
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Anthropogenic and biotic disturbances have the potential to interact, generating cumulative impacts on animal movement or, alternatively, counterbalancing or masking each other. Despite their importance, those interactions have not been investigated thoroughly. Our study aimed to fill this knowledge gap by assessing the combined effects of a human activity—that is, military exercises—and a biotic disturbance—that is, insect harassment—on movement rates of free-ranging semidomesticated reindeer (Rangifer tarandus tarandus). From 2010 to 2012, we analyzed location data from fifty-one Global Positioning System (GPS)-collared female reindeer in the largest European military test range, situated in northern Sweden. In the presence of both military exercises and mosquito harassment, reindeer reacted by increasing their movement rates but not as much as when mosquito harassment occurred alone. Conversely, reindeer reduced their movement rates during military exercises performed with aircraft. Moreover, the effect of military exercises performed with vehicles was evident only when combined with mosquito harassment. These results stress the value of evaluating the effects of the interaction between biotic disturbances and human activities, especially in northern ecosystems, because of the predicted climate warming and the growing interest toward natural resource extraction and other forms of land use.
Introduction
Today, most areas around the world are perturbed by human activities. For example, the contiguous United States are completely lacking areas free from anthropogenic noise (Barber et al. Citation2011). Similarly, the development and expansion of lighting technologies are making light pollution a spreading threat to natural systems around the world (Longcore and Rich Citation2004; Gaston et al. Citation2013). Moreover, the continuous expansion of the transport sector is generating levels of air and noise pollution that are turning into a global threat (Holden and Linnerud Citation2015; Lindgren and Wilewska-Bien Citation2016; Shannon et al. Citation2016). Human activities may displace and disturb animals by altering their habitat or affecting their behavior (Steidl and Powell Citation2006), with possible complex repercussions because animals can perceive human activities through diverse means, such as auditory, visual, and olfactory cues. The response is usually driven by a trade-off between the benefit of responding to the disturbance—that is, an increased chance of survival when the disturbance is perceived as a risk—and its cost; for example, displacement or reduced time available for parental care, mating, or feeding activities (Frid and Dill Citation2002). When the anthropogenic disturbance is strong and causes a reduction in the time spent feeding and resting, the cost of the response can be substantial and severely affect the animal’s survival and fitness (e.g., Luick et al. Citation1996). At the larger scale, human activities can end up affecting entire populations by displacing or confining them to low-quality habitats or habitats with higher predation risk (Frid and Dill Citation2002).
Anthropogenic disturbances can have long-lasting effects on arctic and subarctic ecosystems, due to the low biodiversity and harsh conditions of those systems (Forbes, Ebersole, and Strandberg Citation2001; Willard, Cooper, and Forbes Citation2007; Reynolds and Tenhunen Citation2013). Nonetheless, the rising global interests in natural resources such as forest products, minerals, and gas and oil have driven an intensification in human activities and infrastructure development in those ecosystems (Nellemann et al. Citation2001; Raynolds et al. Citation2014), which imposes costs for the animals inhabiting them (Johnson et al. Citation2005). Moreover, climate change is manifesting in the arctic and subarctic regions at faster rates than in any other area in the world (IPCC Citation2014). Arctic and subarctic ecosystems are shaped by their cold and harsh climate, and global warming is expected to have acute effects on the structure and functioning of their simple but sensitive food webs (Ims and Fuglei Citation2005; Stempniewicz, Błachowiak-Samołyk, and Węsławski Citation2007; Legagneux et al. Citation2014; Barthelemy et al. Citation2017). Even small disturbances directly affecting only one species may have widespread, indirect effects on a variety of organisms (Croll et al. Citation2005; Post et al. Citation2009). Direct anthropogenic disturbances and climate change have the potential to have interacting effects on species living in arctic and subarctic ecosystems, but that interaction has so far received little attention (but see Raynolds et al. Citation2014).
Reindeer (Rangifer tarandus, a species called caribou in North America) are the most abundant and important large herbivore in arctic and subarctic ecosystems. Reindeer regulate plant communities through grazing and are the main prey for several predator species (Pedersen et al. Citation1999; van der Wal Citation2006; Musiani et al. Citation2007; Andren et al. Citation2011; Hobbs et al. Citation2012; Bernes et al. Citation2015). Moreover, reindeer constitute the main source of subsistence for at least twenty indigenous peoples (Turi Citation2002; Forbes et al. Citation2006, Citation2009; Forbes Citation2013). Both wild and semidomesticated reindeer are integral parts of northern ecosystems (Uboni et al. Citation2016), because during most of the year semidomesticated reindeer range freely on natural pastures (Forbes and Kumpula Citation2009). Various human activities and infrastructures affect the behavior, movement, and habitat selection of wild and semidomesticated reindeer and caribou; for example, by increasing movement speed and distance traveled, by preventing animals from visiting high-quality pastures, or by increasing the time spent in vigilance behavior or traveling, which may decrease feeding time (Vistnes and Nellemann Citation2008; Skarin and Åhman Citation2014). Among those activities and infrastructures, we find tourism and hunting (e.g., Vistnes et al. Citation2008; Reimers et al. Citation2009; Skarin et al. Citation2010; Wilson and Wilmshurst Citation2019), vehicle traffic, helicopters and aircraft (Harrington Citation2003; Reimers and Colman Citation2006), and drill sites, mines, dams, and various kinds of power plants (e.g., Fancy Citation1983; Murphy and Curatolo Citation1987; Panzacchi et al. Citation2013; Skarin et al. Citation2015; Skarin, Sandström, and Alam Citation2018). Most studies so far have addressed only the short-term effects of those activities and infrastructures on reindeer behavior, but some have demonstrated that their impact goes beyond movement and habitat selection, affecting reproduction rates and calf survival (Luick et al. Citation1996; Vistnes and Nellemann Citation2008).
Harassment from insects is another well-known factor influencing reindeer behavior and fitness. Mosquitoes (especially Aedes spp., Culicidae), blackflies (Simuliidae), horseflies (Tabanidae), warble flies (Hypoderma tarandi L., Oestridae), and nose bot flies (Cephenemyia trompe L., Oestridae; the latter two hereafter referred to as oestrid flies) commonly attack reindeer in summer. Oestrid flies cause the strongest harassment, considerably affecting reindeer activity patterns (Hagemoen and Reimers Citation2002). Oestrid flies are indeed excellent and fast fliers and easily detect and follow reindeer (A. C. Nilssen and Anderson Citation1995), and reindeer often exhibit violent reactions to fly harassment, running for hours in search for relief (Hagemoen and Reimers Citation2002). Mosquitoes also have the ability to alter reindeer behavior (Fancy Citation1983; Downes, Theberge, and Smith Citation1986; Mörschel and Klein Citation1997; Noel et al. Citation1998; Raponi et al. Citation2018). In northern Scandinavia, oestrid activity occurs approximately between early July and early September (Anderson, Nilssen, and Folstad Citation1994; A. C. Nilssen Citation1997; Anderson et al. Citation2001), and mosquitoes are mainly active between June and August (Schäfer and Lundström Citation2001). During periods of intense insect harassment, reindeer search for relief in fresh and windy areas such as snow patches, hilltops, ridges, mires, open areas, roads, and stream estuaries (Fancy Citation1983; Helle and Aspi Citation1984; Downes, Theberge, and Smith Citation1986; Pollard et al. Citation1996; Noel et al. Citation1998; Skarin et al. Citation2004; Moen Citation2008; Vistnes et al. Citation2008). In those areas, reindeer are expected to move less, or slower, than in areas with high insect harassment. Overall, intense insect harassment may have dramatic consequences for reindeer health, body conditions, and fecundity (Hagemoen and Reimers Citation2002). In some cases, reindeer may even increase their tolerance to anthropogenic disturbances if insect harassment is severe. For example, reindeer are expected to avoid areas used seasonally by humans, such as mountain tourist huts and trails (Skarin Citation2007). However, in the Scandinavian Mountains, reindeer move to higher elevations in July and August to escape insect harassment even when those areas are heavily used by tourists (Skarin et al. Citation2004). Thus, the necessity to escape insects seems to override the need for human avoidance. Fancy (Citation1983) determined that caribou movement rates did not differ between areas characterized by high human disturbance (drill sites) and control areas under either low or high insect activity levels. Additionally, the effects of the disturbance generated by oilfields on caribou behavior are more pronounced in the absence of insects (Murphy and Curatolo Citation1987). Caribou may tolerate some level of human disturbance because drill sites are open and windy areas; that is, optimal insect relief areas (Fancy Citation1983; Pollard et al. Citation1996; Noel et al. Citation1998). Reindeer response to other types of human disturbance may also be overridden by insect harassment, and if the two disturbances are not analyzed simultaneously, their effects may be underestimated or misinterpreted.
