
ABSTRACT
The microenvironment in which cancer resides plays an important role in regulating cancer survival, progression, malignancy and drug resistance. Tumor microenvironment (TME) consists of heterogeneous number and types of cellular and non-cellular components that vary in relation to tumor phenotype and genotype. In recent, non-cellular secreted components of microenvironmental heterogeneity have been suggested to contain various growth factors, cytokines, RNA, DNA, metabolites, structural matrix and matricellular proteins. These non-cellular components have been indicated to orchestrate numerous ways to support cancer survival and progression by providing metabolites, energy, growth signals, evading immune surveillance, drug resistance environment, metastatic and angiogenesis cues. Thus, switching action from pro-cancer to anti-cancer activities of these secreted components of TME has been considered as a new avenue in cancer therapeutics and drug resistance. In this report, we summarize the recent pre-clinical and clinical evidences to emphasize the importance of non-cellular components of TME in achieving precision therapeutics and biomarker study.
Introduction
Many new and existing drugs are undergoing successive clinical trials on various cancer tissue models but a majority of them fail due to one or the other reason. Several reports have documented the occurrence of drug resistance, precision of drugs and relapse by using cancer tissue models.Citation2,39,48,73,76 A major cause that has been suggested behind cancer survival, progression, metastasis, failure of drugs and drug resistance is the heterogeneity of tumor microenvironment (TME).Citation3,11,22,36,44,73,76,105,112,125 Tumor progression is significantly contributed by the non-tumor cells surrounding the tumor and secreted non-cellular components, which collectively form the TME.Citation3,45,73,76,105,125 The TME consists of cellular components like cancer associated fibroblasts (CAF's), pericytes, lymphocytes, adipocytes, neutrophils, T-reg cells, mesenchymal stem cells, mast cells and other immune components which play a crucial role in immunosuppression.Citation3,44,54,73,76,125 Indeed, cellular architecture is central in microenvironmental heterogeneity, and secreted non-cellular components are of significant importance in shaping the tumor phenotypes and drug responses.Citation3,44,54,73,76,84,125 Commonly, non-cellular components of TME have been reported to include various types of molecules such as growth factors, cytokines, extracellular matrix (ECM) structural proteins, secreted matricellular proteins, paracrine signaling mediators such as Wnt, BMP group of proteins, small regulatory RNAs, DNA and metabolites.Citation32,44,73,76,125,Citation126 Currently, several pre-clinical and clinical efforts have been documented to harness non-cellular components as potential therapeutic targets and biomarker tools;Citation48,125 Yuan et al.Citation2,109,110,118,126 This review highlights the area of tumor heterogeneity and roles of various components, ECM, extracellular RNA and DNA, exosome, cytokines, growth factors, pH and metabolites as non-cellular heterogeneity. These together drive tumor physiology and drug resistances of cancer, and these are described in the context of pre-clinical and clinical perspectives.
Tumor heterogeneity as a global factor
Tumor heterogeneity is one of the major and global factors behind driving wheels of tumor progression, metastasis and cancer drug resistance.Citation2,39,48,58,73,76,125 Tumor heterogeneity has been described as inter-tumoral and intra-tumoral heterogeneity. Inter-tumoral heterogeneity is due to the presence of different tumor cells in different sub-populations that vary in genotype and phenotype leading to difference in morphology, physiology and anatomy of the tumor. On the other side, intra-tumoral heterogeneity can occur within the cancer cells of only one tumor.Citation74 Yuan et al.Citation126 Intra-tumoral heterogeneity has been suggested to be due to genetic, epigenetic and microenvironmental factors. The rise of tumor heterogeneity in a carcinoma is widely due to the diverse variety of cells found in the TME which are structurally, genetically, physiologically, functionally and anatomically different. The different types of cells found in the TME secrete various types of factors which help in transforming normal cell phenotype to cancer cell phenotype each with different morphology.Citation58,60,67,Citation107
Genetic heterogeneity
In recent, genetic diversity within and between tumors has been called as genetic heterogeneity, and it has been implicated in several signalling pathways, phenotype variations and a roadblock to personalized medicine.Citation11,37,76,98,112 In this type of tumor heterogeneity, major factors that have been reported to contribute to global genomic instability include chromosomal instability, circular extrachromosomal DNA amplification, instability in gene mutation and expression leading to difference in genotype and phenotype of cells.Citation11,37,76,98,112 These genetic mutations play a key role in contributing to genetic heterogeneity which further leads to intra-tumoral heterogeneity.Citation3,11,44,73,76,112,125
Epigenetic heterogeneity
Epigenetic heterogeneity within tumor has been indicated to arise from non-genetic influences on gene expression.Citation2,28,125 This type of tumor heterogeneity means the state of being diverse in content or character with reference to DNA methylation, histone signature and chromatin remodeling, which are independent of DNA sequence.Citation2,33,125 The origin of intra-tumoral heterogeneity has been based on diverse epigenetic regulation of genes and non-coding DNA.Citation2,28,125 Currently, it is well accepted that epigenetic heterogeneity is a crucial factor behind observed hallmarks of cancer, immunomodulation, drug resistance and relapse of cancer.Citation1,10,75,76
Cellular microenvironment heterogeneity
Heterogeneity of TME is a key to cancer progression and development. Tumor cells have diverse nature in terms of morphology, structure, genotype and phenotype.Citation44,73 Cellular microenvironment heterogeneity includes the different types of cellular components found in TME like cancer and non-cancer affected immune cells, adipocytes, fibroblasts, stromal cells and cancer stem cells.Citation3,58,73,76 These cellular components of tumor have been involved in many pro-cancer activities like tumor survival, growth, progression and metastasis.Citation3,44,73,76,82,125
Non-cellular microenvironment in cancer
The phenotype of a tumor is not only dependent on the internal characters of the tumor cells, but is also dependent on its interaction with components of non-tumor cells in the surroundings. These include cytokines, growth factors, ligands, small RNAs, DNA, soluble factors, metabolite, and the non- cellular solid-state extracellular matrix (ECM) that make the TME more amiable for the progression of the tumor.Citation8,Citation42,52,72,92,99,101 The cellular interactions with the non-cellular microenvironment also play a role in governing the tumor metastasis.Citation23 Tumor-stromal cell crosstalk intensifies the production of chemokines, growth factors, cytokines, matrix metalloproteinases (MMPs) which facilitate the tumor growth and angiogenesis.Citation4,59
ECM and its role in tumor progression
In tumor tissue, ECM component has been called as the collection of secreted extracellular molecules from the cellular community, which extend structural and biochemical support to their microenvironment.Citation8 Roberts et al.Citation52,72,92,101 Proteins, glycoproteins and proteoglycans are the major non cellular components of TME, which make the ECM. It regulates functions of cells both structurally and functionally.Citation72,92 The extent of contribution of ECM could be better realized by the fact that more stiffness of cancer tissue may provide a better platform for the modulation of non-cellular microenvironment, which can support cancer growth and invasion (Roberts et al.Citation19,52,101
Structural matrix factors as network of ECM
ECM includes an array of various structures and components. For example, a compact basement membrane (also called as specialized ECM) has been found to be rich in fibronectin, collagen, & laminin. Another component of ECM is described as interstitial matrix consisting of proteoglycans, glycoproteins and collagen, which contribute to the tensile strength of the tissue (Roberts et al.Citation52,72,101 ECM helps in preventing the penetration of targeted drug inside the cancer cells and its uptake by them, forming a tough semi-impermeameable matrix like a tight mess leading to eventual failure of the drug given. Further, various glycoproteins and proteoglycans in the ECM maintain compact cell-cell adhesion, which pose a challenge in drug delivery.Citation52,72 Harris et al.Citation45 suggested the importance of stromal remodeling in pancreatic ductal adenocarcinoma (PDAC), which in turn is a manifestation of non-cellular heterogeneity that could shape the growth and progression of carcinoma. Their data suggest that Serpin B2, a type of ECM component in the stromal compartment can modulate the stromal remodeling and act as a suppressor for PDAC invasion.
