
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
SARS-CoV-2 and other microbes within aerosol particles can be partially shielded from UV radiation. The particles refract and absorb light, and thereby reduce the UV intensity at various locations within the particle. Previously, we demonstrated shielding in calculations of UV intensities within spherical approximations of SARS-CoV-2 virions within spherical particles approximating dried-to-equilibrium respiratory fluids. The purpose of this paper is to extend that work to survival fractions of virions (i.e., fractions of virions that can infect cells) within spherical particles approximating dried respiratory fluids, and to investigate the implications of these calculations for using UV light for disinfection. The particles may be on a surface or in air. Here, the survival fraction (S) of a set of individual virions illuminated with a UV fluence (F, in J/m2) is assumed described by S(kF) = exp(-kF), where k is the UV inactivation rate constant (m2/J). The average survival fraction (Sp) of the simulated virions in a group of particles is calculated using the energy absorbed by each virion in the particles. The results show that virions within particles of dried respiratory fluids can have larger Sp than do individual virions. For individual virions, and virions within 1-, 5-, and 9-µm particles illuminated (normal incidence) on a surface with 260-nm UV light, the Sp = 0.00005, 0.0155, 0.22, and 0.28, respectively, when kF = 10. The Sp decrease to <10−7, <10−7, 0.077, and 0.15, respectively, for kF = 100. Results also show that illuminating particles with UV beams from widely separated directions can strongly reduce the Sp. These results suggest that the size distributions and optical properties of the dried particles of virion-containing respiratory fluids are likely important to effectively designing and using UV germicidal irradiation systems for microbes in particles. The results suggest the use of reflective surfaces to increase the angles of illumination and decrease the Sp. The results suggest the need for measurements of the Sp of SARS-CoV-2 in particles having compositions and sizes relevant to the modes of disease transmission.
Introduction
Some diseases caused by viruses and bacteria can be transmitted by sneezed or coughed aerosols, droplets (of any size), or by particles remaining after droplets of respiratory or other fluids have dried-to-equilibrium (termed “dried particles” here) (Huang et al. Citation2021; Lu et al. Citation2020; Miller et al. Citation2020). SARS-CoV-2 (Severe Acute Respiratory Syndrome coronavirus 2), the causative agent of COVID-19 (coronavirus disease 2019), is an enveloped, positive-sense, and single-stranded RNA virus, as are other viruses in the coronavirus subfamily (SARS, MERS, and types causing the common cold). SARS-CoV-2 is found in fluids of the nasopharynx (Landry et al. Citation2020; Yilmaz et al. Citation2021), nose (Péré et al. Citation2020), throat (Zou et al. Citation2020), and lung (Wolfel et al. Citation2020). It occurs in saliva (Azzi et al. Citation2020; Wyllie et al. Citation2020) and sputum (Pan et al. Citation2020).
Respiratory fluids may be aerosolized by coughing, sneezing, talking or breathing. Microbes in particles on surfaces may be transferred to humans by direct contact, via fomites, or may be reaerosolized (Fisher et al. Citation2012; Kesavan et al. Citation2017; Krauter and Biermann Citation2007; Paton et al. Citation2015; Qian et al. Citation2014). An approach to reducing transmission of diseases associated with microbes in the respiratory tract is to reduce the exposure of persons to viable microbes in the air (e.g., Cheng et al. (Citation2020) for SARS-CoV-2), and/or on surfaces/fomites (Boon and Gerba Citation2007; Rutala and Weber Citation2019). Face mask mandates and usage lead to lower rates of COVID-19 cases and deaths (Guy et al. Citation2021; Joo et al. Citation2021; Mitze et al. Citation2020).
There is a need for improved designs of buildings and vehicles, and the airflows within them, to reduce the transmission of airborne diseases while allowing human interactions to the extent possible (Augenbraun et al. Citation2020; Morawska et al. Citation2020). There is a need for decontamination of face masks, other personal protective equipment, filters (Fischer et al. Citation2020; Woo et al. Citation2012), and surfaces in many locations.
UV germicidal irradiation (UVGI), i.e., UV light at wavelengths from 200- to 320-nm, has been used for over 100 years to inactivate microbes. Low-pressure mercury lamps with a dominant emission peak at 254 nm have been the most common UVGI source. The use of UVGI for air and surface decontamination (including sources, biophysics, and inactivation systems) has been described in detail (Kowalski Citation2009). Improvements in UVGI sources and application techniques continue to be described as do studies of effectiveness (Anderson et al. Citation2018; Lindsley et al. Citation2018; Morawska et al. Citation2020; Rutala and Weber Citation2019).
A problem with using UVGI to inactivate microbes is that in some cases viruses and bacteria can be partially shielded from UVGI by clumping of microbes (e.g., Coohill and Sagripanti Citation2008; Kesavan et al. Citation2014; Handler and Edmonds Citation2015) or being on or within particles (Doughty et al. Citation2021; Osman et al. Citation2008). This problem is important, in part because inactivation rates obtained with measurements of individual, separated microbes may be higher than rates of inactivation of microbes within clumps or particles. A UV inactivation system may be designed for, and tested with, individual microbes, but then in actual use the microbes may occur within larger particles. In such a case, users and designers may have a false sense of security. Also, system designers may wonder how the problem of shielding from UV could be reduced or circumvented. Removing particles using filters or inactivating particles by increasing the UV intensity are possible solutions. But, questions remain about other possible solutions and tradeoffs.
Modeling the UVGI within microbes in particles can help in understanding shielding quantitatively, and in designing systems which mitigate the effects of shielding from UVGI and thereby achieve a desired level of inactivation. A key objective of modeling is to predict wavelength-dependent UV intensity within particles/droplets in a way that is (1) based on fundamental physical first principles, (2) approximates the detailed geometry of multiple particle systems and variable illumination directions, and yet is (3) computationally tractable. The fundamental physical principles in this case are represented by Maxwell's wave equations which here (with non-magnetic media and no free charges) reduce to the 3D vector partial differential equation (Bohren and Huffman Citation1983):
(1)
(1)
where ∇ × is the curl operator, E is the complex electric field vector for time-harmonic fields, λ is the wavelength in free space (for UVGI applications, typically close enough to the wavelength in air), and m = mr + i mi is the complex refractive index of the material, where mr (or mi) is referred to as the real (or imaginary) part of the complex refractive index. (In another common notation, n is used for mr, and k is used for mi, but here k is used for the UV inactivation rate constant.) Note that m is a material property and thus is position dependent. Quantities of interest, such as fluence and absorption rate, are typically obtained from the squared magnitude of E.
In general, it is possible to solve EquationEquation (1)(1)
(1) , for a given distribution of m and exciting electric field(s) (typically monochromatic plane wave(s)), using purely numerical means (e.g., a discrete dipole method or finite-difference time-domain method). Such approaches, applied to the systems of interest here (which, overall, are on the order of 4–100 wavelengths in diameter), would be computationally demanding.
Methods based upon analytical (as opposed to numerical) solutions to EquationEquation (1)(1)
(1) can be less computationally intensive. For examples relevant to modeling UV in microbes in particles, analytical solutions are possible when m is piecewise-constant, and when the boundaries separating one value of m from another are in the form of (1) spherical surfaces and/or (2) infinite plane surfaces. For such conditions E can be represented exactly in the piecewise-constant regions by fundamental basis functions (which are solutions to EquationEquation [1]
(1)
(1) ), and the mathematical problem reduces to matching the continuity conditions on the boundary surfaces. The most basic case of such solutions is Lorenz-Mie theory, which provides the exact E (derived using separation-of-variables methods to Equation [1]) for a single sphere in an unbounded medium and excited by a plane wave. The extension of this approach to multiple spheres, and to spheres adjacent to plane boundaries is termed the Multi-Sphere T-Matrix (MSTM) method which has been described in detail (Mackowski Citation2008; Mackowski and Mishchenko Citation2011, Citation2013). The method requires the application of non-trivial mathematical techniques and cannot be simply described in a few equations or sentences. The implementation of the solution also requires numerical methods for solving linear equations. The overall computational effort, however, is considerably less than that associated with a direct numerical solution to EquationEquation (1)
(1)
(1) for the system under consideration.
