
ABSTRACT
Simultaneous responses of American (Concord), European (Pinot Noir), and American-European hybrid (Traminette) cultivars to seasonal and diurnal variation in field environmental conditions were assessed to propose suitable cultivars for potential use for selection and breeding in grapevines in dry areas. ‘Pinot Noir’ had higher gas exchange parameters, higher water potential, and chlorophyll fluorescence, while ‘Concord’ had the lowest values in all three seasons and experienced moderate stress by the end of each season. The dry conditions in 2005 caused water potential to decline sharply compared to values recorded in 2004 and 2006. ‘Pinot Noir’ and ‘Traminette’ exhibited a mild stress, while ‘Concord’ exhibited a moderate stress in 2006. Photosynthetic rate decreased slightly during the summer with a slight recovery at the beginning of September, and did not differ across the seasons among cultivars. Stomatal conductance followed the same pattern. However, in both 2004 and 2005, stomatal conductance dropped sharply late in the season because of frequently occurring high temperatures that were greater than 32 °C. Maximum morning values of Fv/Fm were within the optimal range. The study suggests the presence of marginal photoinhibition during mid-summer in Concord cultivars, because no full recovery of photochemical efficiency early in the evening was observed. The European cultivar (Pinot Noir) and the American-European hybrid (Traminette) are more adapted to field conditions in Kansas than Concord cultivars. However, high cold-mortality of Pinot Noir due to high sensitivity to cold winter temperature makes the hybrid cultivars the most suitable cultivar for Midwest regions with relatively cold winters.
Introduction
Viticulture and winemaking are affected by many factors. Climate, soils, and cultivars/genotypes are the most important factors (Fraga et al., Citation2012). The entire range of grape growing climate zones is about 10 °C globally; and can be narrower for cultivars such as Pinot Noir, the range is an even narrower 2 °C (Santisi, Citation2011). The National Academy of Sciences proposed that the general shift of warmer temperatures pole ward will cause a significant change in the geographic distribution of wine production (Lallanilla, Citation2013) in the next half century (Hannah et al., Citation2013). Most of the world’s wine-producing regions experience seasonal drought. Global warming is also affecting grapevine development, as indicated by changes in phenology and earlier harvests observed throughout different regions in the world (Webb et al., Citation2007). Webb et al. (Citation2012) reported that the factors driving early maturation and harvest dates observed in many winemaking regions across the globe are due to increased temperatures and shortages in soil water content. The frequency of such drought events with higher temperatures is likely to increase in the U.S. midwestern regions, with pronounced dry summers, with most of the extra precipitation falling in winter and spring (Mishra et al., Citation2010). The occurrence of high temperatures during growing season may impact water uptake and growth development, resulting in a source-sink imbalance and non-uniform maturity with change in fruit composition (Keller, Citation2010; Santisi, Citation2011). These observed effects vary across grape cultivars (Jones et al., Citation2005; Schultz, Citation2000).
High temperatures can last over several days during the summer period; these high temperatures can impact grape development and fruit quality. They can affect flowering when occurring earlier, slow vegetative growth, and berry development (Greer and Weston, Citation2010; Greer and Weedom, Citation2013), which results in harvest delay, yield loss, and alteration of berry composition (Chaves et al., Citation2007; Greer and Weston, Citation2010). The effect of high temperatures on a number of other grapevine cultivars has been documented, but most of the studies have been carried out in controlled environments with a dominant focus on molecular level or short-time responses (Greer and Weston, Citation2010). When Greer and Weston (Citation2010) exposed potted Semillon grapes to high temperatures, they found that susceptibility to high temperatures was dependent on the stage of bunch/berry development, with flowering, veraison, and mid-ripening being highly susceptible stages while the fruit-set stage was highly tolerant of high temperatures. By contrast, Soar et al. (Citation2009), using growth chambers to increase air temperatures to about 40 °C around Shiraz canopies, showed no impact of the increased temperatures on berry sugar ripening. Thus, there appears to be cultivar differences in the response of vines to high temperatures.
Plants display complex and escalating responses to high temperature and water shortages, depending on the frequency, duration, and intensity of the water deficit (Ha et al., Citation2014; Nishiyama et al., Citation2013). These responses range from the molecular and physiological levels (Nishiyama et al., Citation2013; Osakabe et al., Citation2013a, Citation2013b) to the whole plant (Chaves et al., Citation2003).
An immediate response of the grapevine is a decline in the photosynthetic activity (Chaves et al., Citation2007; Santos et al., Citation2007), which is closely associated with a decline in stomata conductance (gs) (Chaves et al., Citation2010; Damour et al., Citation2010). Both responses take place on a diurnal and seasonal basis (Chaves et al., Citation2010). Summer ambient air temperatures and vapor air pressure deficit can both affect stomatal opening in grapevines (Escalona et al., Citation2003). Grapevine cultivars show different stomatal responses to water deficit and, consequently, have different rates of photosynthesis (Escalona et al., Citation2003; Tomás et al., Citation2014).
