
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Objective: To identify the significantly altered circRNAs and mRNAs in thyroid cancer, investigate their target miRNAs and determine their biological functions.
Methods: The differentially expressed circRNAs, mRNAs and pathways in thyroid cancer were identified by microarray analysis and gene set enrichment analysis (GSEA). The correlative circRNAs and mRNAs were found out through Pearson correlative analysis. The common target miRNAs of circNEK6 and FZD8 related to thyroid cancer was screened out through Targetscan, miRanda and HMDD analysis. The mRNA and protein expressions in thyroid cancer tissues and cells were detected by qRT-PCR and western blot. CircRNA was confirmed by the RNase R digestion and nucleic acid electrophoresis. The target relationships were verified by the dual luciferase reporter assay. Cell viability, invasion and apoptosis were determined by MTT assay, Transwell assay and flow cytometry, respectively.
Results: CircNEK6 and FZD8 were significantly up-regulated in thyroid cancer, with strong correlations. The Wnt signaling pathway was activated in thyroid cancer. MiR-370-3p was the common target miRNA of circNEK6 and FZD8, and it was down-regulated in thyroid cancer. Overexpression of circNEK6 and FZD8 could promote the growth and invasion of thyroid cancer cells, while up-regulation of miR-370-3p could suppress thyroid cancer progression and inhibit the Wnt signaling pathway. MiR-370-3p’s effect on thyroid cancer cells could be rescued by circNEK6 or FZD8.
Conclusion: CircNEK6 promoted the progression of thyroid cancer through up-regulating FZD8 and activating Wnt signaling pathway by targeting miR-370-3p.
Introduction
In recent decades, thyroid cancer has aroused more and more attentions globally because of its fastest increasing rate in incidence among all cancer types.Citation1 As the most common endocrine cancer, the mortality rate of thyroid cancer worldwide is approximately 0.2–0.6 per 100,000 people.Citation2 It has a disparity by sex as its incidence rate is 3 times in women higher than that in men.Citation3 Besides the traditional treatment strategies of surgery, radiotherapy and chemotherapy, researchers are now developing the promising novel molecular targeted therapies for the better treatment of thyroid cancer.Citation4 Therefore, it is of great importance to understand the mechanism of thyroid cancer and find potential targets.
Circular RNAs (circRNAs) are a novel class of long non-coding RNA which are produced co-transcriptionally by the spliceosome at the expense of canonical mRNA isoforms, forming a head-to-tail backsplice to make exons join in a non-linear order.Citation5 The dysregulation of circRNAs have been identified in many types of malignancies, which greatly affected the progression of human cancer. For example, circDOCK1 could suppress the apoptosis of oral squamous cell carcinoma cells through inhibiting the expression of miR-196a-5p.Citation6 Overexpression of circRNA ciRS-7 abrogated the inhibitory effect of miR-7 on gastric cancer.Citation7 The studies on thyroid cancer and circRNA are limited up to now. Peng et al. investigated the microarray profiles of circRNAs in papillary thyroid cancer and totally found 98 significantly dysregulated circRNAs, among which circRNA_100395 showed a noticeable interactive potential with cancer-related miRNAs.Citation8
The influences of circRNAs on thyroid cancer and their interactions with miRNA need to be more clearly elucidated. MiR-370 was reported to be a tumor suppressor in several kinds of malignancies, including hepatocellular carcinoma,Citation9 lung cancerCitation10and breast cancer.Citation11 Therefore, its potential suppressive effect on thyroid cancer was investigated in this study, as well as its association with the key circRNA.
The Wnt signaling pathway was involved in many phases of vertebrate embryonic development.Citation12 Its aberrant activation could facilitate the progression of various human cancers. For instance, the suppression of the Wnt signaling pathway by lncRNA PTCSC3 effectively inhibited the proliferation and invasion of glioma cells.Citation13 On the contrary, the esophageal cancer was accelerated by the activation of Wnt signaling led by the methylation of DACT2.Citation14 In thyroid cancer, it was deactivated by miR-126, a tumor suppressor that promoted the apoptosis and cell cycle arrest of thyroid cancer cells.Citation15 It was also reported to be able to regulate LGR5 in papillary thyroid cancer and promote cellular migration.Citation16 In short, many studies proved that the Wnt signaling pathway was positively related to the progression of human cancers.
Frizzled class receptor 8 (FZD8) was reported to be one of the cell surface receptor of the Wnt signaling pathway, which belongs to the Frizzled family of serpentine proteins.Citation17 The cancer stem-like cells which contributed to head and neck squamous carcinomas could be eliminated through the down-regulation of FZD8 and the Wnt signaling.Citation18 In colon cancer, FZD7 was also up-regulated together with the canonical Wnt signaling which supported colon epithelial tumor expansion.Citation19 Although FZD8’s performance in thyroid cancer has not been identified, its close relationship with the Wnt signaling pathway implied its potential important role in the thyroid cancer.
Therefore, in present study, we identified the significantly up-regulated circNEK6 and FZD8 in thyroid cancer through bioinformatics analysis and investigated their target relationships with miR-370-3p. CircNEK6 not only influenced the progression of thyroid cancer through regulating FZD8, but also activated the Wnt signaling pathway in thyroid cancer. Our discoveries revealed the importance role of circNEK6 in thyroid cancer and its interactions with FZD8 and miR-370-3p.
Methods and materials
Microarray analysis
The expression profiles of mRNAs and circRNAs of thyroid cancer tissues were downloaded from microarray GSE3678 (GPL570) and GSE93522 (GPL19978) respectively through the database of National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO). The screening conditions for differentially expressed mRNAs and circRNAs were |log2(FC)| > 1 and P.adjust < 0.05 (calculated by Benjamini and Hochberg False Discovery Rate method).
Gene set enrichment analysis
The normalized mRNA expression profiles of GSE3678 were uploaded to GSEA v3.0 software to conduct the enrichment analysis of KEGG pathways. The significantly altered pathways were visualized by dotplot and joyplot by using ggplot2, grid, devtools and easygplot2 packages. The genes involved in the Wnt signaling pathway were further analyzed by the GSEABase package.
