Abstract
Scion-rootstock interactions were analyzed to test the interaction between environment and above-ground vine phenology. The experiment employed ‘Marechal Foch’ and ‘Vidal Blanc’ as reciprocally grafted, own-rooted, and self-grafted vines. Results suggest that genetically complex phenological factors were under the control of the scion cultivar. The effect of the scion on fruit composition was related to differences in yield between the cultivars. Other effects are secondary in nature as a result of vine vigor and shoot density. Factors potentially under specific root influence, (e.g., fruit-set, water status, nutrition, soil pH, salinity, or root pests) were not limiting in this experiment.
INTRODUCTION
This analysis focuses on the potential use of grape rootstocks to control vegetative and reproductive activities of the grapevine through the modification of vine physiology. Rootstock studies have become increasingly focused on the assumption that several physiological events could be accounted for by rootstock effects without considering the vine-scion relationship and its regulation of phenology, growth, canopy and root architecture, or fruit production. Studies generally focused on scion and root interactions that have specific regulative mechanisms in key physiological processes for roots in general, e.g., water and mineral absorption, when they operate under limiting conditions due to drought, pests, disease, or other factors (CitationKeller, 2010).
This limited knowledge of rootstock physiology is evident in commercial viticulture where 90% of all the vinifera vines of the world are still grafted to fewer then 10 rootstocks and they are chosen mainly for their tolerance to expected soil conditions particularly related to water availability or soil pH (CitationKeller, 2010). Consequently, the impact of rootstock on the performance of the scion cultivar grafted to it is still one of the areas of viticulture frequently involved in controversy. Determining the effect of rootstocks on important quantifiable viticultural parameters is ambiguous because separating the observables with respect to their cause is often difficult and makes a determination as to the cause of the observation speculative. Additionally, a genotype's performance is intimately tied to the environment of its evaluation and this can influence the performance of the rootstock as well as the scion cultivar grafted to it producing another limitation on the validity of any conclusion drawn about the rootstock-scion interaction.
Consideration of direct root contribution to above ground vine structures provides a physiological basis for root influence on the performance of the scion. Roots anchor the vine to the soil, take up water and nutrients, and produce and transport plant hormones, including abscisic acid, auxins, gibberellins, and ethylene (CitationRom, 1987). Furthermore, roots serve as a repository of stored carbohydrates (CitationEdson et al., 1995) and nitrogenous compounds (CitationWermelinger, 1991), which are important in the spring growth flush prior to full canopy expression. However, determining direct responses to root influences requires an initial defining of terms (CitationStriegler & Howell, 1991). A primary root effect would be one that directly influenced a scion response via an understood aspect of root morphology or physiology, such as the fruit-set response noted above. A secondary response would include an indirect scion response, such as canopy density, as influenced by the rootstock's direct impact on scion vigor. The following experimental efforts were established with a goal to test two hypotheses. The primary viticultural impact of root system choice is its influence on the vigor of the scion cultivar under conditions of culture and the complex above-ground vine observables like date of bud burst, date of crop ripening, and levels of cold resistance are under the genetic control of the scion cultivar as influenced by the ambient environment. To test these two hypotheses, two-hybrid direct producer cultivars (Marechal Foch and Vidal Blanc) were selected based on specific variation in these characteristics.
MATERIALS AND METHODS
Plant Material
Marechal Foch (Kuhlmann 188-2, aka ‘Foch’) and Vidal Blanc (Vidal-256, aka ‘Vidal’) are cultivars capable of economic production levels as own-rooted and they are described by CitationGalet (1979) as follows: “‘Marechal Foch’ is a complex hybrid result of a crossing of 101-14 Mgt. × ‘Goldriesling’ which resulted from a cross between ‘Riesling’ and ‘Courtiller musque’. It produces small blue-black berries on winged cylindrical clusters.” With regard to ‘Vidal Blanc’, CitationGalet (1979) notes: “‘Vidal Blanc’ is also a complex hybrid resulting from a crossing of ‘Ugni Blanc’ (syn. ‘Trebbiano’) and Seibel 4986. It produces small greenish-white berries on medium large cylindrical clusters.” Bud burst for ‘Vidal’ is commonly 10–14 days later than ‘Foch’ and harvest at technological maturity is 4–6 weeks later in Michigan (CitationAnderson et al., 1980). Cold hardiness assessments in both the laboratory and in the vineyard suggest that ‘Foch’ is much more cold hardy in both bud and wood than is ‘Vidal’. Finally, ‘Foch’ is less vigorous at the same locations regardless of soil type (within a range of viticulture suitable soils) and achieves less vine size (expressed as annual cane pruning weights) than does ‘Vidal’ (). Based upon these attributes the two were chosen to demonstrate whether rootstock, in fact, alters phenological characteristics of the scion. The effect would be notable when applied in a reciprocal graft system.