Grapevines are often grown in areas where water deficits develop and constrain vine growth, especially during the establishment of young vines. Grapes grown in Midwestern regions of the U.S. may experience cold winters and warm, dry summers (Kadir, Citation2006). Temperatures differ dramatically during the seasons, and those over 38 °C are not uncommon in Midwestern U.S. during the summer. Therefore, the growing season is often characterized by low soil moisture and high vapor pressure deficit (Kadir, Citation2006). Although American grape cultivars have high tolerance to cold temperature and are more sensitive to high temperatures and heat stress, with the global warming there is always a need to investigate new cultivars, such as the European types, which might have potential to adapt in some areas in the Midwestern states. An example is ‘Pinot Noir’, which is among the most expensive category of table wine produced in North America (Haeger and Storchmann, Citation2006). Considering the importance of introducing new cultivars into the Midwestern U.S., it is essential to evaluate responses of different cultivars to field conditions. Genotypes within grape species differ in response to high temperature that may develop during the growing season. Kadir (Citation2006) found that the European cultivars are relatively less sensitive to high temperatures than the American cultivars. Thus, there is a substantial genetic variability for the temperature traits. To date, there appears to be a paucity of knowledge of the performance of common grape cultivars in vineyard conditions to the high temperatures that occur during the summer advective air heating that occurs in hot climates. There appears to be no or very limited studies on the performance of the American cultivar, European cultivar, and an American-European hybrid grown simultaneously in vineyard conditions to the high temperatures that occur during the summer. Therefore, this study was conducted to examine the responses of three wine grape cultivars (American cultivar, European cultivar, and an American-European hybrid) simultaneously to high temperatures, and to assess the potential for adoption of the European cultivars in south central Kansas where summer temperatures are between 37 °C and 40 °C.
Materials and methods
Location and plant materials
The research was conducted during three seasons (2004–06) under field conditions at the John C. Pair Horticultural Research in Wichita, Kansas, USA (Latitude, 37°, 31 min, 15.3 s North, Longitude 97°, 18 min, 56.2 s West) on a Canadian fine sandy loam soil (Mollisols). The experiment was set up in a completely randomized block design with three replications (three plots per each cultivar). Each plot consists of three grapevines spaced 2.4 m between rows and 2 m within rows. Vines consisted of three grape genotypes: Grafted Vitis vinifera L. (Pinot Noir), own-rooted Vitis labruscana B. (Concord), and own-rooted French American hybrid (Traminette). ‘Pinot Noir’ vines were trained to a low-cordon system, while ‘Traminette’ and ‘Concord’ vines were trained to a high-cordon system. The three cultivars were selected from 24 cultivars being evaluated on the same vineyard, because, in addition to producing different wine types (white versus red), they are the most promising cultivars in the wine region, and they contrast in precocity (Traminette starts vegetative growth earlier than Pinot Noir or Concord) and in cold hardiness with Concord as the most cold-hardy cultivar.
Irrigation design
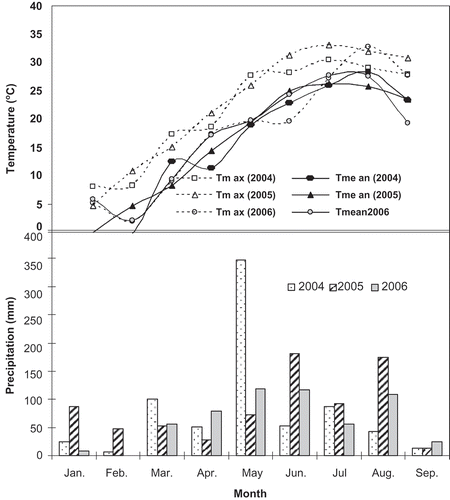
Irrigation water was applied with buried- and above-ground emitters (3 L/h), two per vine, positioned 30 cm from the vine trunk and distributed on both sides of the root system. Water was monitored with flow meters placed in each irrigation line. Water was supplied twice per week from the beginning of berry development in June until July, then three times per week from July to harvest. Near field capacity was monitored by placing tensiometers per each plot. Frequency of irrigation changed slightly from year to year. Cumulative rainfall during the growing season (mid-June until the end of August) was 194, 447, and 147 mm in 2004, 2005, and 2006, respectively. shows monthly rainfall and the mean air temperature at the experimental research station site during the 2004, 2005, and 2006 growing seasons.
Figure 1. Total rainfall (bars) and monthly mean air and maximum air temperature (lines) at the John C. Pair Horticultural Research site, during 2004, 2005, and 2006 growing seasons.
Water relations
Leaf water potential (ΨL) measurements were conducted with a Scholander-type pressure chamber (Model 1000; PMS Instrument Co., Corvallis, OR, USA) during 2005 and 2006 growing seasons. Measurements were taken at 0700 h, midday at 1200–1400 h, and at 1900 h immediately following individual leaf gas exchange measurements in the field. Individual leaves were excised and placed in plastic bags containing moist paper towels and measurements were performed in situ.
Gas exchange parameters
All measurements were taken at five different dates corresponding to: (1) fruit set, (2) pre-veraison, (3) post-veraison, (4) pre-harvest, and (5) post-harvest. Physiological measurements were taken on three fully exposed leaves per each of the three vines per plot.
Photosynthetic CO2 assimilation (A) and stomatal conductance (gs) to water vapor were measured with an open gas exchange system (LI-6400, Li-Cor Inc., Lincoln, NE, USA) on three sun-exposed and fully expanded leaves per vine of three individual vines per plot for each cultivar during the growing seasons of 2004, 2005, and 2006. Red-blue light-emitting diodes maintained incident irradiance at saturating levels (1200 µmol m–2 s–1). The flow rate of air through the sample chamber was set at 400 µmol s–1, and leaf temperature was maintained at 28 ± 0.8 °C by the chamber thermoelectric coolers. The sample chamber CO2 concentration was adjusted to 400 µL L–1 with the system’s CO2 injector (Model 6400-01, LI-Cor Inc.). Vapor pressure deficit (VPD) varied between 1.5 kPa and 2.0 kPa. After the leaf was enclosed in the cuvette, leaves were allowed to reach steady-state values (e.g., when the coefficients of variation of CO2 and H2O inside the chamber were <5%) within 1–4 min. Intrinsec water-use efficiency (WUEi) as the ratio of photosynthesis rate to stomatal closure (A/gs) was calculated. Gas exchange was expressed on a one-sided leaf-area basis for all cultivars.