Correlative analysis
The Pearson correlative analysis was employed to analyze the relationships between the differentially expressed mRNAs in the Wnt signaling pathway and the differentially expressed circRNAs of thyroid cancer tissues. The criteria of closely related mRNAs and circRNAs were Cutoff > 0.8 and P < 0.05. The relationship network of mRNAs and circRNAs was constructed by Cytoscape (version 3.5.1). In addition, the miRNAs that targeted FZD8 and circNEK6 were predicted by Targetscan (http://www.targetscan.org/vert_71/) and miRanda (http://www.microrna.org/microrna/home.do), respectively. The miRNAs that related to thyroid cancer were predicted by the Human MicroRNA Disease Database (HMDD, http://www.cuilab.cn/hmdd). As a result, the common miRNA that simultaneously targeted FZD8 and circNEK6 and related to thyroid cancer was screened out.
Tissues and cells
Forty pairs of thyroid cancer tissues and the corresponding adjacent normal tissues came from patients in the Third Affiliated Hospital of Kunming Medical University (Kunming, China). The informed consent was obtained from all patients. The Human Research Ethics Committee of the Third Affiliated Hospital of Kunming Medical University approved this study.
The thyroid cancer cell lines TPC-1 and FTC-133 together with the normal thyroid cell line Nthy-ori 3–1 were purchased from BeNa Culture Collection (Beijing, China). FTC-133 cells were maintained in the high glucose DMEM, while TPC-1 cells and Nthy-ori 3–1 cells were maintained in the RPMI and F-12K mediums respectively. All mediums were supplemented with 10% fetal bovine serum (FBS, Hyclone, Logan, UT, USA). Cells were cultured in humidity with CO2 at 37°C.
Rnase r digestion
Total RNA (5 μg) was incubated 15 min at 37°C with 3 U/μg of RNase R (Epicentre Biotechnologies, Madison, WI, USA). The RNase R digestion reaction was performed twice following previously published procedures.
Nucleic acid electrophoresis
The agarose gels with TE buffer (Thermo Scientific, Waltham, MA, USA) was used to detect the cDNA and genomic DNA (gDNA) PCR products of circNEK6. DL600 (KeyGen, Nanjing) was served as the DNA marker. The electrophoresis at 110 V was employed to separate the DNA for 30 min, and the results were measured by the UV irradiation.
Qrt-pcr
The total RNA of thyroid cancer cells was extracted by the TRIzol reagent (Life Technologies corporation, Gaithersburg, MD, USA). After the reverse transcription of RNA conducted by using PrimeScript II 1st Strand cDNA Synthesis Kit (TaKaRa, Tokyo, Japan), qRT-PCR was performed for the obtained cDNA through SYBR Premix Ex Taq (TaKaRa). U6 was the internal control for the determination of the relative expressions of miR-370-3p, while GAPDH was the internal control for the detection of circNEK6 and FZD8. The relative expression was calculated by the method. The primers were synthesized by Sangon Biotech (Shanghai, China) and listed in .
Table 1. Sequences for transfection.
Vector construction
To induce NEK6 transcript formation in vitro by nonlinear splicing, circNEK6 over-expression vector was constructed. The specially designed front and back circular frames were synthesized and added to pLO-circ vector for the circulation of transcripts. The front circular frame contained the endogenous flanking genomic sequence with Ndel restriction enzyme site, and the back circular frame contained part of the inverted upstream sequence with EcoRI site. The cDNA encoding circNEK6 in TPC-1 cells was amplified using primers 5ʹ-CATATGGAAGAAGACA-3ʹ and 5ʹ- GAATTCCTCGTACAGCAG-3ʹ. As a result, the sequence contained Ndel site, splice acceptor AG, circNEK6 sequence, splice donor GT and Ndel site. Then the amplified fragment was cloned to the vector between the two frames. Besides, we also established the mock vector only containing a nonsense stuffer between the two circular frames without the circNEK6 encoding cDNA. The result of vector construction was verified by direct sequencing. The vectors were constructed with the help of Guangzhou Geenseed Biotech Co (Guangzhou, China).
Cell transfection
MiR-370-3p mimics, miR-370-3p inhibitor, si-circNEK6, pLO-circNEK6, pcDNA3.1-FZD8, si-FZD8 and the empty vectors were obtained from GenePharma (Shanghai, China). In the first group, cells were divided into NC without treatment, circNEK6(+) transfected with pLO-circNEK6 and circNEK6(-) transfected with si-circNEK6. In the second group, cells were divided into NC without treatment, FZD8(+) transfected with pcDNA3.1-FZD8 and FZD8(-) transfected with si-FZD8. In the third group, cells were divided into NC without treatment, miR-370-3p(+) transfected with miR-370-3p mimics, miR-370-3p(-) transfected with miR-370-3p inhibitor, miR-370-3p(+) + FZDB(+) that were co-transfected with miR-370-3p mimics and pcDNA3.1-FZD8, and miR-370-3p(+) + circNEK6(+) that were co-transfected with miR-370-3p mimics and pLO-circNEK6. The sequences were listed in .
Table 2. Primer Sequences for qRT-PCR.
TPC-1 cells at logarithmic phase were digested by pancreatin and resuspended to prepare the cell suspension at 24 h before transfection. Then they were inoculated into 6-well plates at the density of 1 × 106 cells/well to incubate for 18–24 h to ensure the cell confluence of each well was 80–90% before transfection. The original medium was replaced with fresh medium without serum and antibiotics at 3 h before transfection. Cell transfection was conducted by using Lipofectamine 2000 (Life Technologies), and the transfected cells were cultured for 48 h before further experiments.
Western blot
The total protein of thyroid cancer cells was extracted and quantified by the Bradford protein assay (Bio-Rad, Hercules, CA, USA) before it was separated by the SDS-PAGE and transferred to the PVDF membrane through iBlot Dry Blotting Transfer System (Life Technologies). The membrane was then blocked in PBS buffer solution with 5% skimmed milk and 0.1% Tween-20 for 2 h. It was incubated by the primary antibodies at 4°C overnight, and then washed by PBS supplemented with 0.1% Tween thrice before being further incubated with the secondary antibody IgG-HRP (ab7090, 1:2000) for 1–2 h. After incubation, the washed PVDF membrane was added with DAB developing solution for exposure. Software ImageJ was employed to quantify the visualized protein bands. β-actin was the internal control. The primary antibodies used were anti-FZD8 (ab155650, 1:10000), anti-Cyclin D1 (ab134175, 1:10000), anti-c-myc (ab32072, 1:10000) and anti-β-actin (ab8227, 1:1000). The antibodies were all obtained from Abcam (Cambridge, MA, USA).
