
ABSTRACT
The research was conducted in the pummelo growing areas of Thailand. The results showed that the dropped fruits had a statistically smaller fruit size (7.25 cm) than the retained fruit (9.35 cm; P < 0.05) at 4 months after fruit set. Moreover, the dropped fruits had an abnormal fruit shape when compared to the retained fruits. Consideration of fruit and leaf carbohydrate concentrations showed that the retained fruits had statistically higher carbohydrate concentrations (262 mg g−1) than the dropped fruits (203 mg g−1; P < 0.05). Meanwhile, the dropped fruits had statistically lower leaf carbohydrate concentrations (55 mg g−1) than those in the retained fruits (85 mg g−1; P < 0.05). The retained fruits also had statistically higher N, P, and K concentrations (1.6%, 1.1%, and 1.3%, respectively) than the dropped fruits (1.1%, 0.5%, and 1.0%, respectively; P < 0.05). Therefore, the causes of early fruit drop in pummelo could be insufficient carbohydrate and plant nutrients supplied for fruit growth and development.
Introduction
Pummelo growers in Thailand have been facing preharvest fruit drop problems for many years. Over the last 2–3 years, this problem has become quite serious with growers in Thailand losing both production and income. A shortage of carbohydrates, imbalances in plant nutrition, and plant hormones have all been reported as causes of the fruit drop in citrus species (Davies and Albrigo, Citation1998) and other fruit crops such as sweet cherry (Blanusa et al., Citation2006) and apple (Ward, Citation2004). In favorable conditions, fruit trees set more fruit than in ideal. This phenomenon results in carbohydrate and plant nutrient competition among fruitlets and fruit drop will occur. Many healthy fruit trees drop fruit naturally in early summer in what is known as the “June drop.” Where a heavy crop has set, too many fruitlets may remain on the branches, resulting in a final crop of disappointingly small fruits. Many researchers have reported that fruit thinning decreased fruit drop and increased carbohydrate availability for remaining fruitlets in citrus species and other fruit species (Atkinson et al., Citation2002; Beruter and Droz, Citation1991; Blanusa et al., Citation2006; Ruiz et al., Citation2001). Spiegel-Roy and Goldschmidt (Citation1996) reported that fruit drop in citrus trees consists of two successive abscission waves affecting flowers and developing fruitlets can be distinguished. The first wave massively induces flower and ovary abscission, whereas the second one reduces the number of growing fruitlets during the June drop. Moreover, a strong relationship between the availability of the amounts of carbohydrates for the fruitlets, especially soluble sugars, and their probability of abscission has been suggested by Iglesias et al. (Citation2003). The decline in carbohydrate levels throughout the flowering and fruit set period (Hilgeman et al., Citation1967; Jones and Steinacker, Citation1951), which is accentuated by heavy flowering (Garcia-Luis et al., Citation1988), indicates that reserve carbohydrates are indeed utilized to sustain the early stages of reproductive development (Shimizu et al., Citation1978). Similar trends in carbohydrate cycling have been reported for avocado (Scholefield et al., Citation1985). Experiments with labeled CO2 indicated that reserve carbohydrates were utilized mainly to support reproductive development, while photosynthesis from mature leaves supplied the needs of vegetative growth (Akao et al., Citation1981). While an imbalance of plant nutrients may also be the cause of the fruit drop in citrus and the other fruit crops, it has been reported that the accumulation of all the macro- and microelement within the fruit also increased during fruit growth and development of pomegranate. The concentration of most elements in arils and peel decreased during fruit growth and development (Mirdehghan and Rahemi, Citation2007). In “Newhall” and “Skagg’s Bonanza” Navel Oranges, Ca, B, Fe, and Mn concentrations in rind reached the greatest levels at 80–180 days after full bloom (DAFB), different from K and Mg, which increased at the period of fruitlet growth (before 80 DAFB) and decreased thereafter. In pulp, concentrations of Ca, K, Mg, and Mn declined gradually with time, whereas a small rise in B toward the end of sampling and a clear increase of Fe at 80–180 DAFB were observed (Sheng et al., Citation2009). Moreover, Pechkeo et al. (Citation2007) found that nutrient accumulation in the mangosteen fruit decreased from blooming to harvesting period. Mangosteen (leaf, peel, and flesh) required higher amounts of N, P, K, Ca, and Mg for growth in the early stage of the fruit development period (from bloom to 6th week after bloom) and S and B in the late stage of the fruit development period (from 6th week after bloom) compared with other growth periods. In apple fruits, the initial nutrient concentrations decreased quickly, undergoing slow and continuous decreases and then remaining almost constant until the end of fruit maturation, indicating nutrient dilution, once the total nutrient accumulation increased gradually with fruit growth (Nachtigall and Dechen, Citation2006). While nutrient imbalance causes fruit drop in many fruit crops, Singh et al. (Citation2008) reported in Aonla or Indian gooseberry that the lower productivity is mainly due to heavy early fruit dropping from initiation of fruitlets to fruit maturity, which is due to several factor, viz., moisture stress, nutritional, temperature, hormonal imbalance, higher wind velocity, insect-pests, and diseases. Moreover, citrus orchards in Pakistan are facing problems of fruit size, color, quality, and excessive premature fruit drop which is due to the deficiencies of essential nutrients (Ibrahim et al., Citation2011). Ashraf et al. (Citation2013) reported that fruit dropping was one of the main reasons of low citrus fruit yield in Pakistan, which may occur as a result of nutrient deficiency. It is obvious that changes in plant nutrient concentrations during fruit growth and development can normally be found and balance of plant nutrients is very important for fruit growth to meet nutrient requirements.