Some previous attempts to approximate the UV intensities in clusters or agglomerates have assumed that the UV light travels in a straight line as it passes through the surfaces of spores (as if it were gamma rays), but attenuated by UV absorbing molecules in the material (Handler Citation2016; Handler and Edmonds Citation2015). No reflection or refraction was included in the model. This approach does not suggest the regions within a sphere that are well shielded because the rays refract (bend) away from these regions as they enter the sphere, regions which are clear in exact solutions for spheres illuminated with 266-nm light (Hill et al. Citation2015, Fig. 7), and in intensities calculated using geometrical optics (Chowdhury et al. Citation1992). Kowalski et al. (Citation2019) used Mie Theory to calculate scattering and extinction cross sections for an individual sphere. However, they assumed that the scattered light does not interact with any other spheres in the cluster even if the spheres are in contact. Even for a particle immediately downstream from a given sphere Kowalski et al. (Citation2019) assume that “any forward scattering that reaches the second particle is minor,” and thus they ignore it. These assumptions may be useful for approximations of UVGI in clusters in water in cases where the difference between the refractive index of bacteria and water is small (and so the refraction at the spore surface and the scattering by the individual bacterium tend to be small). However, Kowalski et al. (Citation2019) claimed incorrectly that such an approach was applicable to spores and other microbes in air. In Kowalski et al. (Citation2019, Fig. 2), rays traveling from the first to the second sphere are drawn parallel to the rays incident upon the sphere. In reality, a solid sphere in air, having a typical refractive index (1.5), illuminated by a plane wave, focuses a large fraction of the light impinging upon it into a small region just outside the sphere (Barber and Hill Citation1990, Figs. 4.32 and 4.34). Short-focal-length ball lenses are used in microscopy (Agbana et al. Citation2018). Microbes in air refract and reflect light at their surface.
The MSTM Fortran program, details of which can be found in Mackowski (Citation2008) and Mackowski and Mishchenko (Citation2011, Citation2013) was used by Doughty et al. (Citation2021) to calculate UV intensities in spherical particles which have a diameter (100 nm, in the range of the 60–140 nm reported for SARS-CoV-2 (Zhu et al. Citation2020)) and optical properties selected to approximate SARS-CoV-2 virions. In the rest of this paper, these spherical approximations of virions are termed “virions.” The virions were randomly positioned inside a “host” spherical particle (1-, 5, or 9-µm diameter) with optical properties approximating dried-to-equilibrium respiratory fluid. The 0.1-µm case is the lower-limit of host diameter. The host particles were in some cases resting on a surface. The calculated results showed that (1) virions in particles can be partially shielded from UVGI whether the host particle is on a surface or not; (2) virions can be protected both by absorption of UVGI within the host particle and by refraction at the air-particle surface; (3) the extent of shielding is affected by the wavelength of the UVGI and properties of the dried-droplet host particle; (4) shielding increases as the size of the host particle increases; (5) illumination with incident beams from widely separated directions tends to decrease the shielding from UVGI; and (6) shielding is decreased in particles illuminated uniformly from all directions as may occur for a particle in turbulent air illuminated for enough time to tumble and spin through all orientations relative to the incident beam(s).
In our previous modeling of shielding using the MSTM (Doughty et al. Citation2021), the key parameter calculated for each virion was
(2)
(2)
where Cabs,i is the absorption cross section of the ith virion in a particle (either resting on the surface or not), defined so that Cabs,iI0 is the net rate of energy transfer into the virion (i.e., the integration of the normal component of the Poynting vector over the surface of the ith virion), where I0 is the irradiance illuminating the host sphere. Cabs,single is the absorption cross section of an individual virion (either resting on the surface or not), i.e., defined so that Cabs,singleI0 is the net rate energy transfer into an individual virion. The absorption efficiency of a sphere is Qabs = Cabs/πa2, where a is the radius. A complete derivation of the cross-section formulas for spheres-inside-spheres is in Mackowski (Citation2014).
The Ri is especially useful for understanding the physics of shielding of microbes from UV and how and where it occurs. However, the survival fraction (S) is the more important quantity for modeling and designing systems and procedures for minimizing disease transmission. The simplest common expression for survival fraction (S) is the single-stage decay model (Kowalski Citation2009):
(3)
(3)
where k is the UV rate constant for inactivation (m2/J) and F is the UV-exposure dose (Fluence), energy per unit area (J/m2). Also, F = It, where I is the intensity of the irradiance (W/m2), and t is the time of illumination. For a given virus and set of experimental conditions (humidity, etc.) the S is measured for different F, and then k is determined using EquationEquation (3)
(3)
(3) . The k have been measured for SARS-CoV-2 (Heilingloh et al. Citation2020; Ratnesar-Shumate et al. Citation2020), and other viruses (Sagripanti and Lytle Citation2011; Schuit et al. Citation2020), bacteria, and fungi (Kowalski Citation2009). More complex expressions for S(F) of the survival fraction, such as those which incorporate an additional term or terms to account for shoulders, tails, or other features (Kowalski Citation2009, Ch. 3; Kowalski et al. Citation2019) are beyond the scope of this paper.
To account for shielding of spores within a cluster of spores, Handler and Edmonds (Citation2015) and Handler (Citation2016) estimated the F at each spore and used it to calculate an average S(F) for the spores in the cluster. They, and Kowalski et al. (Citation2019), assumed that microbes not on the illuminated surface of a cluster or particle are illuminated by an attenuated plane wave that travels in the same direction as the incident wave. However, the UV light within particles does not take the form of a simple attenuating plane wave (e.g., a virion in a particle may be at a minimum or maximum of a standing wave within the particle) and so an S based on assuming attenuated plane waves is not an adequate approach in general (see “Methods”).
The objective of this study is to increase understanding of the survival fraction of SARS-CoV-2 virions (and other virions and microbes) when the virions (or other microbes) are within particles which partially shield them from UVGI. The hope is that increased understanding of shielding from UVGI, and the modeling capabilities described, will allow designers and users of germicidal- or solar-UVGI systems to better mitigate the effects of shielding from UVGI. Here, the survival fraction (Si(kF)) for the ith virion in a particle is calculated as
(4)
(4)
(see “Methods”). We calculate and illustrate curves of average survival fractions vs. kF for many of the cases for which we calculated Ri previously (Doughty et al. Citation2021), for virions of SARS-CoV-2 (approximated as 100-nm diameter spherical particles) within otherwise homogeneous spherical particles having optical properties to approximate dried particles of sneezed or coughed respiratory fluids. As in Doughty et al. (Citation2021), the host particles have diameters of 1-, 5- and 9-µm (and 0.1 µm for the individual virion); the UV wavelengths are 260 and 302 nm; the particle is in air or on a surface with m = 1.4 + i0.0001; and there are several combinations of incident beam directions. The UV wavelengths were used because 302-nm light is at the most germicidal end of the solar UV, and 260-nm light is highly damaging to nucleic acids and close to the 254-nm primary emission of low-pressure mercury lamps.
Methods
The problem of calculating the UV intensities and estimating the optical properties of the virions and dried respiratory fluids was described in the Introduction and in Doughty et al. (Citation2021), with details on the MSTM given in Mackowski (Citation2008, Citation2014) and Mackowski and Mishchenko (Citation2011, Citation2013).