Matricellular proteins as components of ECM
In recent, contribution of a plethora of matricellular protein has been viewed as dynamically expressed pool of proteins in the ECM.Citation8 Roberts et al.Citation101 It is believed that these non-structural proteins provide sites for ECM structural proteins and cell surface receptors.Citation8 Roberts et al.Citation101 Additionally, it has been shown that these are involved in the sequestration and modulation of activities of specific growth factors. Among several such matricellular proteins, CCN family of proteins, fibulins, osteopontin, periostin, SPARC family members, tenascin(s), and thrombospondins have been reported to be involved in cancer progression, wound healing and tissue repair.Citation8 Roberts et al.Citation52,101
In a recent clinical study, Psyrri et al.Citation95 suggested that osteopontin, a type of matricellular protein could be a potential therapeutic target and prognostic marker in breast cancer due to its highly elevated level in TME. Among a class of matricellular proteins in TEM, a secreted protein acidic and rich in cysteine (SPARC) has been shown to be linked to aggressiveness of human breast cancer. Güttlein et al.Citation42 revealed the clinical evidence of SPARC in breast carcinoma and highlighted its importance in therapeutics and biomarker study. Recently, Hu et al.Citation49 showed the presence of several secreted proteins, namely SULT2B1, CEACAM5, SPRR3, AGR2, S100P, and S100A14, which can potentially be used as therapeutic targets and biomarkers in non-small cell lung cancer (NSCLC). Tzeng et al.Citation113 supported the notion that modulation of microenvironment could have a significant impact on tumor progression and invasion. Such a study suggests that Rab37-mediated Thrombospondin-1 secretion in cancer cells can suppress metastasis and angiogenesis via a cross-talk with endothelial cells.Citation113 Recently, He et al.Citation47 suggested that embryonic stem cells can release potential factors in conditioned medium. These secreted factors can efficiently suppress activation of signal transducer and activator of transcription 3 pathway in breast cancer cells and act as a significant modulator of microenvironment heterogeneity. In summary, both structural matrix and matricellular components within ECM can modulate the TME in favour of growth, invasion, metastasis and drug evasion. The basic understanding about interplay between ECM and TME has been translated into the therapeutic approaches, which are currently at pre-clinical and clinical stages.
Role of cytokines in shaping non-cellular microenvironment
The non-cellular TME includes various secreted molecules including cytokines, which enhance a two way communication via paracrine signaling between tumor cells and the cancer associated cells in the environment.Citation27,31 These major proteins as cytokines have been suggested to play an important role in the tumor genotype and phenotype.Citation27 A number of cytokines with different functions such as interleukins, tumor necrosis factor family, interferon family and TGF-beta family of proteins have been found in the TME (Dranoff et al.Citation27 Flavio et al., 2007Citation31,44,73 (). Cytokines may influence the formation of tumors by acting directly on the tumor cells as a growth supporting factor and indirectly by evoking inflammatory cell types Flavio et al., 2007.Citation30
Figure 1. This figure depicts the role of cytokines in cancer growth, invasion and drug-resistance. IL-6, an interleukin enhances tumour survival and progression by inducing various DNA damage repair pathways in cancer cells. IL-33 releases various inflammatory factors that cause inflammation in the tumour cells. IL-35 stimulates macrophages to produce many inflammatory signals that cause inflammation.
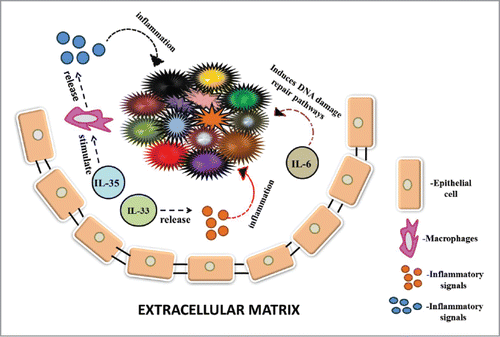
Interleukin-33 (IL-33) has been reported to participate in tumorigenesis through release of inflammatory factors.Citation117 Interleukin 23 (IL-23), a cytokine responsible for causing inflammation is involved in the secretion of TGF-β playing a role in suppressing the immune system and responses. Vascular endothelial growth factor (VEGF) plays a role in angiogenesis. Additionally, IL-10 and IL-23 also play a role in promoting suppression of immune system by decreasing the permeation of CD4+and CD8+T cells into tumor tissues.Citation83 IL-4, a cytokine in the TME of breast cancer induces cancer cell growth, survival and metastasis mediated by MAPK signaling pathway as its blockage leads to decrease in numbers of cancer cell and CSCs.Citation34 IL-10, a growth factor required in signaling of the inflammatory response and invasion of immune cells, acts differently in the TME. It is found to specifically trigger cancer progression and tumor maintenance.Citation7 Interleukin-6 (IL-6) is a major component of almost all TME. Recent study has shed light that IL-6 not only activates inflammation but also controls a number of pro-cancer activities like progression, malignancy and anti-death signaling pathways. IL-6 also acts as a protective shield against DNA damage by inducing pro-cancer signaling repair pathways.Citation61
Transforming growth factor–β (TGF-β) in the TME also plays an essential role in self-renewal, differentiation, maintenance and survival of cancer stem cells which lead to tumor progression and metastasis.Citation91 TGF-β has also been observed to drive breast cancer metastasis by downregulating miR-196a-3p expression.Citation18
A chemokine CCL5 which is secreted by tumor-associated microglia has also been studied and proved to promote the growth of optic glioma in vivo.Citation87 In a clinical study, Cao et al.Citation13 showed evidence about the elevated level of expression of chemokine CXC subfamily of IL-8, GROα, IP-10, and MIG in tumor tissues over tumor-adjacent tissues and normal tissues. Further, pro-inflammatory cytokines, IL-6 and IL-8 have been reported to be linked with senescence-associated secretory phenotype, which act as pro-TME factors in inducing EMT and other pro-cancer activities.Citation106 Another study suggests that purified IL-6 and IL-8 can produce self- and cross-reinforced senescence/inflammatory microenvironment responsible for aggressive phenotypes to a luminal breast cancer cell line.Citation85 There are various types of cytokines like IL-35 and others which suppress or inhibit the normal functioning of immune cells like neutrophils, macrophages thus leading to failure of immunotherapy.Citation129 In summary, cytokines have been perceived as key molecules to establish better cell-cell communication within tumor community, which mostly work in support of pro-cancer microenvironment. Therefore, there is a scope to look for potential mimetics to these growth promoting and inflammatory cytokines as cancer therapeutics.
Exosomes as a messenger in tme
The concept of secretome in shaping TME has seen a wide resurgence. The heterogeneity of secretome has been thought to involve exosomes, small vesicles of endocytic cargo used to ferry bioactive molecules such as proteins, lipids, RNA, DNA and metabolites molecules. Such tumor originated exosomes have been suggested to be important for reprogramming and metastasis of malignant cellsCitation44,70,73 (). The characterization of exosomal content has revealed about the presence of RNA materials including small RNAs, long non-coding RNAs and messenger RNAs.Citation10
Figure 2. This figure illustrates the role of exosomes in cancer progression, malignancy and drug-resistance. Exosomes are secreted by normal and cancer cells. These exosomes act as a mediator of communication between cancer and normal cells acting as a messenger. They are also involved in transport of proteins, nucleic acids and lipids to cancer cells. Exosomes released by cancer cells toughen the extracellular matrix acting as a barrier to entry of drugs inside the cancer cells leading to drug resistance.