Leaf chlorophyll fluorescence was measured by a pulse-modulated fluorometer (Fluorescence Monitoring System [FMS-1]; Hansatech Instruments Ltd., Norfolk, U.K.). The FMS-1 requires no dark adaptation of the leaf because it separates actinic light from the fluorescence signal by using modulated fluorometery. Measurements consisted of exposing a leaf section to a LED light source causing excitation of a pulsed fluorescence signal in the absence of actinic light. Four different values were calculated when measuring chlorophyll fluorescence for this experiment: Fo, Fm, Fv, and Fv/Fm. Fo is the initial chlorophyll fluorescence level. Fm represents the maximum fluorescence yield. Fv, or variable fluorescence level, is calculated by simply subtracting Fo from Fm. The PSII efficiency was calculated as Fv/Fm.
Winter damage
Injury to vascular tissue can occur without apparent trunk splitting, resulting in poor growth or subsequent vine collapse later in the growing season. Assessment of thin longitudinal cuts were made to assess the color of the phloem and xylem. Damaged xylem has a milky-white to brown hue, and damaged phloem tissue has a brown color. The percentage of cold-winter damaged vines was based on the number of collapsed vines per each cultivar during the growing season from 2004 to 2007.
Statistical analysis
Prior to submitting data to an analysis, assumptions were checked on the error terms, using residual-based techniques. The Anderson-Darling test was used to verify normality and Barlett’s test was used to verify homogeneity. All data were analyzed using generalized linear models with SAS Ver. 9.13 (SAS Institute, Cary, NC, USA) and least squares means and standard errors determined. Statistical significances were assessed at the 5% level. Regression analysis was used to relate A to gs
Results and discussion
There were significant effects of season and cultivars on most parameters, and season × cultivar interactions were significant for all variables. Seasonal changes in environmental conditions within the experimental site are shown in . Mean daily air temperature fluctuated around 25 °C in June to August during the 2004, 2005, and 2006 seasons, and it became cooler by September. The growing season of 2006 was the warmest; June and July averaged a temperature of 24 °C and 26 °C, respectively. The maximum temperature of the summer season occurred in August with an average of 32 °C before temperatures became cooler in September. Rainfall was two times less than in 2005, and the most rain occurred at the end of August and in early September.
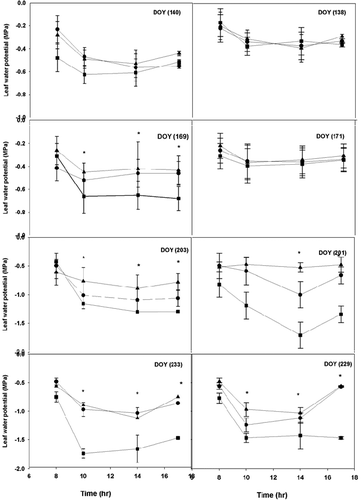
Midday and early morning leaf water potentials were measured only in 2005 and 2006. There was a significance difference between 2005 and 2006 seasons (P < 0.002). The higher temperatures in the summer of 2006 led to higher midday water potentials (more negative values) (ΨL) for all cultivars (). The season of 2005 was marked by heavy rains as compared to 2006. This resulted in relatively less negative ΨL. They can be explained by lower vapor pressure deficits due to mild temperatures. At the end of the season, ‘Concord’ had the highest water potential, but it was not significantly different from ‘Pinot Noir’ and ‘Traminette’ (P < 0.003). Grapevine cultivars did not exhibit any differences in water potential in 2005 across the season (). In 2006, ‘Concord’ and ‘Traminette’ showed a higher midday potential late in the late season compared to ‘Pinot Noir’ (). Significant differences were observed in the last two measurement days of the season. Maximum midday water potential recorded in ‘Concord’, which experienced a moderate stress, are in agreement with other studies (Naor and Wamble, Citation1994). The dry condition in 2006 caused water potential to decline sharply compared to values recorded in 2005. ‘Pinot Noir’ and ‘Traminette’ exhibited a mild stress, while ‘Concord’ exhibited a moderate stress in 2006. ‘Pinor Noir’ and ‘Traminette’ had maximum water potentials of –1.17 MPa and –1.1 MPa, respectively, compared to –1.52 MPa in Concord. We conclude from our study that Concord is the most sensitive among the cultivars. Naor and Wamble (Citation1994) found that leaf water potential of irrigated ‘Concord’ vines decreased from –1.0 MPa to –1.6 MPa during the growing season under field conditions. We believe that the high temperatures had the most impact on ΨL. Grapevine is a water-stress-avoiding species (Chone et al., Citation2001), and stomatal conductivity is regulated to control leaf water deficit. It maintains midday leaf water potential at a constant value (Naor, Citation1998). Diurnal time courses of leaf water potential were typical, declining during the day and with partial recovery in the late afternoon at about 19:00 h (). The differentials between leaf water potentials in the morning and midday were highest at peak temperatures and varied with cultivars (P < 0.0001). Midday values were lower than early morning values for the three species (P < 0.001). In both seasons of 2005 and 2006, Concord recorded the highest values later in the season compared to ‘Pinot Noir’ and ‘Traminete’. Leaf water potential of vines decreased to maintain transpiration (Tognetti et al., Citation2004). Leaf water potential varied between sampling dates and within cultivars (). Although it has been shown that leaf water potential varies with vine water status (Van Leeuven et al., Citation2009) modified by environmental conditions, such as light, temperature, and vapor pressure deficit (Gálvez et al. Citation2014), it is also highly variable depending on the microclimatic environment of each particular leaf (Jones, Citation2004). Jones (Citation2004) reported that temporal fluctuations, such as passing clouds, make the interpretation of ΨL measured during the day as an indicator of irrigation need unsatisfactory. In the semi-arid southern region of Kansas, cloud cover is frequent during the summer months. During the course of this study, none of the measurement days coincided with passing clouds, thus, the measurement of midday ΨL and gs occurred at non-limiting light levels suggesting that ΨL can be used as an indicator of water status for each cultivar.