A way to obtain EquationEquation (4)(4)
(4) from EquationEquation (3)
(3)
(3) is as follows. In EquationEquation (3)
(3)
(3) , F is the same for each virion. We desire an expression for Si(kF) based on the energy absorbed by the ith virion in the host particle when the host particle is illuminated with fluence F. To achieve that expression, the numerator and denominator of the exponent in EquationEquation (3)
(3)
(3) are multiplied by Cabs,single to obtain
(5)
(5)
where FCabs,single = energy absorbed by a single virion illuminated with F, and where k/Cabs,single is the yield for viral inactivation (inactivations/J if k is in J/m2), or the quantum yield for viral inactivation (inactivations/photon if k is in m2/photon and F is in photons/m2 (Rauth Citation1965, EquationEq. 2
(2)
(2) )). Then to obtain the equivalent S(kF) for the ith virion, the FCabs,single in EquationEquation (5)
(5)
(5) is replaced by the energy absorbed by the ith virion when the particle is illuminated by F, that is, by FCabs,i, to obtain
(6)
(6)
which is equivalent to EquationEquation (4)
(4)
(4) . Also in EquationEquation (4)
(4)
(4) , the FRi can be taken to define an Fi for each virion.
The key function illustrated in the figures is Sp(kF), the average of the survival fractions (Si) of all the N modeled virions in all the particles with the same specified composition. It is defined as:
(7)
(7)
The Ri can range over several orders of magnitude in absorbing particles with dimensions of several wavelengths. In such cases, large numbers of Ri are needed to adequately simulate the distribution of Ri. For 9-µm particles, 120 simulations of 100 virions/particle were calculated and combined to approximate the distribution of Ri for randomly positioned virions, and so N = 12,000 for this case. The values of N, and why these N were chosen were discussed in Doughty et al. (Citation2021).
Additional parameters are defined as follows. The penetration depth, δ, is the distance z at which I(z)/Io = 1/e = 0.368 for a UV planewave propagating in a material, where the intensity of the irradiance of the wave at position (z) is
(8)
(8)
where Io is I at z = 0. The δ, mi and UV wavelength λ are related as
(9)
(9)
The notation for angles follows that used in the MSTM codes (Mackowski Citation2008). A line perpendicular to the surface and passing through the center of the particle and through the planar surface defines the z-axis. The zenith angle (β) is the angle from this axis. When there is more than one illumination beam with β ≠ 0˚, the azimuthal angles (α) between these beams are also specified.
provides, for each curve in the figures, the wavelength (λ) in nm, diameter (d) of the host particle in µm, numbers of virions (N) used to obtain the Sp(kF), mi, penetration depth (δ), incident angle(s) β (and α when needed), and Sp(kF) for specific values of kF: 1, 10, 100, 1000, 10,000. The Sp at specific kF are given because extracting accurate values from the Figures is tedious. The mi for the virions is 0.0021 at 260 nm and 0.000092 at 302 nm.
Table 1. Figure numbers, parameters, and average survival fraction of virions (Sp) for virions having UV-inactivation rate constant k within host particles illuminated with fluence F, for selected values of the dimensionless parameter kF.
Each curve in the figures is calculated by selecting a set of kF that are evenly spaced in log space (sufficiently close that the curve is smooth) and then using Equation (7) with the N values of Ri calculated with the parameters given in the specific row in (as in Doughty et al. Citation2021, Table 3), to calculate the Sp for each of the selected kF. Sp is plotted vs. the dimensionless parameter kF to emphasize the relation between k and F. If k = 1 m2/J and the abscissa is F in J/m2, no change in the tick labels would be needed. If k = 0.1 m2/J and the abscissa is F in J/m2, each labeled value for each tick would need to be increased by a factor of 10.
In the “Results and Discussion,” illustrates the effects of host particle diameter (0.1-, 1-, 5-, or 9-µm) on Sp(kF) for two λ, δ, and mi(λ). illustrates the effects of δ on Sp(kF) by using a set of four mi(λ), two at 260 nm and two at 302 nm, for 9-µm host particles. shows the effects of illumination from different incident angles and combinations of these on the shielding from UVGI, as seen in Sp(F), for 9-µm host particles. illustrates the effects on Sp(F) of illumination of particles over all orientations, as compared with illumination from one or a few angles.
Figure 1. Average survival fractions (Sp) vs. kF for 100 nm virions within spherical particles of dried respiratory fluids of the sized indicated and resting on a surface with real refractive index 1.4.
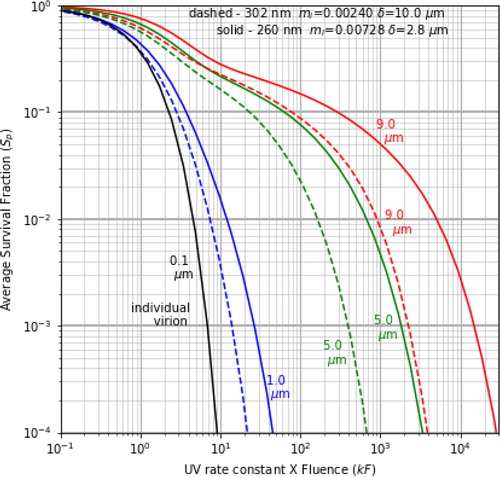
Figure 2. Sp vs. kF for different penetration depth (and mi) for the dried respiratory fluids, and all d = 9 µm except for the 0.1 µm individual virion shown for comparison. The blue and green curves are at 302 nm and the red and purple are at 260 nm.
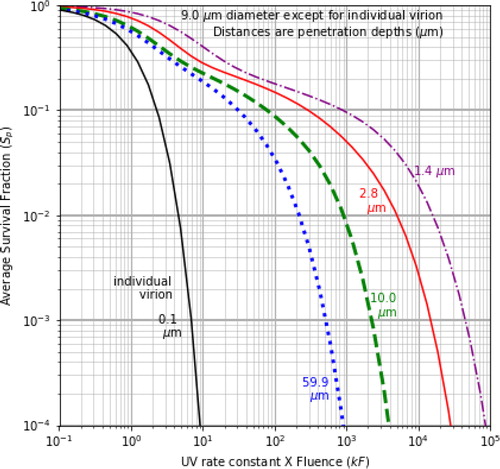
Figure 3. Sp vs. kF, for 9-µm particles, 260 nm UVGI, δ = 2.8 µm, with various combination of illumination directions: the three curves on the right are for a single illumination angle β = 0˚, 40˚ and 80˚; the next two curves have an illumination with β = 0˚ and a second wave, β = 40˚, or 80˚; and the next four curves have more widely separated beams. In each case, the total intensity illuminating the particle is the same.
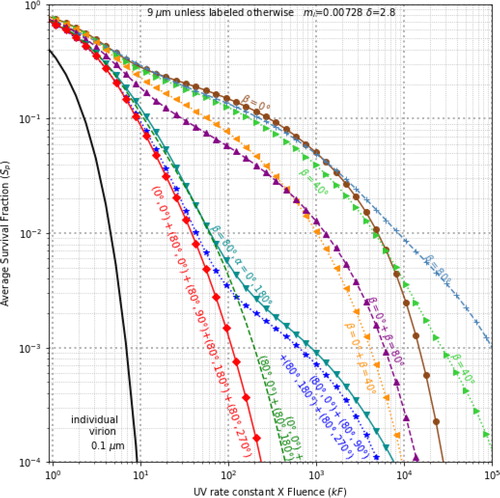
Figure 4. Sp vs. kF for virions in particles where the orientation averaged curves for 5-µm (green) and 9-µm (blue) are in dashed lines and are for particles not on a surface, and the other curves are shown for comparison. All cases are with 260-nm UVGI.