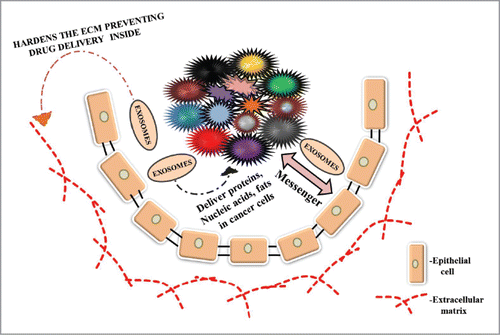
Exosomes have been involved in inter- and intra-cellular communication between cancer cells, normal cells and the environment in which they reside thus acting as carriers of information.Citation109,118 Along with the communication process they are also involved in transporting different types of materials to cancer cells like proteins, fats, different signaling molecules and essential elements.Citation77,103,109,110,115,118
Tumor-derived exosomes also induce NF-κB signaling in macrophages leading to an increase in the activity and release of various inflammatory factors that cause tumor survival, progression and metastasis of gastric carcinoma.Citation121 A recent study has revealed that tumor-derived exosomes carry proteins that have potential to block positive immune response in tumor cells that help to eradicate tumor, thus allowing cancer and normal cells to escape from various immune checkpoints leading to cancer growth and metastasis. They also stimulate various cells to produce different types of signals and factors that help in cracking down the immunosurveillance system leading to cancer progression and malignancy.Citation119 Exosomes are also involved in loss of adhesion of the cancer cells leading to increase in motility of cancer cells resulting in metastasis.Citation110
There are many evidences which support that Tumor-derived (TD) exosomes released from tumor tissue are involved in drug resistance by removing the drug given and also promote other cells to produce factors and signals that negatively regulate immune response leading to jumping of cancer cells from various immune checkpoints.Citation115 Various studies have revealed that exosomes secrete various proteins that are involved in growth of fibrous tissue around tumor cells that help in resisting effective permeability of drugs inside the tumor cells.Citation5 In summary, idea of exosome contribution in shaping TME has received wide attention, due to its ability to facilitate export and import of potential key messengers driving pro-cancer events. Therefore, importance lies here that exosome could be exploited in two ways, first in biomarker study and second as a tool to bring ectopic applications of drugs/inhibitors as anticancer agents.
Extra-cellular RNA and DNA in shaping microenvironmental heterogeneity
In recent, the roles of small non-coding RNAs have been at the forefront in cancer therapeutics due to their role as both tumour suppressors and oncogenes. A predominant class of small RNAs as microRNAs (miRNAs) have been widely reported in cancer progression, invasion and metastasis by regulating expression of various genes.Citation16,29,63,100 More recently, besides intracellular regulation and biological functions, these small RNAs including miRNAs have been considered as a crucial actor in TME based heterogeneity due to differential extracellular space in tumor Evans-Knowell et al.Citation30 These small RNA species have been involved in communication between cancer cells in their respective TME.Citation63,69,114 As an additional evidence Challagundla et al.Citation17 reported that exosomic miRNA-21 and miRNA-155 can be suggested as a communication bridge between neuroblastoma cells and human monocytes and subsequent observations of drug resistance. Another evidence from Chugh et al.Citation21 suggested about the secreted nature of miRNAs from host as miRNA-17-92 cluster and circulating miRNA profiles from KSHV mouse models. MiRNA-210 found in the TME of Osteosarcoma (OS) has been reported to play a role in conversion of Osteosarcoma cells to OS stem cells leading to tumor growth and progression.Citation127 MiRNA-210 has also been found in the TME of breast cancer tissue.Citation6 Bott et al.Citation9 reported the presence of miRNA-1246 in the TME of breast cancer, which can induce the release of inflammatory factors like interleukins such as IL-6 and IL-8, which can participate in tumor inflammation and promotion of NF-κB signaling.
Now, it is a widely accepted notion that DNA can be leaked and possibly exported from the dead and dying cells into the extracellular spaces. In case of a tumor, DNase I can also be secreted to extra-cellular space and it is suggested that it acts as an agent of waste disposal system in the human system to attain degradation of extra-cellular DNA.Citation101 There is also evidence that extracellular DNA may be a component of neutrophil extracellular traps, which may work as an anti-tumor immune response system.Citation43 Besides the possibilities of leakage of DNA to extracellular space, there is an emerging evidence of exosome mediated transport to achieve cell-cell communication in TME.Citation46,53 In summary, the presence of extracellular RNA and DNA in TME has opened up new avenues to strengthen for prognosis and diagnosis, approaches and also has provided a platform to visualize the contribution of these RNA/DNA molecules in pro-cancer activities ().
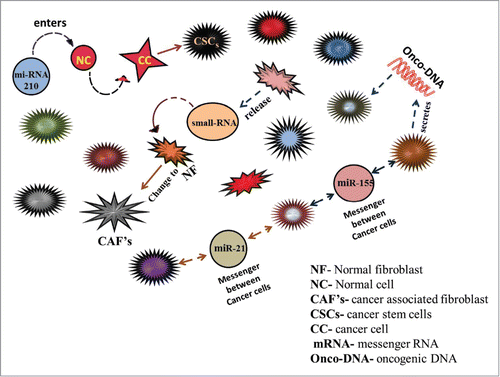
Figure 3. This figure describes the role of extracellular secreted RNAs (small RNAs and mRNA) and DNAs in tumor growth, transformation, progression and communication. MiRNA 210 is released by cancer developing cells and normal cells in tumor derived exosomes. MiRNA 210 is taken by normal cells leading to change in Mi-RNA levels further leading to transformation of normal cells to cancer cells. MiRNA21 and miRNA155 are released by normal and cancer cells which act as messenger between different cells of the tumor playing a role in communication. Cancer cells also secrete oncogenic DNA which converts normal cells to tumor cells.
Modulation of microenvironment heterogeneity by growth factors
A plethora of molecular components in the microenvironment have shown their ability to modulate the growth of tumor cells, which progress and metastasize. A major class of such molecules in TME has been described as polypeptide growth factors that are released from a cell and interact with growth receptors present on target cells. Different types of growth factors mediate cell-cell communications with distinct cellular populations, which is mediated by unique set of growth receptors.
There are various growth factors like vascular endothelial growth factor (VEGF) which plays a major role in angiogenesis providing nutritional supply to cancer cells through blood and metastasis.Citation44,73,76,93,125 Yuan et al.Citation126 Hepatocyte growth factor (HGF) that is involved in intercellular communication via paracrine signaling promotes change of normal functioning of fibroblasts to cancer associated fibroblasts (CAF's) thus helping in tumor heterogeneity, progression and malignancy.Citation122 HGF also causes epithelial to mesenchymal transition (EMT) leading to invasion of cancer cells.Citation86 Various glycoproteins are found in the TME with diverse kind of roles. Fibulin-3, a glycoprotein secreted by glioblastoma cells induces NF-κB signaling leading to proliferation and metastasis of cancer cells along with secretion of tumor necrosis factor alpha (TNFα).Citation81 Progranulin (PGRN), a glycoprotein found in the TME of colorectal cancer (CRC) enhances tumor progression and malignancy by conversion of normal functioning of fibroblasts to cancer associated fibroblasts (CAF's).Citation114
The insulin-like growth factor-1 (IGF-1) and IGF-1 receptor based communication has been suggested to induce cells to undergo epithelial to mesenchymal transition and promote migration and invasion.Citation44,73 Another growth factor, platelet derived growth factor has been suggested to establish cellular communication among stromal cells, notably fibroblasts, myofibroblasts and macrophages for growth, survival and suitable TME.Citation76,125 Yuan et al.Citation126 One of the secreted molecules, sphingosine-1-phosphate produced through the metabolism of cancer and non-cancer cells surrounding the tumor is derived outside the cells and it plays an essential role in tumor survival and progression.Citation80 In summary, the importance of growth factors in TME has been well known. Recent accumulations of evidence in the form of pre-clinical and clinical levels are encouraging and may lead to additional therapeutic tools other than existing drugs/inhibitors regimens.
pH as a factor to turn TME
In view of the Warburg effects, altered pH has been considered as one of the common hallmarks and also a factor for promoting tumour progression;Citation44,76,125 Yuan et al.Citation126 Further, a decrease in the pH causes damage to the ECM proteins which is followed by invasion and metastasis of the cancer cells from the tumour tissue to the external environment. A therapeutic approach in targeting the tumour tissue may include the reversal in the state of pH in and outside the matrix like decrease in the pH intracellularly and increase in the pH extracellularly through any type of drug mechanism.Citation57
Secreted metabolites and microenvironmental heterogeneity
Intratumoral heterogeneity has been perceived as one of the pivotal hurdles in cancer therapy success. Among potential components as the contributor of heterogeneity, extracellular gradients of metabolites can act in creating phenotypic diversity of cells in the TME.Citation44,73,76,125 Yuan et al.Citation97,126 There are evidences to support that extracellular metabolites can behave as tumor morphogens that shape up a unique tumor heterogeneity.Citation14 It is widely accepted that CAFs residing in the TME can promote the growth of cancer cells by secreting essential energy-rich metabolites, including lactate, ketone bodies, fatty acids, glutamine, and other amino acids in extracellular space.Citation97,120 In a recent paper, Loo et al.Citation71 reported on the ability of miRNA-551a and miRNA-483 in regulating creatine kinase, brain-type (CKB), which is responsible for the accumulation of phosphocreatine from metabolite creatine. Such accumulation of phosphocreatine has been indicated to accentuate the metastatic survival.