Because of their essential role in arctic and subarctic ecosystems and their vulnerability to human encroachment in those ecosystems (Uboni et al. Citation2016), we chose free-ranging semidomesticated reindeer as a study species to investigate the potential combined effects of a human activity—that is, military exercises—and insect harassment on animal movements. Our study system was located in a military test range, which also overlapped the reindeer summer grazing grounds. Specifically, our aims were to assess whether (1) reindeer move away from their traditional summer range while military exercises occur and (2) such military exercises and insect harassment interact in affecting reindeer movement. We hypothesized that reindeer would react to human presence and loud noises due to military exercises by relocating from the test range. Moreover, we hypothesized that the effect of military exercises may be masked by insect harassment when the two disturbances occur simultaneously.
Methods
Study area
The study was conducted in the range of the Udtja reindeer herd, belonging to the Udtja reindeer herding community, Norrbotten County, northern Sweden (). In Udtja, reindeer spend all year in the boreal forest. Nonetheless, they have distinct summer and winter ranges and range freely within the borders of the community, controlled by a fence toward the northwest and by active work of the herders in the southeast (Swedish Sami Parliament Citation2019; https://www.sametinget.se/8744). The summer range is dominated by old forest, constituted primarily by Scots pine (Pinus sylvestris) and Norway spruce (Picea abies), interspersed with mires, lakes, and, on mountain tops, downy birch (Betula pubescens) forest. Elevation ranges between 187 and 714 m.a.s.l., and roads are concentrated toward the southeast at rather low densities (gravel roads: 0.25 km/km2; paved roads: 0.02 km/km2). Monthly temperatures in July–September averaged 10.13°C (±3.31) in the period 2006–2016, and the precipitation monthly sum for the same months averaged 69.78 mm (±37.74) in the period 2013–2016 (Swedish Hydrological and Meteorological Institute Citation2017; Tjåkaape weather station, http://opendata-downloadmetobs.smhi.se/explore/). A military test range, the Vidsel Test Range, was established in 1959 within the Udtja spring and summer reindeer ranges. There, the main human activity is represented by military training exercises. With its 1,625 km2 of ground space, this area is Europe’s largest military test range. Roads inside the test range are used only in conjunction with the military exercises. Outside the test range, main roads are used by regular traffic, and most secondary roads are predominantly used in conjunction with forestry activities. Inside the test range, 1,465 km2 were designated in 1995 as a nature reserve where logging activities are not permitted (). The main reindeer predator in the area is the brown bear (Ursus arctos), but European lynx (Lynx lynx), wolverine (Gulo gulo), and golden eagle (Aquila chrysaetos) also make up part of the predation (Sivertsen Citation2017).
Figure 1. This study was conducted in the Udtja reindeer herding community, northern Sweden. © Lantmäteriet, Sametinget (www.sametinget.se/8744). ArcGIS World Imagery basemap source: Esri, DigitalGlobe, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AEX, Getmapping, Aerogrid, IGN, IGP, swisstopo, and the GIS user community
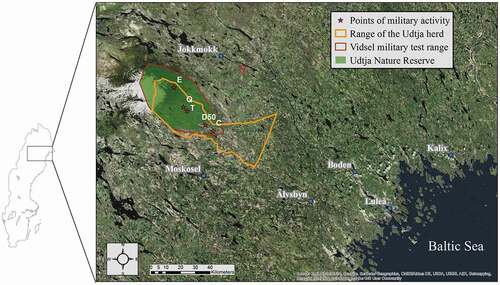
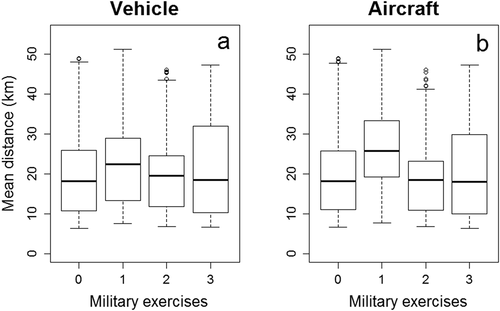
Figure 2. Differences in the daily average Euclidean distance (in kilometers) between reindeer locations and the points of military activity (y-axes) in the absence or presence of military exercises (x-axes) in the Vidsel test range, Sweden, from 1 August to 10 September, 2010–2012. The occurrence of military exercises is noted as follows: 0 = absence; 1 = exercises performed at one activity point; 2 = exercises performed at two activity points; 3 = exercises performed at three activity points. (a) Military exercises performed with terrestrial vehicles and (b) military exercises performed with aircraft
Data collection
Reindeer Global Positioning System data
Between 2010 and 2012, we fitted seventy-one adult female reindeer (n = 19 in 2010, n = 27 in 2011, n = 25 in 2012) with Global Positioning System (GPS) collars (Followit AB, Tellus) in the Utdja reindeer herding community and downloaded the data manually in the field once a year during the winter gatherings of the animals. The collars were programmed to collect GPS locations at 2-hour intervals and the average GPS fix rate was 86.2 percent. Subsequently, we removed six individuals from the analysis because of collar failure (n = 2 in 2010, n = 3 in 2011, n = 1 in 2012) and fourteen individuals because their fix rate was <80 percent (n = 5 in 2010, n = 4 in 2011, n = 5 in 2012), which left fifty-one reindeer for analysis (n = 12 in 2010, n = 20 in 2011, n = 19 in 2012). Of those, forty-nine were fitted with collars for one year and two were followed for two years. We cleaned the GPS data set by removing obvious outliers (i.e., locations falling outside the study area, detected based on latitude and longitude), locations with a two-dimensional accuracy or a dilution of precision > 5, duplicates, and clear erroneous locations identified based on movement speed. Because herders gather their reindeer for calf marking prior to 12 July and insect harassment in northern Scandinavia occurs from early July up to early autumn (Anderson, Nilssen, and Folstad Citation1994; A. C. Nilssen Citation1997; Anderson et al. Citation2001; Skarin et al. Citation2010), we restricted the study period to 12 July to 10 September.
Military exercises
We obtained daily data on military exercises performed at the Vidsel Test Range in 2010–2012 by the Swedish Defense Materiel Administration in the form of occurrence of military exercises aiming at three locations (E, Q, and T) within the test range (), hereafter defined as activity points. Due to the classified nature of those data, the spatial extent of the military exercises was unknown. The exercises were performed with light (e.g., cars and trucks) and heavy terrestrial vehicles (e.g., tanks) or with aircraft. Because military exercises performed with light and heavy terrestrial vehicles always occurred simultaneously, we hereafter refer to those exercises as being performed with vehicles. Vehicles were usually driven toward one of the activity points from points C or D50 (), whereas aircraft flew into the test range from a nearby military airport in the southeast. No mitigation measures were adopted in relation to disturbance to reindeer.
Insect harassment indices
Fly and mosquito harassment are positively correlated with air temperature and negatively correlated with wind speed (e.g., Hagemoen and Reimers Citation2002). Moreover, fly harassment is negatively correlated with cloud cover (Anderson, Nilssen, and Folstad Citation1994; Colman et al. Citation2003). The intensity of harassment by flies and mosquitoes on ungulates can be summarized by an insect harassment index (IHI; Weladji, Holand, and Almøy Citation2003) when the weather conditions of a certain area at a certain time and the threshold of insect activity (in terms of temperature, wind speed, and cloud cover) are known. For example, according to Weladji, Holand, and Almøy (Citation2003), oestrid flies are active at air temperatures ≥13°C, wind speeds <6 m/s, and cloud cover <40 percent. Based on that information, Weladji, Holand, and Almøy (Citation2003) developed an oestrid harassment index (OHI):
where PHD, the harassment day factor, indicates a day with midday temperature above the threshold of fly activity and wind speed below the threshold, and S, the severity factor, reflects cloud cover below the threshold.
