
Abstract
Occupational immune diseases are a serious public health burden and are often a result of exposure to low molecular weight (LMW) chemicals. The complete immunological mechanisms driving these responses are not fully understood which has made the classification of chemical allergens difficult. Antimicrobials are a large group of immunologically-diverse LMW agents. In these studies, mice were dermally exposed to representative antimicrobial chemicals (sensitizers: didecyldimethylammonium chloride (DDAC), ortho-phthalaldehyde (OPA), irritants: benzal-konium chloride (BAC), and adjuvant: triclosan (TCS)) and the mRNA expression of cytokines and cellular mediators was evaluated using real-time qPCR in various tissues over a 7-days period. All antimicrobials caused increases in the mRNA expression of the danger signals Tslp (skin), and S100a8 (skin, blood, lung). Expression of the TH2 cytokine Il4 peaked at different timepoints for the chemicals based on exposure duration. Unique expression profiles were identified for OPA (Il10 in lymph node, Il4 and Il13 in lung) and TCS (Tlr4 in skin). Additionally, all chemicals except OPA induced decreased expression of the cellular adhesion molecule Ecad. Overall, the results from these studies suggest that unique gene expression profiles are implicated following dermal exposure to various antimicrobial agents, warranting the need for additional studies. In order to advance the development of preventative and therapeutic strategies to combat immunological disease, underlying mechanisms of antimicrobial-induced immunomodulation must be fully understood. This understanding will aid in the development of more effective methods to screen for chemical toxicity, and may potentially lead to more effective treatment strategies for those suffering from immune diseases.
Keywords:
Introduction
Millions of workers have the potential for dermal and/or respiratory exposure to low molecular weight (LMW) chemicals which can result in occupational diseases. While the number of chemicals used in industrial applications now exceeds 82,000, according to the EPA, ≈ 3000 additional new chemicals are introduced every year. Workplace exposures can result in a number of allergic diseases; ≈ 11 million American workers are at risk for exposure to agents that can cause allergic disease (Anderson and Meade Citation2014). Exposures to certain LMW chemicals can impact immune function that can result in uncontrolled inflammation, increased susceptibility to infection and disease, or allergic disease. These conditions may be detrimental to a worker’s health and workplace performance, causing significant economic losses (Cashman et al. Citation2012).
Allergic conditions are exaggerated immune responses, resulting in disease outcomes including asthma and allergic contact dermatitis (ACD). These often occur in response to LMW sensitizers found in the workplace. Health experts believe that between 15% and 23% of all cases of asthma may be related to working conditions (Pralong et al. Citation2012; Dotson et al. Citation2015). Contact dermatitis (irritant and allergic) is also a common chemically-induced occupational disease, accounting for 15–20% of all occupational illnesses, with an estimated annual cost of at least $1 billion (Sasseville Citation2012). While many chemicals are known to directly induce allergic disease, there is also the potential for non-allergenic chemicals to function as adjuvants or irritants, augmenting the immune responses induced by other chemical and protein allergens.
Antimicrobials represent a broad class of LMW chemicals with the intended purpose of eliminating or controlling the growth of harmful microorganisms. Exposure to these agents can occur occupationally or via use/consumption of consumer products. The use of antimicrobial agents has been associated with an increased incidence of allergic diseases, including asthma, atopic dermatitis, and less commonly, anaphylaxis. Very diverse immunological mechanisms and mediators have been identified in the sensitization response to antimicrobials (Anderson et al. Citation2019). Specifically, common antimicrobials o-phthalaldehyde (OPA), benzalkonium chloride (BAC), didecyldimethylammonium chloride (DDAC), and triclosan (TCS) have been associated with immunological diseases (Anderson and Meade Citation2014; Anderson et al. Citation2019). Quaternary ammonium compounds (QAC) are a specific class of antimicrobials (used in sprays and wet-wipe products used to disinfect surfaces and floors) and are recognized as common occupational allergens due to their association with both contact dermatitis and occupational asthma (Bernstein et al. Citation1994; Shaffer and Belsito Citation2000; Suneja and Belsito Citation2008; Gonzalez et al. Citation2014; Anderson et al. Citation2016b; Shane et al. Citation2017; Shutty and Scheinman Citation2017). The QAC BAC is commonly associated with asthma in humans; however, it is typically classified as an irritant (or weak sensitizer) in animal studies (Manetz and Meade Citation1999). DDAC, another QAC, is a broad-spectrum bactericidal and fungicidal biocide (Skaliy et al. Citation1980). Allergic contact dermatitis and immediate-type allergic reactions caused by DDAC exposure have been reported (Dejobert et al. Citation1997; Dibo and Brasch Citation2001; Ruiz Oropeza et al. Citation2011; Mowitz and Ponten Citation2015). In animal models, DDAC has been identified as an irritant and strong T-cell-mediated sensitizer based on cellular phenotyping and the lack of identification of serum IgE (Anderson et al. Citation2016b). OPA is an aromatic dialdehyde used as a high-level antimicrobial disinfectant for medical equipment which is sensitive to normal heat or steam sterilization processes. Exposure has been associated with anaphylaxis, occupational asthma, and severe allergic reactions in humans (Sokol Citation2004; Fujita et al. Citation2007). Additionally, animal studies have identified OPA as an IgE-mediated sensitizer (Anderson et al. Citation2010).
While certain antimicrobials – including those described above – are known to induce sensitization, others such as TCS have been associated with allergic disease, though not directly sensitizing. In addition to its clinical use, TCS is used as a preservative, fungicide, and biocide in household and personal care products (Glaser Citation2004; Fang et al. Citation2010; Weatherly and Gosse Citation2017). Research suggests that TCS exposure may be at least in part responsible for recent increases in the frequency of asthma and allergic disease (Savage et al. Citation2012, Citation2014; Anderson et al. Citation2013). Additional studies have revealed that topical exposure to TCS augmented the allergic response to an experimental allergen through a thymic stromal lymphopoietin (TSLP)-mediated signaling pathway in a mouse model of asthma (Anderson et al. Citation2013; Marshall et al. Citation2015).
