Abstract
Eleven yield-related plant traits in 33 accessions selected from 3 Nigerian breeding populations were studied to understand the influence of nut size-number trade-off in cashew tree yield. Three-years of data were statistically analyzed for genetic variability estimates and showed that the accessions were significantly different for all the yield-related traits with highest variability in production of hermaphrodite flowers. The pattern of variation was population-specific with sparsely fruiting Brazilian accessions characterized by large nuts and kernels, while the Indian accessions were prolific but with small–medium nuts. Flowering, fruit-set, and fruit-development differed significantly between years suggesting a strong influence of environment on cashew tree production. Consistent relationships were found between nut number and total nut yield at both inter- and intra-population levels, underscoring the importance of the number to nut yield in cashew. However, regression between nut number and nut size confirmed the existence of the size-number trade-off phenomenon in cashew, but the degree varied between populations. Cashew trees with jumbo nuts (>15 kg) were significantly affected by the trade-off, as they exhibited poor flowering and fruit set, in addition to prolonged fruit development. This category of nut size should be discouraged for commercial planting. These preliminary results showed that cashew cultivars with nut sizes between 9.0 and 12.0 g are better, as such trees produced enough nuts per tree to support profitable investments in cashew farming.
INTRODUCTION
Although cashew (Anacardium occidentale L.) is native to Brazil where it was introduced to Africa and Asia in the 16th century (CitationMitchell and Mori, 1987), the crop has thrived in Africa, and the continent now produces more than one-third of the world's raw cashews, with 75% coming from West Africa. Apart from West Africa, other major producing countries are Vietnam, Indian, Brazil, Tanzania, and Mozambique. Cashew is also grown in some sub-tropical parts of the United States. Cashew is one of the commodity nuts in the global market with worldwide trade estimated at $1.5 billion with the African share being approximately $347 million free on board (FOB). As a smallholder crop in Africa, it has been estimated that produce from the cashew tree, especially nuts, provides income and additional livelihood to about 3 million households in the continent (CitationTopper et al., 1998). In addition to employment generation in rural areas, the economic potential of cashew for industrial uses has also been widely reported. For example, the oil from the cashew shell has potential for the manufacturing of brake linings, paints, and vanishes, etc. Juice, alcohol, vinegars, and wines have been produced from pseudo-fruit (apple). Kernel oil suitable for domestic use has been extracted from the kernel. Health-wise, juice from the cashew apple has been reported to be three times richer in vitamin C than citrus, and the kernels contain unsaturated fatty acids, i.e., with low cholesterol (CitationGrundy, 1987). Cashew is also known to be rich in iron, magnesium, and antioxidants (CitationNandi, 1998).
Over the years, output of cashew plantations has continued to decline steadily, and this trend has been traced to low yield of the existing trees. For example, tree yields in a typical farmer's field were such that only 20% of the tree population produced 70% of the nut yield (CitationMartin and Kasuga, 1995). This situation constitutes a serious challenge for a sustainable nut production sub-sector and global cashew industry. However, yield is a complex trait involving interaction of many reproductive traits, and several factors have been attributed to poor yield of cashew trees. Among them are poor genetic material (CitationFoltan and Ludders, 1995); poor and non-synchronized flowering; and adverse environmental factors, such as temperature, sunlight, rainfall, relative humidity, and soil nutrition (CitationParameswaran et al., 1984). CitationPatnaik et al. (1985) and CitationWunnachit and Sedgley (1992) identified poor fruit set and excessive premature fruit drop among other factors responsible for poor yield in cashew. Fruit set can even vary between different ecologies or locations. CitationRao and Hassan (1957) reported only about 3% fruit set on the West Coast of India, and 6–12% fruit set on the East Coast (CitationMurthy et al., 1984). In reality, the number of fruits that attained maturity in cashew was often very low compared to initial fruit set.
Recurrent selection breeding procedure has long been advocated for yield improvement in a tropical tree crop species (CitationBaudouin et al., 1997). However, such breeding methods require adequate information about the genetic variability of the crop species and relationships between yield-related traits. Although studies on the correlations between yield-related characters have been reported (CitationAliyu, 2006; CitationAzevedo et al., 1998), unfortunately, those studies never accounted for the trade-off between the nut size and nut number that may be crucial to cashew tree yield. There have been contrasting reports in the literature on the seed size and number trade-off in plants. For example, while relationships between the seed size and seed number within individual plants of Ipomopsis aggregate (CitationWolf et al., 1986), Lupinus polyphylluss (CitationAniszewski et al., 2001) were significantly negative; correlations in Claytonia virginica (CitationMorgan, 1998) and Lupinus texensis (CitationSchaal, 1980) ranged from significantly negative to significantly positive. Moreover, CitationKoenig et al. (2009) and CitationParciak (2002) reported neutral relationships between seed size and number in Prunus virginiana and Quercus lobata, respectively. With the recent awareness that size is a major criterion that determines the market value of the nut and kernel in the cashew global trade, cashew farmers are now planting cultivars with extra-large nuts to earn premium price from their farm produce. Unfortunately, information on the implication of such a decision on the total nut yield has not been empirically quantified. To this end, there is the need to examine in detail the trade-off phenomenon between the nut size and nut number vis-à-vis total nut yield in cashew to establish a threshold point for optimal yield that can guarantee profitable investments in the production sector of the cashew industry.
In this study, nut size and nut number trade-off was evaluated through genetic correlation analysis between yield-related components for the 33 cashew accessions from three Nigerian breeding populations over three production seasons (years).