Results and discussion
Average survival fractions are larger in larger particles
shows Sp (average of the Si as in Eq. [7]) as a function of kF for virions within 1-, 5- and 9-µm diameter host particles, and for individual 0.1-µm-diameter virions. In , the Sp increases with host particle size for both wavelengths used (260 and 302 nm). As seen in and the table, when λ = 260 nm and kF = 10 the Sp is: 0.000045 for the individual virion, 0.0156 for the 1-µm particles, 0.217 for 5-µm particles, and 0.28 for the 9-µm particles. When λ = 260 nm and kF = 100 the Sp is: < 0.0000005 for the 1-µm particles, 0.077 for the 5-µm particles, and 0.149 for the 9-µm particles.
In , the Sp are larger in host particles when λ = 260 nm than when λ = 302 nm, but that does not mean that lower F are needed for inactivation. The mi for the virions is 0.0021 at 260 nm and 0.000092 at 302 nm (Doughty et al. Citation2021, Table 2), and so virions modeled here tend to be less absorbing of UVGI and the k tend to be smaller at 302 nm than at 260 nm. Also, the quantum yields for inactivation are lower at 302 nm than at 260 nm for the viruses reported by Rauth (Citation1965, Fig. 5) and that also tends to make the k smaller at 302 nm. Because k tends to be smaller at 302 nm than at 260 nm, a higher F is required to achieve the same kF and Sp.
The curves of Sp(kF) vs kF in are not simple exponentials (except for the individual-virion curves) because they are averages of many (N) simple exponentials. For the ith exponential curve, Si = 1/e when kF = 1/Ri. As kF increases, the Si of the virions with the largest Ri decrease faster than do the Si of virions with smaller Ri. Consequently, the relative contributions of the different virions to the Sp vs kF curve changes with kF. For large kF, the virions with the smallest Ri dominate the Sp.
Average survival fractions vs. kF are larger in particles with higher absorptivity
shows the Sp vs. kF curves for virions in host particles having different UV absorptivities for the material of the dried droplet, as indicated by the mi stated in the caption and . The curves are labeled by the penetration depth δ, which is useful for understanding the shielding. In , the mi was chosen to span a wider range than the estimated “typical” values used in the other figures. In , at 260 nm, in addition to the δ = 2.8 and 10 µm in , there is also a more strongly absorbing particle with δ = 1.4 µm (mi = 0.01456). At 302 nm, there is also a more weak-absorbing particle with δ = 59.9 µm (mi = 0.00040). This larger range of δ is used to better account for the large variations in some of the primary light-absorbing components of saliva or nasal fluids, such as, at 260 nm, nucleic acids (e.g., Poehls et al. Citation2018), and at 302 nm, uric acid (Hawkins et al. Citation1963; Riis et al. Citation2018, ).
In , the Sp of even the least absorbing particle is many times greater than that of the individual virion. For example, even when the δ is 6.7 particle diameters (i.e., the 9-µm particle at 302 nm with δ = 59.9 µm), when kF = 10, the Sp for the virions in the particle is 0.188, which is 4200 times greater than the Sp = 0.000045 for the individual virion. Because the estimated absorption is so low, the increase in Sp occurs primarily because of refractive shielding. For solar wavelengths longer than the 302-nm UVGI used here, the absorption tends to be even lower, with larger δ. However, because of refractive shielding the large increase in Sp remains (as compared to that of the individual virions).
This increase in Sp for higher absorption by the host particle is also seen in , where the Sp are smaller at λ = 302 nm than at λ = 260 nm because at 302 nm the particles are less absorbing of UVGI. For example, when kF = 10, the Sp for 1-µm particles are 0.0156 at 260 nm, but 0.00377 at 302 nm. Also when kF = 10, the Sp for 5-µm particles are 0.217 at 260 nm but 0.164 at 302 nm.
Survival fractions are smaller in particles illuminated from widely separated directions
and illustrate Sp vs. kF for illumination with beams with β = 0˚ or β ≠ 0˚, or with beams from multiple incident directions (with F split equally between the beams). These figures illustrate how illumination with beams from multiple angles can result in reductions in Sp. In , when kF = 10, the Sp in the 9-µm particles range from 0.0876 with 5 widely separated UV beams, to 0.282 with one UV beam at normal incidence. For the individual virion, Sp = 0.000045. When kF = 100, the Sp range from 0.149 with one UV beam with β = 0˚, to 0.00128 with 5 widely separated UV beams, i.e., 116 times smaller than the single-beam case.
In , the curves can be grouped into three main categories: (a) illumination with one beam only, with β = 0˚, 40˚, or 80˚; (b) illumination with multiple beams, but the differences in illumination angles are less than 90˚; and (c) illumination with multiple beams where at least one of the angular differences between two of the angles of incidence is greater than 90˚.
With only one illumination beam the differences in Sp distributions are relatively small, but not negligible. The set of beams providing the lowest Sp includes five beams: one beam heading perpendicular to the surface (β = 0˚), and four beams, each with β = 80˚ from the vertical but approaching from different azimuthal angles, α = 0˚, 90˚, 180˚, 270˚. At kF = 100, the Sp = 0.00128 for this set. A set of four beams with β = 80˚ and α = 0˚, 90˚, 180˚, or 270˚ (similar to the 5-beam case but with no β = 0˚ beam), has Sp = 0.00331.
Highly UV-reflective surfaces (Krishnamoorthy and Tande Citation2016; Lindsley et al. Citation2018; Rutala et al. Citation2013; Ryan et al. Citation2010) can increase overall UVGI and can help in directing UVGI into shaded regions of a room. The results in suggest an additional benefit of highly UV-reflective surfaces, i.e., helping to achieve illumination with multiple beams where the angular differences between the angles of incidence with respect to the surface is relatively large (see Doughty et al. (Citation2021) for the Ri distributions).
In and , the UV illumination energy is split between the beams so that the total intensity in each of the cases in can be directly compared. Potential increases in the overall UV-illumination intensity that could result from generating additional beams using reflective surfaces could very likely decrease the Sp. However, such calculations depend upon factors such as the positions, angles and reflectivities of the relevant surfaces, and are beyond the scope of this paper.
shows the orientation-averaged Sp vs kF for virions in 5- and 9-µm particles (dashed lines). Such orientation averaged results might be obtained with a fixed-orientation particle illuminated equally from all directions or with a particle that rotates (tumbles and spins) through all orientations with respect to one or more UV beams.
In (see also the numbers in ), the reductions in Sp with orientation averaging are remarkable. For 5-µm particles when kF = 100, the Sp is less than 5 × 10−7 for the orientation-averaged case and is 0.0768 for the single beam with β = 0˚. For kF > 9, the orientation-averaged case has a lower Sp than any of the other cases with 9-µm. For 9-µm particles when kF = 100, the Sp is less than 5 × 10−7 for the orientation-averaged case but is 0.00128 for the 5 beam case, and 0.149 for the single beam β = 0˚ case. Also in , when kF > 11 the Sp for the orientation-averaged 5-µm particles is less than the Sp for 1-µm particles (β = 0˚). When kF > 33, the Sp for the orientation-averaged 9-µm particles is less than Sp for 1-µm particles (β = 0˚). Such results may appear confusing. How can orientation averaging be so important compared to a 9-fold difference in diameter when the smaller particle is only 1-µm diameter? The reason is that no virions are refractively shielded well in the orientation-averaged particles. But the wavelength is so short (260 nm), that even a 1-µm particle illuminated from one direction can have 100-nm regions that are refractively shielded (e.g., Hill et al. Citation2015, Figure 7a).