Using hourly weather data provided by the Swedish Hydrological and Meteorological Institute (Citation2017; Tjåkaape weather station, http://opendata-downloadmetobs.smhi.se/explore/), we calculated an OHI and mosquito harassment index (MHI) for our study period and area. However, in order to develop IHIs that are as precise as possible, we adapted Weladji, Holand, and Almøy’s (Citation2003) index by averaging the wide range of values reported in the literature for temperature and wind speed thresholds of oestrid and mosquito activity (Table S1). We calculated the OHI (ranging from 0 to 1.5) based on PHD = 1 on a day with temperature >13° C and wind speed <9.6 m/s and S = 0.5 on days with cloud cover <40 percent. The MHI could only take a value of 1 when the temperature was between 7°C and 17°C and the wind speed was <6.12 m/s or a value of 0 otherwise, because there is no evidence of a relationship between cloud cover and mosquito activity. The IHIs were not validated in our study area. Because oestrid flies are obligate parasites of reindeer and reindeer are the most abundant large mammal that mosquitoes can feed upon in northern Scandinavia (except humans), when those insects are active, reindeer are likely one of their main targets. Moreover, our study covers the period of activity of adult oestrid flies and mosquitoes in northern Scandinavia (Anderson, Nilssen, and Folstad Citation1994; A. C. Nilssen Citation1997; Schäfer and Lundström Citation2001), and insect activity is highly dependent on meteorological conditions, which the indices take into account. Therefore, we assume that the IHIs are accurate proxies for insect harassment on reindeer.
Environmental and anthropogenic variables
Based on previous studies (Skarin et al. Citation2008, Citation2010), we collected six environmental and anthropogenic variables to assess drivers of reindeer movement (see models below): (1) vegetation type; (2) elevation; (3) slope; (4) ruggedness; (5) the Normalized Difference Vegetation Index (NDVI); and (6) the Euclidean distance between each reindeer location and the nearest road. We obtained maps of vegetation type, roads, and elevation (50-m-resolution digital elevation model) from Lantmäteriet (Citation2018; www.lantmateriet.se). We derived slope and ruggedness from the digital elevation model using the slope and vector ruggedness measure (Sappington, Longshore, and Thompson Citation2007) tools in a geographic information systems (GIS) environment (QGIS 3.2.0; QGIS Development Team Citation2018). We obtained twelve sixteen-day composite NDVI maps (MODIS Terra Vegetation Index 250-m resolution) that covered the entire study period from NASA Earthdata Search website (Citation2018; https://search.earthdata.nasa.gov/). Because the original NDVI maps had a 200-m resolution, we resampled them at 50-m resolution.
Data analysis
Reindeer response to military exercises
To assess whether reindeer responded to military exercises by moving away from the military activity points, we developed univariate regression models in which the daily average Euclidean distance (in meters) of each reindeer from the three activity points was the response variable (calculated in QGIS 3.2.0; QGIS Development Team Citation2018). The predictor categorical variable described the occurrence of military exercises performed either with vehicles or with aircraft (Vehi_ALL and Air_ALL, respectively), each ranging from 0 (i.e., no military exercises performed in a given day) to 3 (i.e., military exercises performed at all three activity points). Due to collinearity between Vehi_ALL and Air_ALL (pairwise Pearson’s correlation coefficient: r = 0.81), we did not include both variables in the same models. For each type of military activity, we compared a set of six models: (1) a generalized least squares (GLS) model; (2) a generalized linear mixed effects model (GLMM) with reindeer ID as random intercept term; (3) a GLMM with year as a random term; (4) a GLMM with reindeer ID and year as nested random terms; (5) a GLS with an autoregressive correlation structure, AR1; and (6) a GLMM with reindeer ID as random intercept term and an autoregressive correlation structure, AR1. We evaluated model performance based on the Akaike information criterion (AIC; Burnham and Anderson Citation2002). Because no military exercises occurred in July, we limited this analysis to 1 August to 10 September. Furthermore, we only included those reindeer that spent at least part of the study period inside the test range (n = 10 in 2010, n = 7 in 2011, n = 11 in 2012).
Effects of insect harassment and military exercises on reindeer movement
To assess whether insect harassment and military exercises affected reindeer movement, we fitted GLS models and GLMMs using the Euclidean distance traveled by a reindeer between successive GPS locations (hereafter referred to as movement rate but also defined as step length) as the response variable. The predictor variables were Vehi_ALL, Air_ALL, OHI, MHI, temperature, elevation, slope, ruggedness, vegetation type, NDVI, distance from roads, and area (i.e., a binary variable indicating whether the reindeer was moving inside or outside the military test range). Thus, we did not measure insect harassment directly but inferred it from the IHIs. Movement rates were not estimated if one or more consecutive GPS locations were missing. Because reindeer movement rates were temporally autocorrelated and affected by circadian rhythm (Online Resource, Figures S1 and S2; but see also Erriksson, Källqvist, and Mossing Citation1981; van Oort et al. Citation2005), we followed Dray, Royer-Carenzi, and Calenge (Citation2010) by using the nb2listw function in the R package “spdep” to define a spatial weighting object to represent the temporal structure of the data (Bivand, Hauke, and Kossowski Citation2013; Bivand and Piras Citation2015). Subsequently, we detrended the data through the principal coordinate analysis of neighbor matrices (for details, see Borcard and Legendre Citation2002; Dray, Legendre, and Peres-Neto Citation2006) and used the detrended residuals as response variable in our models. We excluded from detrending twenty reindeer (n = 7 in 2010, n = 9 in 2011, n = 4 in 2012) because the procedure did not support the low temporal autocorrelation in their movement rates. Therefore, the final data set for this analysis included thirty-one female years (corresponding to twenty-nine female reindeer, because for two were followed two consecutive years).
We treated Vehi_ALL, Air_ALL, OHI, and MHI as continuous variables. We averaged OHI, MHI, and temperature on a 2-hour interval to match the GPS fix interval. We calculated elevation, slope, and ruggedness as the mean of the values along each step. We assigned temporally and/or spatially specific values of vegetation type, NDVI, and distance from roads to each step based on the first GPS location in the step. Lastly, we included interaction terms between the IHIs and (1) the military exercises variables, (2) vegetation type, (3) elevation, and (4) distance from roads as well as area and (1) distance from roads and (2) the military exercises variables. This latter term aimed to assess whether reindeer reacted differently to military exercises whether the reindeer was close to or far from the areas where the exercises took place. The interactions between the IHIs, vegetation type, and elevation aimed to determine whether reindeer used insect relief areas in response to insect harassment. We screened all predictor variables for collinearity using a pairwise Pearson’s correlation coefficient (threshold for removal: r ≥ 0.8), followed by the variance inflation factors (threshold for removal: variance inflation factor ≥3; Zuur, Ieno, and Elphick Citation2010). We fitted four separate sets of models, one for each combination of IHI and type of military exercise, because of high correlation between OHI and MHI (r = 0.81; Online Resource, Table S2) and between the military exercises performed with vehicles and aircraft (r = 0.83). Each set of models included (1) a GLS model with no random term, fixed variance, or autocorrelation structure; (2) three GLMMs with either reindeer ID, year, or reindeer ID/year as random intercept terms; (3) two GLS models with either year or month as a fixed variance structure; and (4) a GLS model with year as a fixed variance structure and with an exponential spatial autocorrelation structure applied to the model residuals (range = 850, nugget = 0.4, sill = 1). We tested reindeer ID as a random intercept term to exclude the influence of differences in behavior and sample size among individual animals (Gillies et al. Citation2006). We added year and month as fixed variance structures to address heteroscedasticity and temporal variability (Gillies et al. Citation2006; Uboni, Smith et al. Citation2015; Uboni, Vucetich et al. Citation2015). We tested the exponential spatial correlation structure to address spatial autocorrelation caused by landscape features such as topography and vegetation density, which could restrict or ease reindeer movement (Skarin et al. Citation2010; Panzacchi et al. Citation2016). We identified the best-fit model based on AIC and used restricted maximum likelihood estimation in all models for their AICs to be comparable (Zuur et al. Citation2009).
Once the need for a random term, a variance structure, and a correlation structure were determined, we performed an automatic backward model selection procedure using the stepAIC function in the “MASS” package for R (Venables and Ripley Citation2002). In order to compare models with different fixed terms, we refitted the full model using maximum likelihood estimation. Then, once the most parsimonious fixed structure was found, we refitted the reduced model with restricted maximum likelihood (Zuur et al. Citation2009).
Results
Reindeer response to military exercises
According to the GLS with an autoregressive correlation structure (Online Resource, Table S3), the average distance between reindeer and activity points increased only when military exercises were performed with aircraft at one activity point (β = 1,110.73, SE = 257.26, p = .0005; all other ps > .05; ).
Effects of insect harassment and military exercises on reindeer movement
All best-fit models aimed to assess the effect of insect harassment and military exercises on reindeer movement rates included year as a variance structure and a spatial correlation structure but no random term (Online Resource, Table S4).