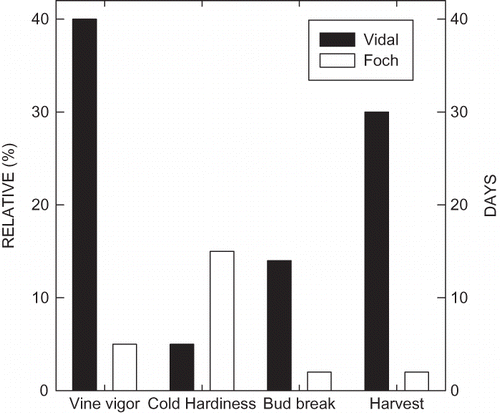
FIGURE 1 Diagrammatic representation of phenological characteristics of the two cultivars utilized in the experiment. On a relative scale, ‘Vidal’ tends to have higher vine vigor (+40%) and reduced cold hardiness (-10%) than Foch. Also, ‘Vidal’ tends to commence bud break 10–14 days later than ‘Foch’ and harvest for ‘Vidal’ is 25–30 days later than Foch in Michigan. Bud break for Foch is between the end of April and the first week of May. Harvest for Foch is between the last week of August and the first week of September (unpublished data from Michigan Lake Shore Appellation).
Grafting Treatments
Mature dormant canes of both cultivars were collected from Michigan State University vineyard at the Horticulture Teaching Research Center (HTRC in East Lansing; N 42.43, W 84.24) and provided to a commercial nursery for bench grafting. All material for scion or rootstock use was from the same source vineyard and collected from healthy, vigorous vines, i.e., exceeding 0.65 kg of cane prunings per meter of row. Vines were grown for a year in the nursery and shipped for planting the following spring as dormant vines. Upon receipt, vines were sorted by fresh weight, and similar weight class groupings were created for each replicate. Thus, initial vine fresh weight was blocked when the replicates were planted. Grafting treatments applied in this study for ‘Marechal Foch’ (F) and ‘Vidal Blanc’ (V) were: F-OR, F-F, F-V, V-OR, V-V, and V-F where the first letter in the treatment code is the scion cultivar, the second letter is the rootstock and OR means Own-Rooted.
Vineyard Establishment
The experiments were conducted at the Horticulture Teaching and Research Center (HTRC) of Michigan State University. Michigan's climate is characterized by a short growing season (150 to 175 days from last spring frost to first fall frost) with cool climate conditions during the summer (1200 ± 300 growing degree days (GDD), base 10°C; CitationBaskerville & Emin, 1969). Yield and quality are often limited by several factors, namely, spring freeze (50% chance of spring frost as late as 15 May), early fall frost, high humidity, and rainfall during the harvest season (CitationHowell & Sabbatini, 2008, CitationSabbatini & Howell, 2010).
Vines were planted to 2.4 × 3.0 m spacing within and between rows, respectively. The soil is a Kalamazoo sandy loam (USDA, 1957). Each vine was trained to Hudson River Umbrella (a bilateral cordon at the top wire) with a height of 1.8 m. Vines were trained with double-trunks with a single cordon arising from each. Production in this system comes from eight-node canes and two-node renewal spurs, spaced at 20 cm along the cordon, with canes and spurs alternated, leaving approximately 60 nodes per vine retained at pruning. Four-vine plots were planted with an un-grafted guard vine between each plot. The blocked vines, based on initial vine fresh weights, were organized to produce five replicates. Each block was arranged as a two-factor factorial with the scion cultivar being one factor and the rootstock cultivar being the other.