MATERIALS AND METHODS
Thirty-three accessions from three cultivated/and breeding populations of cashew conserved and being grown at the research plots of the Cocoa Research Institute of Nigeria, Ibadan, Nigeria (Lat.: 07° 10′ N; Long.: 03° 52′ E) were selected and used for this study. Eleven of these accessions were land races (FA) collected from the farmer's fields in the central states where the crop has thrived over five decades (CitationSanwo et al., 1973). The second population includes 11 open-pollinated progenies selected from Indian accessions (IA) originally introduced by the former Western Nigeria Development Corporation (WNDC) in 1964 (CitationAkinwale and Esan, 1989; CitationSanwo et al., 1973). And the third population was 11 accessions from Brazil (BA) recently imported by a private farmer, Kosoni-Ola Farm Limited, Oro, Kwara State (north-central region). These trees were planted in a randomized complete design of three blocks with each accession represented by three trees per block, thus making a total of nine individuals per accession. The trees were planted at spacing of 9.0 m × 9.0 m, therefore, occupying a total land area of about 2.5 ha. The fields were regularly maintained through cultural and integrated pest management practices to keep pests and disease infestations at a minimum.
To assess the variability between these accessions and examine nut number-size trade-off among other yield-related traits, we selected 11 plant characters (6 fruits, 2 floral, 1 vegetative, and 2 phenological) that have been found to directly and/or indirectly related to nut yield in cashew (CitationAzevedo et al., 1998; CitationMasawe et al., 1998). These include weight of the whole fruit (g) (WWF), nut weight (g) (NWT), nuts/tree (NNT), nut yield/tree (kg) (NYT), kernel weight (g) (KWH), and kernel to nut ratio (KNR). Other characters include the number of hermaphrodite flowers/panicle (HPP), pollen grain fertility (PPS), tree canopy spread (m2) (TCS), days to fruit maturity (DFM), and harvesting period (HPD). To reduce ambiguity due to influence of weather/climatic factors, we collected the data on these 33 accessions for three production seasons (2002/2003, 2003/2004, and 2004/2005). It is important to note that cashew flowering and fruiting in this region starts in September and sometimes continues to April/May of the following year (CitationAliyu and Awopetu, 2003). Data on the fruit characters (WWF, NWT, and KWH) were collected on 50 randomly selected cashew fruits (apple and nut attached) across the four cardinal positions of tree canopy/accession/replicate/year (CitationMasawe et al., 1996). Weighing was done using Sartorius AND-HR-120 model (Sartorius AG, Goettingen, Germany) weighing balance with accuracy (e = 1 mg and d = 0.1 mg). Kernel to nut ratio was derived from the weight of the kernel divided by the weight of the nut to express the relative shelling percentage. Number of nuts per tree (NNT) was the total number of nuts harvested on each tree per year and the weight in kg was recorded as nut yield per tree (NYT). Number of hermaphrodite flowers was scored on 60 randomly selected inflorescences across the four cardinal positions of the tree canopy with the data being collected on alternate days over a period of 16 weeks (October–February) based on our previous work (CitationAliyu and Awopetu, 2003). We adopted a non-destructive technique using permanent marker (ink) to distinguish new flowers from old. Pollen grain fertility was determined from the mean percentage of viable (deeply stained with 2% aceto-carmine stain) pollen grains from anthers (male flowers) collected from twelve inflorescences randomly selected across the four cardinal positions of tree canopy using a binocular Olympus microscope (Olympus Winter & Ibe GmbH., Hamburg, Germany) at 400× magnification. Tree canopy spread (TCS) was the mean of the diameters of the canopy measured in two directions. Number of days to fruit maturity (DFM) was determined when 75% of fruit set from 400 inflorescences (across tree canopy) of the same age attained physiological maturity (see CitationAliyu and Hammed, 2000) and effective maturity date calculated from September 1, i.e., when the earliest flower anthesis is expected in the region (CitationAliyu and Awopetu, 2003). Harvest period (HPD) was calculated from the number of days from the first fruit drop to the last fruit drop of the selected 400 inflorescences for fruit maturity.
Data collected for the three production seasons (years) were summarized and statistically analyzed for the two tailed ANOVA, correlation coefficient, and regression analyses as described by CitationSteel and Torrie (1988) using a Genstat Discovery 3.0 software (VSN International Ltd., Hemel Hempstead, UK) package (www.vsni.co.uk/software/discovery.genstat/).
RESULTS
Variation between the Accessions (G)
The results of the analysis of variance (ANOVA) showed substantial variations (P < 0.01) between the 33 accessions and three production seasons (years) for all 11 plant traits studied (). Significant interactions between the accession and year (accession × year) were recorded in 5 of the 11 traits (see ). An overview of the variation (coefficient of variation) between the 33 accessions showed that the accessions constitute a wide genetic diversity for the flower production, pollen grain fertility, fruit size, nut size, nut yield, kernel size, fruit maturity period, and canopy size (). Mean variation (CV%) ranged from about 19 to 94% in pollen fertility and hermaphrodite flowers production, respectively. Fruit weight ranged between 18.35 g recorded from FA-11 to 260 g in BA-25. Nut weight also ranged between the smallest, 2.50 g obtained from FA-11 to jumbo nut, 19.40 g from BA-23. Among the fruit/nut traits (WWF, NWT, NNT, NYT, KWH, and KNR), nut number per tree (NNT) recorded the largest variation (∼72%), ranging from sparsely fruiting of 160 nuts/tree to a prolific tree of 5,220 nuts/tree for FA-08 and IA-58, respectively. With about 70% variation in nut yield (NYT), the traits ranged from 0.98 kg/tree for the poorest accession, FA-08 to 39.49 kg/tree for FA-06. This type of wide variation in nut yield is common in the cashew field.
