
ABSTRACT
Lichen heaths are declining in abundance while shrubs are increasing their range in alpine and arctic areas due to climate change. This can have a large impact on the surface albedo of these areas. The aim of this article is to quantify the difference in albedo between lichen heaths and shrub-dominated vegetation and the variability within lichen heaths. Several environmental conditions that can influence the albedo measurements are considered. We measured the albedo of twenty lichen- and shrub-dominated plots on an alpine mountain area in southern Norway in the summer of 2018 with two radiometers using a paired plot design. With this design, we ensured similar weather conditions, aspects, and zenith angles between the paired lichen- and shrub-dominated plots. In addition, we collected patches of Cladonia stellaris and Flavocetraria nivalis to measure their albedo. The average difference in albedo between the lichen- and shrub-dominated plots is 0.124. The albedo of the lichen-dominated plots varies between 0.227 and 0.284, and that of the shrub-dominated plots varies between 0.115 and 0.148. This variation in albedo is explained by differences in aspect and vegetation composition. Further studies should focus on the consequences of this decrease in albedo for the microclimate in alpine and arctic areas.
Introduction
Warming causes changes in vegetation composition, especially in alpine and arctic areas (Wilson and Nilsson Citation2009; Vanneste et al. Citation2017; Maliniemi et al. Citation2018). Shrubs, for instance, are extending their range (Sturm, Racine, and Tape Citation2001; Hallinger, Manthey, and Wilmking Citation2010); species richness is locally increasing or decreasing (Michelsen et al. Citation2011); and certain plant species show elevational shifts (Walther et al. Citation2002; Klanderud and Birks Citation2003). One of the vegetation types in alpine and arctic areas most vulnerable to climate change is lichen heaths (Bjerke Citation2011). Recent studies have shown that lichen heaths are decreasing as a response to climate change through an increased competition with vascular plants (e.g., Cornelissen et al. Citation2001; Elmendorf et al. Citation2012) but also because of other factors, such as reindeer grazing and trampling (e.g., den Herder, Kytöviita, and Niemelä Citation2003; Joly, Jandt, and Klein Citation2009).
Lichens can also affect the climate of their direct environment. Several studies suggest that lichens act as a thermally insulating layer for the underlying soil (Beringer et al. Citation2001; Porada, Ekici, and Beer Citation2016). In addition, the relatively high albedo compared to other vegetation types can lead to differences in microclimate. Due to this high albedo, more shortwave radiation will be reflected, leading to a lower net radiation for lichens compared to other vegetation with a lower albedo. A lower net radiation means that less energy is available to heat the environment (Oke Citation2002). Not only will the microclimate be affected by the decrease in lichens but potentially climate on a larger scale because large parts of alpine and arctic areas are covered with lichens. For instance, alpine lichen heaths cover 6 percent of Norway (Bryn et al. Citation2018). Porada, Ekici, and Beer (Citation2016) modeled an average surface coverage of 69 percent for lichens and bryophytes in boreal forest and tundra at high latitudes. If lichen cover is reduced substantially, the albedo will decrease, and this might result in an increase of atmospheric heating for this region, as proposed by others (Chapin et al. Citation2005; Myers-Smith et al. Citation2011). For this reason, the decrease in lichens in alpine and arctic environments as an indirect effect of climate change can act as a positive feedback by further increasing temperatures. Hence, it is important to study how a decrease in lichen heaths affects the radiation budget and climate. To quantify the influence of lichens on the climate, a good estimation of their albedo is necessary.
The albedos of lichen heaths and single lichen species have been measured with both remote sensing and direct observations on the ground (Stoy et al. Citation2012; Cohen et al. Citation2013). Satellite images are often too coarse for albedo measurements of one single vegetation type or species in alpine areas, because vegetation is generally too fragmented in these areas (Virtanen and Ek Citation2014). Moreover, it is difficult to estimate the exact species composition of the area measured. Few studies have tried to quantify the albedo of lichens or pure lichen heaths in the field or in the laboratory (). These measurements indicate a large variation in albedo between lichen species as well.
Table 1. Albedo of lichen species measured in previous studies
The measured albedos of most of these lichen species are higher than the albedos of other alpine vegetation types. For instance, Juszak et al. (Citation2016) measured an albedo of 0.15 for a surface dominated by dwarf birch (Betula nana), one of the shrub species that has been shown to benefit most from global warming in alpine and arctic areas (Sturm, Racine, and Tape Citation2001; D. Walker et al. Citation2003). This lower albedo is caused not only by the higher reflection of lichens but also by the more complex canopy structure of shrubs. The taller shrub branches trap the radiation more easily in their canopy than lichens, leading to a lower albedo (Oke Citation2002; Beringer et al. Citation2005). Several studies have found evidence for marked expansion of shrubs in the Arctic at the expense of lichens (M. D. Walker et al. Citation2006; Fraser et al. Citation2014; Chagnon and Boudreau Citation2019). This shrub expansion results in a lower albedo and a higher absorption of solar radiation with potential impacts on the microclimate (Sturm et al. Citation2005) and climate on a regional scale (Loranty and Goetz Citation2012). The different studies measuring the albedo of lichen species and other vegetation types have provided important insight into the differences in albedo between lichens and other vegetation. However, these albedo values are difficult to compare because the impacts of several environmental factors have not yet been accounted for.
Several environmental factors can affect the measurement of albedo. Cloud cover and the solar zenith angle are two factors that can influence the measurements, even within the same day (Goodin and Isard Citation1989; Yang et al. Citation2008; Juszak et al. Citation2016). Clouds produce more diffuse solar radiation, which is more easily trapped by vegetation canopies than direct solar radiation. Therefore, the presence of clouds can potentially lead to a lower albedo. Other factors can influence albedo measurements as well, such as the aspect of a surface and the day of the year (Sicart et al. Citation2001; Yang et al. Citation2008). If these factors are not taken into account in comparison of albedo measurements of lichens with those of other vegetation, an adequate estimation of the difference in albedo between the surfaces cannot be made.
