Abstract
The commercial bumble bee, Bombus impatiens, has been used in Maine lowbush blueberry since the late 1990s. Studies conducted in 1995–1998 and published in 2001 showed that B. impatiens was a good pollinator of lowbush blueberry. Stocking density was estimated at ¾–1 quad (set of four, 250-bee colonies) per acre. Subsequent research reported here has shown that on an individual bee basis, B. impatiens is 2.5 times more efficient per floral visit in placing pollen on the stigma. Foraging behavior suggests that bumble bees may be more consistent at producing outcrossing among clones than honey bees, although individual honey bees are more likely to be floral constant than bumble bees. By 2004, some blueberry growers questioned the efficacy of commercial bumble bees. To address these concerns, a three-year validation experiment was conducted from 2005–2007. Seven to nine isolated individual small blueberry fields (2–25 acres) were selected each year to compare bumble bees with honey bees or no commercial pollinators (n = 3 to 5 fields per treatment) It was verified that 2.5 quads/ha was a suitable stocking density and did not differ in pollination level from 7.5–10 honey bee hives per ha. A central place foraging behavior could not be demonstrated with bumble bees in these smaller fields. It was established that as the foraging force of bumble bees increased, which varied between hives and fields, better fruit set resulted.
INTRODUCTION
Commercial production of lowbush blueberry, Vaccinium angustifolium Ait., requires high densities of bees for adequate pollination (CitationDelaplane and Mayer, 2000; CitationYarborough, 2006). Prior to the 1960s, most pollination in lowbush blueberry fields was dependent upon native bees (CitationBoulanger et al., 1967). In Maine, over the past 20 years, we have collected more than 75 species of native bee associated with lowbush blueberry in bloom out of a verified taxonomic collection of 217 bee species for the state (Drummond et al., unpublished University of Maine museum notes). The most abundant native bee taxa associated with lowbush blueberry bloom are members of the families Andrenidae, Halictidae, and Megachilidae (CitationBoulanger et al., 1967; CitationFinnamore and Neary, 1978; CitationStubbs et al., 1992; CitationDrummond and Stubbs, 1997; CitationJavorek et al., 2002), although bumble bees (of the genus Bombus) are also common but mostly as queens during bloom (CitationStubbs et al., 1992).
Honey bees have become the pollinator of choice since the 1960s in lowbush blueberry (CitationKarmo, 1957, Citation1958) and in Maine more than 70,000 colonies are moved into the state for blueberry pollination each year (CitationDrummond, 2002). The number of honey bee colonies in the U.S. dropped from 4.5 million managed colonies in 1980 to 2.4 million colonies in 2005 (CitationNational Research Council, 2007). A national survey on winter losses over 2008–2009 showed that beekeepers lost 29% of their hives over the winter (CitationVan Engelsdorp et al., 2010), but even though the percentage of hives that were lost to Colony Collapse Disorder (CCD) decreased compared to previous years, there were still significant losses. Another survey in the mid-Atlantic region of the Northeast indicated that winter losses since 2000 have been roughly 30% per year (CitationBurdick and Caron, 2007). Varroa mites along with high viral incidences are recognized as a major cause of these heavy annual losses and possibly CCD (CitationCox-Foster et al., 2007), although pesticide exposure is frequently cited as a possible contributing factor (CitationNational Research Council, 2007).
It was the decline of honey bee colonies in the U.S. and the resulting fear of blueberry growers in Maine that they would not have a sufficient supply that prompted a study in the 1990s to assess the practical and economic utility of the commercially reared bumble bee, Bombus impatiens, for pollination of lowbush blueberry in Maine. We conducted a four-year study from 1995–1998 in central coastal Maine and showed that bumble bees at the stocking rate of 3–4 colonies (250 bee colonies) per acre provided equivalent pollination and yields to fields solely stocked with honey bees at a rate of 3 hives/acre (CitationStubbs and Drummond, 2001). The yields in those years, compared to the previous six-year average, ranged from 96–118% for the bumble bee stocked fields and 75–104% for the honey bee stocked fields. This research led to an extension fact sheet on the use of commercial bumble bees for lowbush blueberry pollination (CitationStubbs et al., 2001) and a dramatic acceptance and use of bumble bees. However, by 2004 several Maine lowbush blueberry growers questioned the efficacy of bumble bees and so a three-year validation study was initiated from 2005–2007. This article reports on the results of this study and an additional supporting study conducted in 2009 and 2010.