TABLE 1 Mean Square from (ANOVA) and Descriptive Statistics between the 33 Cashew Accessions Showing Variability Extant for the 11 Yield-Related Plant Traits
Following a modified classification described by CitationAzam-Ali and Judge (2001) and CitationOhler (1979) and in the ranking of the cashew kernel into 0–1.0 g = W500, 1.01–1.52 g = W450, 1.53–2.00 g = W320, 2.01–3.00 g = W240, and ≥3.01 g = W180, the kernels from the 33 accessions ranged from 0.65 g (i.e., W500) to 5.05 g (i.e., W180) for FA-11 and BA-23, respectively. Thus, it shows that there is a high degree of concordance between nut weight and kernel weight. However, variation in the ratio of the kernel to nut (KNR) was relatively low (CV = 25%) compared to other fruit and nut related traits. It ranged between 0.14 and 0.41 for BA-20 and FA-06, respectively. Interestingly, FA-06 combines prolificness with higher kernel to nut ratio. Average of 0.24 recorded for this yield related trait signify improvement in these accessions but still fell below the global benchmark of 0.26. The highest variability (∼94%) was recorded for the number of hermaphrodite flowers per panicle, which ranged from 0.60 for FA-08 to 130 for FA-06. Although the 33 accessions showed less variation (19%) for pollen grain fertility, nonetheless, the tree pollens showed fertility ranges from 45% for FA-07 to 100% predominant for BA-03. The 33 accessions recorded moderate variability for tree canopy and period of fruit development. Tree canopy between the accessions varied from a small and compact canopy (about 5 m2) for IA-05 to large-open and spread type (17.8 m2) for IA-67. The period of the fruit maturity could be as early as 15 weeks after first flowering as obtained in BA-13 or delayed for several months in late fruiting trees, for example, FA-04. Significant variation (38%) in the period of fruit harvest was recorded between the 33 accessions. While accession with a short harvestable period as early as two weeks was found (e.g., BA-13), there are accessions with prolonged harvest that lasted more than two months from first fruit drop, for example, FA-04 and FA-07 ().
TABLE 2 Mean, Range, Standard Deviation (sd), and Coefficient of Variation (%) (cv) Showing Variability between the Accessions for the 11 Yield-Related Plant Traits Studied
The pattern of variation was similar between the three cashew breeding populations studied although with slight variation. However, the data showed that IA accessions shared substantial similarities in fruit characteristics with farmer's accessions (FA) than the Brazilian accessions (BA) (). Cashew trees from IA and FA accessions produced predominantly medium fruits and nuts (about 70 and 6.0 g), while fruits and nuts from BAs were predominantly large (about 117 and 11.0 g) (). In terms of nut yield, IA accessions surpass the farmer's and Brazilian accessions in nut production (NNT) and overall nut yield per tree (NYT) (), and the trend was similar in hermaphrodite flower production. Brazilian accessions with average large nuts produced corresponding large kernels. In contrast, kernel to nut ratio was higher (0.27) for IA accessions, implying disproportionate evolution in nut and kernel sizes. The average pollen fertility was relatively high between the three groups (71–88%), although farmer's accessions had more trees with low pollen fertility compared to the other two populations. The data showed that average period of fruit development in farmer's accessions was longer than cashew trees from IA and BA accessions ().
Variation between Years (E)
Mean values for the 11 plant traits across the three production years summarized in showed that production in the 2nd season (2003/2004) of the study was comparably lower than the 1st and 3rd production years () and the trend was the same for the three breeding populations. This is an indication of a strong influence of environment on cashew flowering and fruiting. There was a correlation between climatic variables, especially rainfall and temperature, with yearly performance (data not shown). However, the significant interaction effect (accession × year) recorded in some plant traits () could imply that individual trees responded differently under different environmental instances, and further demonstrate complex nature of yield in cashew.
TABLE 3 Variation in the 11 Yield-Related Plant Characters between the 3 Production Seasons (E) for 33 Accessions and across Each of the 3 Breeding Populations
Correlations among Yield-Related Traits between (Inter-) Populations
Correlation coefficients of the eleven plant traits analyzed for the entire 33 accessions are presented in . Across the 33 accessions (FA/IA/BA, ), the 11 traits investigated demonstrated a high level of inter-relationship with 41 of the 55 inter-trait correlations being significant (r values). Positive significant correlations were recorded between days to fruit development (DFM) and period of harvest (HPD), kernel weight (KWH), nut weight (NWT), and fruit weight (WWF). This implies that accessions producing bigger fruits (apple, nuts, and kernel) tend to have longer maturity and harvest periods than those with smaller or medium fruits. Significant negative correlations between days to fruit maturity (DFM), and hermaphrodite flower production (HPP), kernel to nut ratio (KNR), nuts per trees (NNT), and pollen fertility (PPS), suggest that accession with a prolonged maturity period tends to be less prolific with lower recoverable kernels. The harvest period showed similar significant negative correlations with hermaphrodite flowers (HPP), nuts per tree (NNT), and pollen fertility (PPS), in addition to total nut yield (kg) per tree (NYT). There were negative correlations between production of hermaphrodite flowers and fruit size (WWF, NWT, and KWH) and tree canopy (TCS). However, significant positive correlations (with high coefficient (r) values) between hermaphrodite production, and kernel to nut ratio (KNR), nuts per tree (NNT), total nut yield (kg) per tree (NYT), and pollen fertility (PPS), imply strong relationships between tree prolificness and flower production. Accessions producing larger fruits were accompanied with large nuts and kernels and coincidentally characterized by large canopies as kernel weight was positively correlated to nut weight (NWT, r = 0.93), tree canopy size (TCS), and fruit weight (WWF). There were very strong and positive correlations between nuts per tree and total nut yield (NYT) and pollen fertility (PPS), but negatively correlated with nut weight (NWT), tree canopy size (TCS), and fruit weight (WWF). This observation emphasizes the important role of pollen fertility to yield. However, correlation between nut weight (NWT) and total nut yield (kg) per tree (NYT) was negative but not significant, implying complex relationship between nut size and nut yield. However, total nut yield (kg) per tree (NYT) was positively correlated to pollen fertility (PPS), hermaphrodite flowers (HPP), and nuts per tree (NNT). While a significant negative correlation was recorded between pollen fertility (PPS) and tree canopy size (TCS), correlation between tree canopy (TCS) and fruit weight (WWF) was significantly positive.