Summary: virions in particles tend to have larger UV survival fractions
Virions within UV-illuminated spherical particles (homogeneous except for the virions) tend to have larger Sp than do individual virions, as seen in the calculated values in the figures and table. Several key relations can be seen in the calculated results.
For host particles on a surface with a given mr + i mi, illuminated with normally incident UVGI (zenith angle β = 0˚) at a given wavelength, as the host particle size increases the Sp increases (except when kF<1 for 1-um particles at 302 nm) ( and ). This relation occurs for both wavelengths studied (260 and 302 nm). To achieve Sp = 0.001 in 1-µm particles with 260 nm light, kF must be approximately 4× higher for 1-µm particles, 230× higher for the 5-µm particles and 2000x greater for the 9-µm particles, all with respect to the individual virion. At 302 nm, the increases in kF required are smaller: 2×, 56× and 300×, respectively, because the absorption of UV by the particle is smaller.
For particles on a surface illuminated with normally incident UVGI (β = 0), as the penetration depth in the host particles decreases the Sp increases. This relation is most clear in where the curves are labeled with the four different δ, but is also seen in .
UV illumination of particles from multiple directions, especially widely varying directions, can reduce the Sp ( and , and ). Particles illuminated equally from all directions tend to have especially low Sp (). Particles in outdoor air for a sufficient time on a gusty and sunny day may be examples of particles illuminated from all directions. UVGI systems may be designed to illuminate particles somewhat uniformly from all orientations. Or, all-orientation Sp may be approximated by particles that tumble and spin sufficiently rapidly as they are carried within turbulent airflows through a UVGI system with illumination from one side.
Uncertainties
The compositions of airway fluids may vary with a person’s genetic makeup, disease state (if any), time since last drink or meal, degree of hydration, exertion level, region of the respiratory tract, and other factors. In the estimates of mi from airway-fluid compositions used here, the molecules absorbing the most UVGI at 260-nm are DNA, RNA, and nucleosides/nucleotides. In using calculated Sp to improve the use of UVGI in reducing the transmission of disease, a sense of the ranges of measured concentrations of UV-absorbing molecules such as nucleic acids can be beneficial. Konečná et al. (Citation2020) reported average dsDNA concentrations in whole saliva of 0.24 g/L in healthy subjects and 0.5 g/L in persons with periodontitis. In supernatants from centrifuged (1,600× g) samples they measured average concentrations of DNA of 0.054 g/L in healthy subjects and 0.07 g/L in persons with periodontitis. Fahy et al. (Citation1995) measured a range of cell-free DNA concentrations from 0.06 to 5.8 g/L (average 0.5 g/L) in patients with asthma. Pandit et al. (Citation2013) found the extracellular RNA in the saliva (supernatant after centrifugation) ranged from 0.17 to 0.76 g/L in healthy control subjects and from 0.09 to 0.36 g/L in cancer patients. SARS-CoV-2 and other microbes can induce neutrophils in saliva to release DNA, histones, and proteins to form Neutrophil Extracellular Traps (NETs) (Arcanjo et al., Citation2020; Lachowicz-Scroggins et al. Citation2019). Eosinophil Extracellular Traps (EETs) and Basophile Extracellular Traps (BETs) also contain DNA and contribute to airway defense and inflammation (Yousefi et al. Citation2020). Bacterial and viral DNA and RNA (Haro et al. Citation2020) also occur in airway fluids, but typically in lower concentrations than host DNA and RNA. To partially account for the large variability in DNA and RNA concentrations, a large range of mi are used in the calculations here ().
In this paper, the model host particles are spherical and homogeneous (except for the virions), and the positions of virions within each particle are random. The MSTM can be used to model UVGI in more-complex particles than those investigated here, e.g., particles with additional spherical regions inside and outside the particle, each with its own mr and mi. However, such modeling requires measurements or assumptions of the geometries of the particles and mr and mi of each of the various regions. The homogeneous-sphere-with-virions assumption used here is a useful case to investigate first, given the large numbers of relevant variables: mr and mi of the virion(s), host particle and surface; size(s) of the virions and the host particle; virion locations; wavelengths; number and angles of beams; presence of a surface or not. However, for more accurate modeling of the Sp of more complex particles, additional studies along the lines of those of Vejerano and Marr (Citation2018) and Walker et al. (Citation2021) would likely be needed.
Vejerano and Marr (Citation2018) collected droplets of artificial saliva on a superhydrophobic surface and used optical microscopy to examine the particles as they dried as the RH was lowered. In their fluorescence images, particles that were less than roughly 20 µm appear to us as approximately spherical, but inhomogeneous. The nonsphericities appear to arise from the crystallization/precipitation of different materials in different locations. Walker et al. (Citation2021, their Fig. 5c) collected dried particles of artificial saliva on a surface and examined them with scanning electron microscopy. The particles generally look smooth on the order of the apparent resolution, except for some unclear features. The smooth appearance suggests that particles remaining after drying in air (as in Walker et al. Citation2021) or after drying on a superhydrophobic surface can be quite different from the material remaining after a droplet of a liquid or liquid/solid mixture is deposited on a not-superhydrophobic surface and then dried (as in Papineni and Rosenthal Citation1997). Walker et al. (Citation2021) also state that light scattering patterns indicate that the particles are inhomogeneous. Vejerano and Marr (Citation2018) and Walker et al. (Citation2021) are the only studies (to the authors' knowledge) with as much information related to morphologies of small particles from dried saliva or respiratory fluids with sizes in the range of interest (e.g., Fennelly Citation2020) and which dry in air or on a superhydrophobic surface. However, there are uncertainties in how to apply these studies to modeling the Sp of dried saliva or respiratory particles in the 0.5- to 9-µm size range because the sizes, positions and the optical properties (or compositions) of internal structures (if any) are not specified, and because respiratory fluids tend to be more complex than the artificial fluids used in some parts of the studies mentioned. Differences between human and model saliva, and between gastric and submaxillary mucin, are not insignificant (Sarkar et al. Citation2019). Because of the additional complexity of actual saliva and other respiratory fluids (containing DNA, RNA, mucins, and other proteins, carbohydrates, etc.), there is cause to wonder if crystallization/precipitation may be more likely to occur as smaller crystals in dried coughed/sneezed droplets, as compared with fewer larger crystals in the artificial salivas. For their fluorescence images, Vejerano and Marr (Citation2018, Figs. 4 and 5) used an aqueous solution of gastric mucin, NaCl, and a phospholipid surfactant. Walker et al. (Citation2021), following (Woo et al. Citation2012), used an aqueous solution of Na+, K+, Mg2+, Ca2+, Cl-, HCO3 -, H2PO4 -, SCN-, amylase, mucin from porcine stomach, glucose and very small concentrations of glucose, amino acids, vitamins, cofactors, and sulfate. Neither includes DNA or RNA, which may affect the diffusion rates of various solutes (Dix and Verkman Citation2008) or particles (Linssen et al. Citation2021) within droplets.
Conclusions
Average survival fractions (Sp) of virions in particles of dried respiratory fluids exposed to UVGI can be much larger than survival fractions of virions not within such particles, as illustrated in . The Sp were calculated using UV intensities within virions calculated with the MSTM, an exact method (within numerical error). The model particles are spherical and homogeneous, except for the virions. The particles may be on a surface and may be illuminated with any number of incident beams.
The key findings are:
The Sp increases as the particle size increases, where the smallest relevant particle is an individual virion ( and ).
The Sp increases as the UV-absorptivity of the material comprising the particle increases ( and ). However, even when the material of the particle absorbs very little UV light, the Sp increases as the particle size increases.