Crop Control
After 3 years of establishment, mature vines were pruned to five-node canes and two-node renewal spurs for the subsequent experimental years. ‘Foch’ was pruned to 30 nodes for the initial 0.5 kg of cane pruned and 10 additional nodes for each subsequent 0.5 kg. ‘Vidal’ was pruned similarly except that the severity of the pruning was greater based on its much greater bud fruitfulness (CitationHowell et al., 1987). A 15+10 pruning severity was employed. After pruning, no additional crop control or canopy management was imposed.
Data Collection
Yield was measured at harvest on a weight per vine basis, and cluster number per vine counted. Samples of 250 random berries per replicate were also collected and weighed at harvest for each treatment and mean berry weight per vine calculated. These yield and berry weight values were used to calculate cluster weight and berries per cluster, respectively. Calculation of the fruitfulness parameter (yield/nodes retained) was used to determine the amount of fruit an average node produced. The Ravaz Index (RI) was used to describe the ratio of reproductive to vegetative growth (yield/vine size indexed as pruning weight) that occurred over the season, thereby providing a post-season assessment of vine balance (CitationRavaz, 1911). Chemical composition of fruit was analyzed using the 250-count berry sample per vine collected on the day of harvest and frozen for later analysis (CitationIland et al., 2004). Prior to analysis, berries were thawed at 24°C for 24 h. Grape juice soluble solids were analyzed using an Atago refractometer (Kirkland, WA, USA), and pH was measured using a 370 Thermo Orion (Beverly, MA, USA) pH meter. An automatic titrator, coupled to an autosampler and control unit (Titroline 96, Schott, Germany) was used to determine titratable acidity.
Cold Hardness Assessment
In the dormant season following the first year of data collection for the experiment, cane and bud tissues were collected and subjected to a controlled freezing regime as previously reported (Stergios & CitationHowell, 1977). Canes were selected based on previously reported criteria such that each sample was taken as a stratified random sample to insure appropriate comparisons among treatments (CitationCochran, 1977; CitationCochran & Cox, 1957). Selected cane tissues were transported in an insulated chest from the vineyard to the laboratory where they were cut into single node sections, and bundled with a thermocouple in each bundle to monitor tissue temperature. Bundles were placed in thermos-type containers to dampen the rate of temperature decline and subjected to a ramping down at 3oC/h. A range of temperatures was pre-selected so that the warmest temperature would cause no injury to buds or canes and the coldest temperature would cause 100% mortality. When each container reached the pre-selected temperature, it was removed from the freezer and placed in a refrigerator at 4°C for 24 h. Tissues were then removed from the containers and placed in a chamber having 100% relative humidity at ambient room temperature (≈20°C) for 7 days and were examined under binocular scopes for damage to critical tissues and to identify any browning of the primary bud and/or phloem and cambium tissues of the cane (CitationStergios & Howell, 1977). Assessment was made on a qualitative basis; tissues were either alive or dead. Determination of the T50 was done using the Spearman-Karber equation (CitationBittenbender & Howell, 1974).
Statistical Analysis
After a 3-year establishment/training period, the vines were cropped and pruned for an additional 3 years. Data were collected for 4 years subsequent to the 6th growing season after planting. Basic statistics, analysis of variance, and correlation analysis were performed using SAS (SAS version 9.1.3; SAS Institute Inc., Cary, NC, USA). Results were tested for homogeneity of variance and subjected to analysis of variance (ANOVA) as a 2×2 factorial (CitationSteel & Torrie, 1980) annually and with the 4-year mean values being for the two factors: above ground variety and below ground variety. Main effects and interactions were assessed and selected relationships subjected to orthogonal contrasts and correlation analysis.