Reindeer movement rates increased in the predicted presence of mosquitoes, as they did, though to a lesser extent, when mosquito harassment and military exercises occurred simultaneously. On the contrary, reindeer movement rates decreased in response to military exercises performed with aircraft. Reindeer generally moved faster outside the test range and at higher elevations. Reindeer movement rates decreased with increasing distance to roads and steeper slopes, the former especially outside the test range (). Vegetation type, temperature, ruggedness, and NDVI did not seem to influence reindeer movement rates in the Udtja reindeer herding community.
Table 1. Effects of military exercises, insect harassment, and environmental and anthropogenic predictor variables on reindeer movement rates
Discussion
Movement rates of free-ranging, semidomesticated reindeer increased in the predicted presence of mosquito harassment, which was inferred from the MHI, whereas they decreased in response to the military exercises. When those two disturbances occurred simultaneously, reindeer response was intermediate. We hypothesize that in the presence of military exercises reindeer were not able to move as fast as they usually do when escaping harassing insects. At the same time, the effect of military exercises was masked when occurring simultaneously with insect harassment. These results stress the importance of assessing the impact of anthropogenic disturbances on animal movement behavior in relation to other biotic or abiotic stressors. Indeed, during our study, military exercises not only altered reindeer movement directly but also altered the reindeer response to insect harassment. A wide range of research has explored the ways in which human activities affect animal behavior (e.g., Longcore and Rich Citation2004; Steidl and Powell [Citation2006] and references therein; Gaston et al. Citation2013; Shannon et al. Citation2016); similarly, many studies have assessed the effects of biotic and abiotic factors on animal movement (e.g., Avgar et al. Citation2013; Michel et al. Citation2013). However, few studies have investigated the co-occurrence of both sets of stressors on animal movement (Fancy Citation1983; Pollard et al. Citation1996; Noel et al. Citation1998), especially when their intensity varies over time. Our study reveals that the effects of those processes on animal movement can interact with each other and that analyzing them separately may lead to a misinterpretation of the complexity of their effects (Sih, Bell, and Kerby Citation2004). Moreover, the impact of human activities may be overlooked when those activities are performed at a time of high environmental stress for the animals. The biology of the animal should thus be considered in relation to the timing of an anthropogenic disturbance. In our study area, for example, the disturbance created by military exercises comes into play during a time when female reindeer are particularly susceptible to disturbance, because they are building up fat reserves for the winter and they are still lactating to feed their calves (Vistnes and Nellemann Citation2001; Harrington Citation2003; Skarin et al. Citation2008, Citation2015). Lactation requires a high amount of energy (White Citation1992); nonetheless, lactating female reindeer prefer an undisturbed environment at the expense of forage quality (Maier et al. Citation1998; Helle et al. Citation2012). If human disturbances occur at the time and in the areas of severe insect harassment, the consequences for female reindeer and their calves may be detrimental. Their increased movement rates translate into increased energy expenditure and, more important, harassed reindeer spend significantly less time feeding (Colman et al. Citation2003), decreasing their possibility to build up energy reserves in preparation for winter (Åhman and White Citation2018). As a consequence, heavily harassed reindeer may enter the winter in poor body conditions, with implications for their survival and reproductive success (Luick et al. Citation1996; Mallory and Boyce Citation2018). Moreover, cohorts that suffer nutritional deficit in their first year of life may experience reduced lifetime reproductive success (Albon, Clutton-Brock, and Guinness Citation1987; Gaillard et al. Citation2000; Couturier et al. Citation2009).
The repercussions of the interacting effects of anthropogenic and biotic disturbances on reindeer behavior and health may be exacerbated in the future as a consequence of the expansion of human activities and the increase in temperature predicted in northern ecosystems (Witter et al. Citation2012). Currently, insect harassment in arctic and subarctic regions is most severe in the central and hottest summer months, July and August. If the projected increase in temperatures occurs, the time of insect harassment may expand to late spring, when the calves are small and most vulnerable. Moreover, climate change is predicted to favor the expansion of new parasitic organisms into those regions (Mallory and Boyce Citation2018). Increased harassment may then start a negative feedback loop, because animals are even more sensitive to disturbances when they suffer nutritional stress. Coupled with increasing combined disturbances caused by various human activities (Vistnes and Nellemann Citation2008; Skarin and Åhman Citation2014) and the negative effects of increased temperatures on reindeer physiology (K. J. Nilssen et al. Citation1984), increased harassment by insects and other parasites may have severe consequences for the species. Such disturbances may have complex repercussions on a wide range of species and environmental processes because of the key role that reindeer play in northern ecosystems (Pedersen et al. Citation1999; van der Wal Citation2006; Musiani et al. Citation2007; Andren et al. Citation2011; Hobbs et al. Citation2012; Bernes et al. Citation2015).
During the study period, female reindeer did not generally react to the military exercises by moving away from the areas where those activities were performed. On the one hand, reindeer may have traded off a peaceful environment for good quality pastures, especially because the disturbance created by military exercises was occasional and might not have been frequent enough to cause relocation of the reindeer (Gill, Norris, and Sutherland Citation2001). Woodland caribou (Harrington and Veitch Citation1991), peregrine and prairie falcons (Falco peregrinus and Falco mexicanus, respectively; Ellis, Ellis, and Mindell Citation1991), harlequin ducks (Histrionicus histrionicus; Goudie and Jones Citation2004), and red-cockaded woodpeckers (Picoides borealis; Delaney et al. Citation2011) tend to react similarly to military exercises. On the other hand, reindeer in the Udtja reindeer herding community may have developed some level of habituation to the military exercises, especially those performed with vehicles. However, habituation should not be considered as a positive outcome, because animals that are habituated to vehicles are more likely to be victims of traffic accidents (Bejder et al. Citation2009). Additional studies are needed to determine whether reindeer have developed habituation to the military exercise but also to assess whether the effects of long and repeated military exercises on movement rates may translate into long-term consequences for the animal health, body conditions, and reproductive success, which could influence population growth and, in the case of reindeer husbandry, the local economy based on meat production.
Reindeer response to insect harassment (in terms of movement rates) did not differ depending on vegetation type, elevation, and distance from roads. Indeed, none of the interactions between the IHIs and those variables were retained in the reduced regression models (). Reindeer often use open areas, higher elevations, and roads as insect relief areas (Helle and Aspi Citation1984; Downes, Theberge, and Smith Citation1986; Skarin et al. Citation2004; Moen Citation2008; Vistnes et al. Citation2008). In our study area, the maximum elevation is rather low (714 m.a.s.l.) and reindeer may not have access to many treeless hilltops. Indeed, in northern Scandinavia the treeline occurs at around 500–700 m. Moreover, roads have low density, which probably prevents reindeer from using those features to escape insect harassment. If roads had been used as insect relief areas, reindeer movement rates should have been lower in their proximity. On the contrary, reindeer movement rates increased with decreasing distance from roads (). We suggest that reindeer may use roads as movement corridors or they may perceive roads as a source of disturbance.
Female reindeer reacted to military exercises performed with aircraft by reducing their movement rates. This behavior may have originated as a response to a potential predator flying overhead. Golden eagles may indeed attack both adult females and calves (Nybakk, Kjelvik, and Kvam Citation1999; Nybakk et al. Citation2002; Gustine et al. Citation2006; Norberg et al. Citation2006; Nieminen, Norberg, and Maijala Citation2011). Studies analyzing the antipredator response of reindeer to attacks by raptors are lacking in the literature, but attacks on other ungulates have been observed in some instances. Females of several ungulate species, such as Thomson’s gazelle (Gazella thomsonii), pronghorn (Antilocapra americana), bighorn sheep, mountain goat (Oreamnos americanus), chamois (Rupicapra rupicapra), and Dall’s sheep (Ovis dalli dalli), protect their young from raptor attacks by standing still (Nette, Burles, and Hoefs Citation1984; Locati Citation1990; Byers Citation1997; Bertolino Citation2003; Hamel and Côté Citation2009; Roberts Citation2014). Moreover, forest-dwelling reindeer are less gregarious than their conspecifics living in the tundra, which respond to predator attacks by grouping together (Baskin Citation1986; Bergerud Citation1988; Skarin and Åhman Citation2014). Dissimilarly, woodland and barren-ground caribou usually react to aircraft with a panic response, which results in overall increased activity levels and movement rates (Calef, DeBock, and Lortie Citation1976; Maier et al. Citation1998; Harrington Citation2003). Forest-dwelling reindeer might feel more protected from a predator flying above while hiding in the forest and move less to decrease the chances of being detected. Eagles usually attack and successfully kill ungulates in open areas (but see Kerley and Slaght Citation2013), and most predation events caused by eagles on reindeer occur in open areas (Nieminen, Norberg, and Maijala Citation2011). Ultimately, the response of reindeer to aircraft observed in this study may also depend on the occasional herding of the reindeer with helicopter performed by the herders in the study area, which may have made the animals less prone to acute responses to flights, as well as on the domestication process (Reimers, Røed, and Colman Citation2012). Although the behavior observed in our study area does not necessarily imply increased energy expenditure due to increased movement, reindeer may reduce feeding time by staying alert until the flight activity ends, which may negatively affect their body conditions (Åhman and White Citation2018).