The Sp tends to decrease as more UV beams, from more widely separated directions, illuminate the particle, where the total UV energy is the same for each case ( and ). In the limiting case of equal illumination from all angles, the Sp tend to be especially low, e.g., Sp for the orientation-averaged 5-µm particles is smaller than the Sp for fixed orientation 1-µm particles unless kF<11 (). The orientation-averaged case is equivalent to the one where particles rotate (spin and tumble) through all orientations with respect to the illumination beams during their time of exposure to UVGI.
Increases in Sp for virions in particles can be mitigated by using more intense UVGI sources, and/or more UVGI sources, and/or with highly UV-reflective surfaces (Lindsley et al. Citation2018; Rutala et al. Citation2013; Ryan et al. Citation2010). An advantage of using highly UV-reflective surfaces is that in addition to increasing the UV intensity, they can also be positioned to increase the numbers of directions from which particles are illuminated, and the angular separation between the various angles of illumination, and thus substantially reduce the Sp as compared to illumination with a single-beam or beams from similar directions.
Detailed, accurate modeling of the Sp of virions in particles requires knowledge of the morphologies of the virion-containing particles, i.e., the 3-D shapes, positions, and compositions or optical properties of each region of the particle.
Recommendations
In designing, testing, and using UVGI systems to inactivate viruses, recognize that virions within particles larger than a very few UV wavelengths tend to have larger survival fractions than do individual virions, and that the survival fractions increase with particle size.
In evaluating UVGI systems for efficacy of inactivation, test for inactivation with virion-containing particles having sizes and compositions representative of the particles that need to be inactivated. For example, if tests were done only with virion-containing particles smaller than 1 µm, but the system may be used with a significant fraction of particles as large as 4 to 5 µm, then recognize that the results shown here suggest that the measured survival fractions will very likely be higher for the 4- to 5-µm particles.
In designing and deploying UVGI systems for virions in particles sufficiently large- and/or absorptive to shield virions, consider, to the extent possible, systems that illuminate particles from many, widely separated, angles (see and ). Consider employing systems with surfaces that are highly reflective at the relevant UV wavelengths, because such surfaces can increase both the UV intensities and the ranges of angles of incidence of the UV illumination, and thereby decrease the survival fractions. For systems in which particles of the relevant sizes spin and tumble through all orientations as they are illuminated, the need for illumination from many directions, including reflected beams, is not necessary, but it should be recognized that verification of adequate tumble/spin rates is not trivial for many rapid flow-through systems.
Code availability
Researchers interested in the MSTM codes to calculate the results Qabs for virions may email D.W. Mackowski ([email protected]). Those interested in using the MSTM to generate the Ri for the cases studied here may email D. C. Doughty ([email protected]). Those interested in using the Ri to generate the Figures or may email S. C. Hill.
Acknowledgments
Hill and Doughty acknowledge partial support from DEVCOM ARL. All authors acknowledge partial support from the Defense Threat Reduction Agency.
References
- Agbana TE, Diehl J-C, van Pul F, Khan SM, Patlan V, Verhaegen M, Vdovin G. 2018. Imaging & identification of malaria parasites using cellphone microscope with a ball lens. PLOS One. 13(10):e0205020. doi:10.1371/journal.pone.0205020
- Anderson DJ, Moehring RW, Weber DJ, Lewis SS, Chen LF, Schwab JC, Becherer P, Blocker M, Triplett PF, Knelson LP, et al. 2018. Effectiveness of targeted enhanced terminal room disinfection on hospital-wide acquisition and infection with multidrug-resistant organisms and Clostridium difficile: a secondary analysis of a multicentre cluster randomised controlled trial with crossover design (BETR Disinfection). Lancet Infect Dis. 18(8):845–853. doi:10.1016/S1473-3099(18)30278-0
- Arcanjo A, Logullo J, Barreto Menezes CC, Chrispim de Souza Carvalho Giangiarulo T, Carneiro dos Reis M, Menezes Migliani de Castro G, da Silva Fontes Y, Regina Todeschini A, Freire‑de‑Lima L, Decoté‑Ricardo D, et al. 2020. The emerging role of neutrophil extracellular traps in severe acute respiratory syndrome coronavirus 2 (COVID-19). Sci Rep. 10(1):19630. doi:10.1038/s41598-020-76781-0
- Augenbraun BL, Lasner ZD, Mitra D, Prabhu S, Raval S, Sawaoka H, Doyle JM. 2020. Assessment and mitigation of aerosol airborne SARS-CoV-2 transmission in laboratory and office environments. J Occup Environ Hyg. 17(10):447–456. doi:10.1080/15459624.2020.1805117.
- Azzi L, Carcano G, Gianfagna F, Grossi P, Dalla Gasperina D, Genoni A, Fasano M, Sessa F, Tettamanti L, Carinci F, et al. 2020. Saliva is a reliable tool to detect SARS-CoV-2. J Infect. 81(1):e45–e50. doi:10.1016/j.jinf.2020.04.005
- Barber PW, Hill SC. 1990. Light scattering by particles: computational methods. Singapore: World Scientific. p. 235–237.
- Bohren CF, Huffman DR. 1983. Absorption and scattering of light by small particles. New York: Wiley.
- Boone SA, Gerba CP. 2007. Significance of fomites in the spread of respiratory and enteric viral disease. Appl Env Microbiol. 73(6):1687–1696. doi:10.1128/AEM.02051-06
- Cheng VC-C, Wong S-C, Chuang VW-M, So SY-C, Chen JH-K, Sridhar S, To KK-W, Chan JF-W, Hung IF-N, Ho P-L, et al. 2020. The role of community-wide wearing of face mask for control of coronavirus disease 2019 (COVID-19) epidemic due to SARS-CoV-2. J Infect. 81(1):107–114. doi:10.1016/j.jinf.2020.04.024
- Chowdhury DQ, Barber PW, Hill SC. 1992. Energy-density distribution inside large nonabsorbing spheres by using Mie theory and geometrical optics. Appl Opt. 31(18):3518–3523. doi:10.1364/AO.31.003518
- Coohill TP, Sagripanti J-L. 2008. Overview of the inactivation by 254 nm ultraviolet radiation of bacteria with particular relevance to biodefense. Photochem Photobiol. 84(5):1084–1090. doi:10.1111/j.1751-1097.2008.00387.x
- Dix JA, Verkman AS. 2008. Crowding effects on diffusion in solutions and cells. Annu Rev Biophys. 37:247–263. doi:10.1146/annurev.biophys.37.032807.125824