We aim to quantify the difference in albedo between lichen heaths and shrub-dominated vegetation in an alpine mountain area in southern Norway. During a field study, we simultaneously measured the albedo on lichen- and shrub-dominated plots with two radiometers. This paired study design ensured similar weather conditions, landscape position, zenith angles, and aspects for the lichen and the corresponding shrub plot. A second aim was to study the causes of the variation in albedo between the measured lichen heaths. In an additional controlled experiment, we measured the difference in albedo between two lichen species, Cladonia stellaris and Flavocetraria nivalis. Subsequently, we related the albedo of these two species to the findings in the field. With this study, we advance the understanding of the impact of the decrease in lichens in alpine and arctic areas on climate. Furthermore, the measured albedo values can be used for modeling purposes in order to model future climatic conditions in these areas.
Methods
Field study
Study area
The study area is part of the Imingfjell mountains (60.1901° N, 8.5724° E), located in Buskerud County, east of the Hardangervidda mountain plateau in southern Norway (). This area is located above the tree line and has an elevation of 1,150 ± 10 m. The area stretches approximately 2.5 km along a county road and 200 m from this road into the field. The landscape is characterized by an undulating topography with about 5 m height difference between the ridgetops and the low-lying parts. The area is dominated by low alpine vegetation with lichen heaths covering the numerous ridgetops (). The nearby but slightly lower elevated (800 m) climate station of Dagali reported an average yearly temperature of 0.5°C and an average yearly precipitation of 550 mm (MET Norway Citation2019). However, the period May to July of 2018, when the fieldwork was conducted, was relatively warm and dry. The average temperature of this period was 4.3°C above normal and only 60 percent of the normal precipitation was registered in this part of Norway (Skaland et al. Citation2019).
Figure 1. Location of Imingfjell in southern Norway and a picture of the study area showing that the lichen heaths are mainly found on the ridgetops and the shrubs on the ridgetops and midslopes

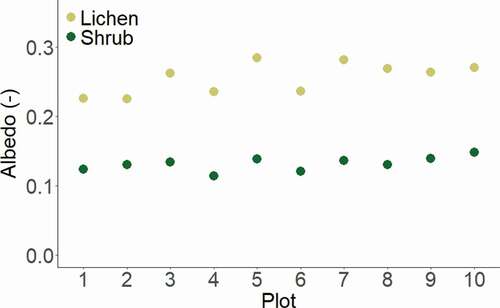
Figure 2. Two-day average albedo of the lichen and corresponding shrub plots. The mean (±SE) albedo for the lichen plots is 0.255 (±0.007) with a minimum of 0.227 and a maximum of 0.284. The mean (±SE) albedo for the shrub plots is 0.132 (±0.003) with a minimum of 0.115 and a maximum of 0.148
Sample design and measurements
As a first step, we delineated the lichen patches in ArcMap (Environmental Systems Research Institute Citation2019) using aerial images of the study area obtained from Geonorge (Citation2018). This resulted in a total area of 15.3 ha of lichen patches. We then randomly selected twenty locations within these lichen patches. While in the field, at each location we chose first a circular plot that was dominated by lichens and a circular plot that was dominated by shrubs, based on the criteria listed in . By using these criteria, we ensured that the lichen and shrub plot at one location had the same landscape position, aspect, zenith angle, and weather conditions. We placed a calibrated Kipp & Zonen CNR4 net radiometer (pyranometer accuracy: 5 percent) in each plot at a height of 30 cm in a horizontal position and measured the incoming and reflected shortwave radiation simultaneously at both plots (see Supplementary Information 1 for a picture of the setup). We calculated the albedo by dividing the reflected shortwave radiation by the incoming shortwave radiation. At 30 cm above the surface, the radiometer has a measurement radius of 112 cm, implying a plot area of 4 m2. The radiometers measured every five seconds and the two data loggers (Kipp & Zonen LOGBOX SE) recorded five-minute averages. On ten of the twenty paired plots, we measured the albedo for two days per paired plot, leading to twenty days of measurements between 4 July and 13 August 2018. To minimize the effect of precipitation on the albedo measurements, we ensured that none of these twenty days contained more than 30 minutes of precipitation . Due to time restrictions, we measured the albedo of the other ten paired plots for only 30 minutes per plot distributed over two days (26 and 27 July 2018). All ten plots were measured between 9:00 a.m. and 6:00 p.m. (zenith angle between 37° and 63°) to ensure that the zenith angle had a minimum effect on the albedo (Lafleur, Wurtele, and Duguay Citation1997). Plots that were measured for two days are hereafter referred to as two-day plots, and plots that were measured for 30 minutes are hereafter referred to as 30-minute plots.
Table 2. Criteria for the selection of lichen-dominated plots and corresponding shrub-dominated plots
In each plot, we visually estimated the percentage cover of all vascular plant and lichen species. The nomenclature followed Artsdatabanken (Citation2018). The slope angle and aspect of the plots were determined using a clinometer.
Data analysis
We calculated a two-day average albedo for every two-day plot. Only measurements between 9:00 a.m. and 6:00 p.m. were used because deviations in the measurements increase with larger zenith angles (Kipp & Zonen Citation2014; see Supplementary Information 2 for examples of time series of the measurements). We conducted a Wilcoxon signed-rank test between the average albedo of the ten two-day lichen and corresponding shrub plots to analyze the difference in albedo between the two plot types. The paired design controls for differences in zenith angle, aspect, and weather conditions between the lichen- and shrub-dominated plots.