Therapeutic approaches to modulate tme
The TME can affect the success of chemotherapy, as the resistance of cancer drugs has been focused on cancer cells.Citation1,39,48,55,68,78,84,86,88,89 There are various ways in which TME plays a role in drug resistance such as by not allowing the drug to enter inside the tumor tissue by formation of a tough ECM network, modulation of cell-cell communication and changes in the level of soluble components including matricellular proteins, cytokines, metabolites, RNA, DNA and lipid.Citation2,46,48,79,97 Various components present in the TME help the tumor tissue to switch on to some other survival pathways thus leading to failure of the drug given as there are many ways for survival of cancer cells.Citation73,76,125 Yuan et al.Citation126
An important category of secreted components within TME as nano-vesicle packaged miRNAs have been suggested to act as tumor suppressors, oncogenic and immune modulators.Citation35,40,56,69,104,124 Based on these attributes, secreted exosomal miRNAs have been viewed as significant players in tumor prognosis, diagnosis, therapeutics and post therapy monitoring of cancer patients (Seliger et al.Citation104 One of them is miRNA-7 that suppresses autophagy inducing factors in the TME of pancreatic carcinoma along with glycolysis metabolism leading to inhibition of carcinoma growth and malignancy.Citation40 MiRNA-199a-3p in the TME of prostate carcinoma blocks specifically the progression of cancer stem cells of prostate gland and also stops tumor formation serving as therapeutic approach in targeting it.Citation69 MiRNA-29b stops tumor progression, invasion and malignancy by blocking Akt3 which induces cancer cell growth and metastasis as it is found in the TME and tumor cells.Citation63 Downregulation of various miRNAs have been linked with various types of cancers like decrease of miRNA-126 and miRNA-7 level is linked to endocrine cancer.Citation64 While under expression of miRNA-125b, miRNA-145 and miRNA-21 is linked to breast cancer and many more. MiRNA-126 (miRNA-126) is found in the TME of almost all major types of tumors. This can serve as a biomarker for TME based targeted therapy.Citation124
A number of cytokines are discovered for their anti-oncogenic role.Citation38,44,73,76,125, Yuan et al.Citation24,79,126 One of them is Interleukin-31 (IL31) that reduces the number of tumor cells by controlling blood vessel formation and malignancy.Citation24 Among a pool of secreted factors in TME, a form cytokine as CC chemokine ligands (CCL)-5, 20 has been associated with tumor progression and drug resistance. These secreted forms of cytokines as CCL-5 and CCL-20 have been presented as promising cancer therapeutic targets by using small RNAs interference and pharmacological inhibitory approaches.Citation79 A new class of cytokine osteoprotegerin has been reported to modulate TME by displaying crosstalk with fatty acid synthase (FASN) and, cycloxygenase-2. Collectively, osteoprotegerin, fatty acid synthase and cycloxygenase-2 have been suggested as therapeutic targets and suggest the combinatorial anti-cancer treatment options.Citation38
Peptide (along with derivatives) based therapy supported with nano-modifications can serve as a better therapeutic approach to target the TME's cellular and non- cellular components.Citation12,15,20,96,123 These classes of drugs are known for their selectivity, efficiency, amphoteric nature and these are modifiers of biological processes, acting as signalling molecules and passing easily through plasma membrane.Citation96 A most common and effective biomarker in cancer is the low pH of cancer cells. pH (low) insertion peptides (pHLIP®s) serve as new and promising approach in targeting as it helps in imaging of tumor cells, transport of drug molecules across the lipid bilayer and absorption of the targeted drug given.Citation123 Evidence has shown that a newly developed peptide with heparin-binding activity suppresses tumor progression and metastasis by blocking angiogenesis, acting as an angiogenesis inhibitor in targeting the TME.Citation20 Among other secreted proteins, evidences have supported the importance of extracellular secreted exosomal heat shock protein 60 (HSP60) as a modulator of TME. These HSP60 have been linked to the anticancer effects of Suberoylanilide hydroxamic acid, an inhibitor of histone deacetylase family of enzyme.Citation12,15 Another chaperon-like enhancer molecule anterior gradient-2 secreted from tumor cells has been shown to activate VEGF and fibroblast growth factor and suggested as a potential therapeutic target.Citation41
Other than exosomal miRNAs, secreted cytokines and extracellular peptides, tumor secreted metabolites have been seen as a new class of therapeutic targets and tumor biomarkers.Citation26,97,102,108 For an example, Salimian Rizi et al.Citation102 reported that use of inhibitor of arginine, secreted metabolite in TME can serve as therapeutic by dismantling the metabolic crosstalk between developmental adipose stromal cells and endometrial and ovarian tumor cells. Additional evidence by Stadler et al.Citation108 suggested that colorectal cancer cell secreted 12-Hydroxyeicosatetraenoic acid (12-HETE), a product of lipid metabolism can change TME in favor of enhanced invasiveness. Further, authors proposed the therapeutic use of inhibitors to calcium signaling pathways provoked by 12-HETE.
Nanoparticle based therapeutic approach towards targeting the TME is a better option due to drug resistance offered by tumour cells (Adjei and Blanka 2015Citation1; Praneeth et al., 2016.Citation25,51,90,95 These nanoparticles due to their highly selective nature towards targeting particular cellular and non-cellular components in the TME are effective in treatment options.Citation90 Drug delivery systems made from natural and artificial sources deliver drugs to kill the stromal cells and reschedule the microenvironment for tumor development (Adjei and Blanka 2015Citation1). Synthetic miRNA can be used for targeting miRNA to stromal cells as they have a therapeutic possibility to modify the TME (Praneeth et al., 2016). Recent studies and evidences prove that exosomes can act as a biomarker to target the TME.Citation50 Not only exosomes but exosomes loaded with various particles like miRNAs and proteins can also serve as a potent and functional biomarker in targeting TME as their expression levels vary in different types of cancers.Citation25,51
In summary, numerous pre-clinical and clinical data supported the notion that modulation of non-cellular microenvironment can bring promising cancer therapy outcomes. Among potential secreted molecules within TME, nano-vesicle packaged miRNAs, extracellular peptides, growth factors, cytokines, chaperones and metabolites have been widely viewed as therapeutic targets to disrupt the pro-tumor niches. Importantly, pharmacological inhibitors, small RNA based interference and mimetic agents to target non-cellular components could be presented as promising combinatorial therapeutic options in combination with existing drugs such as genotoxic agents, epigenetic modulators and inhibitors to signalling pathways. However, these therapeutic options could also face certain bottlenecks in terms of intra-cellular and inter-organelle delivery, stability, drug dose related side effects, harm to the normal tissues, need of prior knowledge of genetic and epigenetic status of patient, cost effectiveness, potential drug resistance and relapse of cancer. Therefore, therapeutics aimed at non-cellular components within tumor should be critically evaluated with potential promises and pitfalls, so that success of these drugs in combinations with existing anti-cancer agents will be a reality at preclinical and clinical stages.
Conclusion
In conclusion, understanding the cellular and molecular pathways unleashed by secreted components in TME has paved the way to achieve cancer treatment options beyond the scope of genotoxic drug therapy. Several mimetic drug approaches have been endeavored to modulate friendly TME in such a way that it becomes hostile and limiting for growth, progression and metastasis. These therapeutic options centered on non-cellular components have been given preference over commonly used cancer treatment regimens because of their scope to enter to the precision drug therapy and personalize treatment. Interestingly, selective drugs to modulate non-cellular TME have been suggested for combinatorial drug therapy for complete and sustainable eradication of tumor cells as well as non-tumor supporting neighboring cells. Despite tremendous progress in understanding of non-cellular microenvironmental heterogeneity, there is a need for a better cancer model, where a true representative microenvironment could be employed and tested for the success of drugs.
Conflict of interest
The authors convincingly declare no conflict of interest.
Acknowledgment
The authors acknowledge financial support from DST-SERB, Government of India, New Delhi, India (SERB/LS-1028/2013) and Dr. D. Y Patil, Vidyapeeth, Pune, India (DPU/05/01/2016).
References
- Adjei IM, and Blanka S. Modulation of the Tumor Microenvironment for Cancer Treatment: A Biomaterials Approach. J Funct Biomater. 2015;6(1):81–103. doi:10.3390/jfb6010081.