Dual luciferase reporter assay
The empty plasmid pmirGLO (Promega, Madison, WI, USA) and the recombinant plasmids pmirGLO-circNEK6-wt, pmirGLO-circNEK6-mut, pmirGLO-FZD8 -wt and pmirGLO-FZD8-mut from GenePharma were co-transfected with miR-NC or miR-370-3p mimics respectively into the 293T cells (BeNa Culture Collection) by using Lipofectamine 2000 (Life Technologies). After 48 h, the luciferase activity of 293T cells was detected by the Dual Luciferase Reporter Gene Detection System Kit (Promega) according to the instructions.
MTT assay
Cell proliferation was determined by the MTT Kit (Promega) in accordance with manufacturer’s instructions. In brief, single cell suspension at the density of 5 × 104 cells/ml was prepared by the culture medium with 10% FBS. The suspension was added into 96-well plates (200 μl/well) to incubate for 3–5 days. After transfection, the cells were further incubated for 24, 48 and 72 h before being added with 20 μl MTT (5 mg/ml). The supernatant was removed after 4 h incubation with MTT, and 200 μl DMSO was added into the plates to solubilize the formazan. The optical density at 490 nm was measured by the microplate reader, representing the cell viability.
Transwell assay
Fifty μl Matrigel (BD Bioscience, San Jose, CA, USA) diluted with serum free medium was added into the Transwell chamber and solidified at 37°C. After 2 h, 200 μl serum free medium was added into the chamber to hydrate the Matrigel. The transfected cells digested with pancreatin were added into the upper chamber of the Transwell (5 × 103 cells in 200 μl cell suspension). The lower chamber was filled with 500 μl culture medium with 10% FBS. After incubation for 24 h, the chambers were washed by PBS twice and fixed with 4% paraformaldehyde for 20 min. The cells in the lower chamber were stained with 0.1% crystal violet for 30 min. The Matrigel and cells in the upper chamber was removed by the cotton wool. The invasive cells in the lower chamber were photographed by the microscope and counted.
Cell apoptosis assay
Cells were collected and resuspended for the apoptosis detection 48 h after transfection. The apoptosis was determined following the instructions of the PE Annexin V Apoptosis Detection Kit (BD Bioscience). The flow cytometry FACS Calibur was applied to detect the cells apoptosis. The software FACS Diva was employed to analyze the data.
Statistical analysis
The data were expressed by mean ± standard deviation (x ± s). Comparisons between two groups were conducted by the paired t test. Comparisons among multiple groups were conducted by the One-Way ANOVA. The statistical analysis was performed the software GraphPad Prism 6.0.
Results
1. The differentially expressed mrnas, circrnas and signaling pathway in thyroid cancer
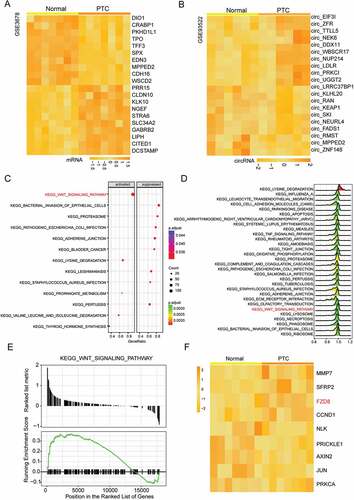
Ten most significantly up-regulated and down-regulated mRNAs in 7 pairs of thyroid cancer tissues and adjacent normal tissues (GSE3678) were displayed in . Meanwhile, through GSE93522, the differentially expressed circRNAs in 6 thyroid cancer tissues were screened out (). CircNEK6 (hsa_circ_0088483) was one of the most up-regulated circRNAs in thyroid cancer tissues (). According to the database circBase, circNEK6 is a kind of exonic circRNA, encoding the mRNA NEK6. The dotplot and joyplot of GSEA showed that the Wnt signaling pathway was activated in thyroid cancer (-). Its normalized enrichment score (NES) was higher than 0, and the majority of genes in this pathway were up-regulated (), illustrating that the Wnt signaling pathway was significantly activated in thyroid cancer. exhibited the differentially expressed mRNAs in the Wnt signaling pathway. FZD8, the upstream gene of the pathway, was up-regulated in thyroid cancer tissues ().
Figure 1. The differentially expressed mRNAs, circRNAs and signaling pathway in thyroid cancer. (a) The heat map of top 10 most up-regulated and down-regulated mRNAs in thyroid cancer tissues analyzed from microarray GSE3678. (b) The heat map of top 10 most up-regulated and down-regulated circRNAs in thyroid cancer tissues analyzed from microarray GSE93522. CircNEK6 was up-regulated in thyroid cancer. (c) The dotplot of significantly altered pathways in thyroid cancer tissues analyzed by GSEA. The Wnt signaling pathway was activated. The enrichment significance (the adjusted P value) was reflected by the color intensity of the nodes. The node size represented the gene count in the analyzed gene set. The gene ratio in the horizontal axis represented the proportion of differential genes in the gene set. (d) The joyplot of significantly altered pathways in thyroid cancer tissues analyzed by GSEA. The Wnt signaling pathway was activated. The enrichment significance (the adjusted P value) was reflected by the color intensity of the peaks. Pathways with ridges on the right side of 0 were up-regulated. (e) The GSEA enrichment plots of genes involved in the Wnt signaling pathway analyzed by GSEA. Most genes were up-regulated in thyroid cancer tissues as the ranked list metric and running enrichment score of them were larger than 0. (f) The heat map of differentially expressed mRNAs in the Wnt signaling pathway analyzed from microarray GSE3678. FZD8 was up-regulated in thyroid cancer tissues. The screening conditions of microarray analysis were |log2(FC)| > 1 and P.adjust < 0.05.