Exposure to antimicrobial chemicals can result in multiple hypersensitivity pathways/disease outcomes (i.e. both IgE-mediated/T-cell-mediated; asthma/allergic contact dermatitis), reflecting an increased complexity of immunological mechanisms driving these response. Further research is needed to evaluate the hazard-potential associated with antimicrobials and to fully understand the immunologic mechanisms that induce and exacerbate immune and allergic diseases. Moreover, identification of specific biomarkers would help to identify potential immune responses resulting from exposure. Ultimately, a complete understanding of mecha-nisms of allergic diseases resulting from antimicrobial exposure will allow for surveillance, proper treatment and/or prevention, while hazard identification will lead to risk assessment, which will ensure safe environments and exposure limits.
In the studies described here, gene expression profiles were examined using real-time qPCR following dermal exposure to the above-mentioned antimicrobial chemicals to identify unique profiles that could potentially aid in hazard classification and provide a better understand-ing of mechanisms involved. Based on human and animals study findings, the antimicrobials used were a weak sensitizer/irritant (BAC), IgE-mediated sensitizer (OPA), T-cell-mediated sensitizer (DDAC), or an adjuvant (TCS). Expression of cytokines and cellular mediators were then analyzed in mouse skin, draining lymph nodes (dLN), blood, and lungs after repeated chemical exposures. It is hoped the findings here will contribute to a more complete understand-ing of mechanisms of immune diseases resulting from antimicrobial exposure, and will help to ensure safe workplace environments and effective exposure limits.
Materials and methods
Animals
BALB/c mice (female, 7–8-week-old) were purchased from Taconic (Germantown, NY). Upon arrival, mice were allowed to acclimate for a minimum of 5 days. Each shipment of mice was randomly assigned to an exposure group and identified with tail markings made by a permanent marker. Mice were housed (five/cage) in ventilated plastic shoebox cages with hardwood chip bedding. Harlan NIH-31 modified 6% irradiated rodent diet and filtered tap water were available ad libitum. Housing facilities were maintained at 68–72 °F and at a 36–57% relative humidity, with a 12 hour light/dark cycle. All animal experiments were performed in the AAALAC International accredited National Institute for Occupational Safety and Health (NIOSH) animal facility in accordance with an animal protocol approved by the CDC-Morgantown Institutional Animal Care and Use Committee (IACUC).
Test chemicals
Benzalkonium chloride (BAC, CAS# 63449-41-2) and o-phthalaldehyde (OPA, CAS# 643-79-8) were purchased from Sigma (St. Louis, MO). Didecyldimethylammonium chloride (DDAC, CAS# 7173-51-5) was purchased from AKSci (Union City, CA). Triclosan (TCS, CAS# 3380-34-5) was purchased from Calbiochem (Burlington, MA). Acetone (CAS# 67-41-1) was purchased from Acros (Waltham, MA).
Chemical exposures
Mice (five/group) were exposed once per day for either 1, 2, 4, or 7 consecutive days to vehicle (acetone) or to one of three concentrations of test chemical (BAC 0.5%, 1%, 2%; OPA 0.25%, 0.5%, 0.75%; DDAC 0.125%, 0.25%, 0.5%; TCS 0.75%, 1.5%, 3%) on the dorsal surface of each ear (25 µl/ear). Concentrations were selected based on previous study findings (Anderson et al. Citation2010, Citation2016a, Citation2016b). Acetone was selected as the vehicle based on solubility and previous use in evaluations of chemical sensitization (). Expression of cytokines and cellular mediators was analyzed in the mouse skin, dLN, blood, and lung 24 hour after the final exposure (see below). Antimicrobials were selected based on their classification as irritant, IgE mediated sensitizer, T-cell-mediated sensitizer, or adjuvant (). For the sensitizing chemicals, an irritating/sensitizing and nonirritating/sensitizing concentration were included.
Table 1. Classification of antimicrobial chemicals used in this study.
Euthanasia and tissue collection
Animals were euthanized by CO2 inhalation 24 hour after the final exposure. Left and right ears, left and right auricular dLN, and lung were collected into tubes containing 500 µl RNAlater (Invitrogen, Carlsbad, CA)). Blood was collected from the abdominal aorta and placed into tubes containing 700 µL QIAzol Lysis Reagent (Qiagen, Germantown, MD)). Samples were frozen at −80 °C until processed.
RNA isolation and reverse transcription
Total RNA was isolated from the ear (RNeasy kit), dLN (miRNeasy kit), blood (miRNeasy kit), and lung (miRNeasy kit for OPA and DDAC; RNeasy kit for BAC and TCS) according to manufacturer protocols (Qiagen). A QIAcube (Qiagen) automated RNA isolation machine was used in conjunction with the specified RNA isolation kit. The concentration and purity of the isolated RNA was determined using a NanoDrop Spectrophotometer (Thermo Scientific, Waltham, MA). Reverse transcription was performed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) according to manufacturer recommendations.
Gene expression analysis
TaqMan Fast Universal PCR Master Mix (Applied Biosystems), cDNA, and gene-specific primers (TaqMan Gene Expression Assays) were combined and real-time quantitative PCR was performed according to the manufacturer’s directions. Genes tested include: S100a8, Tslp, Il13, Il33, Il4, Tnip1, Tnfaip3 (lung); S100a8, Rage, Tnip1, Tnfaip3 (blood); Tslp, Foxp3, Cdh1, Tlr4, Il4, Il13, Il22, (ear), and Ifng, Il-4, Il-5, Il10, Foxp3 (dLN). Actb was used as the reference gene. Genes were selected based on known or suspected immunological roles in the specified tissue. MicroAmp Fast Optical 96-Well Reaction Plates were analyzed in a 7500 Fast Real-Time PCR System (Applied Biosystems) according to manufacturer directions. Data was collected and represented as the relative fold-change compared to vehicle control using the cycle threshold (Ct) and the 2−ΔΔCt method.
Statistical analysis
The PCR data generated from these experiments were analyzed using SAS/STAT for Windows (v9.4) and JMP for Windows (v13). For each chemical utilized in these studies, two-way (Concentration by Day) analysis of variance was performed for each molecule using Proc Mixed in SAS. Data derived using the 2−ΔΔCt method were log-transformed prior to analysis to meet the assumption of homogeneous variance for the statistical model. Significant differences across days, and concentrations were assessed using Fishers LSD test. All differences were considered significant at p < 0.05. Heat maps were generated using JMP version 13.
Results
Antimicrobial chemical exposure increases danger signal expression
Several factors and molecular signals play a role in whether chemical exposure leads to sensitization. The two signals that are essential in order for a chemical to result in sensitization are T-cell activation and the presence of danger signals (Shane et al. Citation2019a). Thymic stromal lymphopoietin (TSLP) is a danger signal typically associated with the activation of Type 2 helper T-cell (TH2) responses. Previous work in our laboratory has shown that TCS augments the allergic response through a TSLP-mediated pathway (Marshall et al. Citation2015).