TABLE 4 Correlation Coefficient Analyses of the Yield Components between the 33 Accessions and within Each of the 3 Cashew Populations
Correlations among Yield-Related Traits within (Intra-) Populations
For future selection procedures for yield in cashew, we ran another correlation coefficient analysis for each of the three breeding populations separately to have a better understanding of the relationships between these traits in cashew. Fewer significant relationships were recorded for IA compared to the Brazilian accessions (BA) that displayed strong coefficient (r) values for 53 out of 55 inter-trait relationships (). This implies that a population with more variability could experience severe consequences of the relationship (negative or positive) as shown by BAs (). There were both consistent and contradictory relationships between the three breeding populations, which has a serious implication for selection work in cashew. For example, fruit development (DFM), kernel weight (KWH), and nuts per tree (NNT) consistently showed positive correlations with harvest period (HPD), nut weight (NWT), and total nut yield (NYT), respectively, across the three populations. Similarly, increasing nut weight (NWT) was negatively correlated to kernel to nut ratio (KNR) for the three populations. Interestingly, while farmers' (FA) and Brazilian (BA) accessions showed very strong positive correlations between hermaphrodite flowers (HPP), pollen grain fertility (PPS), and nut yield (NNT and NYT), Indian accessions (IA) indicated no significant relationships between the number of hermaphrodite flowers and pollen grain fertility. And prolonged fruit development (DFM) has a greater negative consequence on cashew population characterized by jumbo nuts than smaller nuts, as revealed by BA with very strong negative correlations for HPP, NNT, and NYT.
Number of Nuts per Tree vs. Total Nut Yield per Tree (Kg) vs. Nut Weight
3-D regression analyses were plotted for the correlations between nut size (NWT), nut number (NNT), and total nut yield (kg) per tree (NYT) (a–1d) to illustrate these relationships for a better understanding of their role in cashew nut yield. Relationships between number of nuts per tree and total nut yield (kg) per tree were unambiguously positive and consistent across the three populations, either pooled or singly, thus, clearly demonstrating the strong influence of nuts per tree on total nut yield. However, correlations between nut weight (NWT) and nut yield (kg) per tree (NYT) were complicated (a–1d). For example, the pooled accessions showed two distinct patterns of correlations (a, NWT vs. NYT), with accessions having nuts ranging between 2.5 and 12.5 g showing a pattern of positive correlation with total nut yield, in contrast to the negative correlation displayed by those with jumbo nuts (15.0–19.50 g). Not surprising, the non-significant correlation (r = − 0.09) recorded in the pooled accessions implies that both the two correlations cancel the effect of each other. Regression of NWT vs. NYT at the intra-population level clearly showed that the relationship between nut weight and nut yield is positive but additive in nature until the threshold point is reached, afterward the relationship turns negative significantly as depicted graphically in BA (d, NWT vs. NYT).
FIGURE 1 Plots of regression analyses between nut size (g) (NWT) and nut number per tree (NNT), nut number per tree (NNT) and nut yield (kg) per tree (NYT), and nut size (g) (NWT) and nut yield (kg) per tree (NYT) for (a): between 33 Accessions, (b):11 Farmers Accessions (FA), (c): 11 Indian Accessions (IA) and, (d): 11 Brazilian Accessions (BA) (color figure available online).
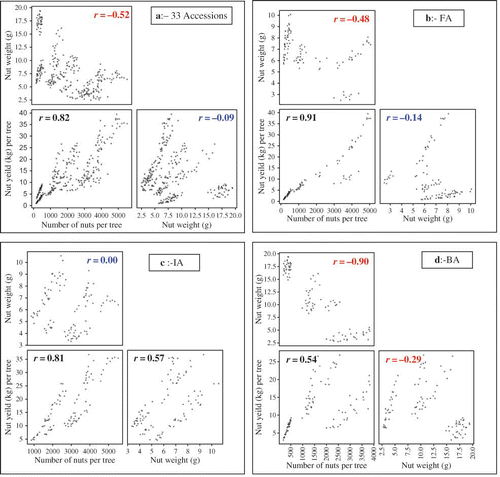
Regression analysis between nut number (NNT) and nut weight (NWT) established the existence of size-number trade-off in cashew (a–1d), as significant negative correlations were recorded for pooled (r = -0.52), farmer's (r = -0.48), and Brazilian (r = -0.90) accessions, but neutral for IA (r = 0.00). This implies that population with jumbo nuts (>15.0 g) will have a higher trade-off effect than genotypes with smaller fruits. The contrasting relationships between nut number and nut weight on one hand and the nut weight and nut yield on the other, suggest the need to identify a threshold level between these traits as a guide for selection of improved cashew cultivars. Comparing the patterns of distribution of the nut yield (kg/tree) and nut number per trees across the 3-D regression analyses, especially pooled and Brazilian accessions (a and 1d), it became obvious that accessions with average nut weights ranging between 9.00 and 12.00 g have the capacity to produce 2,500 nuts/tree, while those with nut sizes between 2.88 and 8.99 g have the tendency to produce as high as about 5,000 nuts per tree (a–1c). However, accessions with mean nut size greater than 15 g rarely produce 500 nuts per tree (d and ). Taking together with nut yield per tree (kg), we found that accessions producing >15 kg/tree were dominated by nut sizes ranging between 6 and 13 g (a–1d and ). In other words, nut sizes outside this range would result to a reduction in output per tree. This information is of importance to cashew breeders and growers questing for bold nuts and high yielding cultivars. It was evident that cashew trees that combine prolific flowering with medium to large nuts (6.0–12.0 g) have the capacity to produce a total nut yield up to about 30 kg per tree per year. For example, out of the 33 accessions evaluated, 9 (27%) viz. FA-05, FA-06, IA-36, IA-58, IA-36, IA-67, BA-03, BA-04, and BA-20 produced nut yields greater than 15 kg/tree, i.e., about 2.6 tons/ha if planted at population density of 175 trees/ha (see ). Accessions with heavier nuts >15.0 g rarely exceed 10 kg per tree per year and those trees with light nuts <4.99 g rarely surpass 15 kg nuts per tree.