2. The close relationships between circnek6, FZD8 and mir-370-3p
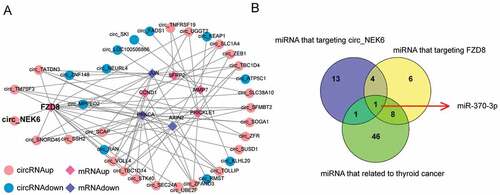
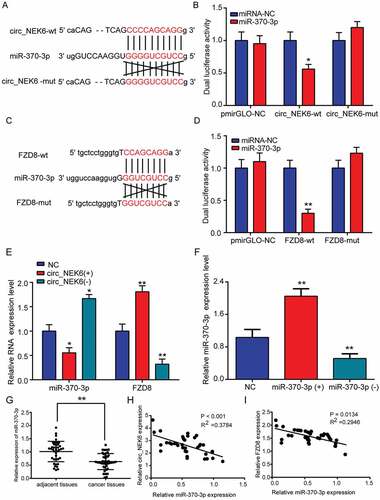
Through the network of differentially expressed circRNAs and differentially expressed mRNAs involved in the Wnt signaling pathway in thyroid cancer, the correlative circRNAs and mRNAs (Cutoff > 0.8 and P < 0.05) were displayed (). CircNEK6 was closely related to FZD8 (). In addition, miRNAs that targeted circNEK6 and FZD8 were predicted in . Among these miRNAs, miR-370-3p was the only thyroid cancer-related miRNA that simultaneously targeted circNEK6 and FZD8 (). Therefore, miR-370-3p was further investigated in the following study.
Figure 2. The close relationships between circNEK6, FZD8 and miR-370-3p. (a) The correlative network of differentially expressed circRNAs and differentially expressed mRNAs involved in the Wnt signaling pathway in thyroid cancer constructed by Cytoscape. CircNEK6 was related to FZD8. (b) The target miRNAs of circNEK6 and FZD8 predicted by miRanda and Targetscan, and the thyroid cancer-related miRNAs predicted by HMDD database. MiR-370-3p was the only common miRNA of them.
3. Circnek6 and FZD8 were up-regulated in thyroid cancer
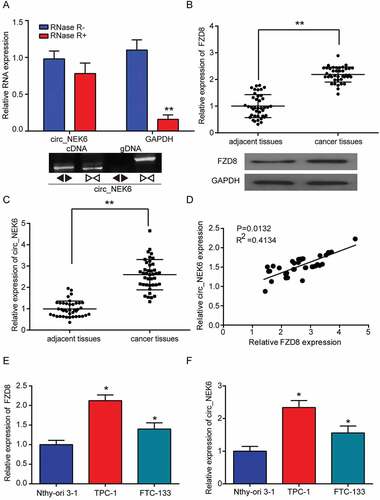
CircNEK6 was the circRNAs as its expression had no significant change when being treated by RNase R (). In addition, the cDNA of circNEK6 was amplified, while the gDNA did not produce amplification with divergent primers, confirming that circNEK6 was a circRNA (). In thyroid cancer tissues, the expressions of FZD8 and circNEK6 were distinctively elevated (-). CircNEK6 expression was positively correlated with that of FZD8 in thyroid cancer tissues (). Their expressions in thyroid cancer cells TPC-1 and FTC-133 were also remarkably higher than those in normal thyroid cell line Nthy-ori 3–1 (-). Since TPC-1 cell line exhibited a more obvious up-regulation of circNEK6 and FZD8 than FTC-133 cell line, it was used in the following in vitro experiments. The results confirmed that circNEK6 and FZD8 were up-regulated in thyroid cancer, and their expressions were positively correlated.
Figure 3. CircNEK6 and FZD8 were up-regulated in thyroid cancer. (a) The relative expression of circNEK6 had no significant change after RNase R digestion. **P < 0.01 compared with RNase R(-). The cDNA of circNEK6 was amplified with divergent primers, while the gDNA was not amplified. (b) The relative expression of FZD8 mRNA in 40 thyroid cancer tissues was significantly higher than that in paired adjacent tissues. The relative expression of FZD8 protein was elevated in cancer tissues. **P < 0.01 compared with adjacent tissues. (c) The relative expression of circNEK6 in 40 thyroid cancer tissues was significantly higher than that in adjacent tissues. **P < 0.01 compared with adjacent tissues. (d) The relative circNEK6 expression was positively correlated with the relative FZD8 expression. (e) The relative expressions of FZD8 mRNA in thyroid cancer cell lines (TPC-1, FTC-133) were significantly higher than that in normal thyroid cell line (Nthy-ori 3–1). *P < 0.05, **P < 0.01 compared with Nthy-ori 3–1. (f) The relative expressions of circNEK6 in thyroid cancer cell lines (TPC-1, FTC-133) were significantly higher than that in normal thyroid cell line (Nthy-ori 3–1). *P < 0.05, **P < 0.01 compared with Nthy-ori 3–1.
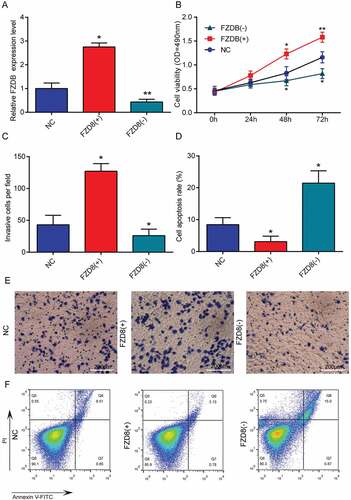
4. Circnek6 and FZD8 promoted the growth and invasion of thyroid cancer cells
CircNEK6 was overexpressed (circNEK6(+)) and silenced (circNEK6(-)) in TPC-1 cells to investigate its biological function in thyroid cancer (). The cell viability and invasion of TPC-1 cells were improved by circNEK6 overexpression but suppressed by circNEK6 down-regulation (-). In contrast, the apoptosis of TPC-1 cells was induced by the silencing of circNEK6, but restrained by circNEK6 overexpression (, ). The FZD8 had a similar effect as circNEK6 on TPC-1 cells. FZD8 was up-regulated by FZD8(+) and knocked down by FZD8(-) (). FZD8 overexpression successfully enhanced the proliferation and invasion of thyroid cancer cells, as well as reduced the apoptosis (). On the other hand, FZD8 knockdown exerted an opposite effect which suppressed the growth and invasion of TPC-1 cells (-. In consequence, both circNEK6 and FZD8 could promote the growth and invasion of thyroid cancer cells.
