOCCO]2.0.CO;2.
- Harsch, M. A., P. E. Hulme, M. S. McGlone, and R. P. Duncan. 2009. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecology Letters 12:1040–49. doi:https://doi.org/10.1111/ele.2009.12.issue-10.
- Hasselquist, N. J., M. J. Germino, T. McGonigle, and W. K. Smith. 2005. Variability of Cenococcum colonization and its ecophysiological significance for young conifers at alpine-treeline. New Phytologist 165:867–73. doi:https://doi.org/10.1111/j.1469-8137.2005.01275.x.
- Heiri, O., A. Lotter, and G. Lemcke. 2001. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. Journal of Paleolimnology 25:101–10. doi:https://doi.org/10.1023/A:1008119611481.
- Holland, W. D., and G. M. Coen. 1982. Ecological (biophysical) land classification of Banff and Jasper National Parks. Edmonton, Alberta: Alberta Institute of Pedology.
- Holtmeier, F.-K. 2009. Mountain timberlines: Ecology, patchiness, and dynamics. Dordrecht, The Netherlands: Springer Science+Business Media.
- Howell, J. D., and S. A. Harris. 1978. Soil-forming factors in the Rocky Mountains of Southwestern Alberta, Canada. Arctic and Alpine Research 10 (2):313–24. doi:https://doi.org/10.2307/1550763.
- Huber, E., W. Wanek, M. Gottfried, H. Pauli, P. Schweiger, S. K. Arndt, K. Reiter, and A. Richter. 2007. Shift in soil–plant nitrogen dynamics of an alpine–nival ecotone. Plant and Soil 301 (1–2):65–76. doi:https://doi.org/10.1007/s11104-007-9422-2.
- IPCC. 2014. Climate change 2014: Synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. Geneva, Switzerland: IPCC.
- Johnson, E. A., and G. I. Fryer. 1996. Why Engelmann spruce does not have a persistent seed bank. Canadian Journal of Forest Research 26:872–78. doi:https://doi.org/10.1139/x26-095.
- Kearney, M. S. 1982. Recent seedling establishment at timberline in Jasper National Park, Alta. Canadian Journal of Botany 60 (11):2283–87. doi:https://doi.org/10.1139/b82-279.
- Kupfer, J. A., and D. M. Cairns. 1996. The suitability of montane ecotones as indicators of global climatic change. Progress in Physical Geography 20:253–72. doi:https://doi.org/10.1177/030913339602000301.
- Lafleur, B., D. Paré, A. D. Munson, and Y. Bergeron. 2010. Response of northeastern North American forests to climate change: Will soil conditions constrain tree species migration? Environmental Reviews 18:279–89. doi:https://doi.org/10.1139/A10-013.
- Laroque, C. P., D. H. Lewis, and D. J. Smith. 2001. Treeline dynamics on Southern Vancouver Island, British Columbia. Western Geography 10/11:43–63.
- Leadem, C. L., S. L. Gillies, H. K. Yearsley, V. Sit, D. L. Spittlehouse, and P. J. Burton. 1997. Field studies of seed biology, 196. Victoria, BC: Research Branch, B.C. Ministry of Forests.
- Liptzin, D., R. L. Sanford, and T. R. Seastedt. 2012. Spatial patterns of total and available N and P at alpine treeline. Plant and Soil 365 (1–2):127–40. doi:https://doi.org/10.1007/s11104-012-1379-0.
- Liptzin, D., and T. R. Seastedt. 2009. Patterns of snow, deposition, and soil nutrients at multiple spatial scales at a Rocky Mountain tree line ecotone. Journal of Geophysical Research 114 (G4). doi:https://doi.org/10.1029/2009JG000941.
- Loomis, P. F., R. W. Ruess, B. Sveinbjörnsson, and K. Kielland. 2006. Nitrogen cycling at treeline: Latitudinal and elevational patterns across a boreal landscape. Ecoscience 13 (4):544–56. doi:https://doi.org/10.2980/1195-6860(2006)13[544:NCATLA]2.0.CO;2.
- Luckman, B. H., and T. Kavanagh. 2000. Impact of climate fluctuations on mountain environments in the Canadian Rockies. Ambio 29:371–80. doi:https://doi.org/10.1579/0044-7447-29.7.371.
- Macias-Fauria, M., and E. A. Johnson. 2013. Warming-induced upslope advance of subalpine forest is severely limited by geomorphic processes. Proceedings of the National Academy of Sciences of the United States of America 110:8117–22. doi:https://doi.org/10.1073/pnas.1221278110.
- Maher, E. L., and M. J. Germino. 2006. Microsite differentiation among conifer species during seedling establishment at alpine treeline. Ecoscience 13 (3):334–41. doi:https://doi.org/10.2980/i1195-6860-13-3-334.1.
- Müller, M., U. Schickhoff, T. Scholten, S. Drollinger, J. Bo Hner, and R. P. Chaudhary. 2015. How do soil properties affect alpine treelines? General principles in a global perspective and novel findings from Rolwaling Himal, Nepal. Progress in Physical Geography 40 (1):135–60. doi:https://doi.org/10.1177/0309133315615802.
- Müller, M., N. Schwab, U. Schickhoff, J. Böhner, and T. Scholten. 2016. Soil temperature and soil moisture patterns in a Himalayan alpine treeline ecotone. Arctic, Antarctic, and Alpine Research 48 (3):501–21. doi:https://doi.org/10.1657/AAAR0016-004.