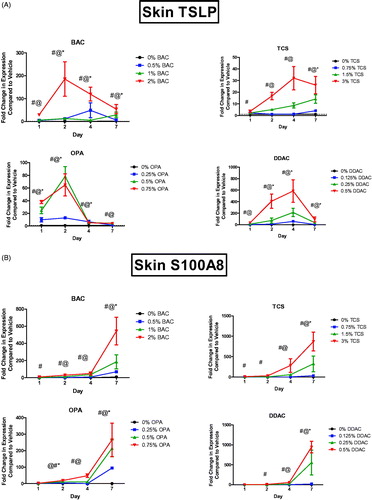
Dermal exposure to all of the antimicrobial chemicals resulted in a significant increase in Tslp mRNA expression at the site of exposure (). DDAC and TCS exposure led to a peak of Tslp expression (580 and 35 fold, respectively) after 4 days, whereas BAC and OPA exposure led to a peak (180 and 80 fold, respectively) of Tslp expression after just 2 days of exposure. For the sensitizing chemicals (DDAC and OPA), both irritating/sensitizing and nonirritating/sensitizing concentrations induced significant changes in Tslp expression. Another danger signal, S100A8, has also been identified as a factor in the adjuvant effect of TCS exposure (Marshall et al. Citation2017). S100A8 forms a heterodimer with S100A9, resulting in the danger signal protein called calprotectin. Dermal exposure to all antimicrobial chemicals resulted in an increase in S100a8 mRNA expression (). However, unlike Tslp expression, S100a8 expression continued to increase during the 7 days of exposure for all concentrations of the tested chemicals. For the sensitizing chemicals (DDAC and OPA), both irritating/sensitizing and nonirritating/sensitizing concentrations induced significant changes in S100a8 expression.
Figure 1. Increases in mRNA expression of danger signals following antimicrobial chemical exposure in the mouse skin. Fold-change in expression of (A) Tslp and (B) S100a8 following 1, 2, 4, and 7 days on exposure. Points represent mean (± SEM) of five mice/group. Low, mid and high concentration of each chemical were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations for each chemical. Dotted line represents an arbitrary value for baseline fold-change.
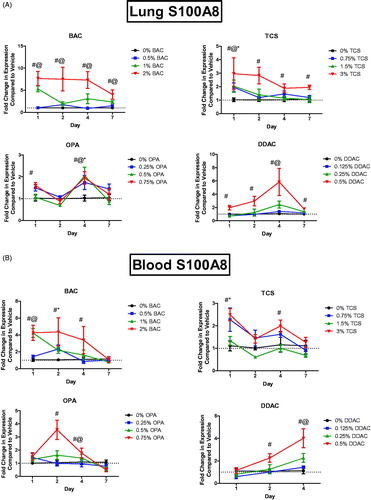
Dermal exposure to chemicals has systemic effects, including enhancing allergic responses in the lungs; thus, expression of danger signals in the lungs of mice following test agent exposure were evaluated. Dermal exposure to the antimicrobial chemicals here did not alter expression of Tslp in the lungs over two-fold (data not shown). Interestingly, the exposures did increase S100a8 expression in the lungs (). Dermal exposure to BAC led to a statistically significant increase in S100a8 expression at mid and high concentrations at all days (). Exposure to the highest TCS concentration significantly increased S100a8 expres-sion in the lungs at all days and just 1 days of TCS exposure increased S100a8 at all concentrations at that site. Exposure to DDAC for 1, 2, 4, and 7 days increased S100a8 expression in the lungs at the highest concentration, and at the mid concentration after 4 days of exposure. Similar responses were observed in the blood for all chemicals evaluated (). Blood following 7 days of DDAC exposure was not evaluated due to equipment failure.
Figure 2. Increases in mRNA expression of S100a8 in the (A) blood (A) and lung (B) following antimicrobial chemical exposure on the mouse skin following 1, 2, 4, or 7 days of exposure. Points represent mean (± SEM) of five mice/group. Low, mid and high concentration of each chemical were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations for each chemical. Dotted line represents an arbitrary value for baseline fold-change.
OPA exposure increases TH2 cytokine levels
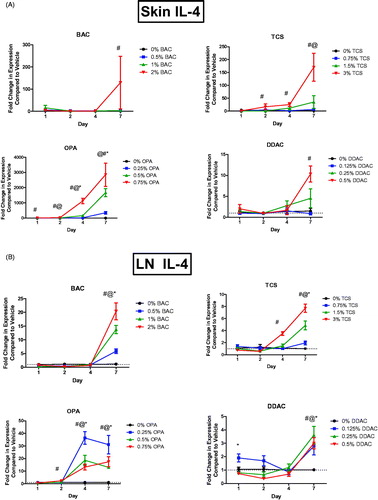
Interleukin (IL)-4 is a central cytokine in TH2 immune responses. Expression of Il4 mRNA was assessed in the skin, dLN, and lungs following dermal exposure to antimicrobial chemicals. OPA exposure significantly increased Il4 mRNA expression in the skin at the low, mid, and high concentrations after 4 and 7 days of exposure, in the skin at the mid and high concentrations after 2 days of exposure, and in the skin at the high concentration after just 1 days of exposure (). Seven days of BAC or DDAC exposure at their highest concentrations led to significant increases in Il4 in the skin. Exposure to the highest concentration of TCS for 2, 4, and 7 days also led to statistically significant increases of Il4 in the skin. Il4 was increased in the dLN following 2 days of the highest OPA exposure and following 4 and 7 days of all tested OPA levels (). Seven days of TCS exposure at all concentrations resulted in a significant increase in Il4 in the dLN and 4 days of the highest concentration of TCS led to a significant increase in Il4. Exposure to BAC or DDAC for 7 days resulted in statistically significant increases in Il4 expression at all test concentrations.
Figure 3. Increases in mRNA expression of Il4 following antimicrobial chemical exposure on the mouse skin. Fold-change in expression of Il4 in the (A) skin and (B) dLN following 1, 2, 4, and 7 days of exposure. Points represent mean (± SEM) of five mice/group. Low, mid and high concentration of each chemical were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations for each chemical. Dotted line represents an arbitrary value for baseline fold-change.