MATERIALS AND METHODS
Two studies were designed to better assess the efficacy of bumble bees for pollination of lowbush blueberry. A three-year colony stocking rate study was conducted in 2005–2007. In 2009 and 2010, flower fidelity of honey bees and bumble bees was compared in two blueberry fields in Jonesboro, Maine.
Stocking Density 2005–2007
In each of the three years, isolated commercial blueberry fields in Maine surrounded by forested habitat were selected and they were stocked with either commercial bumble bees at a rate of 10 colonies/ha or commercial honey bee colonies at 10 hives/ha. In 2005, the study was centered in Centerville and Columbia, Washington Co. Eight fields were selected. Three fields were stocked with commercial honey bees: 0.8 ha field with 8 hives, 0.9 ha field with 9 hives, and a 3.4 ha field with 32 hives. Four fields were selected for stocking with bumble bees: 0.8 ha field with 8 colonies or 2 quads (a quartet of colonies), 2.3 ha field with 6 quads, 2.4 ha field with 6 quads, and a 2.7 ha field with 7 quads. In addition, a single 1.3 ha field was selected that had no managed bees and relied solely on wild, native bees for pollination. In 2006, the study was conducted in Union, Jefferson, Appleton, and Washington in Knox and Lincoln counties. Nine fields were selected. Three fields were stocked with commercial honey bees: 3.0 ha field with 30 hives, 4.2 ha field with 40 hives, and 2.7 ha field with 24 hives. Five fields were selected for stocking with bumble bees: 8.0 ha field with 20 quads, 3.0 ha field with 8 quads, 4.5 ha field with 12 quads, 2.5 ha field with 6 quads, and 4.0 ha field with 10 quads. In addition, a single 1.4 ha field was selected that had no managed bees and relied solely on wild, native bees for pollination. In 2007, nine fields were selected (six in Lincoln Co. and three in Washington Co.) for the study. Four fields were stocked with commercial honey bees: 1.5 ha field with 18 hives, 10 ha field with 102 hives, 6.7 ha field with 66 hives, and 3.3 ha field with 30 hives. Four fields were selected for stocking with bumble bees: 5.0 ha field with 12 quads, 4.0 ha field with 10 quads, 6.7 ha field with 16 quads, and 3.5 ha field with 9 quads. The 3.5 ha bumble bee field had to be removed from the study as additional honey bee colonies were added to the field mid-way through bloom, thus, resulting in four honey bee and three bumble bee fields for 2007. In addition, a single 3.4 ha field was selected that had no managed bees and relied solely on wild, native bees for pollination.
In all three years, three linear transects were laid out from the cluster of honey bee hives or bumble bee quads in each field. The transects were 150 m long and at distances of 10, 25, 50, 100, and 150 m from the colonies. A 1 m2 quadrat was staked out in an adjacent blueberry clone for recording bee activity and, in addition, five stems within each selected clone were randomly chosen, marked with a string lasso around the base, and flowers were counted on each stem for assessing subsequent fruit set. Twelve of the largest fields had an extra sampling station at the end of each transect that was 250 m from the hives. Bee activity was recorded during bloom in each quadrat on each of several visits to each field. One minute counts of honey bees, B. impatiens, other bumble bee species, and other native bees were recorded. After bloom, initial fruit set was estimated by counting the set flowers on each stem and calculating a proportion of green fruit relative to the initial flowers/stem. Prior to commercial harvest, fruits were counted on each stem and the proportion of flowers that produced mature fruit was calculated. Fruit were then harvested and taken back to the laboratory where they were weighed and seeds were counted in a subset of dissected fruits (n = 12 randomly selected berries/field). Proportion fruits set and mature berries were arcsine square root transformed prior to MANOVA and ANOVA. It was not necessary to transform berry weight.