TABLE 1 Main Effects of 4 Years of Data Averages on Influence of Reciprocal Grafts of ‘Marechal Foch’ (F) and ‘Vidal Blanc’ (V) on Vine Growth Characteristics, Yield, Yield Components, and Basic Fruit Chemistry
RESULTS AND DISCUSSION
Results are presented as the average values for 4 years of data collected in mature vines after the first 6 years of vine establishment (). Individual season data have been analyzed and they are identical in conformity with the mean data. Data show that when ‘Marechal Foch’ (F) or ‘Vidal Blanc’ (V) vines were own-rooted (F-OR and V-OR) or self-grafted (F-F and V-V), none of the viticultural parameters collected were affected by the grafting treatments () suggesting no impact of the grafting process in this experiment. However, data illustrated that roots had primary influence on vine size, and that F vine size was increased by ≈80% (mean of F-OR and F-F and percentage calculation) when grafted on the more vigorous V. Contrarily, mean vine size of V was decreased by ≈10% when grafted on the less vigorous F (), although no statistically significant difference with the alternative rootstock was observed. Both root and scion influenced number of nodes retained based on the impact on vine size by roots and the different pruning severity employed for the two scion cultivars (30+10 for Marechal Foch and 15+10 for Vidal Blanc). Different degrees of pruning severity, and the resulting number of retained nodes, were utilized as guided by balanced pruning methodology. Shootless nodes, i.e., blind nodes, data are a general bud hardiness indicator and the cold hardiness test show this to be exclusively under the control of the scion cultivar, with ‘Foch’ more cold hardy than ‘Vidal’, having fewer shootless nodes per vine.
Vine yield, cluster number, berry weight, berries per cluster, and cluster weight are under the control of the scion cultivar. Rootstock had no measurable impact as evidenced by the lack of statistically significant change in these parameters over the same scion on different rootstocks (). Basic fruit characteristics were impacted only when F was grafted on V and probably related to the larger vine size induced by the more vigorous variety, V. Higher levels of titratable acidity as well as lower sugar concentration (°Brix) were observed when the vigorous V was used as scion or rootstock in the F-V combination (). However, when the data were analyzed to determine the main effect, considering F and V above or below ground, the rootstock effect was only evidenced in vine growth and yield (V higher then F), that modified basic fruit chemistry characteristics, reducing sugar and increasing acidity in the grafting combination with V as rootstock. The lower sugar and pH and higher titratable acidity support this interpretation of the data based on scion size differences induced by the root.
Vine phenology was not impacted by the grafting combinations. Results reported in indicate that these phenomena are exclusively under the control of the scion cultivar, with the early variety F not influenced by the late variety V as rootstock and vice versa. The values shown for the root influence are nearly identical for date of bud break, veraison, and date of harvest when calculated as difference from the early variety F. The scion data matched exactly the response of own-rooted vines of that cultivar (data not shown). When the data are elaborated to provide a view of the main effect of the F and V as scion or rootstock (), the above ground effects are clear and due to the genetic difference of the material; meanwhile, the effect of the variety used as rootstock disappears. Cold hardiness data collected during the dormant seasons () are nearly identical to the other parameters collected. V and F differ in cold hardiness (F > V), however the grafting combination did not change the cold resistance of the buds or the canes, showing in the main effect analysis that F had a greater cold hardiness when used as scion, but did not induce higher cold hardiness in V ().