However, information about the causes of preharvest fruit drop in pummelo is still unclear. The objective of this study was to investigate carbohydrate and plant nutrient concentrations in fruits on preharvest fruit drop in pummelo cultivar Thong Dee, which is the most famous pummelo cultivar in Thailand.
Material and method
Plant material
The research was conducted in the pummelo growing areas of Nakhon Pathom province, Thailand during Jan. to Aug. 2013. Ten trees of similar age and size were selected to set up the experiment. Forty inflorescences with the same age in each tree were randomly selected for fruit growth measurement (400 pummelo inflorescences were used for this study in total).
Leaf sample collection
Five pummelo leaves mostly near retained fruits and 5 leaves mostly near dropped fruits (approximately 1–2 in from the fruits) in each pummelo tree were collected for total nonstructural carbohydrate concentration (TNC) at 4 months after fruit set (50 pummelo leaves mostly near retained fruits and 50 pummelo leaves mostly near dropped fruits were used in total). The pummelo leaves were analyzed at 4 months after fruit set due to the pummelo fruits starting to drop obviously at this stage.
Fruit sample collection
Approximately 2 weeks after the selected inflorescences started setting fruits, the pummelo fruits were separated into two groups, normal shaped and abnormally shaped fruits (). The normal shaped fruits were the fruits that would be retained on the tree and had normal fruit growth (the retained fruit), whereas the abnormally shaped fruits were the fruits that would drop from the tree and had slower growth (the dropped fruit). That prediction came from the experience of pummelo growers and the preliminary research. Ten pummelo fruits from each group were collected at 1, 2, 3, and 4 months after fruit set. Peel and pulp of collected fruits were separated and taken to laboratory for TNC and plant nutrient (N, P, and K) concentrations analysis.
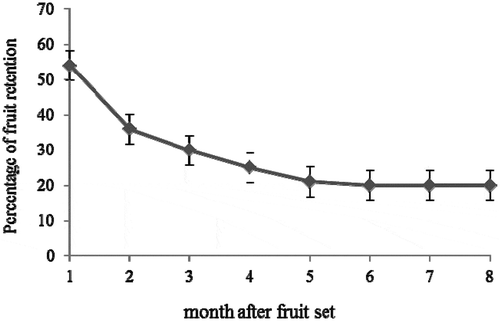
Figure 1. Percentage of fruit retention during fruit growth in pummelo cultivar Thong Dee. The vertical bars presented the standard error of mean.
Total nonstructural carbohydrate analysis
Fruit samples and leaf samples were taken for total nonstructural carbohydrates analysis using Nelson’s procedure (Hodge and Hofreiter, Citation1962). All the samples were analyzed for TNC by washing the sample under tap water, then rinsing with distilled water and finally drying in a hot air oven at 65°C for 72 h. After drying, the material was ground in a Wiley Mill and stored in a desiccator. Total nonstructural carbohydrates were extracted with 0.2 N H2SO4 (Smith, Citation1969) and determined by Nelson’s reducing sugar procedure method at the Central Laboratory, Kasetsart University, Kamphaeng Saen Campus.
Plant nutrient concentration analysis
Pummelo fruit samples were taken for plant nutrient concentration analysis. The plant material was dried and grounded before plant nutrient analysis. From this dried material, the nitrogen (N), phosphorus (P), and potassium (K) concentration were determined. Levels of N were determined using the Microkjeldahl Method. P and K were extracted by HNO3–HClO4 (5:1); then, the solution was left to cool down. Phosphorus in solution was determined calorimetrically by the molybdate-vanadate yellow color method. While K was determined using an atomic absorption spectrophotometer, all plant nutrient analysis was done at the Soil, Plant and Agricultural Material Testing and Research Unit, Central Laboratory, Kasetsart University, Kamphaeng Saen Campus.