- Oren, R., and D. W. Sherriff. 1995. Water and nutrient acquisition by roots and Canopies. In Resource physiology of conifers: Acquisition, allocation, and utilization, edited by W. K. Smith and T. M. Hinckley, 39–74. San Diego, CA: Academic Press.
- Perry, D. A., R. Oren, and S. C. Hart. 2008. Soil: The fundemental resource, forest ecosystems, 256–300. Baltimore: Johns Hopkins University Press.
- Pickles, B. J., B. D. Twieg, G. A. O’Neill, W. W. Mohn, and S. W. Simard. 2015. Local adaptation in migrated interior Douglas-fir seedlings is mediated by ectomycorrhizas and other soil factors. New Phytologist 207 (3):858–71. doi:https://doi.org/10.1111/nph.13360.
- Pinheiro, J., B. Bates, S. Debroy, D. Sarkar, and R Core Team. 2017. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-131. Available: https://CRAN.R-project.org/package=nlme
- Pisaric, M. F. J., C. Holt, J. M. Szeicz, T. Karst, and J. P. Smol. 2003. Holocene treeline dynamics in the mountains of northeastern British Columbia, Canada, inferred from fossil pollen and stomata. The Holocene 13:161–73. doi:https://doi.org/10.1191/0959683603hl599rp.
- Poschlod, P., M. Abedi, M. Bartelheimer, J. Drobnik, S. Rosbakh, and A. Saatkamp. 2013. Seed ecology and assembly rules in plant communities. In Vegetation Ecology, edited by E. van der Maarel and J. Franklin. Chichester: John Wiley & Sons.
- R Core Team. 2015. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
- Rosbakh, S., P. Poschlod, and N. Anten. 2015. Initial temperature of seed germination as related to species occurrence along a temperature gradient. Functional Ecology 29 (1):5–14. doi:https://doi.org/10.1111/1365-2435.12304.
- Roush, W. M. 2009. A substantial upward shift of the alpine treeline ecotone in the Southern Canadian Rocky Mountains, 164. Victoria, BC: Department of Geography, University of Victoria.
- Seastedt, T. R., W. D. Bowman, T. N. Caine, D. McKnight, A. Townsend, and M. W. Williams. 2004. The landscape continuum: A model for high-elevation ecosystems. Bioscience 54:111–21. doi:https://doi.org/10.1641/0006-3568(2004)054[0111:TLCAMF]2.0.CO;2.
- Simberloff, D. 2009. The role of propagule pressure in biological invasions. Annual Review of Ecology, Evolution, and Systematics 40 (1):81–102. doi:https://doi.org/10.1146/annurev.ecolsys.110308.120304.
- Smith, W. K., M. J. Germino, D. M. Johnson, and K. Reinhardt. 2009. The altitude of alpine treeline: A bellwether of climate change effects. The Botanical Review 75 (2):163–90. doi:https://doi.org/10.1007/s12229-009-9030-3.
- Ste-Marie, C., E. A. Nelson, A. Dabros, and M.-E. Bonneau. 2011. Assisted migration: Introduction to a multifaceted concept. The Forestry Chronicle 87 (6):724–30. doi:https://doi.org/10.5558/tfc2011-089.
- Sullivan, P. F., S. B. Ellison, R. W. McNown, A. H. Brownlee, and B. Sveinbjörnsson. 2015. Evidence of soil nutrient availability as the proximate constraint on growth of treeline trees in northwest Alaska. Ecology 96 (3):716–27. doi:https://doi.org/10.1890/14-0626.1.
- Theurillat, J.-P., and A. Guisan. 2001. Potential impact of climate change on vegetation in the European Alps: A review. Climatic Change 50:77–109. doi:https://doi.org/10.1023/A:1010632015572.
- Tingstad, L., S. L. Olsen, K. Klanderud, V. Vandvik, and M. Ohlson. 2015. Temperature, precipitation and biotic interactions as determinants of tree seedling recruitment across the tree line ecotone. Oecologia 179 (2):599–608. doi:https://doi.org/10.1007/s00442-015-3360-0.
- Urli, M., C. D. Brown, R. N. Perez, P.-L. Chagnon, and M. Vellend. 2016. Increased seedling establishment via enemy release at the upper elevational range limit of sugar maple. Ecology 97 (11):3058–69. doi:https://doi.org/10.1002/ecy.1566.
- Wagg, C., B. C. Husband, D. S. Green, H. B. Massicotte, and R. L. Peterson. 2011. Soil microbial communities from an elevational cline differ in their effect on conifer seedling growth. Plant and Soil 340:491–504. doi:https://doi.org/10.1007/s11104-010-0621-x.
- Wang, T., A. Hamann, D. L. Spittlehouse, and T. Q. Murdock. 2012. ClimateWNA: High-resolution spatial climate data for western North America. Journal of Applied Meteorology and Climatology 51:16–29. doi:https://doi.org/10.1175/JAMC-D-11-043.1.
- Westbrook, M. R., 2014. Local scale forest encroachment into alpine habitat: past patterns and future predictions. MSc Thesis, Department of Biological Sciences, University of Cincinnati, 74.
- Winder, R., E. A. Nelson, and T. Beardmore. 2011. Ecological implications for assisted migration in Canadian forests. The Forestry Chronicle 87 (6):731–44. doi:https://doi.org/10.5558/tfc2011-090.