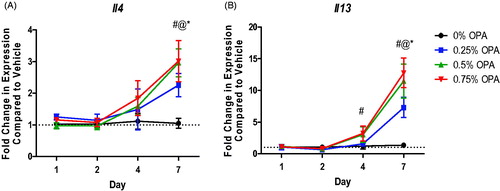
Dermal exposure to OPA for 7 days increased Il4 expression in the lungs at all test levels (). IL-13 is another cytokine central to the TH2 immune response. Dermal exposure to OPA for 7 days also increased Il13 expression in the lungs at all test concentrations (). None of the other antimicrobials altered expression of Il4 or Il13 in the lung (data not shown).
Figure 4. Increases in mRNA expression of TH2 cytokines in lungs following antimicrobial chemical exposure on mouse skin. Fold-change in expression of (A) Il4 and (B) Il13 in the mouse lung following 1, 2, 4, and 7 days of exposure. Points represent mean (± SEM) of five mice/group. Low, mid and high concentrations were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations. Dotted line represents an arbitrary value for baseline fold-change.
OPA exposure influences the regulatory response
Regulatory T-cells (Treg) have previously been found to be involved in the immune response to chemical exposure (Long et al. Citation2016). FoxP3, the critical transcription factor for Treg cell development was assessed in the skin following the chemical exposures. Dermal exposure to BAC, OPA, and DDAC for 7 days increased Foxp3 expression in skin at all tested concentrations (). Exposure to TCS for 7 days at the highest concentration also increased the expression. OPA exposure also significantly increased Foxp3 expression after 4 days of all test concentrations and after 2 days of the mid and high concentrations. Similar, but less dramatic results, were obtained in the dLN (Supplemental Figure 3). Expression of Il10, a cytokine produced by Treg cells and involved in immune regulation, was assessed in the dLN following dermal chemical exposure. OPA exposure significantly increased Il10 expression in the dLN after 7 days of all test levels and after 4 days of exposure to the highest concentration (). Exposure to DDAC for 2 days significantly decreased Il10 expression in the LN at the mid and high concentrations.
Figure 5. Increases in mRNA expression of regulatory genes and TH1 cytokines following anti-microbial chemical exposure in the mouse skin following 1, 2, 4, and 7 days of exposure. Fold-change in the expression of (A) Foxp3 in skin and (B) Il10 in dLN. Points represent mean (± SEM) of five mice/group. Low, mid and high concentration of each chemical were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations for each chemical. Dotted line represents an arbitrary value for baseline fold-change.
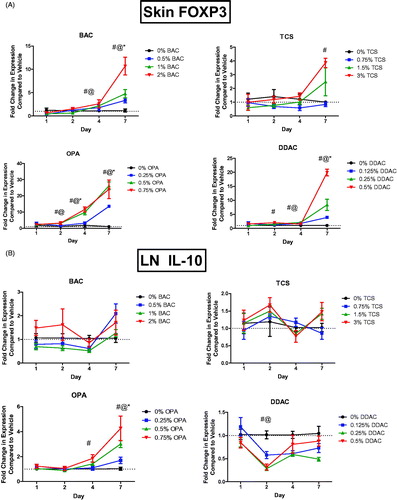
Exposure to TCS uniquely alters Tlr4 expression in skin
Understanding the mediators involved in chemical sensitization and immune responses after dermal chemical exposure is critical in identifying the differences between sensitizers, irritants, and adjuvants. Toll-like receptor 4 (TLR4) has previously been identified to play a role in the immune response to TCS (Marshall et al. Citation2017). Tlr4 expression was therefore assessed in the skin following the dermal chemical exposures. Exposure to 7 days of TCS at the highest concentration increased Tlr4 expression in the skin (). However, no other chemical significantly increased this expression.
Figure 6. Unique changes in mRNA expression of genes following antimicrobial exposure. Fold-change in expression of (A) Tlr4 and (B) Cdh1 in the skin following 1, 2, 4 and 7 days of exposure. Points represent mean (± SEM) of five mice/group. Low, mid and high concentration of each chemical were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations for each chemical.
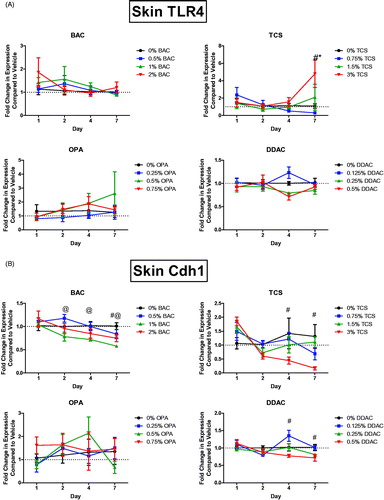
Decreases in E-cadherin expression after antimicrobial exposures
E-cadherin, a cellular adhesion molecule highly expressed in the skin and associated with innate lymphoid cells (ILC), has been shown to suppress TH2 cytokine production by Type 2 innate lymphoid cells (ILC2) through ligation with the co-inhibitory receptor killer-cell lectin like receptor G1 (KLRG1). E-cadherin is also associated with inflammatory skin diseases such as atopic dermatitis (Salimi et al. Citation2013). Exposure for 4 and 7 days to TCS and DDAC at the highest concentration decreased E-cadherin (Cdh1) expression in the skin (). Cdh1 expression was also decreased following exposure to the high concentration of BAC at Day 7 and the mid concentration at Days 2, 4, and 7. OPA exposure did not alter Cdh1 expression.
Chemical exposure increases ifnγ and Il22 expression
Interferon (IFN)-γ is the cytokine central to Type 1 helper T-cell (TH1) responses. OPA exposure significantly increased Ifnγ mRNA expression in the dLN after just 2 and 4 days of exposure at all test concentrations, and after 1 days of exposure to the highest concentration (). Exposure to BAC significantly increased Ifng expression after 1 days of exposure to the high concentration. Interestingly, expression was significantly decreased (mid and high levels) after 7 days of exposure. Exposure to DDAC significantly decreased Ifng expression for all concentrations 2 days post-exposure; this decrease persisted until 7 days post-exposure. No changes in Ifnγ expression were seen following exposure to TCS.