- Ahuja N, Sharma AR, Baylin SB. Epigenetic Therapeutics: A New Weapon in the War Against Cancer. Annu Rev Med. 2016;67:73–89. doi:10.1146/annurev-med-111314-035900.
- Alizadeh AA, Aranda V, Bardelli A, Blanpain C, Bock C, Borowski C. Toward understanding and exploiting tumor heterogeneity. Nat Med. 2015;21(8):846–53. doi:10.1038/nm.3915.
- Andreuzzi E, Colladel R, Pellicani R, Tarticchio G, Cannizzaro R, Spessotto P, Bussolati B, Brossa A, De Paoli P, Canzonieri V, Iozzo RV, Colombatti A, Mongiat M. The angiostatic molecule Multimerin 2 is processed by MMP-9 to allow sprouting angiogenesis. Matrix Biol. 2017. doi: 10.1016/j.matbio.2017.04.002.
- Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev. 2013;32(3–4):623–42. doi:10.1007/s10555-013-9441-9.
- Bar I, Merhi A, Abdel-Sater F, Ben Addi A, Sollennita S, Canon JL, Delrée P. The MicroRNA miR-210 Is Expressed by Cancer Cells but Also by the Tumor Microenvironment in Triple-Negative Breast Cancer. J Histochem Cytochem. 2017;65(6):335–346. doi:10.1369/0022155417702849.
- Berti FC, Pereira AP, Cebinelli GC, Trugilo KP, Brajão de Oliveira K. The role of interleukin 10 in human papilloma virus infection and progression to cervical carcinoma. Cytokine Growth Factor Rev. 2017;34:1–13. doi:10.1016/j.cytogfr.2017.03.002.
- Bornstein P, Sage EH. Matricellular proteins: extracellular modulators of cell function”. Curr. Opin. Cell Biol. 2002;14(5): 608–16. doi:10.1016/S0955-0674(02)00361-7.
- Bott A, Erdem N, Lerrer S, Hotz-Wagenblatt A, Breunig C, Abnaof K, Wörner A, Wilhelm H, Münstermann E, Ben-Baruch A, Wiemann S. miRNA-1246 induces pro-inflammatory responses in mesenchymal stem/stromal cells by regulating PKA and PP2A. Oncotarget. 2017;8(27):43897–43914.
- Braicu C, Tomuleasa C, Monroig P, Cucuianu A, Berindan-Neagoe I, Calin GA. Exosomes as divine messengers: are they the Hermes of modern molecular oncology? Cell Death Differ. 2015;22(1):34–45. doi:10.1038/cdd.2014.130.
- Burrell RA, McGranahan N, Bartek J, Swanton C. The causes and consequences of genetic heterogeneity in cancer evolution. Nature. 2013;501(7467):338–45. doi:10.1038/nature12625.
- Campanella C, D'Anneo A, Marino Gammazza A, Caruso Bavisotto C, Barone R, Emanuele S. The histone deacetylase inhibitor SAHA induces HSP60 nitration and its extracellular release by exosomal vesicles in human lung-derived carcinoma cells. Oncotarget. 2016;7(20):28849–67. doi:10.18632/oncotarget.6680.
- Cao Y, Huang H, Wang Z, Zhang G. The Inflammatory CXC Chemokines, GROαhigh, IP-10low, and MIGlow, in Tumor Microenvironment Can Be Used as New Indicators for Non-small Cell Lung Cancer Progression. Immunol Invest. 2017;46(4):361–374. doi:10.1080/08820139.2017.1280052.
- Carmona-Fontaine C, Deforet M, Akkari L, Thompson CB, Joyce JA, Xavier JB. Metabolic origins of spatial organization in the tumor microenvironment. Proc Natl Acad Sci U S A. 2017;114(11):2934–2939. doi:10.1073/pnas.1700600114.
- Caruso Bavisotto C, Cappello F, Macario AJ, Conway de Macario E, Logozzi M4, Fais S. Exosomal HSP60: a potentially useful biomarker for diagnosis, assessing prognosis, and monitoring response to treatment. Expert Rev Mol Diagn. 2017;17(9):815–822. doi:10.1080/14737159.2017.1356230.
- Ceppi P, Peter ME. MicroRNAs regulate both epithelial-to-mesenchymal transition and cancer stem cells. Oncogene. 2014;33(3):269–78. doi:10.1038/onc.2013.55.
- Challagundla KB, Wise PM, Neviani P, Chava H, Murtadha M, Xu T. et al. Exosome-mediated transfer of microRNAs within the tumor microenvironment and neuroblastoma resistance to chemotherapy. J Natl Cancer Inst. 2015;107(7). doi:10.1093/jnci/djv135.
- Chen Y, Huang S, Wu B, Fang J, Zhu M, Sun L, Zhang L, Zhang Y, Sun M, Guo L, Wang S. Transforming growth factor-β1 promotes breast cancer metastasis by downregulating miR-196a-3p expression. Oncotarget. 2017;8(30):49110–49122.
- Chin L, Xia Y, Discher DE, Janmey PA. Mechanotransduction in cancer. Curr Opin Chem Eng. 2016;11:77–84. doi:10.1016/j.coche.2016.01.011.
- Choi SH, Lee JY, Suh JS, Park YS, Chung CP, Park YJ. Dual-function synthetic peptide derived from BMP4 for highly efficient tumor targeting and antiangiogenesis. Int J Nanomedicine. 2016;11:4643–4656. doi:10.2147/IJN.S115044.
- Chugh PE, Sin SH, Ozgur S, Henry DH, Menezes P, Griffith J. et al. Systemically circulating viral and tumor-derived microRNAs in KSHV-associated malignancies. PLoS Pathog. 2013;9(7):e1003484. doi:10.1371/journal.ppat.1003484.
- Chung HW, Lim JB. Role of the tumor microenvironment in the pathogenesis of gastric carcinoma. World J Gastroenterol. 2014;20(7):1667–80. doi:10.3748/wjg.v20.i7.1667.
- Cretu A, Brooks PC. Impact of the non-cellular tumor microenvironment on metastasis: potential therapeutic and imaging opportunities. J Cell Physiol. 2007;213(2). 391–402. doi:10.1002/jcp.21222.
- Davidi S, Fremder E, Kan T, Raviv Z, Timaner M, Karin N, Hershkovitz D, Arohneim A, Shaked Y. The antiangiogenic role of the pro-inflammatory cytokine interleukin-31. Oncotarget. 2017;8(10):16430–16444.
- Dejima H, Iinuma H, Kanaoka R, Matsutani N, Kawamura M. Exosomal microRNA in plasma as a non-invasive biomarker for the recurrence of non-small cell lung cancer. Oncol Lett. 2017;13(3):1256–1263.
- Devashree J, Purohit S, Jain A, Sharma NK. Export of short RNAs: A bridge between breast carcinoma and their neighboring cells”. Front. Oncol. 2016;6:147.
- Dranoff G. Cytokines in cancer pathogenesis and cancer therapy. Nat Rev Cancer. 2004;4(1):11–22. doi:10.1038/nrc1252.
- Easwaran H, Tsai HC, Baylin SB. Cancer epigenetics: tumor heterogeneity, plasticity of stem-like states, and drug resistance. Mol Cell. 2014;54(5):716–27. doi:10.1016/j.molcel.2014.05.015.
- Eichmüller SB, Osen W, Mandelboim O, Seliger B. Immune Modulatory microRNAs Involved in Tumor Attack and Tumor Immune Escape. J Natl Cancer Inst. 2017;109(10).
- Evans-Knowell A, LaRue AC, Findlay VJ. MicroRNAs and Their Impact on Breast Cancer, the Tumor Microenvironment, and Disparities. Adv Cancer Res. 2017;133:51–76. doi:10.1093/jnci/djx034.
- Fernandes JV, Cobucci RN, Jatobá CA, Fernandes TA, de Azevedo JW, de Araújo JM. The role of the mediators of inflammation in cancer development. Pathol Oncol Res. 2015;21(3):527–34. doi:10.1007/s12253-015-9913-z.
- Salazar-Onfray F, López MN, Mendoza-Naranjo A. Paradoxical effects of cytokines in tumor immune surveillance and tumor immune escape. Cytokine Growth Factor Rev. 2007;18(1-2):171–82. doi:10.1016/j.cytogfr.2007.01.015.