Reindeer moved faster outside compared to inside the military test range. This result may be due to environmental differences between the two areas. Indeed, most of the military test range overlaps with a nature reserve where logging has been prohibited since 1995, making the forests denser and possibly animal movement slower. Additionally, the topography is more complex inside the test range compared to outside. Alternatively, the difference we detected in movement rates between inside and outside the test range may be due to the military exercises. Indeed, generally reindeer moved slower during military exercises performed with aircraft, except when outside the test range. In our study, the spatial extent of the military exercises was unknown. More precise data would allow for a better understanding of the effect of military exercises on reindeer movement, especially in the case of exercises performed with vehicles (where we did not detect any effect on reindeer movement), because they may be performed within a smaller area compared to aircraft exercises.
Based on the results of this study, we suggest that environmental impact assessments should always consider the combined and cumulative effects of planned and preexisting human activities and infrastructures, together with the conditions of the animals that might be affected, the location of insect relief areas, and the projected impacts of climate change. This is particularly important in arctic and subarctic ecosystems, because they are expected to experience some of the most pronounced changes in climatic conditions based on the current predictions (IPCC Citation2014), and the species inhabiting them are adapted to their harsh conditions, such as low nutrient availability and cold temperatures (Williams, Henry, and Sinclair Citation2015). Adding direct anthropogenic disturbance on top of the impacts of climate change may cause irreversible damage to northern species and ecosystems.
Supplemental Material
Download Zip (155.8 KB)Acknowledgments
We thank Stéphane Dray for thorough explanations regarding the detrending procedure, Giovanna Jona Lasinio for suggestions on the statistical analyses, and Pablo Garrido for fruitful discussions about the interpretation of the study results. We are thankful to the Udtja reindeer herding community for providing access to the GPS data and for invaluable information about how reindeer herding was practiced in the area throughout the study period. Moreover, we thank the Swedish Defense Materiel Administration for sharing data on military exercises.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
Related Research Data
References
- Åhman, B., and R. C. White. 2018. Rangifer diet and nutritional needs. In Reindeer and Caribou: Health and disease, ed. M. Tryland and S. J. Kutz. Boca Raton, FL: CRC Press.
- Albon, S., T. Clutton-Brock, and F. Guinness. 1987. Early development and population dynamics in red deer. II. Density-independent effects and cohort variation. The Journal of Animal Ecology 56:69–81. doi:https://doi.org/10.2307/4800.
- Anderson, J. R., A. Nilssen, and I. Folstad. 1994. Mating behavior and thermoregulation of the reindeer warble fly, Hypoderma tarandi L. (Diptera: Oestridae). Journal of Insect Behavior 7:679–706. doi:https://doi.org/10.1007/BF01997439.
- Anderson, J. R., A. C. Nilssen, and W. Hemmingsen. 2001. Use of host-mimicking trap catches to determine which parasitic flies attack reindeer, Rangifer tarandus, under different climatic conditions. The Canadian Field-Naturalist 115:274–86.
- Andren, H., J. Persson, J. Mattisson, and A. C. Danell. 2011. Modelling the combined effect of an obligate predator and a facultative predator on a common prey: Lynx Lynx lynx and wolverine Gulo gulo predation on reindeer Rangifer tarandus. Wildlife Biology 17:33–43. doi:https://doi.org/10.2981/10-065.
- Avgar, T., A. Mosser, G. S. Brown, and J. M. Fryxell. 2013. Environmental and individual drivers of animal movement patterns across a wide geographical gradient. Journal of Animal Ecology 82:96–106. doi:https://doi.org/10.1111/j.1365-2656.2012.02035.x.
- Barber, J. R., C. L. Burdett, S. E. Reed, K. A. Warner, C. Formichella, K. R. Crooks, D. M. Theobald, and K. M. Fristrup. 2011. Anthropogenic noise exposure in protected natural areas: Estimating the scale of ecological consequences. Landscape Ecology 26:1281. doi:https://doi.org/10.1007/s10980-011-9646-7.
- Barthelemy, H., S. Stark, M. M. Kytöviita, and J. Olofsson. 2017. Grazing decreases N partitioning among coexisting plant species. Functional Ecology 31:2051–60. doi:https://doi.org/10.1111/1365-2435.12917.
- Baskin, L. M. 1986. Differences in the ecology and behaviour of reindeer populations in the USSR. Rangifer 6:333–40. doi:https://doi.org/10.7557/2.6.2.667.
- Bejder, L., A. Samuels, H. Whitehead, H. Finn, and S. Allen. 2009. Impact assessment research: Use and misuse of habituation, sensitisation and tolerance in describing wildlife responses to anthropogenic stimuli. Marine Ecology Progress Series 395:177–85. doi:https://doi.org/10.3354/meps07979.
- Bergerud, A. T. 1988. Caribou, wolves and man. Trends in Ecology & Evolution 3:68–72. doi:https://doi.org/10.1016/0169-5347(88)90019-5.
- Bernes, C., K. A. Bråthen, B. C. Forbes, J. D. Speed, and J. Moen. 2015. What are the impacts of reindeer/caribou (Rangifer tarandus L.) on arctic and alpine vegetation? A systematic review. Environmental Evidence 4:4. doi:https://doi.org/10.1186/s13750-014-0030-3.
- Bertolino, S. 2003. Herd defensive behaviour of chamois, Rupicapra rupicapra, in response to predation on the young by a golden eagle, Aquila chrysaetos. Zeitschrift Für Jagdwissenschaft 49:233–36.
- Bivand, R., and G. Piras. 2015. Comparing implementations of estimation methods for spatial econometrics. Journal of Statistical Software 63:1–36. doi:https://doi.org/10.18637/jss.v063.i18.
- Bivand, R., J. Hauke, and T. Kossowski. 2013. Computing the Jacobian in Gaussian spatial autoregressive models: An illustrated comparison of available methods. Geographical Analysis 45:150–79. doi:https://doi.org/10.1111/gean.2013.45.issue-2.
- Borcard, D., and P. Legendre. 2002. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecological Modelling 153:51–68. doi:https://doi.org/10.1016/S0304-3800(01)00501-4.
- Burnham, K. P., and D. R. Anderson. 2002. Model selection and multi-model inference: A practical information-theoretic approach. New York: Springer-Verlag.
- Byers, J. A. 1997. American pronghorn: Social adaptations and the ghosts of predators past. Chicago, IL: University of Chicago Press.
- Calef, G. W., E. A. DeBock, and G. M. Lortie. 1976. The reaction of barren-ground caribou to aircraft. Arctic 29:201–12. doi:https://doi.org/10.14430/arctic2805.
- Colman, J. E., C. Pedersen, D. Ø. Hjermann, Ø. Holand, S. R. Moe, and E. Reimers. 2003. Do wild reindeer exhibit grazing compensation during insect harassment? The Journal of Wildlife Management 67:11–19. doi:https://doi.org/10.2307/3803056.
- Couturier, S., S. D. Côté, R. D. Otto, R. B. Weladji, and J. Huot. 2009. Variation in calf body mass in migratory caribou: The role of habitat, climate, and movements. Journal of Mammalogy 90:442–52. doi:https://doi.org/10.1644/07-MAMM-A-279.1.
- Croll, D. A., J. L. Maron, J. A. Estes, E. M. Danner, and G. V. Byrd. 2005. Introduced predators transform subarctic islands from grassland to tundra. Science 307:1959–61. doi:https://doi.org/10.1126/science.1108485.
- Delaney, D. K., L. L. Pater, L. D. Carlile, E. W. Spadgenske, T. A. Beaty, and R. H. Melton. 2011. Response of red‐cockaded woodpeckers to military training operations. Wildlife Monographs 177:1–38. doi:https://doi.org/10.1002/wmon.3.
- Downes, C., J. Theberge, and S. Smith. 1986. The influence of insects on the distribution, microhabitat choice, and behaviour of the Burwash caribou herd. Canadian Journal of Zoology 64:622–29. doi:https://doi.org/10.1139/z86-092.
- Dray, S., M. Royer-Carenzi, and C. Calenge. 2010. The exploratory analysis of autocorrelation in animal-movement studies. Ecological Research 25:673–81. doi:https://doi.org/10.1007/s11284-010-0701-7.