Figure 7. Increase in mRNA expression of cytokines following chemical exposure. Fold-change in expression of (A) Ifnγ in dLN and (B) Il22 in skin following 1, 2, 4, and 7 days of exposure. Points represent mean (± SEM) of five mice/group. Low, mid and high concentration of each chemical were evaluated. Statistical significance (p < 0.05) compared to 0% is indicated at each timepoint for *low, @mid, and #high concentrations for each chemical. Dotted line represents an arbitrary value for baseline fold-change.
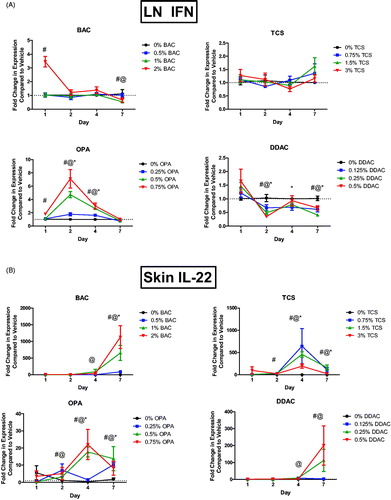
IL-22 is a cytokine expressed by Type 17 helper T-cells (TH17). Il22 mRNA expression was assessed in the skin following dermal exposure to chemicals. All chemicals resulted in significant increases in Il22 expression after exposure. Exposure to TCS for 4 or 7 days significantly increased Il22 expression at all concentrations, with a peak increase after 4 days of exposure (). Dermal exposure to BAC or DDAC increased Il22 expression after 4 and 7 days, but the peak increase occurred after 7 days of exposure (). OPA significantly increased expression of Il22 by 1 days post-exposure, with a peak increase at 4 days that persisted until 7 days post-exposure.
Discussion
Occupational immune diseases are a serious health burden. Thus, the ability to identify chemical hazards and understand immunological mechanisms of disease is critical. Numerous studies have shown that exposure to chemicals can drive the development of allergic diseases, either directly, or indirectly. The results from this study identify unique expression profiles of select cytokines and cellular mediators between different classes of antimicrobial chemicals (Supplemental Figures 1–4). For this study, the representative chemicals were classified based on a specific type of immune response (). It is important to note that this classification scheme represents a simplified approach, as the categorization of immune responses is more complex than what is presented. The wide spectrum of clinical symptoms associated with most of the investigated chemicals suggest mixed immune responses and hypersensitivity classifications that may not be mutually exclusive. However, in the studies described here, irritating/sensitizing and nonirritating/sensitizing concentrations were evaluated for the sensitizing chemicals. Although direct comparisons may be difficult to make due to differences in exposure concentration and potency, unique chemical trends can still be identified.
For this study, OPA was classified as an irritant and IgE-mediated sensitizer based on findings in previous human and animal studies. Studies conducted in our laboratory found that exposure to OPA significantly increased ear swelling and lymphocyte proliferation in the dLN when evaluated in the local lymph node assay (LLNA) (Anderson et al. Citation2010). In addition, 0.5% OPA exposure increased serum IgE along with IL-4 expression at both the gene and protein level in the dLN . Consistent with these findings which further support its TH2 classification, OPA was identified to induce early and persistent expression of Il4, following dermal exposure to multiple concentrations, in the ear and dLN. Interestingly and uniquely, OPA also induced expression of Il4 and Il13 in the lungs. While involvement of the skin is recognized in the development of dermal sensitization, it has recently been implicated in the development of systemic sensitization leading to elicitation responses at various sites in the body, including the respiratory tract (Bello et al. Citation2007). This has been demonstrated in animal studies involving both protein and chemical allergens (Zhang et al. Citation2002; Herrick et al. Citation2003; Redlich Citation2010). In a workplace setting, the like-lihood of dermal contact with low molecular weight (LMW) chemicals is high, further support-ing the idea that dermal exposures may lead to respiratory allergic disease. In addition to their probability of occurrence in the workplace, LMW chemical skin exposures also have a potential for higher dose-delivery in comparison to inhalation exposures (Bello et al. Citation2007). These findings support a very interesting connection and potential discriminating feature between respiratory and contact sensitizers and further suggests that sensitization via the skin may be important for respiratory allergic outcomes.
The development of chemical allergy is immunologically complex and our understanding of the mechanisms driving these responses continue to evolve. Research suggests that dosage, exposure duration, and route of exposure may all influence/alter a developing immune response. Adding to the complexity of defining immune responses is the increased understanding that the development of hypersensitivity responses is not as divergent nor categorical as once thought. It is generally accepted that sensitizing chemicals that induce TH2 and/or IgE-mediated responses will tend to initiate expression of TH2 cytokines and suppress those commonly associated with TH1 effector responses (Dearman and Kimber Citation1991; Kimber and Dearman Citation1992; Anderson et al. Citation2011). Despite this, the specific chemical properties that define each type of sensitizer have not been identified.
In contrast to OPA, DDAC has been identified as an irritant and strong T-cell-mediated sensitizer in mice. Exposure to 0.5% DDAC was previously shown to result in increased ear swelling with a significant increase in lymphocyte proliferation at 0.25%, when evaluated in the LLNA but did not increase serum IgE levels (Anderson et al. Citation2016b). Although classified as a T-cell sensitizer in these studies, here, DDAC induced significant expression of Il4 in the skin and dLN following 7 days of exposure. This is consistent with increases previously identified in IL-4 expression following DDAC exposure at both the transcript and protein level (Shane et al. Citation2019b). It is possible that this early IL-4 production is due to innate mediators such as ILC2, and may contribute to a mixed-type response. Additional studies conducted in our laboratory have also demonstrate that extended dermal exposure to QAC (14 days+) induced production of serum and local IgE (Shane et al. Citation2017, Citation2019b). Interestingly, significant decreases in mRNA expression of the TH1 cytokine Ifng that persisted throughout the course of the study, were identified following DDAC exposure. In contrast, OPA significantly increased expression of Ifng. BAC was included in the present study as an irritant/weak sensitizer based on findings in human and animal studies (Manetz and Meade Citation1999; Isaac and Scheinman Citation2017). For BAC, an immediate increase in Ifng was observed, but this did not persist after the 1 days timepoint. These findings further demonstrate the induction of mixed responses by LMW chemicals and support the impact of exposure duration on the subsequent immunological response.