For further analysis, we combined the albedo of the two-day plots and the 30-minute plots. To create a data set with a consistent measurement duration, a random 30-minute period was taken from every two-day plot. A Wilcoxon signed-rank test between the average albedo of the randomly selected 30-minute and two-day average albedo showed no significant difference (average difference: 0.0075 ± 0.0013; Supplementary Information 3). The 30-minute plots were measured on two days without clouds, whereas the two-day plots were measured during clear and cloudy days. This could potentially bias the analysis because cloud cover might have an influence on the albedo (Goodin and Isard Citation1989). In addition, the variation in zenith angles due to different measurement times of the 30-minute albedos within a day might cause additional noise (Goodin and Isard Citation1989). However, we did not find a systematic influence of cloud cover and zenith angle on the albedo (Supplementary Information 4 and 5); therefore, we concluded that it was reasonable to combine the average albedo of the random 30-minute measurements of the two-day plots and the albedo of the 30-minute plots into one data set.
We performed linear regressions between the albedo and percentage of the most abundant species per lichen-dominated plot to study whether certain species are associated with a higher albedo. The percentage of all species was recalculated to relative percentage so the abundance of all species in one plot summed up to 100 percent. We analyzed the vegetation composition of the lichen-dominated plots with detrended correspondence analysis (DCA) using the Vegan package (J. Oksanen et al. Citation2018) in R. With a DCA, we investigated any patterns of species and plots along underlying environmental gradients. These main patterns can provide information on co-occurrence of the studied species. The data were square root transformed and rare species were downweighted. We fitted the albedo in the DCA diagram using the envfit command of the Vegan package to study whether there was a relation between the species composition of the plots and the albedo. We applied beta regression using the betareg package (Cribari-Neto and Zeileis Citation2009) to study the influence of slope and aspect on the albedo of the lichen-dominated plots. Beta regression was chosen because the response variable (albedo) varies between 0 and 1. The total percentage of all Cladonia spp. was added as an explanatory variable as a proxy for the vegetation composition. All possible models including interactions were built and the one with the lowest Akaike information criterion was selected. One plot was removed from the analysis because it had a high leverage in the model (Cook’s distance = 1.8). All analyses were done using R v3.4.3 (R Core Team Citation2017).
Controlled experiment
To investigate the difference in albedo between lichen species, we collected mats of F. nivalis and C. stellaris on Imingfjell to perform controlled experiments. These two species were chosen because they are among the most abundant lichen species in alpine areas, and plots dominated by either of these two species had different albedos during the field study (see Results). We placed both species on two circular boards and measured the albedo with the same two radiometers used in the field study on six clear days in April 2019 near the city of Bø in southern Norway (59.409156° N, 9.056464° E; 81 m a.s.l.; see Supplementary Information 6 for a picture of the setup). Because we expect, based on the field study, a small difference in albedo between the two species, the radiometers were changed regularly between both lichen plots to exclude potential bias caused by differences between the radiometers (Eugster, Mcfadden, and Chapin Citation1997). We measured the albedo of both species simultaneously for 30 minutes in the morning and 30 minutes in the afternoon. The solar zenith angle was kept constant within a day and between days during all measurements (57°–60°).
To study the impact of aspect on the albedo of the two species, we measured the albedo of the lichen species for 30 minutes in a north-facing position and 30 minutes in a south-facing position in the morning. During the same solar zenith angles in the afternoon (46°–57°), we measured the albedo again using the same procedure. We used a constant slope angle of 8°.
Due to constraints of time and resources, it was not possible to gather more specimens in the field and therefore only one plot of C. stellaris and one plot of F. nivalis was available. Hence, we calculated only the average albedo of the six measurement days for these two lichen species and the average albedo of the different aspects. We conducted no further statistical tests.
Results
Field study
The studied lichen-dominated plots consist of at least 85 percent lichens and the studied shrub-dominated plots consist of at least 90 percent shrubs. The main genera observed in the lichen-dominated plots are Cladonia, Flavocetraria, Cetraria, and Alectoria. The main shrub species in the plots are B. nana and Empetrum nigrum (see Supplementary Information 7 and 8 for the vegetation composition of the plots). The albedo of the lichen-dominated plots was, on average (±SE), 0.124 ± 0.005 higher than the albedo of the shrub-dominated plots (p = .002). The mean (±SE) albedo for the ten two-day lichen-dominated plots was 0.255 ± 0.007 and that for the ten two-day shrub-dominated plots was 0.132 ± 0.003 ().
Lichen-dominated plots with a relatively high abundance of F. nivalis and Alectoria ochroleuca have a higher albedo compared to plots with a relatively high abundance of lichen species of Cladonia spp., Cetraria ericetorum, and the dwarf birch B. nana (). Flavocetraria cucullata (R2 = 0.052, p = .3; ) and E. nigrum (R2 = 0.009, p = .7; ) have no significant relationship with albedo. The ordination separates the Cladonia spp. and F. nivalis and A. ochroleuca, which implies that they are not likely to occur abundantly together (). The albedo increases toward F. nivalis and A. ochroleuca (r = 0.520, p = .004), indicating that plots dominated by these species have a higher albedo than plots dominated by Cladonia spp. This is in agreement with the relationships found in . The DCA diagram also indicates that Cetraria ericetorum and B. nana are more likely to occur in plots dominated by Cladonia spp. than in plots dominated by F. nivalis and A. ochroleuca.
Figure 3. Linear regression between albedo and the percentage of most abundant species within all lichen plots. Plots (a)–(f) are light lichens, plot (g) is a dark lichen, and plots (h) and (i) are shrub species
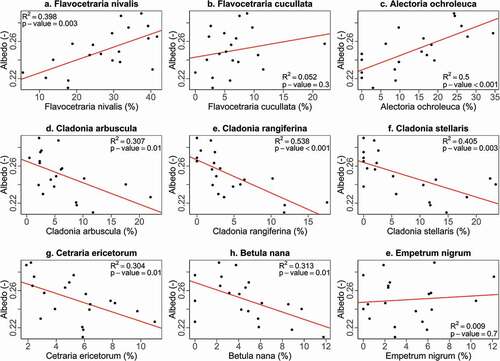
The best model to explain the variation in albedo between the lichen plots includes all of the explanatory variables but no interactions. South- and west-facing slopes have a significantly higher albedo compared to north-facing slopes, and relatively steep slopes have a lower albedo (). No east-facing slopes were present in this study. The model implies that a larger percentage of total Cladonia spp. is associated with a lower albedo, which is in line with the linear regressions () and the DCA ().