DISCUSSION
The main product driving the cashew industry today is the kernel derived from the drupaceous nut, which is typically graded into 26 different classes based on size and color. While color is a qualitative trait, kernel size is quantitative and as such varies significantly and would require a long time for improvement. Such variation has resulted in the classification of cashew kernels into 0–1.0 g = W500, 1.01–1.52 g = W450, 1.53–2.00 g = W320, 2.01–3.00 g = W240, and ≥3.01 g = W180 (CitationAzam-Ali and Judge, 2001) for the whole kernel and prices paid accordingly. In other words, processors are usually looking forward to bigger nuts that will give kernels of higher grades. In this value chain, farmers are conscious of the premium paid for bigger nuts and are, thus, eager to replace old trees producing small nuts with the jumbo nut cultivars. Unfortunately, none of the studies in the past have been able to account for the relationship between the jumbo cultivars and output per tree to guide farmers on profitable investments. Here, we selected three different cashew breeding populations that can provide sufficient variability to study the relationships between yield-related traits and more importantly size-number trade-off in cashew.
Generally, variation patterns recorded in this study demonstrated that the selected accessions were of wider variability in yield-related components and phenological traits, which not only supported previous studies (CitationAliyu and Awopetu, 2007; CitationMartin and Kasuga, 1995; CitationMasawe et al., 1998), but also indicate that the materials are suitable for achieving the objective of this study. Mean values for the fruit size across the 33 accessions suggest that the Brazilian accessions (BA) could be a repository of genes for bolder “jumbo” nuts and kernels of higher grades, and IA and FA accessions for the medium nuts. Since fruit characteristics have been previously shown as a good delineating attribute in cashew (CitationAliyu and Awopetu, 2007), the 33 accessions displayed source-of-origin/population-specific pattern in the fruit (apple, nut, and kernel) characteristics. The accessions introduced from Brazil were unique for extra-large fruits (jumbo) while Indian accessions were distinct for prolific fruiting. CitationPeixoto (1960), quoted by CitationOhler (1979) earlier, reported cashew trees with individual nuts weighing 30 g in Brazil. Moreover, plants growing in the same ecology over a long period are known to evolve common adaptive and unique morphological features (CitationLachenaud and Oliver, 2001; CitationPaudel et al., 2002).
Variation recorded in total nut yield per tree (0.16–39.49 kg/tree) in these accessions falls within the ranges previously reported by other workers (CitationDamodaran et al., 1978; CitationNalini and Santhakumari, 1991; CitationOhler, 1979). Such a wide range in the nut yield is a reflection of variability that exists in cashew farmers' fields and most often the ratio of high to poor yielding trees is typically about 1:8 (CitationOhler, 1979). However, identification of candidate trees with very high yielding and the occurrence of Brazilian accessions capable of producing high premium kernels (W180–W240) indicate a good opportunity for nut yield and kernel size improvement through a recurrent selection breeding program. Strong correlation between nut and kernel sizes as found in this study indicates that nut size could be a reliable selection marker for kernel size in cashew (CitationSanwo, 1980; CitationAliyu, 2006). Prolific flowering and fruiting associated with IA accessions demonstrated that they are advanced generations from a pre-breeding program focusing on selections for high yield in the past (CitationAkinwale and Esan, 1989; CitationSanwo et al., 1973). Variation pattern recorded for vegetative (TCS) and phenological characters (DFM and HPD) showed less polymorphism compared to nut yield characters (CitationAkinwale, 1990; CitationAliyu and Awopetu, 2007), nonetheless, there are trees with compact canopy that can be selected as parents in a breeding for dwarf-compact cultivars that will be suitable for high density planting.
Development of cultivars that combine desirable nut characteristics, i.e., capable of producing high premium kernels and high yielding remain a huge challenge for the global cashew industry. Strategy to harness these two important quality characteristics (size and number) to efficiently balance the effect of trade-off to achieve optimum outputs is crucial to future growth of the cashew economy. Unambiguous positive and significant correlations between number of nuts per tree (NNT) and nut yield (kg) per tree (NYT), irrespective of fruit/nut sizes and populations as recorded in this study, confirm fruiting prolificness is a major determinant for high yielding in cashew. In contrast, nut size (NWT) recorded negative significant correlation to nuts per tree, thus, confirming the existence of a size-number trade-off phenomenon in cashew. These contradicting results pose a great challenge to cashew farmers questing to plant “jumbo” cultivars. However, it was instructive to know that the effect of size-number trade-off was not significant among prolific trees with medium nuts (6.0–9.0 g) as demonstrated by (b and 1c) and FA-05, FA-06, IA-31, IA-58, and IA-66 with production exceeding 3,000 nuts per tree (). CitationVenable (1992) remarked that while negative correlations are sometimes found for seed size-number trade-off among populations or species, some intraspecific correlations have been found to be neutral or positive (CitationMarshall et al., 1986; CitationMazer, 1987; CitationWinn, 1988) and such data were interpreted as a violation of the trade-off assumption. In other words, cashew trees producing nuts of moderate size can circumvent the size-number trade-off rule. Alternatively, trees with these categories of nut sizes can efficiently mobilize and allocate resources for optimum outputs (yield).