Figure 2. Seasonal pattern of mid-day leaf water potential of grapevine cultivars: Concord (square), Pinot noir (triangle), and Traminette (circle) in 2005 (A) and in 2006 (B) growing seasons. Bars not visible indicate standard error smaller than symbol. Asterisks denote significant difference among cultivars at p < 0.05. DOY: Day of the year.
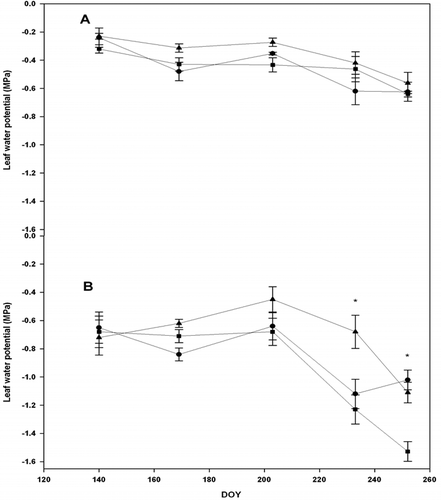
Figure 3. Diurnal course of leaf water potential measured at different times (DOY) in grapevine cultivars: Concord (square), Pinot noir (triangle), and Traminette (circle) in 2005 (right) and 2006 (left) growing seasons. Bars not visible indicate standard error smaller than symbol. Asterisks denote significant differences among cultivars at p < 0.05. DOY: Day of the year.
There were correlations between ΨL and gs for ‘Pinot Noir’ and ‘Traminette’ only with R2 of 0.81 and 0.69, respectively. No correlation was observed in Concord cultivar (data not reported); Lovisolo et al. (Citation2010), who after a review of the literature, stated that there is no clear relationship between gs and ΨL in grapevines. However, Shakel (Citation2007) reported linear relationship (R2 = 0.88) between gs and ΨL of ‘Pinot Noir’ grapevines.
The results from this experiment study would indicate that the measurement of midday ΨL is a sensitive indicator of vine water status and a reflection of daily water use of grapevines under the semi-arid conditions, such as the southern regions of the Midwest. Williams et al. (Citation2010a, Citation2010b) found that both vegetative and reproductive growth of ‘Thompson Seedless’ grapevines grown in the San Joaquin Valley were linearly related to seasonal, mean values of midday ΨL. Therefore, measurements of midday ΨL could be used in a vineyard irrigation management program in the Midwest regions. It is recognized that stomata of isohydric plants are highly regulatory and prevent fluctuations in ΨL. Conversely, anisohydric plants typically have large fluctuations in ΨL and this is usually attributed to poor stomatal control over evaporative demand (Schultz, Citation2003). ‘Pinot Noir’ has previously been characterized as isohydric based on its response to slowly developing soil moisture stress (Schultz, Citation2003). In our case, vines of both ‘Pinot Noir’ and ‘Traminette’ grown with changes in evaporative demand indicated relatively low stomatal sensitivity. By contrast, Concord vines grown in identical conditions were highly responsive to evaporative demand, and exhibited substantially more negative midday ΨL compared with the other cultivars under a similar irrigation regime. Furthermore, midday ΨL of ‘Concord’ vines was strongly influenced by midday VPD, an outcome of the intimate response to evaporative demand. On the basis of these considerations, ‘Traminette’ can be considered as isohydric, while ‘Concord’ can be classified as anisohydric.
The seasonal trends of A and gs in response to environmental conditions are similar, and this results in the close coupling of gas exchange components (Moriana et al., Citation2002). The photosynthetic rate decreased slightly during the summer with a slight recovery at the beginning of September (). The photosynthetic rate at the beginning of summer did not differ across the seasons among cultivars. No significant effect of season was observed on this parameter. Rainfall events, which occurred at the beginning of summer in 2004 and 2005, increased the soil moisture around the entire root system. This avoided restriction of canopy gas exchange due to chemical signals (hormones) produced in roots under drought (Wahbi et al., Citation2005). Thus, photo-assimilation rates in the spring were generally higher than those observed during the rest of the season for each year of the study. ‘Pinot Noir’ always had the highest photosynthetic rate followed by the hybrid ‘Traminette’ and then ‘Concord’. In the 2004 season, photosynthesis of all three cultivars recovered well by the end of the season. In contrast, 2005 revealed a slow recovery, while in 2006, as the warmest growing season of the study, none of the cultivars recovered well. Greer (Citation2012) reported that the leaves appearing in the mid- and late season did not reach the highest rates of photosynthesis compared to rates observed in the early emerging leaves. Similar conclusions were reported on Actinidia deliciosa vines and elsewhere (Greer, Citation2012). This study showed that recovery relies not only on leaf development, but also on cultivar and environmental conditions. All cultivars recovered well in the 2004 season but not in 2006. However, during the 3-year study, Vitis vinifera (Pinot Noir) recovered well, compared to the hybrid and American cultivars. Stomatal conductance followed the same pattern. Stomatal conductance is also known to change with leaf development during the growing season, increasing initially but generally declining with leaf age (Greer, Citation2012).
Figure 4. Seasonal course of photosynthesis, and conductance in grapevine cultivars: Concord (square), Pinot noir (triangle), and Traminette (circle) in Citation2004 (A), 2005 (B), and 2006 (C) growing seasons. Bars not visible indicate standard error smaller than symbol. DOY: Day of the year. Asterisks denote significant differences between treatments for each cultivar.