Although chemicals can directly affect the immune system and subsequently influence allergic disease, there is also the potential for indirect affects through mechanisms involving irritation/inflammation and adjuvancy. While associated with allergic disease in humans, in vivo hazard identification models have not identified TCS as a sensitizer or irritant (Anderson et al. Citation2016a). However, dermal TCS exposure has been shown to augment the allergic response to an experimental allergen in a mouse model of asthma; thus, TCS was classified as an adjuvant for this study (Anderson et al. Citation2013, Citation2016a). While TCS exposure induced expression profiles that were similar to the sensitizers (Tslp, S1000a8, Il22, Il4, Foxp3, Cdh1), unique to exposure were increases in expression of Tlr4 in the skin. A similar finding has been previously described in our laboratory (Marshall et al. Citation2017). More specifically, in the current and previous studies, TCS was seen to induce abundant expression of S100A8 in the skin; this protein acts as an endogenous ligand for the intracellular signaling receptor TLR4, which is important for activation of the innate immune system.
The skin serves as a protective layer for our bodies from the outside environment. As the largest organ in the body, the skin is an extremely important player in relation to allergic disease. The presence of multiple innate immune factors including leukocytes, complement factors, antimicrobial peptides, and pattern recognition receptors allow the skin to be a site of immune surveillance and tolerance yet these factors may also contribute to the development of allergic disease (Bangert et al. Citation2011). The initiation of sensitization begins with exposure and antigen recognition. In order to gain access to immune cells responsible for commencing sensitization, allergens must penetrate the epithelium. In the skin, LMW chemical allergens may be absorbed through the stratum corneum, hair follicles, and sebaceous glands (Nayak et al. Citation2014), accessing internal cells without physical alteration of the epithelium due to their small size. A widely-accepted concept explaining immunogenicity of LMW chemical allergens involves a haptenation step, i.e. combining with and altering a self-protein, causing an allergic response following presentation by antigen-presenting cells (Landsteiner and Jacobs Citation1935; Kohler et al. Citation1995; Chipinda et al. Citation2011). Because TCS is not a sensitizing chemical, it does not form a hapten. This lack of reactivity is one potential explanation of why TLR signaling may be unique to this class of chemical.
Recently, the emergence of many novel cellular subsets and molecules involved in immunological responses has occurred, shedding light on unexplored realms of the immune system and their potential involvement in a variety of disease states, including allergic disease (Shane et al. Citation2019a). In accordance with these developments, further investigation into these responses demonstrated that DDAC induced high levels of expression of the TH2-skewing cytokine Tslp, which has been shown to activate ILC2 in the skin (Kim et al. Citation2013). ILC2 are a subset of innate lymphocytes that lack rearranged antigen-specific receptors and produce Type 2 cytokines. ILC2 have recently emerged as important mediators of allergic disease (Cosmi et al. Citation2017). Following DDAC exposure, ILC2 in the skin were rapidly activated, and their activation coincided with the production of Type 2 cytokines in the absence of T-cells; this provided a potential mechanism for the initiation of the mixed-type allergic response (Shane et al. Citation2019b). E-cadherin (cellular adhesion molecule highly expressed in skin) has been associated with the suppression of ILC2 function via inhibition of their TH2 cytokine production (Salimi et al. Citation2013). Here, all chemicals except OPA decreased Cdh1 expression in the skin. However, as numerous signals can drive ILC2 regulation and activation (Dahlgren and Molofsky Citation2018), it is possible that the ILC2 contribute to early TH2 cytokine production in the skin following exposure to OPA.
Another newly characterized helper T-cell subset thought to play a role in allergic disease is the TH22 subset. These cells are identified by their production of IL-22 in the absence of IFNγ, IL-4, and IL-17, and are thought to contribute to host defense against microbial pathogens and promote tissue repair or remodeling (Fujita Citation2013). In the skin, IL-22 plays a major role in home-ostasis and pathogenesis of skin diseases by inducing keratinocyte proliferation and epidermal hyperplasia, inhibiting terminal differentiation of keratinocytes, and promoting the production of antimicrobial proteins. While information about the role of TH22 cells in chemical allergy in the skin is lacking, they have been implicated in the pathogenesis of inflammatory skin disorders such as psoriasis and atopic dermatitis (AD) (Mirshafiey et al. Citation2015). Additionally, IL-22 levels were found to be increased in the skin of patients with AD, ACD, and allergic asthma (Jia and Wu Citation2014) and IL-22 has been suggested as a potential biomarker for allergic disease (Zissler et al. Citation2016). In the current study, all chemicals induced Il22 expression. This outcome supports the need for additional research investigating the role of this cytokine in immunological disease.
Our laboratory has previously shown that chemical sensitizers affect Treg cells (Long et al. Citation2016). Following single dermal exposure to the known asthmogen toluene diisocyanate (TDI), the LN Treg cell population expanded significantly at 4, 7, and 9 days. Additionally, Treg cells isolated from TDI-sensitized mice were significantly more suppressive compared with their control cell counterparts, further supporting a functional role for Treg cells during sensitization. While here all the tested chemicals induced expression of Foxp3 in the ear, only OPA increased Il10 in the dLN. OPA also induced the largest and earliest peak expression of Foxp3 in the skin. Although the number of Treg cells was not determined in the present study, the increases in gene expression in addition to our previous findings support a direct role for Treg cells in chemical sensitization. The collection of data regarding Treg cells and chemical allergy is growing but remains limited. In order to elucidate the immunologic mechanisms involved in LMW chemical sensitization, the biological functions of pertinent immune cell subsets, such as Treg cells need to be delineated.
It has long been recognized that the presence of foreign antigens alone is insufficient to generate immune responses: activation of the innate immune system is also required. Research is continuing to bring to light the importance of such “danger signals” in allergic sensitization. In this study, all chemicals induced expression of the danger signal, Tslp in the skin. The highest Tslp expression was identified after DDAC exposure, at later timepoints. OPA-induced elevations in Tslp expression were generally lower compared to the other sensitizers and had almost returned to baseline by 4 days. However, since different concentrations were compared for each chemical, it is difficult to draw specific conclusions, but for the sensitizing chemicals both irritant and nonirritating sensitizing concentrations induced increases in expression. Additionally, S100a8 expression in the skin peaked (all concentrations) at Day 7 and was signifi-cantly elevated for at least one concentration by Day 1 or 2 post-exposure. It is important to note that since the lungs were not perfused prior to collection, there is potential for contribution from the blood in the lung S100A8 response. This is reflected by their similar expression patterns. Neutrophils are known to express high levels of S100a8, and therefore circulating neutrophils could be contributing to the high levels of expression observed in multiple tissues. However, these finding suggest that differential expression patterns (early vs. later) could be a potential way to distinguish immunological mechanisms of disease.