TABLE 2 Main Effects of 4 Years of Data on Reciprocal Grafts of ‘Marechal Foch’ (F) and ‘Vidal Blanc’ (V) on Vine Phenology and the Impact of a Spring Freeze Event in Year 2
TABLE 3 Using Hardy Resistant Cultivars Marechal Foch (F) and Vidal Blanc (V) to Determine the Influence of the Above Ground Primary Bud (10 Bud) and Cane Tissue Cold Hardiness (T50) Influenced by the Above (Scion) or Below Ground (Root) Cultivar, Whether as a Reciprocal Graft of V-F or F-V or Own Rooted (V-V, F-F) in Dormant Season after Year 1 of the Study
In previous reports, we have strived to sort the differences between primary impacts of the rootstock from those deemed secondary, defined as an influence on some other aspect of crop performance (CitationHowell et al., 1987; CitationStriegler et al., 1993). This concern is only magnified as one considers potential sources of variability among vines within a vineyard and, indeed, among characteristics within an individual vine's canopy (CitationHowell & Shaulis, 1980; CitationHowell & Wolpert, 1978). One must account for such sources of variability before it is possible to focus upon a specific viticultural factor, such as rootstock choice, with confidence that the factor is directly responsible for a specific vine response or a set of vine responses. This requires careful experimental design (CitationCochran & Cox, 1957) and the employment of stratified random sampling procedures (CitationCochran, 1977). It is difficult to establish experiments that can effectively distinguish between primary impacts and secondary impacts of rootstocks (CitationStriegler & Howell, 1991). For example, the response of a scion cultivar on different rootstocks as related to ripening date is influenced by yield per vine, shoot density and leaf area-fruit weight ratio. Differences did occur, but none was identifiable as a direct response to the root system. Several reports of root influence on the scion cultivar proved that are readily mediated via water relations and growth regulators (CitationPadgett-Johnson et al., 2000). The data reported here support the hypothesis that most of the complex, multiple gene-influenced phenological responses by scion cultivars grafted to grape rootstocks are controlled by the scion genotype and its expression in the environment of culture. Complex plant factors such as onset of growth in the spring, growing season length, cold resistance and fruit maturity can all be limited by an inadequate root system, but when conditions approach optimum, the genetic characteristics of the scion are paramount.
CONCLUSIONS
The grapevine is a system that starts from the integration of scion and root systems, and impacts several physiological events during the annual cycle. Integration of scion and root activities is complementary, creating correlative interdependence and mutual influence in determining basic physiological events. However, the data reported here suggest that vine phenomena resulting from genetically complex, environmentally-influenced controls are most likely to be under the primary control of the scion cultivar. Scion-rootstock interactions were analyzed to test the complex interaction between the environment and the genetic control of the scion cultivar on the above-ground vine parameters, i.e., date of bud burst, fruit ripening process, and cold resistance. The phylloxera resistant cultivars Marechal Foch (Kuhlmann 188-2—Foch) and Vidal Blanc (Vidal-256—Vidal), phenotypically different, showed that during the 4 years of data collection genetically complex factors of phenology were under the primary control of the scion cultivar. The effect of the scion cultivar on fruit composition was explained by differences in yield and vine size. There are numerous examples of differences among vines grafted to different rootstocks (CitationKeller, 2010). However, this original approach to the analysis of such responses will allow a rigorous determination of the specificity of that response to rootstock. This is important because there are important breeding efforts aimed at solving soil-borne problems (CitationCousins & Walker, 2002) and if direct impacts of rootstock can be effectively demonstrated, then the opportunity to improve vine culture would be greatly enhanced. On the other hand, this study does not rely on rootstocks that are seldom involved in scion regulation and, then, only in limiting growing conditions.
ACKNOWLEDGMENTS
This research was supported by the Michigan Agricultural Experiment Station and the Michigan Grape and Wine Industry Council. We appreciate the help of David Francis in vineyard maintenance, James. A. Wolpert, Oriana Silvestroni, and Alberto Palliotti for extensive discussions, and Alan Green for critical reading of an earlier version of the manuscript.
LITERATURE CITED
- Anderson , K.A. , Howell , G.S. and Wolpert , J.A. 1980 . Phenological development of different Vitis cultivars . Fruit Var. J. , 34 : 5 – 7 .
- Baskerville , G.L. and Emin , P. 1969 . Rapid estimation of heat accumulation from maximum and minimum temperatures . Ecology , 50 : 514 – 517 .
- Bittenbender , H.C. and Howell , G.S. 1974 . Adaptation of the Spearman-Kärber method for estimating the T50 of cold stressed flower buds . J. Amer. Soc. Hort. Sci. , 99 : 187 – 190 .
- Cochran , W.G. 1977 . Sampling techniques , New York : Wiley .
- Cochran , W.G. and Cox , G.M. 1957 . Experimental designs , New York : Wiley .