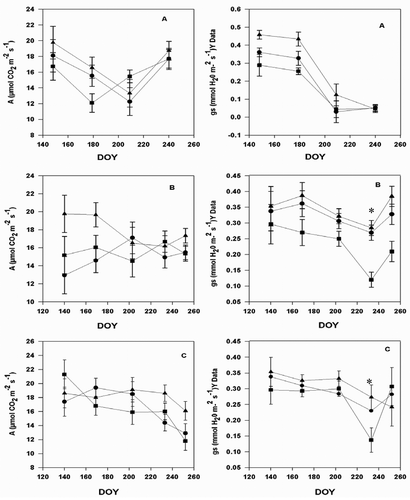
Cultivars exhibited differences in stomatal conductance (P < 0.0001) in the month of July in both 2005 and 2006. ‘Concord’ had a lower gs compared to ‘Pinot Noir’ and ‘Traminette’. This variability in stomatal responses among grape cultivars has been reported (Bota et al., Citation2001). The 2006 growing season was marked with moderate temperatures. In 2006, no significant differences in photosynthetic rate and gs among cultivars were observed, although Pinot Noir had higher values for these parameters than Traminette and Concord. The decline observed at the end of July was less than that observed in 2004 and 2005. Many factors have been implicated in the decrease of gs during the day (Correia et al., Citation1995). In our experiment, the sharp decrease in gs late in the season may be associated with the increase in VPD. As mentioned earlier, there were correlations between ΨL and gs for ‘Pinot Noir’ and ‘Traminette’; this may explain, to a certain extent, the decrease in gs. The decrease in ΨL observed late in the season () in all years may also have affected the decrease in gs through an increased stomatal sensitivity to abscisic acid (Tardieu and Davies, Citation1993).
The current study revealed no consistent relationship between A and gs during mid-season when high temperatures occurred frequently; this might suggest that high temperatures during growing seasons most likely had an impact on non-stomatal limitations of photosynthesis, mainly carboxylation, and ribulose 1, 5-bisphosphate regeneration. Similar findings were reported (Greer and Weedon, Citation2012), which suggest that the activation state of the enzyme Rubisco is inhibited by high temperatures.
In addition, grapevines with gs values decreasing from 200–500 mmol H2O m–2 s–1 to 150 mmol H2O m–2 s–1 are assumed to undergo a mild water stress, while grapevines with gs values between 50 and 150 mmol H2O m–2 s–1 are considered moderately stressed (Lovisolo et al., Citation2010). In this study, all three cultivars experienced moderate stress during 2006, and Concord exhibited stress-sensitivity compared to Traminette and Pinot Noir cultivars. Summer temperatures not only decreased A and gs, but also changed the diurnal patterns of gas exchange (). The general diurnal course of gas exchange followed the pattern mentioned by Zufferey et al. (Citation2011) for different grapevines. Under hotter and drier conditions, as at our experimental site, this pattern changed. In June, the diurnal course of A followed the typical pattern with a midday depression. Two factors are implicated in the reduction of A in the afternoon for similar photon flux density (PFD) conditions. First, there is greater sensitivity of leaf gas exchange to air temperature and VPD under summer conditions (Zufferey et al., Citation2000). Second, gs decreases partly due to an increase in VPD (Düring, Citation1976). This is characteristic of semi-arid environments.
A high correlation was found between intrinsic water use efficiency (WUEi) and stomatal conductance, regardless of the cultivar, mainly in 2004 and 2005 seasons ( and ). This correlation has been described before in grapevine (Pou et al., Citation2008). Intrinsic water use efficiency (A/gs) increased as gs decreased (). A/gs as a function of gs followed the same pattern in 2004 and 2005. A similar response in grapevines was reported by Cuevas et al. (Citation2006) and Cifre et al. (Citation2005).
Figure 5. The relationship between intrinsic water use efficiency (WUEi) and stomatal conductance (gs) in 2004 (A), 2005 (B), and 2006 (C) growing seasons, n = 3.
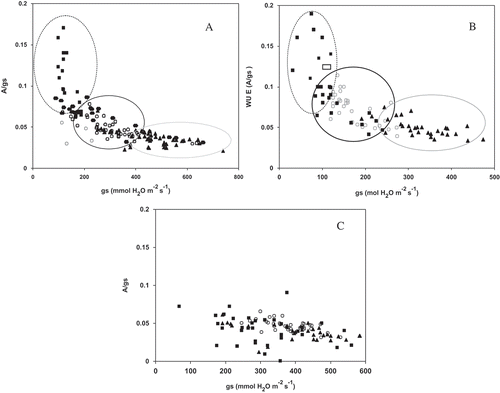
Under low light intensity and high air humidity, as occurs in the morning or evening, grapevine stomata may be widely open at low photosynthetic rates, leading to low WUEi. By contrast, stomatal closure at midday, an important adaptation to high VPD in all three cultivars, may lead to an increase in WUEi when photosynthesis is maintained. This has been observed in grapevines (Souza et al., Citation2003).
Under the field conditions, vines experienced moderate water stress mainly at the end of the season (gs < 50 mmol m−2 s−1). While water stress caused both A and gs to decrease, gs response was more sensitive than A. An increase in A/gs is a mean to offset the decrease in A as stomata close. When gs is higher than 300 mmol m−2 s−1, differences between A/gs in the morning, at midday, and in late afternoon tend to be less (). As water stress increased, differences in A/gs in the morning, midday, and afternoon became more evident. However, based on this study, it is worth reporting that there is a large variability of WUEi within the three cultivars, depending on the environmental conditions. In fact, it has been reported that many cultivars can behave differently with water economy parameters along the growing season (Poni et al., Citation1993) or between experimental years (Pou et al., Citation2012).