The burden of occupational allergic disease is widespread. Occupational allergic conditions are multifactorial and are the result of complicated immunologic events. The results from the studies here suggest unique gene expression profiles are detectable following exposure to various antimicrobial chemicals, indicating potential utility as biomarkers in future risk assessment. Likewise, the data from these studies suggest a high throughput gene expression kinetics screen which can potentially serve as a basis for future investigational studies. The findings also support the need for additional research into mechanisms of disease, the mediators involved, and identification of potential biomarkers. Future studies will need to focus on additional earlier timepoints and evaluation of additional classes of chemicals. A complete understanding of the mechanisms of immune and allergic diseases resulting from LMW chemical exposure will allow for surveillance, proper treatment and/or prevention, while hazard identification will lead to risk assessment, which will ensure safe environments and effective exposure limits.
Supplemental Material
Download MS Word (347.5 KB)Disclosure statement
The authors declare no conflicts of interest. The authors alone are responsible for the content of this manuscript. The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the National Institute for Occupational Safety and Health, Centers for Disease Control and Prevention.
Additional information
Funding
References
- Anderson S, Franko J, Kashon M, Anderson K, Hubbs A, Lukomska E, Meade B. 2013. Exposure to triclosan augments the allergic response to ovalbumin in a mouse model of asthma. Toxicol Sci. 132(1):96–106.
- Anderson S, Meade B. 2014. Potential health effects associated with dermal exposure to occupational chemicals. Environ Health Insights. 8(Suppl 1):51–62.
- Anderson S, Meade B, Long C, Lukomska E, Marshall N. 2016a. Investigations of immunotoxicity and allergic potential induced by topical application of triclosan in mice. J. Immunotoxicol. 13(2):165–172.
- Anderson S, Shane H, Long C, Lukomska E, Meade B, Marshal N. 2016b. Evaluation of the irritancy and hypersensitivity potential following topical application of didecyldimethylammonium chloride. J. Immunotoxicol. 13(4):557–566.
- Anderson S, Siegel P, Meade B. 2011. The LLNA: A brief review of recent advances and limitations. J Allergy. 2011:1–10.
- Anderson S, Umbright C, Sellamuthu R, Fluharty K, Kashon M, Franko J, Jackson L, Johnson V, Joseph P. 2010. Irritancy and allergic responses induced by topical application of o-phthalaldehyde. Toxicol Sci. 115(2):435–443.
- Anderson S, Weatherly L, Shane H. 2019. Contribution of antimicrobials to the development of allergic disease. Curr Opin Immunol. 60:91–95.
- Bangert C, Brunner P, Stingl G. 2011. Immune functions of the skin. Clinics Dermatol. 29(4):360–376.
- Bello D, Herrick C, Smith T, Woskie S, Streicher R, Cullen M, Liu Y, Redlich C. 2007. Skin exposure to isocyanates: Reasons for concern. Environ Health Perspect. 115(3):328–335.
- Bernstein J, Stauder T, Bernstein D, Bernstein I. 1994. A combined respiratory and cutaneous hypersensitivity syndrome induced by work exposure to quaternary amines. J Allergy Clin Immunol. 94:257–259.
- Cashman M, Reutemann P, Ehrlich A. 2012. Contact dermatitis in the United States: Epidemiology, economic impact, and workplace prevention. Dermatol Clin. 30(1):87–98.
- Chipinda I, Hettick J, Siegel P. 2011. Haptenation: Chemical reactivity and protein binding. J Allergy. 2011:1–11.
- Cosmi L, Liotta F, Maggi L, Annunziato F. 2017. Role of Type 2 innate lymphoid cells in allergic diseases. Curr Allergy Asthma Rep. 17(10):66.
- Dahlgren M, Molofsky A. 2018. All along the watchtower: Group 2 innate lymphoid cells in allergic responses. Curr Opin Immunol. 54:13–19.
- Dearman R, Kimber I. 1991. Differential stimulation of immune function by respiratory and contact chemical allergens. Immunology. 72(4):563–570.
- Dejobert Y, Martin P, Piette F, Thomas P, Bergoend H. 1997. Contact dermatitis from didecyldi-methylammonium chloride and bis-(aminopropyl)-lauryl amine in a detergent-disinfectant used in hospital. Contact Dermatitis. 37(2):95–96.
- Dibo M, Brasch J. 2001. Occupational allergic contact dermatitis from N,N-bis(3-amino-propyl)-dodecylamine and dimethyldidecylammonium chloride in two hospital staff. Contact Dermatitis. 45(1):40–40.
- Dotson G, Maier A, Siegel P, Anderson S, Green B, Stefaniak A, Codispoti C, Kimber I. 2015. Setting occupational exposure limits for chemical allergens – understanding the challenges. J Occup Environ Hyg. 12(sup1):S82–S98.
- Fang J, Stingley R, Beland F, Harrouk W, Lumpkins D, Howard P. 2010. Occurrence, efficacy, metabolism, and toxicity of triclosan. Environ Carcinogen Ecotoxicol Rev. 28(3):147–171.
- Fujita H. 2013. The role of IL-22 and TH22 cells in human skin diseases. J Dermatol Sci. 72(1):3–8.
- Fujita H, Sawada Y, Ogawa M, Endo Y. 2007. Health hazards from exposure to o-phthalaldehyde, a disinfectant for endoscopes, and preventive measures for health care workers. SanEiShi. 49(1):1–8.
- Glaser A. 2004. The ubiquitous triclosan: A common antibacterial agent exposed. Pesticides You. 24:12–17.
- Gonzalez M, Jegu J, Kopferschmitt M, Donnay C, Hedelin G, Matzinger F, Velten M, Guilloux L, Cantineau A, de Blay F. 2014. Asthma among workers in healthcare settings: Role of disinfection with quaternary ammonium compounds. Clin Exp Allergy. 44(3):393–406.
- Herrick C, Xu L, McKenzie A, Tigelaar R, Bottomly K. 2003. IL-13 is necessary, not simply sufficient, for epicutaneously-induced TH2 responses to soluble protein antigen. J Immunol. 170(5):2488–2495.
- Isaac J, Scheinman P. 2017. Benzalkonium chloride: An irritant and sensitizer. Dermatitis. 28(6):346–352.