Table 3. Result of the best beta regression model using albedo as dependent variable and aspect, slope, and total percentage of Cladonia spp. as explanatory variables
Figure 4. DCA ordination of all lichen plots. Species labels consist of the first three letters of the genus and the first three letters of the species names (see Supplementary Information 9 for clarification). Arrow indicates the direction of increasing albedo. The ellipses cluster the group of species associated with a low albedo and the group of species associated with a high albedo according to the linear regressions in Figure 3
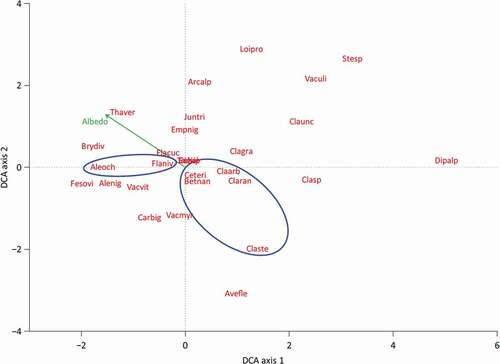
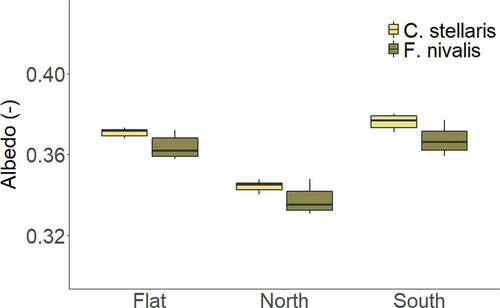
Figure 5. Measured albedo during the controlled experiment of Cladonia stellaris and Flavocetraria nivalis grouped by different aspects (n = 6 days for each box plot)
Controlled experiment
The mean (±SE) albedo of the horizontal surface of C. stellaris is 0.371 ± 0.001 (). This is slightly higher than the mean albedo of the horizontal F. nivalis (0.364 ± 0.002). However, the difference in albedo between the two species falls within the accuracy of the radiometers and is dependent on the distribution of the radiometers (Supplementary Information 10). Hence, it cannot be concluded that there is a substantial difference in albedo between the two lichen species. The north-facing slope has a substantially lower albedo (0.341 ± 0.003) than the horizontal surface (0.367 ± 0.002) and the south-facing slope (0.372 ± 0.003).
Discussion
Difference in albedo between lichens and shrubs
The higher albedo of the lichen-dominated plots is due to the higher reflection of shortwave radiation of the lichens and is in line with previous studies that have measured albedo of lichen heaths (Petzold and Rencz Citation1975; Williamson et al. Citation2016) and shrub vegetation (Juszak et al. Citation2016). The measured albedo of the lichen-dominated plots during the field study was higher than that reported in other studies that measured the albedo of alpine lichen heaths, because our lichen-dominated plots consisted of at least 85 percent lichens. Williamson et al. (Citation2016) measured an albedo of 0.19 on a Dryas–lichen tundra. However, their plot consisted of only 34 percent lichens. Petzold and Rencz (Citation1975) measured an albedo of 0.21 on a lichen heath, which is only slightly lower than the albedo measured in our study. The albedo of plots dominated by B. nana was previously measured by Juszak et al. (Citation2016), who found a slightly higher albedo of 0.15 during the growing season in northeast Siberia, compared to 0.13 in our study. A possible reason for this slight difference is that their measurements were performed during larger zenith angles than our measurements.
Difference in albedo between Flavocetraria nivalis and Cladonia stellaris (controlled experiment)
We measured a difference in albedo of less than 0.01 between the two lichen species, which falls within the accuracy of the radiometers. Therefore, we cannot establish a clear difference in albedo between C. stellaris and F. nivalis. The albedos of C. stellaris and F. nivalis measured during the controlled experiment do partly coincide with other studies that measured the albedo of lichen monocultures (). Petzold and Rencz (Citation1975) measured a lower albedo for C. stellaris (0.22) than this study (0.37). An evident difference is that the study of Petzold and Rencz (Citation1975) measured under natural conditions with stunted black spruce occasionally present in their plots, whereas we measured under partly controlled conditions without any other vegetation mixed in the plot. Therefore, the black spruce could be the reason for the difference in albedo between their plots and our plots.
A limitation of the controlled experiment is that the albedo of only one C. stellaris surface and one F. nivalis surface could be measured. However, Heim and Lundholm (Citation2013) measured the albedo of five different surfaces of a mixture of Cladonia terranova and Cladonia boryi and the difference between the highest and lowest albedo was only 0.015. Therefore, we argue that our controlled experiment provides an accurate approximation for the albedo differences between C. stellaris and F. nivalis.