- Dray, S., P. Legendre, and P. R. Peres-Neto. 2006. Spatial modelling: A comprehensive framework for principal coordinate analysis of neighbour matrices (PCNM). Ecological Modelling 196:483–93. doi:https://doi.org/10.1016/j.ecolmodel.2006.02.015.
- Ellis, D. H., C. H. Ellis, and D. P. Mindell. 1991. Raptor responses to low-level jet aircraft and sonic booms. Environmental Pollution 74:53–83. doi:https://doi.org/10.1016/0269-7491(91)90026-S.
- Erriksson, L.-O., M.-L. Källqvist, and T. Mossing. 1981. Seasonal development of circadian and short-term activity in captive reindeer, Rangifer tarandus L. Oecologia 48:64–70. doi:https://doi.org/10.1007/BF00346989.
- Fancy, S. G. 1983. Movements and activity budgets of caribou near oil drilling sites in the Sagavanirktok River floodplain, Alaska. Arctic 36:193–97. doi:https://doi.org/10.14430/arctic2262.
- Forbes, B. C. 2013. Cultural resilience of social-ecological systems in the Nenets and Yamal-Nenets Autonomous Okrugs, Russia: A focus on reindeer nomads of the tundra. Ecology and Society 18:36. doi:https://doi.org/10.5751/ES-05791-180436.
- Forbes, B. C., F. Stammler, T. Kumpula, N. Meschtyb, A. Pajunen, and E. Kaarlejärvi. 2009. High resilience in the Yamal-Nenets social–Ecological system, West Siberian Arctic, Russia. Proceedings of the National Academy of Sciences of the United States of America 106:22041–48. doi:https://doi.org/10.1073/pnas.0908286106.
- Forbes, B. C., J. J. Ebersole, and B. Strandberg. 2001. Anthropogenic disturbance and patch dynamics in circumpolar arctic ecosystems. Conservation Biology 15:954–69. doi:https://doi.org/10.1046/j.1523-1739.2001.015004954.x.
- Forbes, B. C., M. Bölter, L. Müller-Wille, J. Hukkinen, F. Müller, N. Gunslay, and Y. Konstantinov. 2006. Reindeer management in northernmost Europe. Berlin Heidelberg, Germany: Springer-Verlag.
- Forbes, B. C., and T. Kumpula. 2009. The ecological role and geography of reindeer (Rangifer tarandus) in northern Eurasia. Geography Compass 3:1356–80. doi:https://doi.org/10.1111/geco.2009.3.issue-4.
- Frid, A., and L. Dill. 2002. Human-caused disturbance stimuli as a form of predation risk. Conservation Ecology 6. doi: https://doi.org/10.5751/ES-00404-060111.
- Gaillard, J.-M., M. Festa-Bianchet, N. Yoccoz, A. Loison, and C. Toigo. 2000. Temporal variation in fitness components and population dynamics of large herbivores. Annual Review of Ecology and Systematics 31:367–93. doi:https://doi.org/10.1146/annurev.ecolsys.31.1.367.
- Gaston, K. J., J. Bennie, T. W. Davies, and J. Hopkins. 2013. The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biological Reviews 88:912–27. doi:https://doi.org/10.1111/brv.12036.
- Gill, J. A., K. Norris, and W. J. Sutherland. 2001. Why behavioural responses may not reflect the population consequences of human disturbance. Biological Conservation 97:265–68. doi:https://doi.org/10.1016/S0006-3207(00)00002-1.
- Gillies, C. S., M. Hebblewhite, S. E. Nielsen, M. A. Krawchuk, C. L. Aldridge, J. L. Frair, D. J. Saher, C. E. Stevens, and C. L. Jerde. 2006. Application of random effects to the study of resource selection by animals. Journal of Animal Ecology 75:887–98. doi:https://doi.org/10.1111/j.1365-2656.2006.01106.x.
- Goudie, R. I., and I. L. Jones. 2004. Dose-response relationships of harlequin duck behaviour to noise from low-level military jet over-flights in central Labrador. Environmental Conservation 31:289–98. doi:https://doi.org/10.1017/S0376892904001651.
- Gustine, D. D., K. L. Parker, R. J. Lay, M. P. Gillingham, and D. C. Heard. 2006. Calf survival of woodland caribou in a multi-predator ecosystem. Wildlife Monographs 165:1–32. doi:https://doi.org/10.2193/0084-0173(2006)165[1:CSOWCI]2.0.CO;2.
- Hagemoen, R. I. M., and E. Reimers. 2002. Reindeer summer activity pattern in relation to weather and insect harassment. Journal of Animal Ecology 71:883–92. doi:https://doi.org/10.1046/j.1365-2656.2002.00654.x.
- Hamel, S., and S. D. Côté. 2009. Maternal defensive behavior of mountain goats against predation by golden eagles. Western North American Naturalist 69:115–18. doi:https://doi.org/10.3398/064.069.0103.
- Harrington, F. H. 2003. Caribou, military jets and noise: The interplay of behavioural ecology and evolutionary psychology. Rangifer 23:73–80. doi:https://doi.org/10.7557/2.23.5.1683.
- Harrington, F. H., and A. M. Veitch. 1991. Short-term impacts of low-level jet fighter training on caribou in Labrador. Arctic 44:318–27. doi:https://doi.org/10.14430/arctic1554.
- Helle, T., and J. Aspi. 1984. Do sandy patches help reindeer against insects? Rep. Kevo Subarct. Res. Stn 19:57–62.
- Helle, T., V. Hallikainen, M. Särkelä, M. Haapalehto, A. Niva, and J. Puoskari. 2012. Effects of a holiday resort on the distribution of semi-domesticated reindeer. Annales Zoologici Fennici 49:23–35. doi:https://doi.org/10.5735/086.049.0103.
- Hobbs, N. T., H. Andrèn, J. Persson, M. Aronsson, and G. Chapron. 2012. Native predators reduce harvest of reindeer by Sami pastoralists. Ecological Applications 22:1640–54. doi:https://doi.org/10.1890/11-1309.1.
- Holden, E., and K. Linnerud. 2015. Sustainable mobility. In The Routledge handbook of tourism and sustainability, ed. C. Michael Hall, S. Gossling, and D. Scott, 409–19. Abingdon, UK: Routledge.
- Ims, R. A., and E. Fuglei. 2005. Trophic interaction cycles in tundra ecosystems and the impact of climate change. BioScience 55:311–22. doi:https://doi.org/10.1641/0006-3568(2005)055[0311:TICITE]2.0.CO;2.
- IPCC (Intergovernmental Panel on Climate Change). 2014. Mitigation of climate change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge, UK.
- Johnson, C. J., M. S. Boyce, R. L. Case, H. D. Cluff, R. J. Gau, A. Gunn, and R. Mulders. 2005. Cumulative effects of human developments on arctic wildlife. Wildlife Monographs 160:1–36.
- Kerley, L. L., and J. C. Slaght. 2013. First documented predation of Sika deer (Cervus nippon) by Golden Eagle (Aquila chrysaetos) in Russian far east. Journal of Raptor Research 47:328–30. doi:https://doi.org/10.3356/JRR-12-00008.1.
- Lantmäteriet. 2018. www.lantmateriet.se
- Legagneux, P., G. Gauthier, N. Lecomte, N. M. Schmidt, D. Reid, M.-C. Cadieux, D. Berteaux, J. Bêty, C. J. Krebs, R. A. Ims, et al. 2014. Arctic ecosystem structure and functioning shaped by climate and herbivore body size. Nature Climate Change 4:379. doi:https://doi.org/10.1038/nclimate2168.
- Lindgren, J. F., and M. Wilewska-Bien. 2016. Anthropogenic noise. In Shipping and the environment, ed. K. Andersson, S. Brynolf, J. Lindgren, and M. Wilewska-Bien, 229–235. Berlin Heidelberg, Germany: Springer-Verlag.
- Locati, M. 1990. Female chamois defends kids from eagle attacks. Mammalia 54:155–56.
- Longcore, T., and C. Rich. 2004. Ecological light pollution. Frontiers in Ecology and the Environment 2:191–98. doi:https://doi.org/10.1890/1540-9295(2004)002[0191:ELP]2.0.CO;2.
- Luick, B., J. Kitchens, R. White, and S. Murphy. 1996. Modeling energy and reproductive costs in caribou exposed to low flying military jet aircraft. Rangifer 16:209–12. doi:https://doi.org/10.7557/2.16.4.1244.
- Maier, J. A., S. M. Murphy, R. G. White, and M. D. Smith. 1998. Responses of caribou to overflights by low-altitude jet aircraft. The Journal of Wildlife Management 62:752–66. doi:https://doi.org/10.2307/3802352.