Fruit growth measurement
Fruit diameter of both 40 retained fruits and 40 dropped fruits were measured using Vernier caliper every 2 weeks till 3 months after fruit set and then every month until harvesting. Moreover, external (fruit size and fruit shape) and internal fruit (pulp thickness and pulp distribution between the two sides of fruit) characteristics of both pummelo groups were recorded. In addition, percentage of fruit retention was recorded monthly from the first to the seventh months for 40 retained fruits and 40 dropped fruits.
Statistical analysis
All the data were statistically tested by the independent sample t-test method at the 0.05 significant level.
Result and discussion
Fruit growth measurement showed that the retained fruits had higher fruit diameter than that of the dropped fruits at 3–4 months after fruit set. The mean diameter of retained fruits was 7.10, 9.35, 10.25, 10.75, 10.75, and 10.75 cm at 3, 4, 5, 6, and 7 months after fruit set, respectively, whereas the mean diameter of the dropped fruits was 4.10 and 7.25 cm at 3 and 4 months after fruit set, respectively. However, beyond 4 months after fruit set, all abnormally shaped fruit dropped, and hence, it was not possible to record the measurements resulting in no mean diameter of fruit measurement ().
Table 1. Mean fruit diameter (cm) of the retained fruits and the dropped fruits during fruit growth and development in pummelo cultivar Thong Dee (P > 0.05).
In addition, the percentage of fruit retention reduced continually throughout 4 months after fruit set. However, the reduction of fruit retention started to stabilize at the fifth month after fruit set ().
It was observed that the dropped fruits had an abnormal fruit shape, a distorted shape, or a nonspherical shape, whereas the retained fruits had normal fruit shape. Besides, it was found that abnormally shaped dropped fruits started to drop 3 months after fruit set and they were smaller in size than the retained fruits. Moreover, the dropped fruits had less pulp than the retained fruits and pulp was unevenly distributed between the two sides of the fruit ().
Figure 2. External and internal fruit characteristics of the retained and the dropped fruits 1 month after fruit set.

Both the abnormally shaped fruits external and internal characteristics ( and ) obviously dropped more as fruit age increased.
Figure 3. Fruit characteristics in the retained and the dropped fruits as (a) external characteristics and (b) internal characteristics 2 months after fruit set.

Figure 4. Internal fruit characteristics in the retained and the dropped fruits as (a) 3 months after fruit set and (b) 4 months after fruit set.

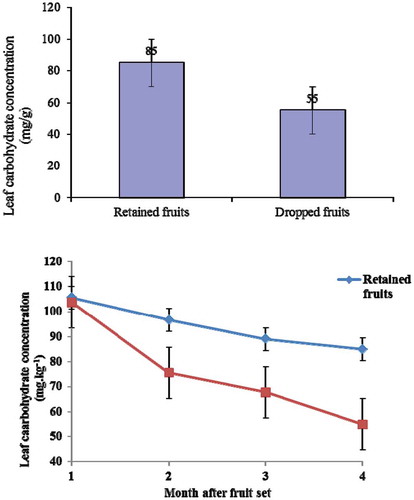
Measurement of fruit carbohydrate concentration showed that the retained fruits had statistically higher carbohydrate concentration (262 mg g−1) than the dropped fruits (203 mg g−1; P < 0.05) (). Meanwhile, the leaf carbohydrate concentration in the dropped fruits had statistically lower carbohydrate concentration (55 mg g−1) than the retained fruits (85 mg g−1; P < 0.05) ().
Figure 5. Fruit carbohydrate concentration in the retained fruit and the dropped fruit of pummelo cultivar Thong Dee at 1–4 months after fruit set (P < 0.05). The vertical bars presented the standard error of mean.
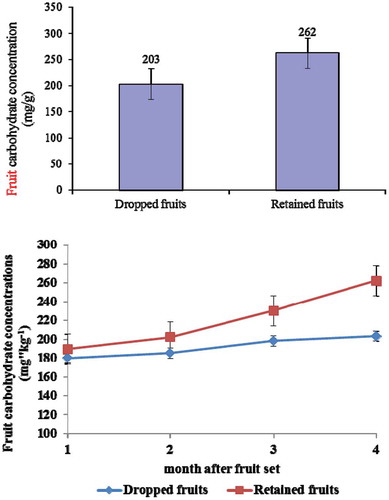
Figure 6. Leaf carbohydrate concentration in the retained fruit and the dropped fruit of pummelo cultivar Thong Dee at 1–4 months after fruit set (P < 0.05). The vertical bars presented the standard error of mean.