Variation in albedo within lichen heaths
The results of the field study show that the variation in albedo between lichen heaths is caused by the vegetation composition. The linear regressions and the DCA diagram show a distinction in albedo between lichen heaths dominated by Cladonia spp. and lichen heaths dominated by F. nivalis and A. ochroleuca. This distinction is most likely not a result of a difference in albedo between these lichen species, because we measured only a minor difference (<0.01) during the controlled experiment between C. stellaris and F. nivalis. Instead, the difference is caused by a higher abundance of the dwarf shrub B. nana and the dark-colored lichen species C. ericetorum in the plots dominated by Cladonia spp. Cladonia spp., B. nana, and C. ericetorum are species that grow on places with some snow cover during the winter months, whereas F. nivalis and A. ochroleuca grow on more exposed ridges with almost no snow cover (L. Oksanen and Virtanen Citation1995; Odland and Munkejord Citation2008). Therefore, B. nana and C. ericetorum more likely occur together with Cladonia spp. than with F. nivalis and A. ochroleuca, as was also the case in the plots that we studied (). The higher abundances of B. nana and the dark-colored lichen species C. ericetorum therefore generate a lower albedo of plots dominated by Cladonia spp. than that plots that are dominated by F. nivalis and A. ochroleuca.
In addition to vegetation composition, the topographical conditions caused variation in the albedo between lichen surfaces. Both the field study and the controlled experiment indicate that a surface on a north-facing slope has a lower albedo than a surface on a south-facing slope, despite a slope angle of less than 10° for all plots. However, the difference in albedo is a result of the measurement procedure rather than a difference in albedo itself. The lower pyranometer of the radiometer measures the reflected shortwave radiation of the sloping surface, and the upper pyranometer measures the incoming solar radiation of a horizontal surface because the sensors were placed in a level position (Supplementary information 11). The incoming solar radiation for a north-facing slope is therefore overestimated, leading to a lower albedo, and the incoming shortwave radiation for a south-facing slope is underestimated, leading to a higher albedo. The difference in incoming shortwave radiation between aspects results in south-facing slopes being warmer than north-facing slopes (Barry Citation2008; Winkler et al. Citation2016). The variation in albedo due to small topographical differences has been considered before and is classified as an important element that affects the measured and modeled albedo in rugged terrain (Hao et al. Citation2018). Our study shows that alpine areas covered with lichen heaths are not an exception. The slopes used in this study did not exceed 10° and it is therefore likely that the difference in albedo measurements between north- and south-facing slopes would be even larger when steeper slopes are measured using the same method. However, lower zenith angles might minimize the effect of aspect on the albedo measurements, leading to a smaller difference in albedo between north- and south-facing slopes. More thorough measurements are needed to quantify the combined effect of aspect and the solar zenith angle on the albedo of lichen surfaces.
Future perspectives
Global warming and changes in precipitation patterns will continue to affect the vegetation composition of alpine and arctic areas (Wehn, Lundemo, and Holten Citation2014). The expansion of shrubs and decrease in lichen cover has the potential to continue under ongoing climate change, as shown by experimental warming studies (M. D. Walker et al. Citation2006; Lang et al. Citation2012). Lichen heaths dominated by Flavocetraria spp. and A. ochroleuca are most vulnerable to the presence of shrubs (Chagnon and Boudreau Citation2019) and changing snow conditions (Wahren, Walker, and Bret‐Harte Citation2005; Odland and Munkejord Citation2008; Bidussi, Solhaug, and Gauslaa Citation2016). Therefore, we expect that lichen heaths dominated by these species will shift to lichen heaths dominated by Cladonia spp., because Cladonia spp. are more tolerant to changing snow conditions and are more likely to occur with B. nana (L. Oksanen and Virtanen Citation1995; Odland and Munkejord Citation2008). Our study shows that such a shift will lower the albedo of these areas substantially. The reason for this is the presence of B. nana and C. ericetorum in the lichen heaths dominated by Cladonia spp. However, we expect that the increase in shrubs will continue and that lichen heaths will turn into shrub-dominated vegetation within decades, as shown by several other studies (Sturm, Racine, and Tape Citation2001; M. D. Walker et al. Citation2006; Fraser et al. Citation2014). Fraser et al. (Citation2014) observed that large continuous lichen mats have disappeared almost completely in the Canadian Arctic since 1980. A global assessment of experimental warming studies showed that shrubs increased with warming, whereas lichens decreased (Elmendorf et al. Citation2012). This shift toward a shrub-dominated landscape will further decrease the albedo and has important consequences for both micro- and macroclimates, because the decrease in albedo will feed back to the climate (Pearson et al. Citation2013).
Multiple papers have mentioned the potential effect that a decrease in albedo due to the loss of lichen abundance will have on other aspects of climate, such as the surface energy balance, soil temperature, and moisture conditions (Stoy et al. Citation2012; Porada, Ekici, and Beer Citation2016; Odland, Sundstøl, and Bjerketvedt Citation2018). Porada, Ekici, and Beer (Citation2016) estimated with a model study that lichens and bryophytes decrease the average soil temperature by 2.7°C at latitudes higher than 50° N. However, their model did not consider the high albedo of lichens but solely the insulating effect of bryophytes and lichens. Therefore, the cooling effect of lichens on the soil might be even larger when the high albedo measured in our study is considered in combination with the insulating characteristics of lichens. Several studies tried to quantify the effect of arctic greening on large-scale albedo and therefore measured the albedo on generalized landscape units; for example, arctic tundra and shrub land (Beringer et al. Citation2005). As a result, a smaller difference in albedo between tundra and shrub vegetation is usually used to predict the impact of albedo change on the future climate. Chapin et al. (Citation2005) reported a potential increase in atmospheric heating of 49.50 MJ m−2 year−1 due to shrub expansion, though they assumed a difference in albedo of only 0.02 between tundra and shrubs. A decrease in albedo of 0.124, as measured during our study, will therefore increase the net radiation substantially. Our vegetation type–specific study shows that a large variation in albedo exists within the landscape units. This variation might subsequently lead to large differences in microclimate within alpine and arctic areas. Therefore, a species-specific approach is required to quantify the exact contribution of several vegetation types to the microclimate and climate of alpine and arctic regions in the future, as proposed by others (Stoy et al. Citation2012; Juszak et al. Citation2014; Williamson et al. Citation2016).