- Cousins , P. and Walker , M.A. 2002 . Genetics of resistance to Meloidogyne incognita in crosses of grape rootstocks . Theor. Appl. Genet. , 105 : 802 – 807 .
- Edson , C.E. , Howell , G.S. and Flore , J.A. 1995 . Influence of crop load on photosynthesis and dry matter partitioning of Seyval grapevines. III. Seasonal changes in dry matter partitioning, vine morphology, yield and fruit composition . Amer. J. Enol. Viticult. , 46 : 478 – 485 .
- Galet , P. 1979 . A practical ampelography , Ithaca , NY : Cornell University Press .
- Howell , G.S. and Wolpert , J.A. 1978 . Nodes per cane, primary bud phenology, and spring freeze damage to Concord grapevines. A preliminary note . Amer. J. Enol. Viticult. , 29 : 229 – 232 .
- Howell , G.S. and Shaulis , N. 1980 . Factors influencing within-vine variation in the cold resistance of cane and primary bud tissues . Amer. J. Enol. Viticult. , 31 : 158 – 161 .
- Howell , G.S. and Sabbatini , P. 2008 . Achieving vine balance with variable annual weather conditions . Proc. of the Midwest Grape and Wine Conf. , : 93 – 103 .
- Howell , G.S , Mansfield , T.K. and Wolpert , J.A. 1987 . Influence of training system, pruning severity and thinning on yield, vine size and fruit quality of Vidal Blanc grapevines . Amer. J. Enol. Viticult. , 38 : 105 – 112 .
- Iland , P. , Bruer , N. , Edwards , G. , Weeks , S. and Wilkes , E. 2004 . Chemical analysis of grapes and wine; techniques and concepts , Adelaide , , Australia : Patrick Iland Wine Promotions Pty Ltd .
- Keller , M. 2010 . The science of grapevines , Academic Press/Elsevier . New York.
- Padgett-Johnson , M. , Williams , L.E. and Walker , M.A. 2000 . The influence of Vitis riparia rootstock on water relations and gas exchange of Vitis vinifera cv Carignane scion under non-irrigated conditions . Amer. J. Enol. Viticult. , 51 : 137 – 143 .
- Ravaz , M.L. 1911 . L'effeuellage de la vigne . Annales de L'Ecole Nationale d'agriculture de Montpellier , 11 : 216 – 244 .
- Rom , R.C. 1987 . “ Roots ” . In Rootstocks for fruit crops , Edited by: Rom , R.C. and Carlson , R.F. 5 – 28 . New York : Wiley .
- Sabbatini , P. and Howell , G.S. 2010 . Effects of early defoliation on yield, fruit composition and harvest season cluster rot complex of grapevines . HortScience , 45 : 804 – 1808 .
- Steel , R. and Torrie , J.H. 1980 . “ Principles and procedures of statistics ” . In A biometrical approach , New York : McGraw-Hill .
- Stergios , B.G. and Howell , G.S. 1977 . Effect of site on cold acclimation and deacclimation of Concord grapevines . Amer. J. Enol. Viticult. , 28 : 43 – 48 .
- Striegler , R.K. and Howell , G.S. 1991 . Influence of rootstock on the cold hardiness of Seyval grapevines. I. Primary and secondary effects on growth, canopy development, yield, fruit quality, and cold hardiness . Vitis , 30 : 1 – 10 .
- Striegler , R.K. , Howell , G.S. and Flore , J.A. 1993 . Influence of rootstock on the response of Seyval to flooding stress . Amer. J. Enol. Viticult. , 44 : 313 – 319 .
- U.S. Department of Agriculture, Soil Conservation Service . 1957 . Major soils of the north central region, U.S.A. Map. Soil Survey, North Central Regional Publication 76 , Washington , DC : Government Printing Office .
- Wermelinger , B. 1991 . “ Nitrogen dynamics in grapevine: Physiology and modeling ” . In Proc. Int. Symp. on Nitrogen in Grapes and Wine, Seattle, WA, 18–19 June 1991 Edited by: Rantz , J.M. 23 – 31 .