- Flower KJ, Ghaem-Maghami S, Brown R. Is there a role for epigenetic enhancement of immunomodulatory approaches to cancer treatment? Curr Cancer Drug Targets. 2017 doi: 10.2174/1568009617666170206105131.
- Gaggianesi M, Turdo A, Chinnici A, Lipari E, Apuzzo T, Benfante A, Sperduti I, Di Franco S, Meraviglia S, Lo Presti E, Dieli F, Caputo V, Militello G, Vieni S, Stassi G, Todaro M. IL-4 primes the dynamics of breast cancer progression via DUSP4 inhibition. Cancer Res. 2017;77(12):3268–3279. doi:10.1158/0008-5472.CAN-16-3126.
- Gandellini P, Doldi V, Zaffaroni N. microRNAs as players and signals in the metastatic cascade: Implications for the development of novel anti-metastatic therapies. Semin Cancer Biol. 2017;44:132–140. doi:10.1016/j.semcancer.2017.03.005.
- Gkretsi V, Stylianou A, Papageorgis P, Polydorou C, Stylianopoulos T. Remodeling Components of the Tumor Microenvironment to Enhance Cancer Therapy. Frontiers in Oncology. 2015;5:214. doi:10.3389/fonc.2015.00214.
- Goncearenco A, Rager SL, Li M, Sang QX, Rogozin IB, Panchenko AR. Exploring background mutational processes to decipher cancer genetic heterogeneity. Nucleic Acids Res. 2017. doi: 10.1093/nar/gkx367.
- Goswami S, Sharma-Walia N. Crosstalk between osteoprotegerin (OPG), fatty acid synthase (FASN) and, cycloxygenase-2 (COX-2) in breast cancer: implications in carcinogenesis. Oncotarget. 2016;7(37):58953–58974. doi:10.18632/oncotarget.9835.
- Gottesman MM. Mechanisms of cancer drug resistance. Annu Rev Med. 2002;53. 615–627. doi:10.1146/annurev.med.53.082901.103929.
- Gu DN, Jiang MJ, Mei Z, Dai JJ, Dai CY, Fang C, Huang Q, Tian L. microRNA-7 impairs autophagy-derived pools of glucose to suppress pancreatic cancer progression. Cancer Lett. 2017;400:69–78. doi:10.1016/j.canlet.2017.04.020.
- Guo H, Zhu Q, Yu X, Merugu SB, Mangukiya HB, Smith N. et al. Tumor-secreted anterior gradient-2 binds to VEGF and FGF2 and enhances their activities by promoting their homodimerization. Oncogene. 2017;36(36):5098–5109. doi: 10.1038/onc.2017.132.
- Güttlein LN, Benedetti LG, Fresno C, Spallanzani RG, Mansilla SF, Rotondaro C. et al. Predictive Outcomes for HER2-enriched Cancer Using Growth and Metastasis Signatures Driven By SPARC. Mol Cancer Res. 2017;15(3):304–316.doi:10.1158/1541-7786.MCR-16-0243-T.
- Halverson TWR, Wilton M, Poon KKH, Petri B, Lewenza S. DNA is an antimicrobial component of neutrophil extracellular traps. PLoS Pathog. 2015;11:e1004593. doi:10.1371/journal.ppat.1004593.
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74. doi:10.1016/j.cell.2011.02.013.
- Harris NL, Vennin C, Conway JR, Vine KL, Pinese M, Cowley MJ, et al. SerpinB2 regulates stromal remodelling and local invasion in pancreatic cancer. Oncogene. 2017;36(30):4288–4298. doi:10.1038/onc.2017.63.
- Hawes MC, Wen F, Elquza E. Extracellular DNA: A Bridge to Cancer. Cancer Res. 2015;75(20):4260–4. doi:10.1158/0008-5472.CAN-15-1546.
- He N, Feng G4, Li Y, Xu Y, Xie X, Wang H, et al. Embryonic stem cell preconditioned microenvironment suppresses tumorigenic properties in breast cancer. Stem Cell Res Ther. 2016;7(1):95. doi:10.1186/s13287-016-0360-x.
- Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG. Cancer drug resistance: an evolving paradigm. Nat Rev Cancer. 2013;13(10):714–26. doi:10.1038/nrc3599.
- Hu R, Huffman KE, Chu M, Zhang Y, Minna JD, Yu Y. Quantitative Secretomic Analysis Identifies Extracellular Protein Factors That Modulate the Metastatic Phenotype of Non-Small Cell Lung Cancer. J Proteome Res. 2016;15(2):477–86. doi:10.1021/acs.jproteome.5b00819.
- Jia Y, Chen Y, Wang Q, Jayasinghe U, Luo X, Wei Q, Wang J, Xiong H, Chen C, Xu B, Hu W, Wang L, Zhao W, Zhou J. Exosome: emerging biomarker in breast cancer. Oncotarget. 2017;8(25):41717–41733.
- Jiao C, Jiao X, Zhu A, Ge J, Xu X. Exosomal miR-34s panel as potential novel diagnostic and prognostic biomarker in patients with hepatoblastoma J Pediatr Surg. 2017;52(4):618–624. doi:10.1016/j.jpedsurg.2016.09.070.
- Jun JI, Lau LF. Taking aim at the extracellular matrix: CCN proteins as emerging therapeutic targets”. Nat Rev Drug Discov. 2011;10(12):945–63. doi:10.1038/nrd3599.
- Kalluri R, LeBleu VS. Discovery of Double-Stranded Genomic DNA in Circulating Exosomes. Cold Spring Harb Symp Quant Biol. 2017;81:275–280. doi:10.1101/sqb.2016.81.030932.
- Kalluri R, Zeisberg M. Fibroblast in cancer. Nat Rev Cancer. 2006;6(5):392–401. doi:10.1038/nrc1877.
- Kenny PA, GY Lee, MJ Bissell. Targeting the tumor microenvironment. Front Biosci. 2007;(12):3468–3474. doi:10.2741/2327.
- Kinoshita T, Yip KW, Spence T, Liu FF. MicroRNAs in extracellular vesicles: potential cancer biomarkers. J Hum Genet. 2017;62(1):67–74. doi:10.1038/jhg.2016.87.
- Koltai T Cancer: fundamentals behind pH targeting and the double-edged approach. Onco Targets Ther. 2016;9:6343–6360. doi:10.2147/OTT.S115438.
- Konrad CV, Murali R, Varghese BA, Nair R. The role of cancer stem cells in tumor heterogeneity and resistance to therapy. Can J Physiol Pharmacol. 2017;95(1):1–15. doi:10.1139/cjpp-2016-0079.
- Koontongkaew S. The tumor microenvironment contribution to development, growth, invasion and metastasis of head and neck squamous cell carcinomas. J Cancer. 2013;4(1). 66–83. doi:10.7150/jca.5112.
- Koren E, Fuchs Y. The bad seed: Cancer stem cells in tumor development and resistance. Drug Resist Updat. 2016;28:1–12. doi:10.1016/j.drup.2016.06.006.
- Kumari N, Dwarakanath BS, Das A, Bhatt AN. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumour Biol. 2016;37(9):11553–11572. doi:10.1007/s13277-016-5098-7.
- Li H, Fan X, and Houghton J. Tumor Microenvironment: The Role of the Tumor Stroma in Cancer. Journal of Cellular Biochemistry. 2007;101(4):805–815. doi:10.1002/jcb.21159.
- Li SC, Vu LT, Luo JJ, Zhong JF, Li Z, Dethlefs BA, Loudon WG, Kabeer MH. Tissue elasticity bridges cancer stem cells to the tumor microenvironment through microRNAs: Implications for a “watch-and-wait” approach to cancer. Curr Stem Cell Res Ther. 2017;12(6):455–470. doi: 10.2174/1574888X12666170307105941.
- Li Y, Cai B, Shen L, Dong Y, Lu Q, Sun S, Liu S, Ma S, Ma PX, Chen J. MiRNA-29b suppresses tumor growth through simultaneously inhibiting angiogenesis and tumorigenesis by targeting Akt3. Cancer Lett. 2017;397:111–119. doi:10.1016/j.canlet.2017.03.032.