Conclusion
The aim of this study was to quantify the difference in albedo between lichen heaths and shrub-dominated vegetation using a paired study design to ensure similar weather conditions, solar zenith angle, and aspects between the paired plots. We measured a difference in albedo of 0.12 between the lichen- and shrub-dominated plots. A second aim of this study was to measure the variation in albedo between lichen heaths. We found differences in albedo of up to 0.06 between lichen heaths, due to the combined effect of differences in vegetation composition (e.g., abundance of B. nana and C. ericetorum in lichen heaths) and topographical differences. The decrease in albedo due to a decrease in lichen heaths may have a large influence on the climate directly near the vegetation but also on a larger scale. However, this has not been thoroughly studied yet, and further research should therefore focus on quantification of the consequences of this decrease in albedo for the local climate and subsequently for large-scale climate.
Supplemental Material
Download Zip (1.9 MB)Acknowledgments
We thank Kristine Hetlesæter and Konstanse Skøyen for their help with the fieldwork and three anonymous reviewers for valuable comments on an earlier version of this article.
Disclosure statement
The authors report no conflicts of interest.
Supplemental data
Supplemental data for this article can be accessed here.
Additional information
Funding
References
- Artsdatabanken. 2018. http://www2.artsdatabanken.no/artsnavn/Contentpages/Hjem.aspx.
- Barry, R. G. 2008. Mountain weather and climate. 3rd ed. Cambridge: Cambridge University Press.
- Beringer, J., F. S. Chapin III, C. C. Thompson, and A. D. Mcguire. 2005. Surface energy exchanges along a tundra-forest transition and feedbacks to climate. Agricultural and Forest Meteorology 131 (3–4):143–61. doi:https://doi.org/10.1016/j.agrformet.2005.05.006.
- Beringer, J., A. H. Lynch, F. S. Chapin, M. Mack, and G. B. Bonan. 2001. The representation of arctic soils in the land surface model: The importance of mosses. Journal of Climate 14 (15):3324–35. doi:https://doi.org/10.1175/1520-0442(2001)014%3c3324:TROASI%3e2.0.CO;2.
- Bidussi, M., K. A. Solhaug, and Y. Gauslaa. 2016. Increased snow accumulation reduces survival and growth in dominant mat-forming arctic-alpine lichens. The Lichenologist 48 (3):237–47. doi:https://doi.org/10.1017/S0024282916000086.
- Bjerke, J. W. 2011. Winter climate change: Ice encapsulation at mild subfreezing temperatures kills freeze-tolerant lichens. Environmental and Experimental Botany 72 (3):404–08. doi:https://doi.org/10.1016/j.envexpbot.2010.05.014.
- Bryn, A., G.-H. Strand, M. Angeloff, and Y. Rekdal. 2018. Land cover in Norway based on an area frame survey of vegetation types. Norsk Geografisk Tidsskrift-Norwegian Journal of Geography 72 (3):131–45. doi:https://doi.org/10.1080/00291951.2018.1468356.
- Chagnon, C., and S. Boudreau. 2019. Shrub canopy induces a decline in lichen abundance and diversity in Nunavik (Québec, Canada). Arctic, Antarctic, and Alpine Research 51 (1):521–32. doi:https://doi.org/10.1080/15230430.2019.1688751.
- Chapin, F. S., M. Sturm, M. C. Serreze, J. P. Mcfadden, J. Key, A. H. Lloyd, A. Mcguire, T. S. Rupp, A. H. Lynch, and J. P. Schimel. 2005. Role of land-surface changes in Arctic summer warming. Science 310 (5748):657–60. doi:https://doi.org/10.1126/science.1117368.
- Cohen, J., J. Pulliainen, C. B. Ménard, B. Johansen, L. Oksanen, K. Luojus, and J. Ikonen. 2013. Effect of reindeer grazing on snowmelt, albedo and energy balance based on satellite data analyses. Remote Sensing of Environment 135:107–17. doi:https://doi.org/10.1016/j.rse.2013.03.029.
- Cornelissen, J. H. C., T. V. Callaghan, J. Alatalo, A. Michelsen, E. Graglia, A. Hartley, D. Hik, S. Hobbie, M. Press, and C. H. Robinson. 2001. Global change and arctic ecosystems: Is lichen decline a function of increases in vascular plant biomass? Journal of Ecology 89 (6):984–94. doi:https://doi.org/10.1111/j.1365-2745.2001.00625.x.
- Cribari-Neto, F., and A. Zeileis. 2009. Beta regression in R. Journal of Statistical Software 34 ((2)). doi: https://doi.org/10.18637/jss.v034.i02.
- den Herder, M., M. M. Kytöviita, and P. Niemelä. 2003. Growth of reindeer lichens and effects of reindeer grazing on ground cover vegetation in a Scots pine forest and a subarctic heathland in Finnish Lapland. Ecography 26 (1):3–12. doi:https://doi.org/10.1034/j.1600-0587.2003.03211.x.
- Elmendorf, S. C., G. H. Henry, R. D. Hollister, R. G. Björk, A. D. Bjorkman, T. V. Callaghan, L. S. Collier, E. J. Cooper, J. H. C. Cornelissen, T. A. Day, et al. 2012. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecology Letters 15 (2):164–75. doi:https://doi.org/10.1111/j.1461-0248.2011.01716.x.
- Environmental Systems Research Institute (ESRI). 2019. ArcGIS release 10.6. Redlands, CA.
- Eugster, W., J. P. Mcfadden, and F. S. Chapin. 1997. A comparative approach to regional variation in surface fluxes using mobile eddy correlation towers. Boundary-Layer Meteorology 85 (2):293–307. doi:https://doi.org/10.1023/A:1000552311805.
- Fraser, R. H., T. C. Lantz, I. Olthof, S. V. Kokelj, and R. A. Sims. 2014. Warming-induced shrub expansion and lichen decline in the Western Canadian Arctic. Ecosystems 17 (7):1151–68. doi:https://doi.org/10.1007/s10021-014-9783-3.