However, to attract profitable investments in cashew industry, high yielding cultivars combining prolific fruiting of not less than 2,500 nuts/trees with nut sizes in the range of 9.0 to 12.0 g is recommended to farmers, because such trees have the capacity to produce a minimum of 2 tons of nuts per hectare and attract better prices for the premium kernels. Nut size would serve as a good indicator for selecting cashew kernels of higher grades (CitationSanwo, 1980). Occurrence of size-number trade-off, especially in jumbo accessions, implies the significant role of nutrition and resource allocation in fruit-set and fruit development vis-à-vis a total yield in cashew. An inverse relationship between size and number at inter-specific and inter-population levels has been widely reported in plants (CitationParciak, 2002; CitationWarne and Charnov, 2008). Allocation of resources is known to enhance flowering and fruiting in plants (CitationStephenson, 1981). Alluding to the possible role of nutrition, the proportion of the kernel to nut decrease with an increasing nut size across the three populations, and highest ratios were recorded among prolific trees. For example, FA-06 with the best nut yield coincidentally had the highest ratio of a kernel to nut. This could suggest a differential genotypic efficiency for resources uptake and allocation (CitationVenable, 1992).
Negative correlations between phenological attributes and nut yield were indirect effects through the hermaphrodite flowers per panicle and pollen grain fertility as shown in our data. Suffice that trees with delay flowering are less prolific for hermaphrodite flowers with reduced pollen fertility. Also, negative correlations between nut yield (NNT and NYT) and phenological characters (DFM and HPD) indicates that trees with delay fruiting and a prolonged harvesting period are characteristically low in nut yield, as none of the late fruiters produce up to 8 kg nuts/tree. Similar negative correlation between floral characters (HPP and PPS) and tree canopy size (TCS) suggests that trees with large canopies tend to produce few hermaphrodite flowers with less fertile pollens, therefore, indirectly reducing total nut yield. A tree with a large canopy is often characterized by overcrowding of branches, poor light penetration, poor aeration, and high humidity, which can significantly affect flower production and the fertility of pollen grains. The work of CitationGomez (2004)on Quercus ilex demonstrated empirical evidence of dependence of offspring fitness on seed size and that larger seeds benefit seedlings against competition (CitationJakobsson and Eriksson, 2000) herbivory (CitationBonfil, 1998) shading (CitationBond et al., 1999), drought (CitationSeiwa et al., 2002), and nutrient limitation (CitationSeiwa, 2000) In other words, selection pressure for offspring fitness through increased nut size (bigger endosperm-cotyledon) can lead to evolution of plants with a large and bushy canopy with fewer flowers and fruits attracting fewer predators. Interestingly, positive correlation between nut size and plant vigor (fitness) has been reported in the cashew (CitationAliyu and Akintaro, 2007; CitationAzevedo et al., 1998). Increased cell size and cell number, for example, in stomata, a consequence of genomic plasticity (CitationBeaulieu et al., 2008), have direct influence on plant organs and size (CitationChung et al., 1998) and help in plant adaptation (CitationHodgson et al., 2010). Large seeded soybean characterized by a large cotyledon “kernel” was associated with a large number of cells (CitationSwank et al., 1987). Evolution of large seeded may be an adaptive trait for the cashew plant in its natural habitat in the Brazilian Amazonian forest.
Variation in the performance of the accessions between the production seasons (years) signifies the important role of climatic factors, such as rainfall, sunlight, temperature, and humidity, etc. on cashew yield (CitationOhler, 1979). According to a study by CitationTolla (2004), available soil moisture can cause up to 17% variability in cashew tree yield between years. The indirect effect of adverse weather on flowering (hermaphrodite flowers and pollen fertility) manifested on yield. Moreover, some tropical tree species are known to exhibit biennial fruiting characteristics. The interaction between accessions and years further demonstrate the complexity between environmental indices and genetic expression in cashew. It is, therefore, instructive that selection for nut yield quality, i.e., size and number, should be based on single tree basis over generations.
CONCLUSIONS
Although these results are preliminary, it is believed that they can guide cashew breeders faced with the task of development of improved cultivars that combine extra-large nut size with high yields. Planting of cultivars that are entirely jumbo nuts (>15 g) should be discouraged for commercial cashew orchards because of the strong effect of nut size-number trade-off on nut yield. There is a need for further studies across different ecologies to have a broad data base for the validation of these preliminary results.
ACKNOWLEDGMENTS
The authors are grateful to the Management and staff of Kosoni-Ola Farms Limited, Oro, Kwara State and the Executive Director of the Cocoa Research Institute of Nigeria for the plant materials. Also acknowledged are Messrs S. O. Odeleye and B. A. Adigun of the Plant Breeding Research Group, CRIN, Ibadan, for support on the field data collection.
Related Research Data
LITERATURE CITED
- Akinwale , S.A. 1990 . Biological characterization of cashew (Anacardium occidentale L.) , 211 Nigeria : Ph.D. Thesis, submitted to the Department of Botany, Obafemi Awolowo University, Ile–Ife .
- Akinwale , S.A. and Esan , E.B. 1989 . Advances in cashew breeding in Nigeria , 2nd , 177 – 174 . Ibadan, , Nigeria : Cocoa Research Institute of Nigeria . Progress in tree crop research
- Aliyu , O.M. and Akintaro , G.O. 2007 . Studies on the effect of storage period and nut-size on the seedling vigour of cashew . American-Eurasia J. Sci. Res. , 2 ( 1 ) : 68 – 74 .
- Aliyu , O.M. and Awopetu , J.A. 2007 . Multivariate analyses of cashew (Anacardium occidentale L.) germplasm in Nigeria . Silvae Genetica , 56 ( 3–4 ) : 170 – 179 .
- Aliyu , O.M. 2006 . Phenotypic correlation and path coefficient analysis of nut yield and yield components in cashew . Silvae Genetica , 55 ( 1 ) : 19 – 24 .
- Aliyu , O.M. and Awopetu , J.A. 2003 . Studies on flowering pattern in cashew (Anacardium occidentale L.) . Nigerian J. Genet. , 18 : 29 – 35 .