- Doughty DC, Hill SC, Mackowski DW. 2021. Viruses such as SARS-CoV-2 can be partially shielded from UV radiation when in particles generated by sneezing or coughing: numerical simulations. J Quant Spectrosc Radiat Transf. 262:107489. doi:10.1016/j.jqsrt.2020.107489
- Fahy JV, Kim KW, Liu J, Boushey HA. 1995. Prominent neutrophilic inflammation in sputum from subjects with asthma exacerbation. J Allergy Clin Immunol. 95(4):843–852. doi:10.1016/s0091-6749(95)70128-1
- Fennelly KP. 2020. Particle sizes of infectious aerosols: implications for infection control. Lancet Respir Med. 8(9):914–924. doi:10.1016/S2213-2600(20)30323-4
- Fischer RJ, Morris DH, van Doremalen N, Sarchette S, Matson MJ, Bushmaker T, Yinda CK, Seifert SN, Gamble A, Williamson BN, et al. 2020. Effectiveness of N95 respirator decontamination and reuse against SARS-CoV-2 virus. Emerg Infect Dis. 26(9):2253. doi:10.3201/eid2609.201524
- Fisher EM, Richardson AW, Harpest SD, Hofacre KC, Shaffer RE. 2012. Reaerosolization of MS2 bacteriophage from an N95 filtering facepiece respirator by simulated coughing. Ann Occup Hyg. 56(3):315–325. doi:10.1093/annhyg/mer101
- Guy GP, Lee FC, Sunshine G, McCord R, Howard-Williams M, Kompaniyets L, Dunphy C, Gakh M, Weber R, Sauber-Schatz E, et al. 2021. Association of state-issued mask mandates and allowing on-premises restaurant dining with county-level COVID-19 case and death growth rates - United States, March 1-December 31, 2020. MMWR Morb Mortal Wkly Rep. 70(10):350–354. doi:10.15585/mmwr.mm7010e3
- Handler FA. 2016. Modeling ultraviolet dose-response of Bacillus spore clusters in air. Aerosol Sci Technol. 50(2):148–159. doi:10.1080/02786826.2015.1137556
- Handler FA, Edmonds JM. 2015. Quantitative analysis of effects of UV exposure and spore cluster size on deposition and inhalation hazards of Bacillus spores. Aerosol Sci Techol. 49(11):1121–1130. doi:10.1080/02786826.2015.1102857
- Haro K, Ogawa M, Saito M, Kusuhara K, Fukuda K. 2020. Bacterial composition of nasal discharge in children based on highly accurate 16S rRNA gene sequencing analysis. Sci Rep. 10(1):20193. doi:10.1038/s41598-020-77271-z
- Hawkins GR, Zipkin I, Marshall LM. 1963. Determination of uric acid, tyrosine, tryptophan, and protein in whole human parotid saliva by ultraviolet absorption spectrophotometry. J Dent Res. 42(4):1015–1022. doi:10.1177/00220345630420040301
- Heilingloh CS, Aufderhorst UW, Schipper L, Dittmer U, Witzke O, Yang D, Zheng X, Sutter K, Trilling M, Alt M, et al. 2020. Susceptibility of SARS-CoV-2 to UV irradiation. Am J Infect Control. 48(10):1273–1275. doi:10.1016/j.ajic.2020.07.031
- Hill SC, Williamson CC, Doughty DC, Pan Y-L, Santarpia JL, Hill HH. 2015. Size-dependent fluorescence of bioaerosols: mathematical model using fluorescing and absorbing molecules in bacteria. J Quant Spectrosc Radiat Transf. 157:54–70. doi:10.1016/j.jqsrt.2015.01.011
- Huang N, Pérez P, Kato T, Mikami Y, Okuda K, Gilmore RC, Conde CD, Gasmi B, Stein S, Beach M, et al. 2021. SARS-CoV-2 infection of the oral cavity and saliva. Nat Med. 27(5):892–903. doi:10.1038/s41591-021-01296-8
- Joo H, Miller GF, Sunshine G, Gakh M, Pike J, Havers FP, Kim L, Weber R, Dugmeoglu S, Watson C, et al. 2021. Decline in COVID-19 hospitalization growth rates associated with statewide mask mandates — 10 states, March–October 2020. MMWR. 70(6):212–216. doi:10.15585/mmwr.mm7006e2
- Kesavan J, Schepers D, Bottiger J, Edmonds J. 2014. UV-C decontamination of aerosolized and surface-bound single spores and bioclusters. Aerosol Sci Technol. 48(4):450–457. doi:10.1080/02786826.2014.889276
- Kesavan JS, Humphreys PD, Bottiger JR, Valdes ER, Rastogi VK, Knox CK. 2017. Deposition method, relative humidity, and surface property effects of bacterial spore reaerosolization via pulsed air jet. Aerosol Sci Technol. 51(9):1027–1034. doi:10.1080/02786826.2017.1335389
- Konečná B, Gaál Kovalčíková A, Pančíková A, Novák B, Kovaľová E, Celec P, Tóthová Ľ. 2020. Salivary extracellular DNA and DNase activity in periodontitis. Appl Sci. 10(21):7490. doi:10.3390/app10217490
- Kowalski W. 2009. Ultraviolet germicidal irradiation handbook: UVGI for air and surface disinfection. New York: Springer. doi:10.1007/978-3-642-01999-9
- Kowalski WJ, Bahnfleth WP, Raguse M, Moeller, R. 2019. The cluster model of ultraviolet disinfection explains tailing kinetics. J Appl Microbiol. 128:1003–1014. doi:10.1111/jam.14527
- Krauter P, Biermann A. 2007. Reaerosolization of fluidized spores in ventilation systems. Appl Environ Microbiol. 73(7):2165–2172. doi:10.1128/AEM.02289-06
- Krishnamoorthy G, Tande BM. 2016. Improving the effectiveness of ultraviolet germicidal irradiation through reflective wall coatings: experimental and modeling based assessments. Indoor Built Environ. 25(2):314–328. doi:10.1177/1420326X14547785
- Lachowicz-Scroggins ME, Dunican EM, Charbit AR, Raymond W, Looney MR, Peters MC, Gordon ED, Woodruff PG, Lefrançais E, Phillips BR, et al. 2019. Extracellular DNA, neutrophil extracellular traps, and inflammasome activation in severe asthma. Am J Respir Crit Care Med. 199(9):1076–1085. doi:10.1164/rccm.201810-1869OC
- Landry ML, Criscuolo J, Peaper DR. 2020. Challenges in use of saliva for detection of SARS CoV-2 RNA in symptomatic outpatients. J Clin Virol. 130:104567. doi:10.1016/j.jcv.2020.104567
- Lindsley WG, McClelland TL, Neu DT, Martin SB, Mead KR, Thewlis RE, Noti JD. 2018. Ambulance disinfection using ultraviolet germicidal irradiation (UVGI): effects of fixture location and surface reflectivity. J Occup Environ Hyg. 15(1):1–12. doi:10.1080/15459624.2017.1376067
- Linssen RS, Chai G, Ma J, Kummarapurugu AB, van Woensel JBM, Bem RA, Kaler L, Duncan GA, Zhou L, Rubin BK, et al. 2021. Neutrophil extracellular traps increase airway mucus viscoelasticity and slow mucus particle transit. Am J Respir Cell Mol Biol. 64 (1):69–78. doi:10.1165/rcmb.2020-0168OC
- Lu J, Gu J, Li K, Xu K, Su W, Lai Z, Zhou D, Yu C, Xu B, Yang Z. 2020. COVID-19 outbreak associated with air conditioning in restaurant, Guangzhou, China, 2020. Emerg Infect Dis. 26(7):1628–1631. doi:10.3201/eid2607.200764
- Mackowski DW. 2008. Exact solution for the scattering and absorption properties of sphere clusters on a plane surface. J Quant Spectrosc Radiat Transf. 109(5):770–788. doi:10.1016/j.jqsrt.2007.08.024
- Mackowski DW. 2014. A general superposition solution for electromagnetic scattering by multiple spherical domains of optically active media. J Quant Spectrosc Radiat Transf. 133:264–270. doi:10.1016/j.jqsrt.2013.08.012
- Mackowski DW, Mishchenko MI. 2011. A multiple sphere T-matrix Fortran code for use on parallel computer clusters. J Quant Spectrosc Radiat Transf. 112(13):2182–2192. doi:10.1016/j.jqsrt.2011.02.019