Figure 4. CircNEK6 promoted the growth and invasion of thyroid cancer cells. (a) The relative circNEK6 expression level was increased in circNEK6(+) group but decreased in circNEK6(-) group. (b) The cell viability detected by MTT assay was increased in circNEK6(+) group but decreased in circNEK6(-) group. (c, e) The number of invasive TPC-1 cells per field investigated by Transwell assay was increased in circNEK6(+) group but decreased in circNEK6(-) group. (d, f) The cell apoptosis rate of TPC-1 cells detected by flow cytometry was decreased in circNEK6(+) group but increased in circNEK6(-) group. *P < 0.05, **P < 0.01 compared with NC.
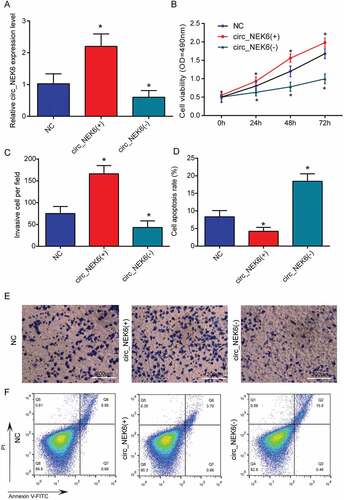
Figure 5. FZD8 promoted the growth and invasion of thyroid cancer cells. (a) The relative FZD8 expression level was increased in FZD8(+) group but decreased in FZD8(-) group. (b) The cell viability detected by MTT assay was increased in FZD8(+) group but decreased in FZD8(-) group. (C, E) The number of invasive TPC-1 cells per field investigated by Transwell assay was increased in FZD8(+) group but decreased in FZD8(-) group. (d, f) The cell apoptosis rate of TPC-1 cells detected by flow cytometry was decreased in FZD8(+) group but increased in FZD8(-) group. *P < 0.05, **P < 0.01 compared with NC.
5. The target relationships of circnek6-mir-370-3p and mir-370-3p-fzd8
MiR-370-3p was predicted to be the downstream target miRNA of circNEK6 (), which was verified by the reduced dual luciferase activity of cells co-transfected with circNEK6-wt and miR-30-3p (). Similarly, FZD8 was predicted as the target gene of miR-370-3p, which could bind miR-370-3p with the 3ʹUTR region (). The dual luciferase activity of FZD8-wt in miR-370-3p group was decreased while that of FZD8-mut showed insignificant variation, confirming the target relationship between miR-370-3p and FZD8 (). After circNEK6 overexpression, the miR-370-3p expression was decreased while that of FZD8 was increased, suggesting that circNEK6 could up-regulate FZD8 through targeting miR-370-3p (). The relative miR-370-3p expression in TPC-1 cells was increased by transfection of miR-370-3p mimics (miR-370-3p(+)) but decreased by the transfection of miR-370-3p inhibitor (miR-370-3p(-)) (). In thyroid tissues, the miR-370-3p expression was significantly lower than that in adjacent tissues (). The expression of miR-370-3p had a negative correlation with both circNEK6 and FZD8 expressions (-), verifying miR-370-3p’s target relationship with circNEK6 and FZD8.
Figure 6. The target relationships of circNEK6-miR-370-3p and miR-370-3p-FZD8. (a) The target relationship between circNEK6 and miR-370-3p predicted by Targetscan. (b) The dual luciferase activity of cells co-transfected with circNEK6-wt and miR-370-3p was decreased, while that of cells co-transfected with circNEK6-mut and miR-370-3p had no significant change. *P < 0.05 compared with miRNA-NC. (c) The target relationship between FZD8 and miR-370-3p predicted by Targetscan. (d) The dual luciferase activity of cells co-transfected with FZD8-wt and miR-370-3p was decreased, while that of cells co-transfected with FZD8-mut and miR-370-3p had no significant change. **P < 0.01 compared with miRNA-NC. (e) The relative expression of miR-370-3p was decreased in circNEK6(+) group but increased in circNEK6(-) group. The relative mRNA expression of FZD8 was increased in circNEK6(+) group but decreased in circNEK6(-) group. *P < 0.5, **P < 0.01 compared with NC. (f) The relative miR-370-3p expression was increased in miR-370-3p(+) group but decreased in miR-370-3p(-) group. **P < 0.01 compared with NC. (g) The relative expression of miR-370-3p in 40 thyroid cancer tissues was significantly lower than that in adjacent tissues. **P < 0.01 compared with adjacent tissues. (h-i) The relative expressions of circNEK6 and FZD8 were negatively correlated with the expression of miR-370-3p in thyroid cancer tissues.
6. MiR-370-3p inhibited the growth and invasion of thyroid cancer cells while miR-370-3p + FZD8 rescued the inhibitory effect
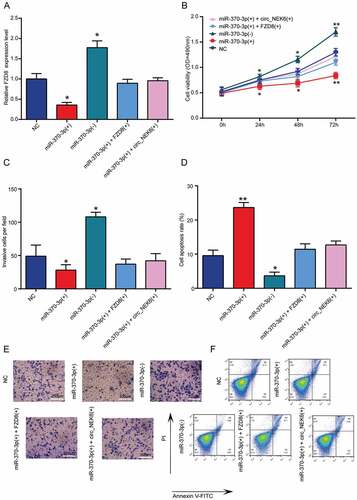
The overexpression of miR-370-3p could elevate the FZD8 expression in thyroid cancer cells, while silencing miR-370-3p could reduce the mRNA expression of FZD8 (). However, co-transfection of miR-370-3p and FZD8 or miR-370-3p and circNEK6 in TPC-1 cells had negligible effect on FZD8 expression (). The cell viability and invasion of TPC-1 cells was decreased by miR-370-3p overexpression, increased by miR-370-3p down-regulation and returned to the similar level as NC by miR-370-3p(+) + FZD8(+) or miR-370-3p(+) + circNEK6(+) (-, es). On the contrary, the apoptosis of TPC-1 cells was induced by miR-370-3p overexpression but suppressed by miR-370-3p down-regulation (, ). MiR-370-3p’s effect on cell apoptosis was also rescued by FZD8 or circNEK6 (, ). These results indicated that miR-370-3p could restrain the deterioration of thyroid cancer.