- Mallory, C., and M. Boyce. 2018. Observed and predicted effects of climate change on Arctic caribou and reindeer. Environmental Reviews 26:13–25.
- Michel, C. J., A. J. Ammann, E. D. Chapman, P. T. Sandstrom, H. E. Fish, M. J. Thomas, G. P. Singer, S. T. Lindley, A. P. Klimley, R. B. MacFarlane, et al. 2013. The effects of environmental factors on the migratory movement patterns of Sacramento River yearling late-fall run Chinook salmon (Oncorhynchus tshawytscha). Environmental Biology of Fishes 96:257–71. doi:https://doi.org/10.1007/s10641-012-9990-8.
- Moen, J. 2008. Climate change: Effects on the ecological basis for reindeer husbandry in Sweden. AMBIO: A Journal of the Human Environment 37:304–11. doi:https://doi.org/10.1579/0044-7447(2008)37[304:CCEOTE]2.0.CO;2.
- Mörschel, F. M., and D. R. Klein. 1997. Effects of weather and parasitic insects on behavior and group dynamics of caribou of the Delta Herd, Alaska. Canadian Journal of Zoology 75:1659–70. doi:https://doi.org/10.1139/z97-793.
- Murphy, S. M., and J. A. Curatolo. 1987. Activity budgets and movement rates of caribou encountering pipelines, roads, and traffic in northern Alaska. Canadian Journal of Zoology 65:2483–90. doi:https://doi.org/10.1139/z87-375.
- Musiani, M., J. A. Leonard, H. Cluff, C. C. Gates, S. Mariani, P. C. Paquet, C. Vilà, and R. K. Wayne. 2007. Differentiation of tundra/taiga and boreal coniferous forest wolves: Genetics, coat colour and association with migratory caribou. Molecular Ecology 16:4149–70. doi:https://doi.org/10.1111/j.1365-294X.2007.03458.x.
- NASA Earthdata Search. 2018. https://search.earthdata.nasa.gov/
- Nellemann, C., L. Kullerud, I. Vistnes, B. Forbes, E. Husby, G. Kofinas, B. Kaltenborn, J. Rouaud, M. Magomedova, et al. 2001. GLOBIO: Global methodology for mapping human impacts on the biosphere: The Arctic 2050 scenario and global application. Nairobi, Kenya: United Nations Environment Programme, UNEP-DEWA.
- Nette, T., D. Burles, and M. Hoefs. 1984. Observations of golden eagle, Aquila chrysaetos, predation on Dall sheep, Ovis dalli dalli, lambs. Canadian Field-Naturalist 98:252–54.
- Nieminen, M., H. Norberg, and V. Maijala. 2011. Mortality and survival of semi-domesticated reindeer (Rangifer tarandus tarandus L.) calves in northern Finland. Rangifer 31:71–84. doi:https://doi.org/10.7557/2.31.1.2029.
- Nilssen, A. C. 1997. Effect of temperature on pupal development and eclosion dates in the reindeer oestrids Hypoderma tarandi and Cephenemyia trompe (Diptera: Oestridae). Environmental Entomology 26:296–306. doi:https://doi.org/10.1093/ee/26.2.296.
- Nilssen, A. C., and J. R. Anderson. 1995. Flight capacity of the reindeer warble fly, Hypoderma tarandi (L.), and the reindeer nose bot fly, Cephenemyia trompe (Modeer)(Diptera: Oestridae). Canadian Journal of Zoology 73:1228–38. doi:https://doi.org/10.1139/z95-147.
- Nilssen, K. J., H. K. Johnsen, A. Rognmo, and A. S. Blix. 1984. Heart rate and energy expenditure in resting and running Svalbard and Norwegian reindeer. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 246:R963–R967. doi:https://doi.org/10.1152/ajpregu.1984.246.6.R963.
- Noel, L. E., R. H. Pollard, W. B. Ballard, and M. A. Cronin. 1998. Activity and use of active gravel pads and tundra by caribou, Rangifer tarandus granti, within the Prudhoe Bay oil field, Alaska. Canadian Field-Naturalist 112:400–09.
- Norberg, H., I. Kojola, P. Aikio, and M. Nylund. 2006. Predation by golden eagle Aquila chrysaetos on semi-domesticated reindeer Rangifer tarandus calves in northeastern Finnish Lapland. Wildlife Biology 12:393–402. doi:https://doi.org/10.2981/0909-6396(2006)12[393:PBGEAC]2.0.CO;2.
- Nybakk, K., O. Kjelvik, and T. Kvam. 1999. Golden eagle predation on semidomestic reindeer. Wildlife Society Bulletin 27:1038–42.
- Nybakk, K., O. Kjelvik, T. Kvam, K. Overskaug, and P. Sunde. 2002. Mortality of semi-domestic reindeer Rangifer tarandus in central Norway. Wildlife Biology 8:63–68. doi:https://doi.org/10.2981/wlb.2002.009.
- Panzacchi, M., B. van Moorter, O. Strand, M. Saerens, I. Kivimäki, C. C. St Clair, I. Herfindal, and L. Boitani. 2016. Predicting the continuum between corridors and barriers to animal movements using step selection functions and randomized shortest paths. Journal of Animal Ecology 85:32–42. doi:https://doi.org/10.1111/1365-2656.12386.
- Panzacchi, M., B. van Moorter, P. Jordhøy, and O. Strand. 2013. Learning from the past to predict the future: Using archaeological findings and GPS data to quantify reindeer sensitivity to anthropogenic disturbance in Norway. Landscape Ecology 28:847–59. doi:https://doi.org/10.1007/s10980-012-9793-5.
- Pedersen, V. A., J. D. Linnell, R. Andersen, H. Andrén, M. Lindén, and P. Segerström. 1999. Winter lynx Lynx lynx predation on semi-domestic reindeer Rangifer tarandus in northern Sweden. Wildlife Biology 5:203–11.
- Pollard, R., W. Ballard, L. Noel, and M. Cronin. 1996. Summer distribution of caribou, Rangifer tarandus granti, in the area of the Prudhoe Bay oil field, Alaska, 1990–1994. Canadian Field-Naturalist 110:659–74.
- Post, E., M. C. Forchhammer, M. S. Bret-Harte, T. V. Callaghan, T. R. Christensen, B. Elberling, A. D. Fox, O. Gilg, D. S. Hik, T. T. Hoye, et al. 2009. Ecological dynamics across the Arctic associated with recent climate change. Science 325:1355–58. doi:https://doi.org/10.1126/science.1173113.
- QGIS Development Team. 2018. QGIS geographic information system. Open Source Geospatial Foundation Project. http://qgis.osgeo.org
- Raponi, M., D. V. Beresford, J. A. Schaefer, I. D. Thompson, P. A. Wiebe, A. R. Rodgers, and J. M. Fryxell. 2018. Biting flies and activity of caribou in the boreal forest. The Journal of Wildlife Management 82:833–39. doi:https://doi.org/10.1002/jwmg.v82.4.
- Raynolds, M. K., D. A. Walker, K. J. Ambrosius, J. Brown, K. R. Everett, M. Kanevskiy, G. P. Kofinas, V. E. Romanovsky, Y. Shur, P. J. Webber, et al. 2014. Cumulative geoecological effects of 62 years of infrastructure and climate change in ice‐rich permafrost landscapes, Prudhoe Bay Oilfield, Alaska. Global Change Biology 20:1211–24. doi:https://doi.org/10.1111/gcb.2014.20.issue-4.
- Reimers, E., and J. E. Colman. 2006. Reindeer and caribou (Rangifer tarandus) response towards human activities. Rangifer 26:55–71. doi:https://doi.org/10.7557/2.26.2.188.
- Reimers, E., K. Røed, and J. Colman. 2012. Persistence of vigilance and flight response behaviour in wild reindeer with varying domestic ancestry. Journal of Evolutionary Biology 25:1543–54. doi:https://doi.org/10.1111/jeb.2012.25.issue-8.
- Reimers, E., L. E. Loe, S. Eftestøl, J. E. Colman, and B. Dahle. 2009. Effects of hunting on response behaviors of wild reindeer. The Journal of Wildlife Management 73:844–51. doi:https://doi.org/10.2193/2008-133.
- Reynolds, J. F., and J. D. Tenhunen. 2013. Landscape function and disturbance in arctic tundra. Berlin Heidelberg, Germany: Springer-Verlag.