- Mackowski DW, Mishchenko MI. 2013. Direct simulation of extinction in a slab of spherical particles. J Quant Spectrosc Radiat Transf. 123:103–112. doi:10.1016/j.jqsrt.2013.02.008
- Miller SL, Nazaroff WW, Jimenez JL, Boerstra A, Buonanno G, Dancer SJ, Kurnitski J, Marr LC, Morawska L, Noakes C. 2020. Transmission of SARS-CoV-2 by inhalation of respiratory aerosol in the Skagit Valley Chorale superspreading event. Indoor Air. 31(2):314–310. doi:10.1111/ina.12751
- Mitze T, Kosfeld R, Rode J, Walde K. 2020. Face masks considerably reduce COVID-19 cases in Germany. Proc Natl Acad Sci USA. 117(51):32293–32301. doi:10.1073/pnas.2015954117
- Morawska L, Tang JW, Bahnfleth W, Bluyssen PM, Boerstra A, Buonanno G, Cao J, Dancer S, Floto A, Franchimon F, et al. 2020. How can airborne transmission of COVID-19 indoors be minimised? Environ Int. 142:105832. doi:10.1016/j.envint.2020.105832
- Osman S, Peeters Z, La Duc MT, Mancinelli R, Ehrenfreund P, Venkateswaran K. 2008. Effect of shadowing on survival of bacteria under conditions simulating the Martian atmosphere and UV radiation. Appl Environ Microbiol. 74(4):959–970. doi:10.1128/AEM.01973-07
- Pan Y, Zhang D, Yang P, Poon LLM, Wang Q. 2020. Viral load of SARS-CoV-2 in clinical samples. Lancet Infect Dis. 20(4):411–412. doi:10.1016/S1473-3099(20)30113-4
- Pandit P, Cooper-White J, Punyadeera C. 2013. High-yield RNA-extraction method for saliva. Clin Chem. 59(7):1118–1122. doi:10.1373/clinchem.2012.197863
- Papineni RS, Rosenthal FS. 1997. The size distribution of droplets in the exhaled breath of healthy human subjects. J Aerosol Med. 10(2):105–116. doi:10.1089/jam.1997.10.105
- Paton S, Thompson K-A, Parks SR, Bennett AM. 2015. Reaerosolization of spores from flooring surfaces to assess the risk of dissemination and transmission of infections. Appl Environ Microbiol. 81(15):4914–4919. doi:10.1128/AEM.00412-15.10.1128/AEM.00412-15
- Péré H, Podglajen I, Wack M, Flamarion E, Mirault T, Goudot G, Hauw-Berlemont C, Le L, Caudron E, Carrabin S, et al. 2020. Nasal swab sampling for SARS-CoV-2: a convenient alternative in times of nasopharyngeal swab shortage. J Clin Microbiol. 58:e00721-20. doi:10.1128/JCM.00721-20
- Poehls UG, Hack CC, Ekici AB, Beckmann MW, Fasching PA, Ruebner M, Huebner H. 2018. Saliva samples as a source of DNA for high throughput genotyping: an acceptable and sufficient means in improvement of risk estimation throughout mammographic diagnostics. Eur J Med Res. 23(1):20. doi:10.1186/s40001-018-0318-9
- Qian J, Peccia J, Ferro AR. 2014. Walking-induced particle resuspension in indoor environments. Atmos Environ. 89:464–481. doi:10.1016/j.atmosenv.2014.02.035
- Ratnesar-Shumate S, Williams G, Green B, Krause M, Holland B, Wood S, Bohannon J, Boydston J, Freeburger D, Hooper I, et al. 2020. Simulated sunlight rapidly inactivates SARS-CoV-2 on surfaces. J Infect Dis. 222(2):214–222. doi:10.1093/infdis/jiaa274
- Rauth AM. 1965. The physical state of viral nucleic acid and the sensitivity of viruses to ultraviolet light. Biophys J. 5(3):257–273. doi:10.1016/S0006-3495(65)86715-7
- Riis JL, Bryce CI, Matin MJ, Stebbins JL, Kornienko O, Huisstede LV, Granger DA. 2018. The validity, stability, and utility of measuring uric acid in saliva. Biomark Med. 12(6):583–596. doi:10.2217/bmm-2017-0336
- Rutala WA, Gergen MF, Tande BM, Weber DJ. 2013. Rapid hospital room decontamination using ultraviolet (UV) light with a nanostructured UV-reflective wall coating. Infect Control Hosp Epidemiol. 34(5):527–529. doi:10.1086/67021.
- Rutala WA, Weber DJ. 2019. Best practices for disinfection of noncritical environmental surfaces and equipment in health care facilities: a bundle approach. Am J Infect Cont. 47:A96–A105. doi:10.1016/j.ajic.2019.01.014
- Ryan K, McCabe K, Clements N, Hernandez M, Miller SL. 2010. Inactivation of airborne microorganisms using novel ultraviolet radiation sources in reflective flow-through control devices. Aerosol Sci Technol. 44(7):541–550. doi:10.1080/02786821003762411
- Sarkar A, Xu F, Lee S. 2019. Human saliva and model saliva at bulk to adsorbed phases - similarities and differences. Adv Colloid Interface Sci. 273:102034. doi:10.1016/j.cis.2019.102034
- Sagripanti J-L, Lytle CD. 2011. Sensitivity to ultraviolet radiation of Lassa, vaccinia, and Ebola viruses dried on surfaces. Arch Virol. 156(3):489–494. doi:10.1007/s00705-010-0847-1
- Schuit M, Gardner S, Wood S, Bower K, Williams G, Freeburger D, Dabisch P. 2020. The influence of simulated sunlight on the inactivation of influenza virus in aerosols. J Infect Dis. 221(3):372–378. doi:10.1093/infdis/jiz582
- Vejerano EP, Marr LC. 2018. Physico-chemical characteristics of evaporating respiratory fluid droplets. J R Soc Interface. 15(139):20170939. doi:10.1098/rsif.2017.0939
- Walker JS, Archer J, Gregson FKA, Michel SES, Bzdek BR, Reid JP. 2021. Accurate representations of the microphysical processes occurring during the transport of exhaled aerosols and droplets. ACS Cent Sci. 7(1):200–209. doi:10.1021/acscentsci.0c01522
- Wolfel R, Corman VM, Guggemos W, Seilmaier M, Zange S, Muller MA, Niemeyer D, Jones TC, Vollmar P, Rothe C, et al. 2020. Virological assessment of hospitalized patients with COVID-2019. Nature. 581(7809):465–469. doi:10.1038/s41586-020-2196-x
- Woo M-H, Grippin A, Anwar D, Smith T, Wu C-Y, Wander JD. 2012. Effects of relative humidity and spraying medium on UV decontamination of filters loaded with viral aerosols. Appl Environ Microbiol. 78(16):5781–5787. doi:10.1128/AEM.00465-12
- Wyllie AL, Fournier J, Casanovas-Massana A, Campbell M, Tokuyama M, Vijayakumar P, Warren JL, Geng B, Muenker MC, Moore AJ, et al. 2020. Saliva or nasopharyngeal swab specimens for detection of SARS-CoV-2. N Engl J Med. 383(13):1283–1286. doi:10.1056/NEJMc2016359
- Yilmaz A, Marklund E, Andersson M, Nilsson S, Andersson L-M, Lindh M, Gisslen M. 2021. Upper respiratory tract levels of severe acute respiratory syndrome coronavirus 2 RNA and duration of viral RNA shedding do not differ between patients with mild and severe/critical coronavirus disease 2019. J Infect Dis. 223(1):15–18. doi:10.1093/infdis/jiaa632
- Yousefi S, Simon D, Stojkov D, Karsonova A, Karaulov A, Simon H-U. 2020. In vivo evidence for extracellular DNA trap formation. Cell Death Dis. 11(4):300. doi:10.1038/s41419-020-2497-x
- Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, Zhao X, Huang B, Shi W, Lu R, et al. 2020. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med. 382(8):727–733. doi:10.1056/NEJMoa2001017
- Zou L, Ruan F, Huang M, et al. 2020. SARS-CoV-2 viral load in upper respiratory specimens of infected patients. N Engl J Med. 382:12. doi:10.1056/NEJMc2001737