Fv/Fm is a good index of photoinhibition in grapevines, and its decrease is generally attributed to damage in PSII (Nunez-Olivera et al., Citation2006). In our study, maximum morning values of Fv/Fm were within the optimal range reported by Björkman and Demming (Citation1987) and they occurred early in the summer in all 3 years. This suggested that there was relatively low stress during these periods (). Indeed, many studies have described reductions in photochemical efficiency and low photosynthetic rates during summer droughts (Valladares and Pearcy, Citation1997). Conversely, morning values of Fv/Fm in July every year were lower than 0.78, with minimum values of 0.67 ± 0.02 for ‘Pinot Noir’, 0.62 ± 0.02 for ‘Traminette’, and 0.57 ± 0.01 for ‘Concord’. These values are in agreement with previous studies on Mediterranean vegetation, which also reported lower Fv/Fm values during the growing seasons (Oliveira and Peñuelas, Citation2001). The diurnal course of Fv/Fm showed a rapid decrease in the morning. Then it became relatively constant and then increased again in the early evening. Higher values in the morning compared to the rest of the day can be explained by the recovery from photoinhibition as nights are warmer after spring (Goethem et al., Citation2013). Thus, our results suggest the presence of marginal photoinhibition during mid-summer in Concord cultivars, because no full recovery of photochemical efficiency early in the evening was observed. In mid-summer, the relatively high assimilation rates of Concord cultivars favor the hypothesis that the observed photoinhibition was due to photoprotection rather than photodamage. Kadir (Citation2006) reported a minor decline of photosystem efficiency in Pinot Noir compared to other cultivars when subjected to higher temperatures. Prolonged exposure to 40/35 °C led to 78% decrease in Fv/Fm in ‘Cynthiana’, compared with 8% decrease in ‘Pinot Noir’, thus corroborating the behavior of ‘Pinot Noir’ under field conditions. The change during the course of the season of Fv/Fm has been reported (Nesterenko et al., Citation2006). The effects of the environmental weather conditions in both seasons may explain the occurrence of a seasonal and diurnal variation in Fv/Fm of our data as reported in other studies (Goethem et al., Citation2013).
Figure 6. Diurnal course of quantum efficiency (Fv/Fm) measured at different times (DOY) in grapevine cultivars: Concord (square), Pinot noir (triangle), and Traminette (circle) in 2005 (A) and 2006 (B) growing season. Bars not visible indicate standard error smaller than symbol. Asterisks denote significant difference among cultivars at p < 0.05. DOY: Day of the year.
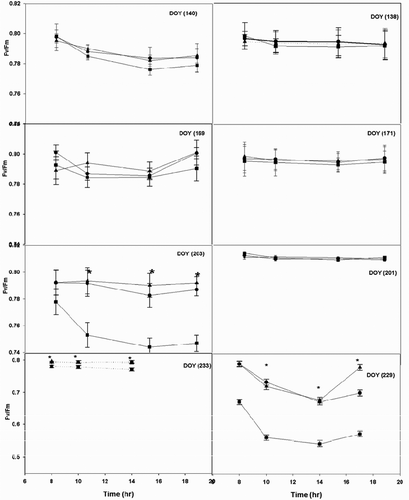
Higher net photosynthetic rates in ‘Pinot Noir’ and ‘Traminette’ than in ‘Concord’ early in the summer would create a higher photochemical sink for the electrons, which protect the photosynthetic apparatus against the accumulation of excessive excitation energy. In addition, the high values of Fv/Fm measured throughout the day, and their almost complete recovery in the evening, suggest that no photoinhibition or chronic damage occurred to vines in this study. These findings suggest that the vines in this experiment were well adapted to their environment and that the environmental conditions that they experienced during the experiment were not so adverse as to cause photoinhibition. Absence of photoinhibition has been frequently found in field-grown vines (Flexas et al., Citation1998).
During the 3 years, it has been observed that the survival of Pinot Noir vines was lower than that of Traminette and Concord; Pinot Noir recorded a 30% of totally collapsed vines while no vines mortality were recorded in neither Traminette nor Concord cultivars.
Conclusion
Regardless of soil water content, a relative water stress developed in the vineyard late in the growing season as a consequence of higher ambient temperatures and VDP. Pinot Noir had higher gas exchange parameters along with higher water potential and chlorophyll fluorescence, while Concord had the lowest values in all three seasons and experienced a moderate stress by the end of the season. The Pinot Noir and Traminette cultivars are more adapted to field conditions in Kansas than Concord. ‘Concord’ is reported to be sensitive to water stress, which we also observed in our experiments. Because ‘Pinot Noir’ is well known to be very sensitive to cold, therefore, we recommend ‘Traminette’ as a potential candidate for field conditions in the Midwestern regions of the U.S.
Literature cited
- Björkman, O., and B. Demming. 1987. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170:489–504.
- Bota, J., J. Flexas, and H. Medrano. 2001. Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Ann. Appl. Biol. 138:353–361.
- Chaves, M.M., J.P. Maroco, and J.S. Pereira. 2003. Understanding plant responses to drought: From genes to the whole plant. Funct. Plant Biol. 30:239–264.
- Chaves, M.M., O. Zarrouk, R. Francisco, M.J. Costa, T. Santos, P.A. Regalado, L.M. Rodrigues, and M.C. Lopes. 2010. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Appl. Biol. 105:661–676.
- Chaves, M.M., T.P. Santos, and C.R. Souza. 2007. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol. 150:237–252.
- Chone, X., C. van Leeuwen, D. Dubourdieu, and J.P. Gaudillere. 2001. Stem water potential is a sensitive indicator of grapevine water status. Ann. Bot. 87:477–483.
- Cifre, J., J. Bota, and J.M. Escalona. 2005. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.): An open gate to improve water-use efficiency?. Agr. Ecosyst. Environ. 106:59–170.
- Correia, M.J., J.S Pereira, M.M. Chaves, M.L. Rodrigues, and C.A. Pacheco. 1995. ABA xylem concentrations determine maximum daily leaf conductance of field-grown Vitis vinifera L. plants. Plant Cell Environ. 18:511–521.
- Cuevas, E., P. Baeza, and J.R. Lissarrague. 2006. Variation in stomatal behaviour and gas exchange between mid-morning and mid-afternoon of north-south oriented grapevines (Vitis vinifera L. cv. Tempranillo) at different levels of soil water availability. Sci. Hort. 108:173–180.