- Li Y, Su J, Li F, Chen X, Zhang G. MiR-150 regulates human keratinocyte proliferation in hypoxic conditions through targeting HIF-1α and VEGFA: Implications for psoriasis treatment. PLoS One. 2017;12(4):e0175459. doi:10.1371/journal.pone.0175459.
- Lima CR, Gomes CC, Santos MF. Role of microRNAs in endocrine cancer metastasis. Mol Cell Endocrinol. 15(456):62–75. doi: 10.1016/j.mce.2017.03.015.
- Lindeman GJ, Visvader JE. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer.2008;8(10):755–768. doi:10.1038/nrc2499.
- Liotta LA, Kohn E. Anoikis: cancer and the homeless cell. Nature. 2004;430(7003):973–974. doi:10.1038/430973a.
- Liu R, Liu C, Zhang D, Liu B, Chen X, Rycaj K, Jeter C, Calhoun-Davis T, Li Y, Yang T, Wang J, Tang DG. MiR-199a-3p targets stemness-related and mitogenic signaling pathways to suppress the expansion and tumorigenic capabilities of prostate cancer stem cells. Oncotarget. 2016;7(35):56628–56642. doi:10.18632/oncotarget.10652.
- Lobb RJ, Lima LG, Möller A. Exosomes: Key mediators of metastasis and pre-metastatic niche formation. Semin Cell Dev Biol. 2017;67:3–10. doi:10.1016/j.semcdb.2017.01.004.
- Loo JM, Scherl A, Nguyen A, Man FY, Weinberg E, Zeng Z, et al. Extracellular metabolic energetics can promote cancer progression. Cell. 2015;160(3):393–406. doi:10.1016/j.cell.2014.12.018.
- Lu P, Weaver VM, Werb Z. The extracellular matrix: a dynamic niche in cancer progression. J Cell Biol. 2012;196(4):395–406. doi:10.1083/jcb.201102147.
- Marusyk A, Almendro V, Polyak K. Intra-tumour heterogeneity: a looking glass for cancer? Nat Rev Cancer. 2012;12(5):323–34. doi:10.1038/nrc3261.
- Marusyk A, Polyak K. Tumor heterogeneity: causes and consequences. Biochim Biophys Acta. 2010;1805(1):105–17.
- Mazor T, Pankov A, Song JS, Costello JF. Intratumoral Heterogeneity of the Epigenome. Cancer Cell. 2016;29(4):440–51. doi:10.1016/j.ccell.2016.03.009.
- Meacham CE, Morrison SJ. Tumour heterogeneity and cancer cell plasticity. Nature. 2013;501(7467):328–37. doi:10.1038/nature12624.
- Meehan K, Vella LJ. The contribution of tumour-derived exosomes to the hallmarks of cancer. Crit Rev Clin Lab Sci. 2016;53(2):121–31. doi:10.3109/10408363.2015.1092496.
- Mumenthaler SM, Foo J, Choi NC, Heise N, Leder K, Agus DB, Pao W, Michor F, Mallick P. The Impact of Microenvironmental Heterogeneity on the Evolution of Drug Resistance in Cancer Cells. Cancer Inform. 2015;14(Suppl 4):19–31.
- Muscella A, Vetrugno C, Marsigliante S. CCL20 promotes migration and invasiveness of human cancerous breast epithelial cells in primary culture. Mol Carcinog. 2017;56(11):2461–2473. doi: 10.1002/mc.22693.
- Nakajima M, Nagahashi M, Rashid OM, Takabe K, Wakai T. The role of sphingosine-1-phosphate in the tumor microenvironment and its clinical implications. Tumour Biol. 2017;39(4):1010428317699133. doi:10.1177/1010428317699133.
- Nandhu MS, Kwiatkowska A, Bhaskaran V, Hayes J, Hu B, Viapiano MS. Tumor-derived fibulin-3 activates pro-invasive NF-κB signaling in glioblastoma cells and their microenvironment. Oncogene. 2017;36(34):4875–4886. doi:10.1038/onc.2017.109.
- Natrajan R, Sailem H, Mardakheh FK, Arias Garcia M, Tape CJ, Dowsett M, Bakal C, Yuan Y. Microenvironmental Heterogeneity Parallels Breast Cancer Progression: A Histology-Genomic Integration Analysis. PLoS Med. 2016; 16;13(2):e1001961 doi:10.1371/journal.pmed.1001961.
- Nie W, Yu T, Sang Y, Gao X. Tumor-promoting effect of IL-23 in mammary cancer mediated by infiltration of M2 macrophages and neutrophils in tumor microenvironment. BiochemBiophys Res Commun. 2017;482(4):1400–1406. doi:10.1016/j.bbrc.2016.12.048.
- Nilendu P, Kumar A, Kumar A, Pal JK, Sharma NK. Breast cancer stem cells as last soldiers eluding therapeutic burn: A hard nut to crack. International Journal of Cancer. 2017. DOI: 10.1002/ijc.30898.
- Ortiz-Montero P, Londoño-Vallejo A, Vernot JP. Senescence-associated IL-6 and IL-8 cytokines induce a self- and cross-reinforced senescence/inflammatory milieu strengthening tumorigenic capabilities in the MCF-7 breast cancer cell line. Cell Commun Signal. 2017;15(1):17. doi:10.1186/s12964-017-0172-3.
- Owusu BY, Galemmo R, Janetka J, Klampfer L. Hepatocyte Growth Factor, a Key Tumor-Promoting Factor in the Tumor Microenvironment. Cancers (Basel). 2017;9(4). doi:10.3390/cancers9040035.
- Pan Y, Smithson LJ, Ma Y, Hambardzumyan D, Gutmann DH. Ccl5 establishes an autocrine high-grade glioma growth regulatory circuit critical for mesenchymal glioblastoma survival. Oncotarget. 2017;8(20):32977–32989.
- Park SA, Surh YJ. Modulation of tumor microenvironment by chemopreventive natural products. Ann N Y Acad Sci. 2017. doi: 10.1111/nyas.13395.
- Patrice JM. The Microenvironment and Drug Resistance. Cancer Drug Resistance.2006;(2):201–210.
- Pautu V, Leonetti D, Lepeltier E, Clere N, Passirani C. Nanomedicine as a potent strategy in melanoma tumor microenvironment. Pharmacol Res. 2017;pii: S1043-6618(17):30015–4.
- Pickup M, Novitskiy S, Moses HL. The roles of TGFβ in the tumour microenvironment. Nat Rev Cancer. 2013;13(11):788–799. doi:10.1038/nrc3603.
- Pickup MW, Mouw JK, Weaver VM. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 2014;15(12):1243–53. doi:10.15252/embr.201439246.
- Pike KA, Hui C, Krawczyk CM. Detecting Secreted Analytes from Immune Cells: An Overview of Technologies. Methods Mol Biol. 2016;1458:111–24. doi:10.1007/978-1-4939-3801-8_9.
- Kuninty Praneeth R, Schnittert Jonas, Storm Gert, and Prakash Jai. MicroRNA Targeting to Modulate Tumor Microenvironment. Front Oncol. 2016;6:3. doi:10.3389/fonc.2016.00003.
- Psyrri A, Kalogeras KT, Wirtz RM, Kouvatseas G, Karayannopoulou G, Goussia A. et al. Association of osteopontin with specific prognostic factors and survival in adjuvant breast cancer trials of the Hellenic Cooperative Oncology Group. J Transl Med. 2017;15(1):30. doi:10.1186/s12967-017-1134-7.
- Qin H, Ding Y, Mujeeb A, Zhao Y, Nie G. Tumor Microenvironment Targeting and Responsive Peptide-based Nano-formulations for Improved Tumor Therapy. Mol Pharmacol. 2017;92(3):219–231. doi:10.1124/mol.116.108084.
- Reina-Campos M, Moscat J, Diaz-Meco M. Metabolism shapes the tumor microenvironment. Curr Opin Cell Biol. 2017;48:47–53. doi:10.1016/j.ceb.2017.05.006.
- Richard G, Brown N, Rouan F, Van der Schroeff JG, Bijlsma E, Eichenfield LF, et al.. Genetic heterogeneity in erythrokeratodermia variabilis: novel mutations in the connexin gene GJB4 (Cx30.3) and genotype-phenotype correlations. J Invest Dermatol. 2003;120(4):601–609. doi:10.1046/j.1523-1747.2003.12080.x.