Figure 7. MiR-370-3p inhibited the growth and invasion of thyroid cancer cells while miR-370-3p + FZD8 rescued the inhibitory effect. (a) The relative FZD8 expression in TPC-1 cells was decreased in miR-370-3p(+) group, increased in miR-370-3p(-) group and rescued in miR-370-3p(+) + FZD8(+) and miR-370-3p(+) + circNEK6(+) groups. (b) The cell viability of TPC-1 cells detected by MTT assay was decreased in miR-370-3p(+) group, increased in miR-370-3p(-) group and rescued in miR-370-3p(+) + FZD8(+) and miR-370-3p(+) + circNEK6(+) groups. (c, e) The number of invasive TPC-1 cells in per field detected by Transwell assay was decreased in miR-370-3p(+) group, increased in miR-370-3p(-) group and rescued in miR-370-3p(+) + FZD8(+) and miR-370-3p(+) + circNEK6(+) groups. (d, f) The apoptosis rate of TPC-1 cells detected by flow cytometry was increased in miR-370-3p(+) group, decreased in miR-370-3p(-) group and rescued in miR-370-3p(+) + FZD8(+) and miR-370-3p(+) + circNEK6(+) groups. *P < 0.05, **P < 0.01 compared with NC.
7. Circnek6 targeted mir-370-3p to up-regulate FZD8 and activate the wnt signaling pathway
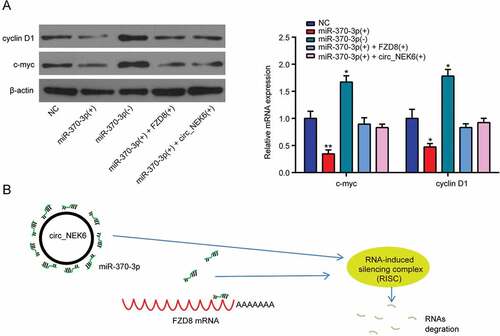
The proteins related to the Wnt signaling pathway, cyclin D1 and c-myc were down-regulated in TPC-1 cells after miR-370-3p overexpression, but up-regulated after miR-370-3p down-regulation (). The promotive effect of miR-370-3p overexpression on the protein expressions could be rescued by simultaneously overexpressing FZD8 or circNEK6 (). Therefore, the Wnt signaling pathway in thyroid cancer could be activated by the up-regulation of circNEK6 which resulted in the down-regulation of miR-370-3p and the overexpression of FZD8. The RNA-induced silencing complex (RISC) led by miR-370-3p could degrade the mRNA expression of FZD8, while circNEK6 bound miR-370-3p to reduce the FZD8 degration (). CircNEK6 up-regulated FZD8 and the Wnt signaling pathway in thyroid cancer through targeting miR-370-3p, thus influenced the progression of thyroid cancer.
Figure 8. CircNEK6 targeted miR-370-3p to up-regulate FZD8 and activate the Wnt signaling pathway. (a) The relative protein expressions of cyclin D1 and c-myc were decreased in miR-370-3p(+) group, increased in miR-370-3p(-) group and rescued in miR-370-3p(+) + FZD8(+) and miR-370-3p(+) + circNEK6(+) groups. *P < 0.5, **P < 0.01 compared with NC. (b) The mechanism diagram illustrated that the RNA-induced silencing complex (RISC) of miR-370-3p that degrade the FZD8 mRNA would be broke by the circNEK6 which bound miR-370-3p.
Conclusion
In this study, circNEK6 was identified to be up-regulated in thyroid cancer, together with the Wnt signaling pathway. FZD8 was the gene involved in the Wnt signaling pathway, with a distinctive elevated expression in thyroid tissues and cells. Both circNEK6 and FZD8 promoted the proliferation and invasion of thyroid cancer cells, whereas suppressed their apoptosis. As the common target miRNA of circNEK6 and FZD8, miR-370-3p restrained the progression of thyroid cancer in vitro. CircNEK6 targeted miR-370-3p to up-regulate FZD8 and activate the Wnt signaling pathway in thyroid cancer.
Microarray and gene set enrichment analyses were applied to screen out the differentially expressed circRNAs, mRNAs and signaling pathways in thyroid cancer. They were effective means for identifying key genes and pathways in human cancers. For instance, Ma et al. revealed that genes associated with the p53 signaling pathway and the JAK/STAT signaling pathway were remarkable enriched in papillary thyroid tumors with LINC00271 down-regulation through GSEA.Citation20 GSEA also clearly showed that OxPhos and MRP genes sets were distinctively down-regulated in thyroid cancer tissues.Citation21 Through bioinformatics analysis, Lee et al. identified that the mitochondrial calcium transporter LETM1 was associated cellular growth signals such as the insulin signaling pathway, and it was up-regulated in aggressive thyroid cancer.Citation22 Detailed studies about important circRNAs in thyroid cancer are limited. Here we adopted the bioinformatics method to analyze the expression profiles of thyroid cancer tissue, successfully screening out circNEK6 as an important circRNA for the first time.
The strong association between circNEK6 and FZD8 was found as they both targeted miR-370-3p. The axis circNEK6/miR-370-3p/FZD8 profoundly influenced the development of thyroid cancer. CircNEK6 has not been well studied before, so that its positive effect on the growth and metastasis of thyroid cancer was revealed for the first time. MiR-370 was a widely accepted tumor suppressor in various human cancers, although its influence in thyroid cancer has not aroused attention before. The meta-analysis of miRNA expression profiles revealed that miR-370 was down-regulated in recurrent prostate cancer.Citation23 It was among the tumor suppressor miRNAs that were frequently methylated in cholangiocarcinoma.Citation24 The cell apoptosis in hepatocellular carcinoma was induced by miR-370,Citation25 whereas the proliferation of ovarian cancer cells was suppressed by it.Citation26 Our findings also confirmed that miR-370-3p was down-regulated in thyroid cancer and its overexpression could attenuate the growth and invasion of cancer cells, which were in line with other studies. FZD8 was reported to be involved in several types of cancers, such as head and neck squamous carcinomas,Citation18 lung cancerCitation27 and breast cancer,Citation28 to facilitate the progression of them. In thyroid cancer, it was up-regulated and exerted similar promotive effects, suggesting that it was an oncogene.