- Geonorge. 2018. Norge i bilder WMS-Ortofoto.
- Goodin, D., and S. Isard. 1989. Magnitude and sources of variation in albedo within an alpine tundra. Theoretical and Applied Climatology 40 (1–2):61–66. doi:https://doi.org/10.1007/BF00867792.
- Hallinger, M., M. Manthey, and M. Wilmking. 2010. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytologist 186 (4):890–99. doi:https://doi.org/10.1111/j.1469-8137.2010.03223.x.
- Hao, D., J. Wen, Q. Xiao, S. Wu, X. Lin, B. Dou, D. You, and Y. Tang. 2018. Simulation and analysis of the topographic effects on snow-free albedo over rugged terrain. Remote Sensing 10 (2):278. doi:https://doi.org/10.3390/rs10020278.
- Heim, A., and J. Lundholm. 2013. Cladonia lichens on extensive green roofs: Evapotranspiration, substrate temperature, and albedo. F1000Research (2):274. doi:https://doi.org/10.12688/f1000research.2-274.v2.
- Joly, K., R. R. Jandt, and D. R. Klein. 2009. Decrease of lichens in Arctic ecosystems: The role of wildfire, caribou, reindeer, competition and climate in north‐western Alaska. Polar Research 28 (3):433–42. doi:https://doi.org/10.1111/j.1751-8369.2009.00113.x.
- Juszak, I., A. M. Erb, T. C. Maximov, and G. Schaepman-Strub. 2014. Arctic shrub effects on NDVI, summer albedo and soil shading. Remote Sensing of Environment 153:79–89. doi:https://doi.org/10.1016/j.rse.2014.07.021.
- Juszak, I., W. Eugster, M. M. P. D. Heijmans, and G. Schaepman-Strub. 2016. Contrasting radiation and soil heat fluxes in Arctic shrub and wet sedge tundra. Biogeosciences 13 (13):4049. doi:https://doi.org/10.5194/bg-13-4049-2016.
- Kipp & Zonen. 2014. Instruction manual: CNR-4 net radiometer.
- Klanderud, K., and H. J. B. Birks. 2003. Recent increases in species richness and shifts in altitudinal distributions of Norwegian mountain plants. The Holocene 13 (1):1–6. doi:https://doi.org/10.1191/0959683603hl589ft.
- Lafleur, P. M., A. B. Wurtele, and C. R. Duguay. 1997. Spatial and temporal variations in surface albedo of a subarctic landscape using surface-based measurements and remote sensing. Arctic and Alpine Research 29 (3):261–69. doi:https://doi.org/10.1080/00040851.1997.12003244.
- Lang, S. I., J. H. Cornelissen, G. R. Shaver, M. Ahrens, T. V. Callaghan, U. Molau, C. J. Ter Braak, A. Hölzer, and R. Aerts. 2012. Arctic warming on two continents has consistent negative effects on lichen diversity and mixed effects on bryophyte diversity. Global Change Biology 18 (3):1096–107. doi:https://doi.org/10.1111/j.1365-2486.2011.02570.x.
- Loranty, M. M., and S. J. Goetz. 2012. Shrub expansion and climate feedbacks in Arctic tundra. Environmental Research Letters 7 (1):011005. doi:https://doi.org/10.1088/1748-9326/7/1/011005.
- Maliniemi, T., J. Kapfer, P. Saccone, A. Skog, and R. Virtanen. 2018. Long‐term vegetation changes of treeless heath communities in northern Fennoscandia: Links to climate change trends and reindeer grazing. Journal of Vegetation Science 29 (3):469–79. doi:https://doi.org/10.1111/jvs.12630.
- MET Norway. 2019. Meteorological data. www.met.no.
- Michelsen, O., A. O. Syverhuset, B. Pedersen, and J. I. Holten. 2011. The impact of climate change on recent vegetation changes on Dovrefjell, Norway. Diversity 3 (1):91–111. doi:https://doi.org/10.3390/d3010091.
- Myers-Smith, I. H., B. C. Forbes, M. Wilmking, M. Hallinger, T. Lantz, D. Blok, K. D. Tape, M. Macias-Fauria, U. Sass-Klaassen, and E. Lévesque. 2011. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environmental Research Letters 6 (4):045509. doi:https://doi.org/10.1088/1748-9326/6/4/045509.
- Odland, A., and H. K. Munkejord. 2008. Plants as indicators of snow layer duration in southern Norwegian mountains. Ecological Indicators 8 (1):57–68. doi:https://doi.org/10.1016/j.ecolind.2006.12.005.
- Odland, A., S. Sundstøl, and D. Bjerketvedt. 2018. Alpine lichen-dominated heaths: Ecology, effects of reindeer grazing, and climate change. A review. Oecologia Montana 27 (2):30–50.
- Oke, T. R. 2002. Boundary layer climates. London: Routledge.
- Oksanen, J., F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. R. Minchin, R. B. O'Hara, G. L. Simpson, P. Solymos, et al. 2018. Vegan: Community ecology package. R Package Version 2:4–6.
- Oksanen, L., and R. Virtanen. 1995. Topographic, altitudinal and regional patterns in continental and suboceanic heath vegetation of northern Fennoscandia. Acta Botanica Fennica 153:1–80.
- Pearson, R. G., S. J. Phillips, M. M. Loranty, P. S. Beck, T. Damoulas, S. J. Knight, and S. J. Goetz. 2013. Shifts in Arctic vegetation and associated feedbacks under climate change. Nature Climate Change 3 (7):673–77. doi:https://doi.org/10.1029/2005JG000013.