- Roberts DD. Emerging functions of matricellular proteins. Cell. Mol. Life Sci. 2011;68(19):3133–6. doi:10.1007/s00018-011-0779-2.
- Romano G, Veneziano D, Acunzo M, Croce CM. Small non-Coding RNA and Cancer. Carcinogenesis. 2017;38(5):485–491. doi:10.1093/carcin/bgx026.
- Rosner K, Kasprzak MF, Horenstein ACJ, Thurston HL, Abrams J, Kerwin LY, et al. Engineering a waste management enzyme to overcome cancer resistance to apoptosis: adding DNase1 to the anti-cancer toolbox. Cancer Gene Ther. 2011;18:346–57. doi:10.1038/cgt.2010.84.
- Salimian Rizi B, Caneba C, Nowicka A, Nabiyar AW, Liu X, Chen K2. Nitric oxide mediates metabolic coupling of omentum-derived adipose stroma to ovarian and endometrial cancer cells. Cancer Res. 2015;75(2):456–71. doi:10.1158/0008-5472.CAN-14-1337.
- Santangelo L, Battistelli C, Montaldo C, Citarella F, Strippoli R, Cicchini C. Functional Roles and Therapeutic Applications of Exosomes in Hepatocellular Carcinoma. Biomed Res Int. 2017;2017:2931813. doi:10.1155/2017/2931813.
- Seliger B. Immune modulatory microRNAs as a novel mechanism to revert immune escape of tumors. Cytokine Growth Factor Rev. 2017;36:49–56. doi:10.1016/j.cytogfr.2017.07.001.
- Sharma SH, Thulasingam S, Nagarajan S. Chemopreventive agents targeting tumor microenvironment. Life Sci. 2016;145:74–84. doi:10.1016/j.lfs.2015.12.016.
- Shintani Y, Fujiwara A, Kimura T, Kawamura T, Funaki S, Minami M, Okumura M. IL-6 Secreted from Cancer-Associated Fibroblasts Mediates Chemoresistance in NSCLC by Increasing Epithelial-Mesenchymal Transition Signaling. J Thorac Oncol. 2016;11(9):1482–92. doi:10.1016/j.jtho.2016.05.025.
- Simmons AJ, Lau KS. Deciphering tumor heterogeneity from FFPE tissues: Its promise and challenges. Mol Cell Oncol. 2016;4(1):e1260191. doi:10.1080/23723556.2016.1260191.
- Stadler S, Nguyen CH, Schachner H, Milovanovic D, Holzner S, Brenner S, Eichsteininger J. et al. Colon cancer cell-derived 12(S)-HETE induces the retraction of cancer-associated fibroblast via MLC2, RHO/ROCK and Ca2+ signalling. Cell Mol Life Sci. 2017;74(10):1907–1921. doi:10.1007/s00018-016-2441-5.
- Steinbichler TB, Dudás J, Riechelmann H, Skvortsova II. The Role of Exosomes in Cancer Metastasis. Semin Cancer Biol. 2017. doi: 10.1016/j.semcancer.2017.02.006.
- Sung BH, Weaver AM. Exosome secretion promotes chemotaxis of cancer cells. Cell Adh Migr. 2017;11(2):187–195. doi:10.1080/19336918.2016.1273307.
- Torres CM, Biran A, Burney MJ, Patel H, Henser-Brownhill T. et al. The linker histone H1.0 generates epigenetic and functional intratumor heterogeneity. Science. 2016;353(6307). doi:10.1126/science.aaf1644.
- Turner KM, Deshpande V, Beyter D, Koga T, Rusert J, Lee C. Extrachromosomal oncogene amplification drives tumour evolution and genetic heterogeneity. Nature. 2017;543(7643):122–125. doi:10.1038/nature21356.
- Tzeng HT, Tsai CH, Yen YT, Cheng HC, Chen YC, et al. Dysregulation of Rab37-Mediated Cross-talk between Cancer Cells and Endothelial Cells via Thrombospondin-1 Promotes Tumor Neovasculature and Metastasis. Clin Cancer Res. 2017;23(9):2335–2345. doi:10.1158/1078-0432.CCR-16-1520.
- Wang L, Yang D, Tian J, Gao A, Shen Y, Ren X, Li X, Jiang G, Dong T. Tumor necrosis factor receptor 2/AKT and ERK signaling pathways contribute to the switch from fibroblasts to CAFs by progranulin in microenvironment of colorectal cancer. Oncotarget. 2017;8(16):26323–26333.
- Wang Z, Chen JQ, Liu JL, Tian L. Exosomes in tumor microenvironment: novel transporters and biomarkers. J Transl Med. 2016; 19;14(1):297. doi:10.1186/s12967-016-1056-9.
- Wang Z, Tan Y, Yu W, Zheng S, Zhang S, Sun L, Ding K. Small role with big impact: miRNAs as communicators in the cross-talk between cancer-associated fibroblasts and cancer cells. Int J Biol Sci. 2017;13(3):339–348. doi:10.7150/ijbs.17680.
- Wasmer MH, Krebs P. The Role of IL-33-Dependent Inflammation in the Tumor Microenvironment. Front Immunol. 2017;7:682. doi:10.3389/fimmu.2016.00682.
- Weidle UH, Birzele F, Kollmorgen G, Rüger R. The Multiple Roles of Exosomes in Metastasis. Cancer Genomics Proteomics. 2017;14(1):1–15. doi:10.21873/cgp.20015.
- Whiteside TL. Exosomes carrying immunoinhibitory proteins and their role in cancer. Clin Exp Immunol. 2017;189(3):259–267. doi:10.1111/cei.12974.
- Wu D, Zhuo L, Wang X. Metabolic reprogramming of carcinoma-associated fibroblasts and its impact on metabolic heterogeneity of tumors. Semin Cell Dev Biol. 2017;64:125–131. doi:10.1016/j.semcdb.2016.11.003.
- Wu L, Zhang X, Zhang B, Shi H, Yuan X, Sun Y, Pan Z, Qian H, Xu W. Exosomes derived from gastric cancer cells activate NF-κB pathway in macrophages to promote cancer progression. Tumour Biol. 2016;37(9):12169–12180. doi:10.1007/s13277-016-5071-5.
- Wu X, Chen X, Zhou Q, Li P, Yu B, Li J, Qu Y, Yan J, Yu Y, Yan M, Zhu Z, Liu B, Su L. Hepatocyte growth factor activates tumor stromal fibroblasts to promote tumorigenesis in gastric cancer. Cancer Lett. 2013;335(1):128–35. doi:10.1016/j.canlet.2013.02.002.
- Wyatt LC, Lewis JS, Andreev OA, Reshetnyak YK, Engelman DM. Applications of pHLIP Technology for Cancer Imaging and Therapy. Trends Biotechnol. 2017;35(7):653–664. doi:10.1016/j.tibtech.2017.03.014.
- Yan J, Ma S, Zhang Y, Yin C, Zhou X, Zhang G. Potential role of microRNA-126 in the diagnosis of cancers: A systematic review and meta-analysis. Medicine (Baltimore). 2016;95(35):e4644. doi:10.1097/MD.0000000000004644.
- Yap TA, Gerlinger M, Futreal PA, Pusztai L, Swanton C. Intratumor heterogeneity: seeing the wood for the trees. Sci Transl Med. 2015;4(127):127ps10.
- Yuan Y. Spatial Heterogeneity in the Tumor Microenvironment. Cold Spring Harb Perspect Med. 2016;6(8): doi:10.1101/cshperspect.a026583.
- Zhang H, Mai Q, Chen J. MicroRNA-210 is increased and it is required for dedifferentiation of osteosarcoma cell line. Cell Biol Int. 2017;41(3):267–275. doi:10.1002/cbin.10721.
- Zhang M, Zhang L, Cui M, Ye W, Zhang P, Zhou S. Wang J miR-302b inhibits cancer-related inflammation by targeting ERBB4, IRF2 and CXCR4 in esophageal cancer. Oncotarget. 2017;8(30):49053–49063.
- Zou JM, Qin J, Li YC, Wang Y, Li D, Shu Y, Luo C, Wang SS, Chi G, Guo F, Zhang GM, Feng ZH. IL-35 induces N2 phenotype of neutrophils to promote tumor growth. Oncotarget. 2017;8(20):33501–33514.