- Aliyu , O.M. and Hammed , L.A. 2000 . A study of nut and apple development in cashew (Anacardium occidentale L.) . Nigerian J. Tree Crop Res. , 4 ( 2 ) : 1 – 10 .
- Aniszewski , T. , Kupari , M.H. and Leinonen , A.J. 2001 . Seed number, seed size and seed diversity in Washington lupin (Lupinus polyphyllus Lindl.) . Ann. Bot. , 87 : 77 – 82 .
- Azam-Ali, S.H. and E.C. Judge. 2001. Small scale cashew nut processing. United Nations Food and Agriculture Organization. www.fao.org/ag/ags/agsi/Cashew/Cashew.htm (http://www.fao.org/ag/ags/agsi/Cashew/Cashew.htm)
- Azevedo , D.M.P. , Crisóstomo , J.R. , Almeida , F.C.G. and Rossetti , A.G. 1998 . Estimates of genetic correlations and correlated responses to selection in cashew (Anacardium occidentale L.) Genet . Mol. Biol. (online) , 21 ( 3 ) doi: DOI: 10.1590/S1415–47571998000300019
- Baudouin , L. , Baril , C. , Clément-Demange , A. , Leroy , T. and Paulin , D. 1997 . Recurrent selection of tropical tree crops . Euphytica , 96 ( 1 ) : 101 – 114 .
- Beaulieu , J.M. , Leitch , I.J. , Patel , S. , Pendharkar , A. and Knight , C.A. 2008 . Genome size is a strong predictor of cell size and stomatal density in angiosperms . New Phytologist , 173 : 422 – 437 .
- Bond , W.J. , Honig , M. and Maze , K.E. 1999 . Seed size and seedling emergence: An allometric relationship and some ecological implications . Oecologia , 120 : 132 – 136 .
- Bonfil , C. 1998 . The effect of seed size, cotyledon reserves, and herbivory on seedling survival and growth in Quercus rugosa and Q. laurina (Fagaceae) . Amer. J. Bot. , 85 : 79 – 87 .
- Chung , J. , Lee , J.H. and Arumuganathan , K. 1998 . Relationships between nuclear DNA content and seed and leaf size in soybean . Theor. Appl. Genet. , 96 : 1064 – 1068 .
- Damodaran , V.K. , Veeraraghavan , P.G. and Vasavan , M.G. 1978 . “ Variation in F1 population of cashew (Anacardium occidentale L.) ” . Vellanikkara, Kerala : M.S. (Hort.) Thesis, Kerala Agricultural University .
- Foltan , H. and Ludders , P. 1995 . Flowering, fruit-set and genotype compatibility in cashew . Angewadte Botanik , 69 : 215 – 220 .
- Gomez , J.M. 2004 . Bigger is not always better: Conflicting selective pressures on seed size in Quercus ilex . Evolution , 58 : 71 – 80 .
- Grundy , S.M. 1987 . Monounsaturated fatty acids, plasma cholesterol, and coronary heart disease . Amer. J. Clinical Nutr. , 45 : 1168 – 1175 .
- Hodgson , J.G. , Sharafi , M. , Jalil , A. , Diaz , S. , Montserrat-Marti , G. and Palmer , C. 2010 . Stomatal vs. genome size in angiosperms: The somatic tail wagging the genomic dog? . Ann. Bot. , 105 : 573 – 584 .
- Jakobsson , A. and Eriksson , O. 2000 . A comparative study of seed number, seed size, seedling size and recruitment in grassland plants . Oikos , 88 : 494 – 502 .
- Koenig , W.D. , Knopsy , J.M.N. , Carmenz , W.J. and Sage , R.D. 2009 . No trade-off between seed size and number in the valley oak Quercus lobata . Amer. Naturalist , 173 ( 5 ) : 1 – 8 .
- Lachenaud , P.H. and Oliver , G. 2001 . Variability in various agronomic traits of wild cocoa trees (Theobromacacao L.) from the Campoi and Tanpok basins . Plant Genet. Resources Nwsl. , 128 : 35 – 40 .
- Marshall , D.L. , Levin , D.A. and Fowler , N.L. 1986 . Plasticity of yield components in response to stress in Sesbania marcrocarpa and Sesbania vesicaria (Leguminosae) . Amer. Naturalist , 127 : 508 – 521 .
- Martin , P.J. and Kasuga , L.J. 1995 . Variation in cashew tree yield in South-east Tanzania and the implication for management of cashew smallholding . Trop. Agr. , 72 ( 4 ) : 261 – 268 .
- Masawe , P.A.L. , Cundall , E.P. and Caligari , P.D.S. Performance in South-eastern Tanzania of local cashew germplasm and selected clones . Proceedings of the International Cashew and Coconut Conference . February 17–21 1997 , Dar es Salam, Tanzania. pp. 146 – 150 . BioHybrids International Ltd., Reading, UK .
- Masawe , P.A.L. , Cundall , E.P. and Caligari , P.D.S. 1996 . Distribution of cashew flower sex-types between clones and sides of tree canopies in Tanzania . Ann. Bot. , 78 : 553 – 558 .
- Mazer , S.J. 1987 . The quantitative genetics of life history and fitness components in Raphanus raphanistrum L. (Brassicaceae): Ecological and evolutionary consequences of seed mass variation . Amer. Naturalist , 130 : 891 – 914 .
- Mitchell , J.D. and Mori , S.A. 1987 . The cashew and its relatives (Anacardium, Anacardiaceae) . Memoir. New York Bot. Gard. , 42 : 1 – 76 .
- Morgan , M.T. 1998 . Female fertility per flower and trade-offs between size and number in Claytonia virginica (Portulacaceae) . Amer. J. Bot. , 85 : 1231 – 1236 .