- Peltoniemi, J. I., T. Manninen, J. Suomalainen, T. Hakala, E. Puttonen, and A. Riihelä. 2010. Land surface albedos computed from BRF measurements with a study of conversion formulae. Remote Sensing 2 (8):1918–40. doi:https://doi.org/10.3390/rs2081918.
- Petzold, D., and A. Rencz. 1975. The albedo of selected subarctic surfaces. Arctic and Alpine Research 7:393–98. doi:https://doi.org/10.2307/1550183.
- Porada, P., A. Ekici, and C. Beer. 2016. Effects of bryophyte and lichen cover on permafrost soil temperature at large scale. The Cryosphere 10 (5):2291. doi:https://doi.org/10.5194/tc-10-2291-2016.
- R Core Team. 2017. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
- Sicart, J. E., P. Ribstein, P. Wagnon, and D. Brunstein. 2001. Clear‐sky albedo measurements on a sloping glacier surface: A case study in the Bolivian Andes. Journal of Geophysical Research: Atmospheres 106 (D23):31729–37. doi:https://doi.org/10.1029/2000JD000153.
- Skaland, R. G., H. Colleuille, A. S. H. Andersen, J. Mamen, L. Grinde, H. T. T. Tajet, E. Lundstad, L. F. Sidselrud, K. Tunheim, I. Hanssen-Bauer, et al. 2019. Tørkesommeren 2018. Oslo: Meteorologisk institutt.
- Stoy, P. C., L. E. Street, A. V. Johnson, A. Prieto-Blanco, and S. A. Ewing. 2012. Temperature, heat flux, and reflectance of common subarctic mosses and lichens under field conditions: Might changes to community composition impact climate-relevant surface fluxes? Arctic, Antarctic, and Alpine Research 44 (4):500–08. doi:https://doi.org/10.1657/1938-4246-44.4.500.
- Sturm, M., T. Douglas, C. Racine, and G. E. Liston. 2005. Changing snow and shrub conditions affect albedo with global implications. Journal of Geophysical Research: Biogeosciences 110:G1. doi:https://doi.org/10.1038/nclimate1858.
- Sturm, M., C. Racine, and K. Tape. 2001. Increasing shrub abundance in the Arctic. Nature 411:546. doi:https://doi.org/10.1038/35079180.
- Vanneste, T., O. Michelsen, B. J. Graae, M. O. Kyrkjeeide, H. Holien, K. Hassel, S. Lindmo, R. E. Kapás, and P. De Frenne. 2017. Impact of climate change on alpine vegetation of mountain summits in Norway. Ecological Research 1–15. doi:https://doi.org/10.1007/s11284-017-1472-1.
- Virtanen, T., and M. Ek. 2014. The fragmented nature of tundra landscape. International Journal of Applied Earth Observation and Geoinformation 27:4–12. doi:https://doi.org/10.1016/j.jag.2013.05.010.
- Wahren, C. H., M. Walker, and M. Bret‐Harte. 2005. Vegetation responses in Alaskan arctic tundra after 8 years of a summer warming and winter snow manipulation experiment. Global Change Biology 11 (4):537–52. doi:https://doi.org/10.1111/j.1365-2486.2005.00927.x.
- Walker, D., H. Epstein, G. Jia, A. Balser, C. Copass, E. Edwards, W. Gould, J. Hollingsworth, J. Knudson, and H. Maier. 2003. Phytomass, LAI, and NDVI in northern Alaska: Relationships to summer warmth, soil pH, plant functional types, and extrapolation to the circumpolar Arctic. Journal of Geophysical Research: Atmospheres 108:D2. doi:https://doi.org/10.1029/2001JD000986.
- Walker, M. D., C. H. Wahren, R. D. Hollister, G. H. Henry, L. E. Ahlquist, J. M. Alatalo, M. S. Bret-Harte, M. P. Calef, T. V. Callaghan, A. B. Carroll, et al. 2006. Plant community responses to experimental warming across the tundra biome. Proceedings of the National Academy of Sciences 103 (5):1342–46. doi:https://doi.org/10.1073/pnas.0503198103.
- Walther, G.-R., E. Post, P. Convey, A. Menzel, C. Parmesan, T. J. Beebee, J.-M. Fromentin, O. Hoegh-Guldberg, and F. Bairlein. 2002. Ecological responses to recent climate change. Nature 416 (6879):389–95. doi:https://doi.org/10.1038/416389a.
- Wehn, S., S. Lundemo, and J. I. Holten. 2014. Alpine vegetation along multiple environmental gradients and possible consequences of climate change. Alpine Botany 124 (2):155–64. doi:https://doi.org/10.1007/s00035-014-0136-9.
- Williamson, S. N., I. C. Barrio, D. S. Hik, and J. A. Gamon. 2016. Phenology and species determine growing‐season albedo increase at the altitudinal limit of shrub growth in the sub‐arctic. Global Change Biology 22 (11):3621–31. doi:https://doi.org/10.1111/gcb.13297.
- Wilson, S. D., and C. Nilsson. 2009. Arctic alpine vegetation change over 20 years. Global Change Biology 15 (7):1676–84. doi:https://doi.org/10.1111/j.1365-2486.2009.01896.x.
- Winkler, M., A. Lamprecht, K. Steinbauer, K. Hülber, J. P. Theurillat, F. Breiner, P. Choler, S. Ertl, A. Gutiérrez Girón, and G. Rossi. 2016. The rich sides of mountain summits–a pan‐European view on aspect preferences of alpine plants. Journal of Biogeography 43 (11):2261–73. doi:https://doi.org/10.1111/jbi.12835.
- Yang, F., K. Mitchell, Y.-T. Hou, Y. Dai, X. Zeng, Z. Wang, and X.-Z. Liang. 2008. Dependence of land surface albedo on solar zenith angle: Observations and model parameterization. Journal of Applied Meteorology and Climatology 47 (11):2963–82. doi:https://doi.org/10.1175/2008JAMC1843.1.