Flower Fidelity 2009–2010
Flower fidelity was measured by using an index of the relative abundance of pure Vaccinium spp. pollen within a bee pollen load. Honey bees and bumble bees (B. impatiens) from commercial hives were collected with a sweepnet on blueberry flowers in each of two fields in Jonesboro, Maine in each year. In both years, one field was an organic field that had a high diversity of flowering plant species within and along the field edge and the second field was a field that was managed with medium intensity at the University of Maine Blueberry Hill Research Farm. In each year, bee collections were conducted on several days during blueberry bloom, removed from the net and the pollen load was extracted from the corbiculae of the hind leg with forceps and then individually placed in a vial and labeled. Forceps were wiped clean with ETOH between bee collections to avoid contamination. In the laboratory, individual pollen loads were placed in a drop of water on a glass slide so that the pollen grains would disperse. Then under a compound microscope 100 pollen grains were inspected and categorized either as tetrad ericaceous pollen or non-ericaceous pollen (CitationStubbs et al., 1994). A percentage of ericaceous pollen was used as the index of fidelity (CitationWhidden, 1996).
RESULTS AND DISCUSSION
A MANOVA evaluating fruit set, proportion of mature berries, and fruit weight provides evidence for a bee treatment effect (F(2,16) = 5.785, P = 0.013), a year effect (F(2,16) = 4.787, P = 0.024), but not a year × bee treatment interaction (P = 0.958). Separate ANOVAs for each pollination measure suggests that there were both year and bee treatment effects for initial fruit set, proportion mature fruit, seed number/berry, and berry weight. None of the pollination measures were characterized by a significant year × bee treatment interaction. lists the means for initial fruit set, proportion of mature fruit, seed number/berry, and berry weight for each of the main effects. Berry weight and seed number were highly related. The number of seeds in a berry explained 28% of the variation in berry weight (F(1,300) = 53.969, P < 0.001). In 2006 and 2007, initial fruit set was significantly higher in honey bee fields than bumble bee fields ( and ), but by harvest the proportion of mature fruit was not significantly different suggesting that “June drop” was higher in honey bee fields. This could be due to a lower number of pollen grains placed on stigmas in a single visit by honey bees than by bumble bees, as has been documented by CitationStubbs and Drummond (1997). This trend was not observed in 2005, where initial fruit set was not significantly different in honey bee pollinated fields compared to honey bee fields. Putting honey bees or bumble bees on a field always resulted in higher fruit set than no deployment of managed bees ().
FIGURE 1 Three measures of pollination for 2006 (A) and 2007 (B). The same letters for a given pollination measure within a year denote means that are not significantly different (α = 0.05).
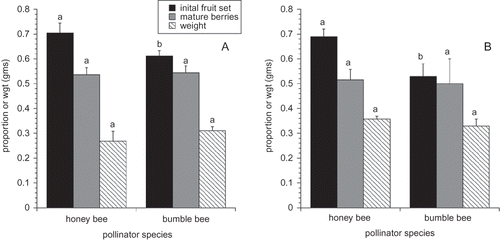
TABLE 1 Measures of Pollination over Three Years in Lowbush Blueberry Fields Supplemented with Honey Bees, Bumble Bees, or Not Supplemented with Commercial Bees (Measures within Parentheses are Standard Errors of the Mean)
It was hypothesized at the beginning of the study that bee forager density would be greatest close to the hives (both bumble bee and honey bee hives are aggregated on the edge of fields near road access) and would decrease as one moved out across the field away from the hives. A split plot ANOVA over the three years (only for the honey bee and bumble bee fields) suggested that bee forager density and the resulting fruit set did not decline as a simple function of distance from the hives (F(4,220) = 0.344, P = 0.848 and F(4,220) = 1.11, P = 0.352; fruit set and bee density, respectively) nor for the interaction bee treatment × distance (F(4,220) = 0.393, P = 0.853 and F(4,220) = 0.778, P = 0.541) or year × bee treatment × distance (F(7,220) = 0.998, P = 0.434 and F(4,220) = 0.566, P = 0.783). This suggests that for a half-circle area of 7.4 acres adjacent to the honey bee and bumble bee hives there was no evidence of central place foraging for these two bee species nor a resulting decline in fruit set. Twenty-two fields had either honey bees or bumble bees stocked. For about 60% of these fields (n = 13), the 7.4-acre area made up 90–100% of the field area. The 7.4-acre sampled area made up 30–70% of the other larger sized fields. I did sample a larger transect (out to 250 m) on 12 of the larger fields. When this data was analyzed separately there was still no evidence of a decline in fruit set or bee forager density (P > 0.05). This is contrary to a previously described decline in honey bee foragers observed in lowbush blueberry by CitationAras et al. (1996) as a function of the distance from the hive, although the single field in this Canadian study was 6.7 km in the longest dimension.