- Murthy , K.N. , Vijayakumar , K. , Pillai , R.S.N. and Kumaran , P.M. Aspects of flowering behaviour and correlation studies in cashew tree (Anacardium occidentale L.) . Proceedings of the International Cashew Symposium, Cochin, Central Plantation Crop Research Institute (ICAR) . Kerala, India. pp. 279 – 280 .
- Nalini , P.V. and Santhakumari , S. 1991 . Study on performance of selected types of cashew at Cashew Research Station, Ankayam, Kerala . The Cashew , 5 ( 3 ) : 3 – 6 .
- Nandi , B.K. Cashew nut nutritional aspects. In: and (eds.). Integrated production practices of cashew in Asia. Food and Agriculture Organization of the United Nations Regional Office for Asia and the Pacific . March 23 2011 , Bangkok, Thailand. Edited by: Papademetriou , M.K. and Herath , E.M. FAO/RAP Publication: 1998/12 . Accessed
- Ohler , J.G. 1979 . Cashew , Amsterdam, , The Netherlands : Koninklijk Instituut voor de Tropen; Teskin, Zutphen Co .
- Parameswaran , N.K. , Daramodaran , V.K. and Prabhakaran , P.V. 1984 . Factors influencing yield in cashew (Anacardium occidentale L) . Indian Cashew J. , 16 : 15 – 19 .
- Parciak , W. 2002 . Environmental variation in seed number, size, and dispersal of a fleshy-fruited plants . Ecology , 83 : 780 – 793 .
- Patnaik , H.P. , Das , M.S. and Panda , J.M. 1985 . Studies on fruit set and fruit drop in cashew (Anacardium occidentale L.) under Orissa conditions . Cashew Causerie , 7 : 7 – 8 .
- Paudel , K.C. , Pieber , K. , Klumpp , R. , Laimer , M. and Camara-Machado , D.A. 2002 . Collection of germplasm of Lapsi (Choerospondias axillaries [Roxb.], B.L. Burtt and A.W. Hill), an indigenous fruit tree of Nepal . Plant Genet. Resources Nwsl. , 130 : 36 – 46 .
- Peixoto , A. 1960 . Cashew, Technical Bulletin on Production , 60 Brazil : Ministry of Agriculture, Rio de Janeiro .
- Rao , V.N.M. and Hassan , M.V. 1957 . Preliminary studies on the floral biology of cashew (Anacardium occidentale L.) . Indian J. Sci. , 7 : 277 – 288 .
- Sanwo , J.O. , Kuti , B.O. and Osundolire , J. 1973 . Germplasm collections. 1972/73 . Annual report of Cocoa Research Institute of Nigeria, Ibadan, Nigeria , : 101 – 103 .
- Sanwo , J.O. 1980 . Correlation coefficient between “fruits” traits as possible aids in the selection of desirable cashew (Anacardium occidentale L.) . Nigerian J. Sci. , 14 ( 1 ) : 225 – 233 .
- Schaal , B.A. 1980 . Reproductive capacity and seed size in Lupinus texensis . Amer. J. Bot. , 67 : 703 – 709 .
- Seiwa , K. 2000 . Effects of seed size and emergence time on tree seedling establishment: Importance of developmental constraints . Oecologia , 123 : 208 – 215 .
- Seiwa , K. , Watanabe , A. , Saitoh , T. , Kanno , H. and Akasaka , S. 2002 . Effects of burying depth and seed size on seedling establishment of Japanese chestnuts, Castanea crenata . For. Ecol. Mgt. , 164 : 149 – 156 .
- Steel , R.G. and Torrie , J.H. 1988 . Principles and procedures of statistics , New York : McGraw Hill .
- Stephenson , A.G. 1981 . Flower and fruiting abortion: Proximate causes and ultimate functions . Annu. Rev. Ecol. System. , 12 : 253 – 279 .
- Swank , J.C. , Egli , D.B. and Pfeiffer , T.W. 1987 . Seed growth characteristics of soybean genotypes differing in duration of seed fill . Crop Sci. , 27 : 85 – 89 .
- Tolla, T.D. 2004. Effects of moisture conditions and management on production of cashew: A case study in the Lower Limpopo basin, Mozambique. M.S. Thesis, submitted to the International Institute for Geo-information Science and Earth Observation, Enschede, The Netherlands, p. 95. www.itc.nl/library/papers_2004/msc/.../teshome_demissie_tolla.pdf (http://www.itc.nl/library/papers_2004/msc/.../teshome_demissie_tolla.pdf)
- Topper , C.P. , Martin , P.J. , Katinila , N. , Kikoka , L.P. , Lamboll , R. , Masawe , P.A.L. and Shomari , S.H. The historical and institutional background of the Tanzanian cashew industry . Proceedings of the International Cashew and Coconut Conference: Trees for Life, the Key to Development . Edited by: Topper , C.P. , Caligari , P.D.S. , Kullaya , A.K. , Shomari , S.H. , Kasuga , L.J. , Masawe , P.A.L. and Mpunali , A.A. pp. 76 – 83 . BioHybrid International, Reading, UK .
- Venable , D.L. 1992 . Size-number trade-off and variation of seed size with plant resource status . Amer. Naturalist , 140 : 287 – 304 .
- Warne , R.W. and Charnov , E.L. 2008 . Reproductive allometry and the size-number trade-off for lizards . Amer. Naturalist , 172 : E80 – E98 .
- Winn , A.A. 1988 . Ecological and evolutionary consequences of seed size in Prunella vulgaris . Ecology , 69 : 1537 – 1544 .
- Wolf , L.L. , Hainsworth , F.R. , Mercier , V. and Benjamin , R. 1986 . Seed size variation and pollinator uncertainty in Ipomopsis aggregate (Polemoniaceae) . J. Ecol. , 74 : 361 – 371 .
- Wunnachit , W. and Sedgley , M. 1992 . Floral structure and phenology of cashew in relation to yield . J. Hort. Sci. , 67 : 769 – 777 .