Accumulated evidence indicated that circRNAs participated the progression of human cancers through the interaction with miRNAs and their target genes. For example, circ_0067934 enhanced the cancer cell viability and metastasis of hepatocellular carcinoma through up-regulating FZD5, which was the target gene of miR-1324.Citation29 CircDOCK1 suppressed the cell apoptosis of oral squamous cell carcinoma via the inhibition of miR-196a-5p, which targeted the tumor suppressor BIRC3.Citation6 Circ_0012673 functioned as a sponge of miR-22, thus accelerating the deterioration of lung adenocarcinoma.Citation30 Similarly, since miR-370-3p had a target relationship with both circNEK6 and FZD8, circNEK6 could indirectly elevate the expression of FZD8 by targeting miR-370-3p. CircNEK6 acted as a competing endogenous RNA (ceRNA) that sponged miR-370-3p in thyroid cancer, leading to the up-regulation of FZD8.
FZD8 was a key gene in the Wnt signaling pathway, which was activated in several types of cancers. In prostate cancer, FZD8 was robustly up-regulated and promoted the bone-metastasis by activating the Wnt signaling pathway.Citation31 In thyroid cancer, lncRNA PTCSC3 and miR-574-5p inhibited the activity of the Wnt signaling pathway to suppress the cancer cell proliferation and migration.Citation32 The main oncogenic driver in thyroid cancer, RAS, was positively related to the activation of the Wnt signaling pathway.Citation33 Therefore, it could be speculated that the activation of the Wnt signaling pathway promoted the development of thyroid cancer, and the up-regulation of FZD8 mediated by the circNEK6/miR-370-3p/FZD8 axis led to the Wnt signaling pathway activation.
The present study had some limitations should be improved in future. For example, the mechanism of how the activated Wnt signaling pathway affected thyroid cancer was not fully elucidated. In addition, in vivo experiments should be supplemented to confirm our results. The association between circNEK6/miR-370-3p/FZD8 and clinical prognosis of thyroid cancer is worth to be explored to evacuate the potential biomarkers.
In conclusion, the significant up-regulation of circNEK6 in thyroid cancer elevated the expression of FZD8 through targeting miR-370-3p and activated the Wnt signaling pathway. CircNEK6 up-regulation promoted the suppressed cancer cell apoptosis but promoted the proliferation and invasion of thyroid cancer cells.
Additional information
Funding
Related Research Data
References
- Park JY, Yi JW, Park CH, Lim Y, Lee KH, Lee KE, Kim JH. Role of BRAF and RAS mutations in extrathyroidal extension in papillary thyroid cancer. Cancer Genomics Proteomics. 2016;13:171–181.
- La Vecchia C, Malvezzi M, C Bosetti, Garavello W, Bertuccio P, Levi F, Negri E. Thyroid cancer mortality and incidence: a global overview. Int J Cancer. 2015;136:2187–2195. doi:10.1002/ijc.29251.
- Siegel RL, Miller, KD, Jemal, A. Cancer Statistics, 2017. CA Cancer J Clin. 2017;67:7–30. doi:10.3322/caac.21387.
- Molinaro E, Romei, C, Biagini, A, Sabini, E, Agate, L, Mazzeo, S, Materazzi, G, Sellari-Franceschini, S, Ribechini, A, Torregrossa, L, et al. . Anaplastic thyroid carcinoma: from clinicopathology to genetics and advanced therapies. Nat Rev Endocrinol. 2017;13:644–660. doi:10.1038/nrendo.2017.76.
- Ashwal-Fluss R, Meyer M, Pamudurti NR, Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N, Kadener S. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell.2014; 56:55–66. doi:10.1016/j.molcel.2014.08.019.
- Wang L, Wei Y, Yan Y, Wang H, Yang J, Zheng Z, Zha J, Bo P, Tang Y, Guo X, et al. CircDOCK1 suppresses cell apoptosis via inhibition of miR196a5p by targeting BIRC3 in OSCC. Oncol Rep. 2018;39:951–966. doi:10.3892/or.2017.6174.
- Pan H, Li T, Jiang Y, Pan C, Ding Y, Huang Z, Yu H, Kong D. Overexpression of circular RNA ciRS-7 abrogates the tumor suppressive effect of miR-7 on gastric cancer via PTEN/PI3K/AKT signaling pathway. J Cell Biochem. 2018;119:440–446. doi:10.1002/jcb.26201.
- Peng N, Shi L, Zhang Q, Hu Y, Wang N, Ye H. Microarray profiling of circular RNAs in human papillary thyroid carcinoma. PLoS One. 2017;12:e0170287. doi:10.1371/journal.pone.0170287.
- Pan XP, Wang HX, Tong DM, Li Y, Huang LH, Wang C. miRNA-370 acts as a tumor suppressor via the downregulation of PIM1 in hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 2017;21:1254–1263.
- Li C, Ge Q, Liu J, Zhang Q, Wang C, Cui K, Chen Z. Effects of miR-1236-3p and miR-370-5p on activation of p21 in various tumors and its inhibition on the growth of lung cancer cells. Tumour Biol. 2017;39:1010428317710824. doi:10.1177/1010428317710824.
- Lv J, Xia K, Xu P, Sun E, Ma J, Gao S, Zhou Q, Zhang M, Wang F, Chen F, et al. miRNA expression patterns in chemoresistant breast cancer tissues. Biomed Pharmacother. 2014;68:935–942. doi:10.1016/j.biopha.2014.09.011.
- Gillhouse M, Wagner Nyholm M, Hikasa H, Sokol SY, Grinblat Y. Two Frodo/Dapper homologs are expressed in the developing brain and mesoderm of zebrafish. Dev Dyn. 2004;230:403–409. doi:10.1002/dvdy.20060.