- Roberts, B. A. 2014. The trials of motherhood: Maternal behavior patterns and antipredator tactics in Thomson’s gazelle (Gazella thomsonii), a hiding ungulate. Dissertation, Princeton, NJ: Princeton University.
- Sappington, J. M., K. M. Longshore, and D. B. Thompson. 2007. Quantifying landscape ruggedness for animal habitat analysis: A case study using bighorn sheep in the Mojave Desert. The Journal of Wildlife Management 71:1419–26. doi:https://doi.org/10.2193/2005-723.
- Schäfer, M., and J. O. Lundström. 2001. Comparison of mosquito (Diptera: Culicidae) fauna characteristics of forested wetlands in Sweden. Annals of the Entomological Society of America 94:576–82. doi:https://doi.org/10.1603/0013-8746(2001)094[0576:COMDCF]2.0.CO;2.
- Shannon, G., M. F. McKenna, L. M. Angeloni, K. R. Crooks, K. M. Fristrup, E. Brown, K. A. Warner, M. D. Nelson, C. White, J. Briggs, et al. 2016. A synthesis of two decades of research documenting the effects of noise on wildlife. Biological Reviews 91:982–1005. doi:https://doi.org/10.1111/brv.2016.91.issue-4.
- Sih, A., A. M. Bell, and J. L. Kerby. 2004. Two stressors are far deadlier than one. Trends in Ecology & Evolution 19:274–76. doi:https://doi.org/10.1016/j.tree.2004.02.010.
- Sivertsen, T. R. 2017. Risk of brown bear predation on semi-domesticated reindeer calves. Dissertation, Swedish University of Agricultural Sciences, Uppsala, Sweden.
- Skarin, A. 2007. Habitat use by semi-domesticated reindeer, estimated with pellet-group counts. Rangifer 27:121–32. doi:https://doi.org/10.7557/2.27.2.167.
- Skarin, A., Ö. Danell, R. Bergström, and J. Moen. 2004. Insect avoidance may override human disturbances in reindeer habitat selection. Rangifer 24:95–103. doi:https://doi.org/10.7557/2.24.2.306.
- Skarin, A., Ö. Danell, R. Bergström, and J. Moen. 2008. Summer habitat preferences of GPS-collared reindeer Rangifer tarandus tarandus. Wildlife Biology 14:1–15. doi:https://doi.org/10.2981/0909-6396(2008)14[1:SHPOGR]2.0.CO;2.
- Skarin, A., Ö. Danell, R. Bergström, and J. Moen. 2010. Reindeer movement patterns in alpine summer ranges. Polar Biology 33:1263–75. doi:https://doi.org/10.1007/s00300-010-0815-y.
- Skarin, A., and B. Åhman. 2014. Do human activity and infrastructure disturb domesticated reindeer? The need for the reindeer’s perspective. Polar Biology 37:1041–54. doi:https://doi.org/10.1007/s00300-014-1499-5.
- Skarin, A., C. Nellemann, L. Rönnegård, P. Sandström, and H. Lundqvist. 2015. Wind farm construction impacts reindeer migration and movement corridors. Landscape Ecology 30:1527–40. doi:https://doi.org/10.1007/s10980-015-0210-8.
- Skarin, A., P. Sandström, and M. Alam. 2018. Out of sight of wind turbines—Reindeer response to wind farms in operation. Ecology and Evolution 8:9906–19. doi:https://doi.org/10.1002/ece3.2018.8.issue-19.
- Steidl, R. J., and B.F.Powell. 2006. Assessing the effects of human activities on wildlife. The George Wright Forum 23(2):50–58. http://www.georgewright.org/232.pdf#page=52
- Stempniewicz, L., K. Błachowiak-Samołyk, and J. M. Węsławski. 2007. Impact of climate change on zooplankton communities, seabird populations and arctic terrestrial ecosystem—A scenario. Deep Sea Research Part II: Topical Studies in Oceanography 54:2934–45. doi:https://doi.org/10.1016/j.dsr2.2007.08.012.
- Swedish Hydrological and Meteorological Institute. 2017. http://opendata-downloadmetobs.smhi.se/explore/
- Swedish Sami Parliament. 2019. https://www.sametinget.se/8744
- Turi, J. 2002. The world reindeer livelihood - current situation, threats and possibilities. In Northern timberline forests: Environmental and socio-economic issues and concerns, ed. S. Kankaanpää, L. Müller-Wille, P. Susiluoto, and M.-L. Sutinen, 70–75. Helsinki, Finland: Finnish Forest Research Institute, Kolari Research Station.
- Uboni, A., D. W. Smith, J. S. Mao, D. R. Stahler, and J. A. Vucetich. 2015. Long‐and short‐term temporal variability in habitat selection of a top predator. Ecosphere 6:51. doi:https://doi.org/10.1890/ES14-00419.1.
- Uboni, A., J. A. Vucetich, D. R. Stahler, and D. W. Smith. 2015. Interannual variability: A crucial component of space use at the territory level. Ecology 96:62–70. doi:https://doi.org/10.1890/13-2116.1.
- Uboni, A., T. Horstkotte, E. Kaarlejärvi, A. Sévêque, F. Stammler, J. Olofsson, B. Forbes, and J. Moen. 2016. Long-term trends and role of climate in the population dynamics of Eurasian reindeer. PLoS One 11:e0158359. doi:https://doi.org/10.1371/journal.pone.0158359.
- van der Wal, R. 2006. Do herbivores cause habitat degradation or vegetation state transition? Evidence from the tundra. Oikos 114:177–86. doi:https://doi.org/10.1111/oik.2006.114.issue-1.
- van Oort, B. E., N. J. Tyler, M. P. Gerkema, L. Folkow, A. S. Blix, and K.-A. Stokkan. 2005. Circadian organization in reindeer. Nature 438:1095–96. doi:https://doi.org/10.1038/4381095a.
- Venables, W. N., and B. D. Ripley. 2002. Modern applied statistics with S. 4th ed. New York: Springer Science & Business Media.
- Vistnes, I., and C. Nellemann. 2001. Avoidance of cabins, roads, and power lines by reindeer during calving. The Journal of Wildlife Management 65:915–25. doi:https://doi.org/10.2307/3803040.
- Vistnes, I., and C. Nellemann. 2008. The matter of spatial and temporal scales: A review of reindeer and caribou response to human activity. Polar Biology 31:399–407. doi:https://doi.org/10.1007/s00300-007-0377-9.
- Vistnes, I. I., C. Nellemann, P. Jordhøy, and O.-G. Støen. 2008. Summer distribution of wild reindeer in relation to human activity and insect stress. Polar Biology 31:1307. doi:https://doi.org/10.1007/s00300-008-0468-2.
- Weladji, R. B., Ø. Holand, and T. Almøy. 2003. Use of climatic data to assess the effect of insect harassment on the autumn weight of reindeer (Rangifer tarandus) calves. Journal of Zoology 260:79–85. doi:https://doi.org/10.1017/S0952836903003510.
- White, R. G. 1992. Nutritional in relation to season, lactation, and growth of north temperate deer. In The Biology of Deer, ed. R. Brown, 407–17. New York: Springer-Verlag.
- Willard, B. E., D. J. Cooper, and B. C. Forbes. 2007. Natural regeneration of alpine tundra vegetation after human trampling: A 42-year data set from Rocky Mountain National Park, Colorado, USA. Arctic, Antarctic, and Alpine Research 39:177–83. doi:https://doi.org/10.1657/1523-0430(2007)39[177:NROATV]2.0.CO;2.
- Williams, C. M., H. A. Henry, and B. J. Sinclair. 2015. Cold truths: How winter drives responses of terrestrial organisms to climate change. Biological Reviews 90:214–35. doi:https://doi.org/10.1111/brv.12105.
- Wilson, S. F., and J. F. Wilmshurst. 2019. Behavioural responses of southern mountain caribou to helicopter and skiing activities. Rangifer 39:27–42. doi:https://doi.org/10.7557/2.39.1.4586.
- Witter, L. A., C. J. Johnson, B. Croft, A. Gunn, and L. M. Poirier. 2012. Gauging climate change effects at local scales: Weather‐based indices to monitor insect harassment in caribou. Ecological Applications 22:1838–51. doi:https://doi.org/10.1890/11-0569.1.
- Zuur, A. F., E. N. Ieno, and C. S. Elphick. 2010. A protocol for data exploration to avoid common statistical problems. Methods in Ecology and Evolution 1:3–14. doi:https://doi.org/10.1111/j.2041-210X.2009.00001.x.
- Zuur, A. F., E. N. Ieno, N. Walker, A. A. Saveliev, and G. M. Smith. 2009. Mixed effects models and extensions in ecology with R. New York: Springer.