I did find a relationship between fruit set and bumble bee forager density at the whole field level (F(1,10) = 11.807, P = 0.006; ). This relationship was not observed with honey bees (F(1,8) = 0.768, P = 0.406; ). This suggests that increasing hive density of bumble bees may increase forager density and result in increased fruit set. Unfortunately, some of the bumble bee fields also were observed to have very low levels of honey bee visitation, presumably from nearby local hives. This “contamination” could have contributed some of the variation in the regression between bumble bee foragers and fruit set, which explained 58% of the variation. However, a fruit set response may also be the case with honey bees. A few reasons why this might not have been observed in this study are: (1) only a small proportion of honey bee foragers in a colony actually forage (or forage effectively) on the blueberry crop (see below); (2) the variation between foragers in a blueberry field from day to day or between fields is high and while a trend is suggested, the analysis did not have the power to detect it; or (3) that the hive stocking rate for honey bees is approaching the optimal level and this results in little variation in fruit set. Two fields in 2005 would tend to suggest that this might not be the case, although 2005 had the lowest overall fruit set of the three years and for several days during the pollination period the weather was rainy and cold.
FIGURE 2 The relationship between bumble bee forager density (A) and honey bee forager density (B) measured 2–3 times during bloom in each field during each year. The numeral adjacent to each data point denotes the year.
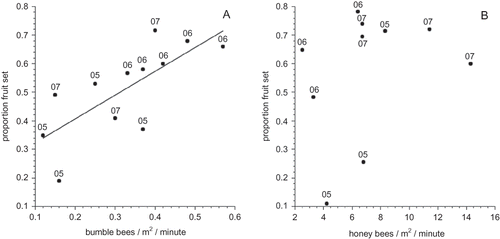
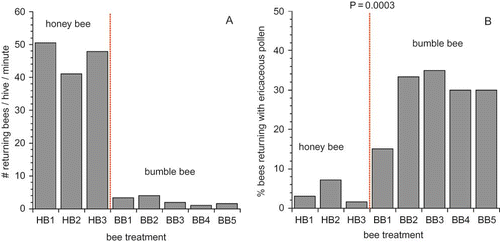
So why do we get such large variation in fruit set and forager density as depicted by and when all the fields were stocked at essentially the same bee density per bee treatment? Certainly landscape characteristics outside of a blueberry field can affect bee foraging and resulting fruit set in a blueberry field. It can be speculated that high abundances of co-flowering plants that secrete high amounts of nectar and high amounts of pollen can easily attract the foraging force out of a blueberry field. Our own data has shown that lowbush blueberry secretes very low levels of nectar in most years (CitationStubbs et al., 2008). In addition, it is known that due to the anatomical structure of blueberry poricidal anthers (CitationDrummond, 2002), bees such as honey bees that do not buzz pollinate have a difficult time collecting pollen from blueberry flowers (CitationThorpe, 2000). Unfortunately, we do not know what characterized the surrounding landscape of each of our fields at the time of bloom between 2005 and 2007. However, to partially address the question of forager and fruit set variation between fields of equal stocking rate, we performed several other measures in 2006 and 2009–2010. In 2006, the colony foraging rate was measured by recording the number of bees returning to the hive in several one-minute periods during bloom. In addition, the proportion of bees returning to the hive with blueberry pollen (tetrad ericaceous pollen) was also recorded. In 2009 and 2010, measures of pollen load purity of bees collected on blueberry while foraging provides a measure of floral fidelity of those bees that are in blueberry fields.