- Xia S, Ji R, Zhan W. Long noncoding RNA papillary thyroid carcinoma susceptibility candidate 3 (PTCSC3) inhibits proliferation and invasion of glioma cells by suppressing the Wnt/beta-catenin signaling pathway. BMC Neurol. 2017;17:30. doi:10.1186/s12883-017-0813-6.
- Zhang M, Linghu E, Zhan Q, He T, Cao B, Brock MV, Herman JG, Xiang R, Guo M. Methylation of DACT2 accelerates esophageal cancer development by activating Wnt signaling. Oncotarget. 2016;7:17957–17969. doi:10.18632/oncotarget.7647.
- Wen Q, Zhao J, Bai L, Wang T, Zhang H, Ma Q. miR-126 inhibits papillary thyroid carcinoma growth by targeting LRP6. Oncol Rep. 2015;34:2202–2210. doi:10.3892/or.2015.4165.
- Michelotti G, Jiang X, Sosa JA, Diehl AM, Henderson BB. LGR5 is associated with tumor aggressiveness in papillary thyroid cancer. Oncotarget. 2015;6:34549–34560. doi:10.18632/oncotarget.5330.
- MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17:9–26. doi:10.1016/j.devcel.2009.06.016.
- Sun S, Liu S, Duan SZ, Zhang L, Zhou H, Hu Y, Zhou X, Shi C, Zhou R, Zhang Z. Targeting the c-Met/FZD8 signaling axis eliminates patient-derived cancer stem-like cells in head and neck squamous carcinomas. Cancer Res. 2014;74:7546–7559. doi:10.1158/0008-5472.CAN-14-0826.
- Chang B, Tessneer KL, McManus J, Liu X, Hahn S, Pasula S, Wu H, Song H, Chen Y, Cai X, et al. Epsin is required for Dishevelled stability and Wnt signalling activation in colon cancer development. Nat Commun. 2015;6:6380. doi:10.1038/ncomms7380.
- Ma B, Liao T, Wen D, Dong C, Zhou L, Yang S, Wang Y, Ji Q. Long intergenic non-coding RNA 271 is predictive of a poorer prognosis of papillary thyroid cancer. Sci Rep. 2016;6:36973. doi:10.1038/srep36973.
- Lee J, Seol MY, Jeong S, Lee CR, Ku CR, Kang SW, Jeong JJ, Shin DY, Nam KH, Lee EJ, et al. A metabolic phenotype based on mitochondrial ribosomal protein expression as a predictor of lymph node metastasis in papillary thyroid carcinoma. Medicine (Baltimore). 2015;94:e380. doi:10.1097/MD.0000000000000874.
- Lee J, Lee WK, Seol MY, Lee SG, Kim D, Kim H, Park J, Jung SG, Chung WY, Lee EJ, et al. Coupling of LETM1 up-regulation with oxidative phosphorylation and platelet-derived growth factor receptor signaling via YAP1 transactivation. Oncotarget. 2016;7:66728–66739. doi:10.18632/oncotarget.11456.
- Pashaei E, Pashaei E, Ahmady M, Ozen M, Aydin N. Meta-analysis of miRNA expression profiles for prostate cancer recurrence following radical prostatectomy. PLoS One. 2017;12:e0179543. doi:10.1371/journal.pone.0179543.
- Nakaoka, T, Saito, Y, Saito, H. Aberrant DNA methylation as a biomarker and a therapeutic target of cholangiocarcinoma. Int J Mol Sci.2017; 18.
- Pan X, Wang H, Tong D, Wang C, Sun L, Zhao C, Li Y, Zhu L, Wu D. Physcion induces apoptosis in hepatocellular carcinoma by modulating miR-370. Am J Cancer Res. 2016;6:2919–2931.
- Chen XP, Chen YG, Lan JY, Shen ZJ. MicroRNA-370 suppresses proliferation and promotes endometrioid ovarian cancer chemosensitivity to cDDP by negatively regulating ENG. Cancer Lett. 2014;353:201–210. doi:10.1016/j.canlet.2014.07.026.
- Wang HQ, Xu ML, Ma J, Zhang Y, Xie CH. Frizzled-8 as a putative therapeutic target in human lung cancer. Biochem Biophys Res Commun. 2012;417:62–66. doi:10.1016/j.bbrc.2011.11.055.
- Jiang Q, He M, Guan S, Ma M, Wu H, Yu Z, Jiang L, Wang Y, Zong X, Jin F, et al. MicroRNA-100 suppresses the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting Wnt/beta-catenin signaling pathway. Tumour Biol. 2016;37:5001–5011. doi:10.1007/s13277-015-4342-x.
- Zhu Q, Lu G, Luo Z, Gui F, Wu J, Zhang D, Ni Y. CircRNA circ_0067934 promotes tumor growth and metastasis in hepatocellular carcinoma through regulation of miR-1324/FZD5/Wnt/beta-catenin axis. Biochem Biophys Res Commun. 2018;497:626–632. doi:10.1016/j.bbrc.2018.02.119.
- Wang X, Zhu X, Zhang H, Wei S, Chen Y, Chen Y, Wang F, Fan X, Han S, Wu G. Increased circular RNA hsa_circ_0012673 acts as a sponge of miR-22 to promote lung adenocarcinoma proliferation. Biochem Biophys Res Commun. 2018;496:1069–1075. doi:10.1016/j.bbrc.2018.01.126.
- Li Q, Ye L, Zhang X, Wang M, Lin C, Huang S, Guo W, Lai Y, Du H, Li J, et al. FZD8, a target of p53, promotes bone metastasis in prostate cancer by activating canonical Wnt/beta-catenin signaling. Cancer Lett. 2017;402:166–176. doi:10.1016/j.canlet.2017.05.029.
- Wang X, Lu X, Geng Z, Yang G, Shi Y. LncRNA PTCSC3/miR-574-5p governs cell proliferation and migration of papillary thyroid carcinoma via Wnt/beta-Catenin signaling. J Cell Biochem. 2017;118:4745–4752. doi:10.1002/jcb.26142.
- Sastre-Perona A, Riesco-Eizaguirre G, Zaballos MA, Santisteban P. β-catenin signaling is required for RAS-driven thyroid cancer through PI3K activation. Oncotarget. 2016;7:49435–49449. doi:10.18632/oncotarget.10356.