- Damour, G., T. Simonneau, H. Cochard, and L. Urban. 2010. An overview of models of stomatal conductance at the leaf level. Plant Cell Environ. 33:1419–1438.
- Düring, H. 1976. Studies on the environmentally controlled stomatal transpiration in grape vines. I. Effects of light intensity and air humidity. Vitis 15:82–87.
- Escalona, J.M., J. Flexas, J. Bota, and H. Medrano. 2003. Distribution of leaf photosynthesis and transpiration within grapevine canopies under different drought conditions. Vitis 42:57–64.
- Flexas, J., J.M. Escalona, and H. Medrano. 1998. Down-regulation of photosynthesis by drought under field conditions in grapevine leaves. Aust. J. Plant Physiol. 25:893–900.
- Fraga, H., A.C. Malheiro, J. Moutinho-Pereira, and J.A. Santos. 2012. An overview of climate change impacts on European viticulture. Food Energy Secur. 1:94–110.
- Gálvez, R., R. Callejas, G. Reginato, and M.C. Peppi. 2014. Irrigation schedule on table grapes by stem water potential and vapor pressure deficit allows to optimize water use. Ciência Téc. Vitiv. 29(2):60–70.
- Goethem, D., S. De Smedt, R. Valcke, G. Potters, and R. Samson. 2013. Seasonal, diurnal and vertical variation of chlorophyll fluorescence on Phyllostachys humilis in Ireland. PLoS One 8(8):15.
- Greer, D.H. 2012. Modelling leaf photosynthetic and transpiration temperature-dependent responses in Vitis vinifera cv. Semillon grapevines growing in hot, irrigated vineyard conditions. AoB Plants 2012:pls009.
- Greer, D.H., and C. Weston. 2010. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Funct. Plant Biol. 37:206–214.
- Greer, D.H., and M.M. Weedon. 2013. The impact of high temperatures on Vitis vinifera cv. Semillon grapevine performance and berry ripening. Front. Plant Sci. 4:491.
- Ha, C.V., M.A. Leyva-Gonzalez, Y. Osakabe, U.T. Tran, R. Nishiyama, Y. Watanabe, M. Tanaka, M. Seki, S. Yamaguchi, N.V. Dong, K. Yamaguchi-Shinozaki, K. Shinozaki, L. Herrera-Estrella, and L.S. Tran. 2014. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. U.S.A. 111:581–856.
- Haeger, J.W., and K. Storchmann. 2006. Prices of American Pinot Noir wines: Climate, craftsmanship, critics. Agr. Econ. 35:67–78.
- Hannah, L., P. Roehrdanz, M. Ikegami, A. Shepard, M. Shaw, G. Tabo, L. Zhi, P. Marquet, and R. Hijmans. 2013. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. U.S.A. 110(17):6907–6912.
- Jones, G.V., M.A. White, R.C. Owen, and C. Storchmann. 2005. Climate change and global wine quality. Clim. Change 73:319–343.
- Jones, H. 2004. What is water use efficiency?, p. 27–40. In M.A. Bacon (ed.). Water use efficiency in plant biology. CRC Press, Boca Raton, FL.
- Kadir, S. 2006. Thermostability of photosynthesis of Vitis aestivalis and V. vinifera. J. Amer. Soc. Hort. Sci. 131:476–483.
- Keller, M.M. 2010. Managing grapevines to optimise fruit development in a challenging environment: a climate change primer for viticulturists. Aust. J. Grape Wine Res. 16:56–69.
- Lallanilla, M. 2013. Will global warming crush the wine industry? Live Science. 12 January 2016. <http://www.livescience.com/28577-wine-global-warming.html>
- Lovisolo, C., I. Perrone, A. Carra, A. Ferrandion, J. Flexas, H. Medrano, and A. Schubert. 2010. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 37:98–116.
- Mishra, V., K.A. Cherkauer, and S. Shukla. 2010. Assessment of drought due to historic climate variability and projected future climate change in the midwestern United States. J. Hydrometeorol. 11:46–68.
- Moriana, A., A. Villalobos, and E. Fereres. 2002. Stomatal and photosynthetic responses of olive (Olea europea L.) leaves to water deficits. Plant Cell Environ. 25:395–405.
- Naor, A. 1998. Relations between leaf and stem water potentials and stomatal conductance in three field-grown woody species. J. Hort. Sci. Biotech. 73:431–436.
- Naor, A., and R.L. Wample. 1994. Gas exchange and water relations of field-grown Concord (Vitis labruscana Bailey) grapevines. Am. J. Enol. Vitic. 45:333–337.
- Nesterenko, T.V., A.A. Tikhomirov, and V.N. Shikhov. 2006. Onto-genetic approach to the assessment of plant resistance to prolonged stress using chlorophyll fluorescence induction method. Photosynthetica 44:321–332.
- Nishiyama, R., Y. Watanabe, M.A. Leyva-Gonzalez, C. Van Ha, Y. Fujita, M. Tanaka, M. Seki, K. Yamaguchi-Shinozaki, K. Shinozaki, L. Herrera-Estrella, and L.S. Tran. 2013. Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response. Proc. Natl. Acad. Sci. U.S.A. 110:4840–4845.
- Nunez-Olivera, E., J. Martinez-Abaigar, and R. Tomas. 2006. Physiological effects of solar ultraviolet-B exclusion on two cultivars of Vitis vinifera L. from La Rioja, Spain. Am. J. Enol. Vitic. 57:441–448.
- Oliveira, G., and J. Peñuelas. 2001. Allocation of absorbed light energy into photochemistry and dissipation in a semi-deciduous and an evergreen Mediterranean woody species during winter. Aust. J. Plant Physiol. 28:471–480.