depicts the return rate to the hive for one-minute periods. In general, honey bees were 20 times more abundant in returning to the hive than bumble bees. However, these data should only be compared between hives of the same bee species because the return rate will greatly be affected by the amount of time that an individual bee spends foraging in the blueberry field before the return. A bee that stays longer in the field will produce a lower return rate, all other factors being equal. The important aspect of the data in is that the difference in honey bee activity between the three honey bee fields was 20%, but the difference between bumble bee hives was 400% (4×). Thus, there appears to be a large difference in bumble bee foraging between hives within a field (within field data averaged a smaller level of difference, but it was still high at 297%) and between different fields (as shown in ). The percentage of bees returning to the hive with blueberry pollen reflects both the efficiency between the two species of bee, but also possibly the proportion of the population that is foraging in the blueberry field. Only about an average of 3.1% of the honey bees returning to the hives possessed blueberry pollen (assuming all ericaceous tetrad pollen to be blueberry pollen, a reasonable assumption in these field locations for the time of year), while an average of 31.2% of the bumble bees observed during the same time of day were observed bringing back blueberry pollen (). I found that, of those bees foraging on blueberry flowers in the field, fidelity was significantly different between bumble bees and honey bees during both 2009 and 2010 (F(1,99) = 8.69, P = 0.004). Over both years, the average % ericaceous pollen taken from bee corbiculae was 65.7 ± 5.1 (se)% for bumble bees and 87.1 ± 4.8% for honey bees. Another way to look at the data was the percent of bees that had a high percent of ericaceous pollen (>90%) compared to mixed loads of pollen. The two species were also not similar for this measure: 13.0% for bumble bees and 64.2% for honey bees. CitationWhidden (1996) found much higher rates (87.5%) for bumble bees foraging in blueberry in Canada.
FIGURE 3 Measures of the number of foragers returning to the hive (A) and the percent of bees returning with ericaceous pollen (assumed to be mostly blueberry pollen) for both honey bees and bumble bees used in the study in 2006 (color figure available online).
CONCLUSIONS
In summary, research conducted between 2005 and 2007 confirms our earlier findings (CitationStubbs and Drummond, 2001) and those of CitationDesjardins and Olivereira (2006) that commercial bumble bees of the species B. impatiens are good pollinators of lowbush blueberry. This research also confirms that stocking densities of at least 10 colonies/ha will provide equal levels of pollination to similar stocking densities of honey bees. Current rental prices of honey bees and purchase prices of bumble bees suggest an economic benefit in favor of bumble bees.
ACKNOWLEDGMENTS
I would like to thank Judith Collins, Kerry Blenk, and Sara Hoffmann for helping me with measurements of fruit set and bee density. Many Maine blueberry growers were involved in this study and appreciation goes out to them for allowing us to set up this research project in their fields. Special thanks go out to Gordon Colby of Allen's Blueberry Farms, John Campbell of Cherryfield Foods, Inc., and the Passamaquoddy tribe/Northeast Blueberry Company. I also want to acknowledge the assistance from Mr. John Wolf of the Koppert Company. I also wish to acknowledge the Maine Wild Blueberry Commission for funding this research. This is journal article number 3255 of the Maine Agricultural and Forestry Experiment Station.
LITERATURE CITED
- Aras , P. , de Oliveira , D. and Savoie , L. 1996 . Effect of a honey bee gradient on the pollination and yield of lowbush blueberry . J. Econ. Entomol. , 89 ( 5 ) : 1080 – 1083 .
- Boulanger , L.W. , Wood , G.W. , Osgood , E.A. and Dirks , C.O. 1967 . Native bees associated with the lowbush blueberry in Maine and Eastern Canada . Bull. T26 Tech. Ser., Maine Agr. Expt. Sta., Orono, ME and Can. Agr. Res. Sta., Fredericton, New Brunswick ,
- Burdick, E. and D.M. Caron. 2007. MAAREC Beekeeper Survey, University of Delaware http://maarec.cas.psu.edu/pdfs/MAARECSurveyPub.pdf (http://maarec.cas.psu.edu/pdfs/MAARECSurveyPub.pdf)
- Cox-Foster , D.L. , Conlan , S. , Holmes , E.C. , Palacios , G. , Evans , J.D. , Moran , N.A. , Quan , P. , Briese , T. , Hornig , M. and Geiser , D.M. 2007 . A metagenomic survey of microbes in honey bee colony collapse disorder . Sci. Express. DOI: 10.1126/science.1146498. ,
- Delaplane , K.S. and Mayer , D.F. 2000 . Crop pollination by bees , New York : CABI Publishers .
- Desjardins , E.C. and Olivereira , D.D. 2006 . Commercial bumble bee Bombus impatiens (Hymenoptera: Apidae) as a pollinator in lowbush blueberry (Ericales: Ericaceae) fields . J. Econ. Entomol. , 99 ( 2 ) : 443 – 449 .