- Jia L, Wu C. 2014. The biology and functions of TH22 cells. Adv Exp Med Biol. 841:209–230.
- Kim B, Siracusa M, Saenz S, Noti M, Monticelli L, Sonnenberg G, Hepworth M, van Voorhees A, Comeau M, Artis D. 2013. TSLP elicits IL-33-independent innate lymphoid cell responses to promote skin inflammation. Sci Transl Med. 5:170ra116.
- Kimber I, Dearman R. 1992. The mechanisms and evaluation of chemically-induced allergy. Toxicol. Lett. 64–65:79–84.
- Kohler J, Martin S, Pflugfelder U, Ruh H, Vollmer J, Weltzien H. 1995. Cross-reactive trinitro-phenylated peptides as antigens for Class II major histocompatibility complex-restricted T-cells and inducers of contact sensitivity in mice. Limited T-cell receptor repertoire. Eur J Immunol. 25(1):92–101.
- Landsteiner K, Jacobs J. 1935. Studies on the sensitization of animals with simple chemical compounds. J Exp Med. 61(5):643–656.
- Long C, Marshall N, Lukomska E, Kashon M, Meade B, Shane H, Anderson S. 2016. A role for regulatory T-cells in a murine model of epicutaneous toluene diisocyanate sensitization. Toxicol Sci. 152(1):85–98.
- Manetz T, Meade B. 1999. Development of a combined irritancy/phenotypic analysis assay for the identification and differentiation of chemicals with the potential to elicit irritation, IgE-mediated, or T-cell-mediated hypersensitivity responses. Am J Ind Med. 36(S1):136–138.
- Marshall N, Lukomska E, Long C, Kashon M, Sharpnack D, Nayak A, Anderson K, Meade B, Anderson S. 2015. Triclosan induces thymic stromal lymphopoietin in skin promoting TH2 allergic responses. Toxicol Sci. 147(1):127–139.
- Marshall N, Lukomska E, Nayak A, Long C, Hettick J, Anderson S. 2017. Topical application of anti-microbial chemical triclosan induces immunomodulatory responses through the S100A8/A9-TLR4 pathway. J Immunotoxicol. 14(1):50–59.
- Mirshafiey A, Simhag A, El Rouby N, Azizi G. 2015. TH22 cells as a new player in chronic inflammatory skin disorders. Int J Dermatol. 54(8):880–888.
- Mowitz M, Ponten A. 2015. Foot dermatitis caused by didecyldimethylammonium chloride in a shoe refresher spray. Contact Dermatitis. 73(6):374–376.
- Nayak A, Hettick J, Siegel P, Anderson S, Long C, Green B, Beezhold D. 2014. Toluene diisocyanate (TDI) disposition and co-localization of immune cells in hair follicles. Toxicol Sci. 140(2):327–337.
- Pralong J, Cartier A, Vandenplas O, Labrecque M. 2012. Occupational asthma: New low-molecular-weight causal agents, 2000–2010. J Allergy. 2012:1–10.
- Redlich C. 2010. Skin exposure and asthma: Is there a connection? Proc Am Thor Soc. 7(2):134–137.
- Ruiz Oropeza A, Fischer Friis U, Duus Johansen J. 2011. Occupational contact urticaria caused by didecyl dimethyl ammonium chloride. Contact Dermatitis. 64(5):297–298.
- Salimi M, Barlow J, Saunders S, Xue L, Gutowska-Owsiak D, Wang X, Huang L, Johnson D, Scanlon S, McKenzie A, et al. 2013. A role for IL-25 and IL-33-driven Type-2 innate lymphoid cells in atopic dermatitis. J Exp Med. 210(13):2939–2950.
- Sasseville D. 2012. Acrylates in contact dermatitis. Dermatitis. 23(1):6–16.
- Savage J, Johns C, Hauser R, Litonjua A. 2014. Urinary triclosan levels and recent asthma exacerbations. Ann Allergy Asthma Immunol. 112(2):179–181.
- Savage J, Matsui E, Wood R, Keet C. 2012. Urinary levels of triclosan and parabens are associated with aeroallergen and food sensitization. J Allergy Clin Immunol. 130(2):453–460.
- Shaffer M, Belsito D. 2000. Allergic contact dermatitis from glutaraldehyde in healthcare workers. Contact Dermatitis. 43(3):150–156.
- Shane H, Long C, Anderson S. 2019a. Novel cutaneous mediators of chemical allergy. J Immunotoxicol. 16(1):13–27.
- Shane H, Lukomska E, Kashon M, Anderson S. 2019b. Topical application of the quaternary ammonium compound didecyldimethylammonium chloride activates Type 2 innate lymphoid cells and initiates a mixed-type allergic response. Toxicol Sci. 168(2):508–518.
- Shane H, Lukomska E, Stefaniak A, Anderson S. 2017. Divergent hypersensitivity responses following topical application of the quaternary ammonium compound, didecyldimethyl-ammonium bromide. J Immunotoxicol. 14(1):204–214.
- Shutty B, Scheinman P. 2017. Occupationally-induced allergic contact dermatitis to aerosolized quaternary ammonium compounds. Dermatitis. 28(6):369.
- Skaliy P, Thompson T, Gorman G, Morris G, McEachern H, Mackel D. 1980. Laboratory studies of disinfectants against Legionella pneumophila. Appl Environ Microbiol. 40(4):697–700.
- Sokol W. 2004. Nine episodes of anaphylaxis following cystoscopy caused by Cidex OPA (o-phthalaldehyde) high-level disinfectant in four patients after cytoscopy. J Allergy Clin Immunol. 114(2):392–397.
- Suneja T, Belsito D. 2008. Occupational dermatoses in healthcare workers evaluated for suspected allergic contact dermatitis. Contact Dermatitis. 58(5):285–290.
- Weatherly L, Gosse J. 2017. Triclosan exposure, transformation, and human health effects. J Toxicol Environ Health. 20(8):447–469.
- Zhang X, Murray D, Lewis D, Siegel P. 2002. Dose-response and time course of specific IgE and IgG after single and repeated topical skin exposure to dry trimellitic anhydride powder in a Brown Norway rat model. Allergy. 57(7):620–626.
- Zissler U, Esser-von Bieren J, Jakwerth C, Chaker A, Schmidt-Weber C. 2016. Current and future biomarkers in allergic asthma. Allergy. 71(4):475–494.