- Osakabe, Y., N. Arinaga, T. Umezawa, S. Katsura, K. Nagamachi, H. Tanaka, H. Ohiraki, K. Yamada, S.U. Seo, M. Abo, and E. Yoshimura. 2013a. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 25:609–624.
- Osakabe, Y., K. Yamaguchi-Shinozaki, K. Shinozaki, and L.S. Phan Tran. 2013b. Sensing the environment: Key roles of membrane-localized kinases in plant perception and response to abiotic stress. J. Exp. Bot. 64:445–458.
- Poni, S., L. Marchiol, C. Intrieri, and G. Zerbi. 1993. Gas exchange response of grapevine leaves under fluctuating light. Vitis 32:137–143.
- Pou, A., J. Flexas, M.M. Alsina, J. Bota, C. Carambula, F. De Herralde, J. Galmés, C. Lovisolo, M. Jiménez, B. Ribas, M. Carbo, D. Rusjan, F. Secchi, M. Tomás, Z. Zsofi, and H. Medrano. 2008. Adjustments of water use efficiency by stomatal regulation during drought and recovery in the drought adapted Vitis hybrid. Physiol. Plant. 134:313–323.
- Pou, A., H. Medrano, M. Tomás, S. Martorell, M. Ribas-Carbó, and J. Flexas. 2012. An anisohydric grapevine variety performs better under moderate water stress and recovery than isohydric cultivars. Plant Soil 359:335–349.
- Santisi, J. 2011. Warming up the wine industry. Environ. Mag. 22(6):15–17.
- Santos, T.P., C.M. Lopes, M.L. Rodrigues, C.R. Souza, J.M. Ricardo-da-Silva, J.P. Maroco, J.S. Pereira, and M.M. Chaves. 2007. Effect of deficit irrigation strategies on cluster microclimate for improving fruit composition of Moscatel field-grown grapevines. Sci. Hort. 112:321–330.
- Schultz, H.R. 2000. Physiological mechanisms of water use efficiency in grapevines under drought conditions. Acta Hort. 526:115–136.
- Schultz, H.R. 2003. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 26:1393–1405.
- Shakel, K. 2007. Water relations of woody perennial plant species. J. Intl. Sci. Vigne Vin 41:121–129.
- Soar, C.J., M.J. Collins, and V.O. Sadras. 2009. Irrigated Shiraz vines (Vitis vinifera) upregulate gas exchange and maintain berry growth in response to short spells of high maximum temperature in the field. Funct. Plant Biol. 36:801–814.
- Souza, C.R., J.P. Maroco, T. Santos, P. Tiago, M. Santos, L. Rodrigues, C.M. Lopes, and M. Chaves. 2003. Partial rootzone-drying: Regulation of stomatal aperture and carbon assimilation in field grown grapevines (Vitis vinifera cv Moscatel). Funct. Plant Biol. 30:653–662.
- Tardieu, F., and W.J. Davies. 1993. Integration of hydraulic and chemical signalling in the control of stomatal conductance and water status of droughted plants. Plant Cell Environ. 16:341–349.
- Tognetti, R., R. d’Andria., G. Morelli, D. Calandrelli, and F. Fragnito. 2004. Irrigation effects on daily and seasonal variations of trunk sap flow and leaf water relations in olive trees. Plant Soil 263:249–264.
- Tomás, M., H. Medrano, E. Brugnoli, J.M. Escalona, S. Martorell, A. Pou, M. Ribas-Carbó, and J. Flexas. 2014. Variability of mesophyll conductance in grapevine cultivars under water stress conditions in relation to leaf anatomy and water use efficiency. Aust. J. Grape Wine Res. 20:272–280.
- Valladares, F., and R.W. Pearcy. 1997. Interactions between water stress, sun–shade acclimation, heat tolerance and photoinhibition in the sclerophyll Heteromeles arbutifolia. Plant Cell Environ. 20:25–36.
- Van Leeuwen, C., O. Tregoat, X. Choné, B. Bois, D. Pernet, and J.P. Gaudillère. 2009. Vine water status is a key factor in grape ripening and vintage quality for red Bordeaux wine. How can it be assessed for vineyard management purposes? J. Intl. Sci. Vigne Vin 43:121–134.
- Wahbi, S., R. Wakrim, B. Aganchich, H. Tahi, and R. Serraj. 2005. Effects of partial rootzone drying (PRD) on adult olive tree (Olea europea) in field conditions under arid climate—I. Physiological and agronomic responses. Agr. Ecosyst. Environ. 106:289–301.
- Webb, L.B., P.H. Whetton, and E.W.R. Barlow. 2007. Modeled impact of future climate change on the phenology of winegrapes in Australia. Aust. J. Grape Wine Res. 13:165−175.
- Webb, A.A. 2012. Can timber and water resources be sustainably co-developed in south-eastern New South Wales, Australia? Environ. Develop. Sustain. 14:233−252.
- Williams, L.E., D.W. Grimes, and C.J. Phene. 2010a. The effects of applied water at various fractions of measured evapotranspiration on water relations and vegetative growth of Thompson Seedless. Irrig. Sci. 43:221–232.
- Williams, L.E., D.W. Grimes, and C.J. Phene. 2010b. The effects of applied water at various fractions of measured evapotranspiration on reproductive growth and water productivity of Thompson Seedless. Irrig. Sci. 28:233–243.
- Zufferey, V., F. Murisier, and H.R. Schultz. 2000. A model analysis of the photosynthetic response of Vitis vinifera L. cvs Riesling and Chasselas leaves in the field: I. Interactions of age, light and temperature. Vitis 39:19–26.
- Zufferey, V., H. Cochard, T. Ameglio, J.L. Spring, and O. Viret. 2011. Diurnal cycles of embolism formation and repair in petioles of grapevine (Vitis vinifera cv. Chasselas). J. Exp. Bot. 62:3885–3894.