- Drummond , F.A. 2002 . Honeybees and blueberry pollination , University of Maine Coop. Ext. Wild Blueberry Bulletin No. 629 .
- Drummond , F.A. and Stubbs , C.S. 1997 . Sampling bee populations in lowbush blueberry in Maine. Proc. 6th Internat. Symp on Vaccinium Culture . Acta Hort. , 446 : 101 – 108 .
- Finnamore , A.T. and Neary , M.E. 1978 . Blueberry pollinators of Nova Scotia, with a checklist of blueberry pollinators in eastern Canada and northeastern United States . Ann. Soc. Entomol. Que. , 23 : 168 – 181 .
- Javorek , S.K. , Mackenzie , K.E. and Vander Kloet , S.P. 2002 . Comparative pollination effectiveness among bees (Hymenoptera: Apoidea) on lowbush blueberry (Ericaceae: Vaccinium angustifolium) . Ann. Entomol. Soc. Amer. , 95 ( 3 ) : 345 – 351 .
- Karmo , E.A. 1957 . Pollination of the lowbush blueberry . Nova Scotia Fruit Grower's Assoc. Annu. Rept. , 94 : 93 – 97 .
- Karmo , E.A. August 17–25, 1956 1958 . Honey bees as an aid in orchard and blueberry pollination in Nova Scotia , August 17–25, 1956 , 955 – 959 . Montreal , , Canada : Proceedings of the 10th International Congress of Entomology .
- National Research Council . 2007 . Status of pollinators in North America , Washington , DC : National Academies Press .
- Stubbs , C.S. and Drummond , F.A. 1997 . Blueberry and cranberry (Vaccinium spp.) pollination: A comparison of managed and native bee foraging behavior. Proceedings of the International Symposium on Pollination . Acta Hort. , 437 : 341 – 343 .
- Stubbs , C.S. and Drummond , F.A. 2001 . Bombus impatiens (Hymenoptera: Apidae): An alternative to Apis mellifera (Hymenoptera: Apidae) for lowbush blueberry pollination . J. Econ. Entomol. , 94 ( 3 ) : 609 – 616 .
- Stubbs , C.S. , Drummond , F.A. and Yarborough , D. 2001 . Commercial bumble bee, Bombus impatiens, management for lowbush blueberry. Wild Blueberry Fact Sheet No. 302 (Bull. No. 2421) , 4 Univ. Maine Coop. Ext. Publ .
- Stubbs , C.S. , Drummond , F.A. and Osgood , E. 1994 . Osmia ribifloris biedermannii and Megachile rotundata (Hymenoptera: Megachilidae) introduced into the lowbush blueberry agroecosystem in Maine . J. Kansas Entomol. Soc. , 67 ( 2 ) : 173 – 185 .
- Stubbs , C.S. , Drummond , F.A. and Ginsberg , H. 2008 . Effects of invasive plant species on pollinator service and reproduction in native plants at Acadia National Park , Tech. Report NPS/NER/NRTR–2007/096 .
- Stubbs , C.S. , Jacobson , H.A. , Osgood , E.A. and Drummond , F.A. 1992 . Alternative forage plants for native (wild) bees associated with lowbush blueberry, Vaccinium spp., in Maine . MAES Tech. Bull. , 148 : 54
- Thorpe , R.W. 2000 . “ The collection of pollen by bees ” . In Pollen and pollination , Edited by: Dafni , A. , Hesse , M. and Pacini , E. 211 – 224 . New York , NY : Springer .
- Van Engelsdorp , D. , Hayes , J. , Underwood , R. and Pettis , J.S. 2010 . A survey of honey bee colony losses in the United States, fall 2008 to spring 2009 . J. Apicul. Res. , 49 ( 1 ) : 7 – 14 .
- Whidden , T.L. 1996 . The fidelity of commercially reared colonies of Bombus impatiens Cresson (Hymenoptera: Apidae) to lowbush blueberry in southern New Brunswick . Can. Entomol. , 128 : 957 – 958 .
- Yarborough , D. 2006 . “ Blueberry pruning and pollination ” . In Blueberries for growers, gardeners, promoters , Edited by: Childers , N.F. and Lyrene , P.M. 75 – 83 . DeLeon Springs , FL : E.O. Painter Printing Co .