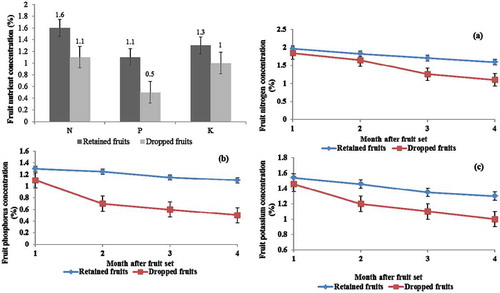
In addition, the retained fruits had statistically higher N, P, and K concentration (1.6%, 1.1%, and 1.3%, respectively) compared to those of the dropped fruits (1.1%, 0.5%, and 1.0%, respectively; P < 0.05) ().
Figure 7. Plant nutrient concentrations in the retained fruits and the dropped fruits on pummelo cultivar Thong Dee at 1–4 months after fruit set (P < 0.05) as (a) nitrogen, (b) phosphorus, and (C) potassium. The vertical bars presented the standard error of mean.
This study found that the dropped fruits had a more abnormal fruit shape and smaller fruit size compared to the retained fruits. These abnormal characteristics could be used to predict which fruits would drop from the tree. In addition, consideration of fruit carbohydrate concentration showed that the dropped fruits had lower carbohydrate concentration than those in the retained fruits. Therefore, the cause of fruit drop in pummelo could be insufficient carbohydrate supply for fruit growth and development. This is consistent with the research of Goldschmidt (Citation1999) who reported that carbohydrate limitation is apparently the primary cause of fruitlet abscission in citrus. This would be due to the carbohydrate competition between developing pummelo fruits. Ruiz et al. (Citation2001) reported that during late fruitlet abscission sucrose concentration in the leaves falls to a low value demonstrating a limitation in supply and competition among the developing fruitlets for carbohydrate. This corresponded to the lower carbohydrate concentration in the mature pummelo leaves from the dropped fruits than those from the retained fruits. Moreover, a correlation between abscission and the carbohydrate levels in leaves and fruitlets at the end of the period of abscission has been found in trees. Thus, fruit thinning altered carbohydrate competition among fruitlets which reduced fruit drop (Garcia-Luis et al., Citation1988). Gonzalez-Carranza et al. (Citation1998) also reported that carbohydrate limitation is apparently the primary cause of fruitlet abscission.
The dropped fruits had a lower N, P, and K concentration than those in the retained fruits. It could be that the abnormal growths of dropped fruits were due to lower nutrient mobilization in the fruit compared to those in the normal fruits. However, leaf nutrient analysis in this study () found that the leaf N, P, and K concentrations were suitable range compared to the nutrient standard value of pummelo for N (2.7–2.9%), P (0.18–0.21%), and K (2.4–3.5%) (Nartvaranant, Citation2010). Thus, the preharvest fruit drop in pummelo may be due to less nutrient mobilization to the dropped fruit more than nutrient shortage. This was in accordance to the physiological role of N, P, and K in fruit growth and development. N plays a major role in agriculture by increasing crop yield and improves fruit quality, fruit number, relating to photosynthetic process, leaf area production, leaf area duration as well as net assimilation rate. N also plays a key role in physiological processes. It imparts the dark green color of plants and promotes leaves, stem, and other vegetative parts’ growth and development. N produces rapid early growth, improves fruit quality, enhances the growth of leafy vegetables, and increases protein content. Thus, balanced amount of N is required for vigorous growth and development processes (Leghari et al., Citation2016). Along with N, P is involved in several key plant functions, including energy transfer, the maintenance of cell membranes and as a bridging element for genetic information, photosynthesis, transformation of sugars and starches, nutrient movement within the plant, and the transfer of genetic characteristics. It has a direct effect on yield and tree health, but it is also important in determining fruit size, firmness, color, storage potential, root growth, seed development, and disease resistance. Adequate P allows the processes described above to operate at optimum rates and growth and development of the plant to proceed at a normal pace (Zhu et al., Citation2017). While K has a key role in many key physiological processes like water relations, opening and closing of stomata, cell division, formation of sugars and starch, and synthesis of proteins, it also improves fruit quality by enhancing fruit size, juice contents, color, size, and juice flavor. So, limited supply of K may affect the yield and quality of citrus fruit and accelerate the fruit dropping (Ashraf et al., Citation2010). From all the literature, it is obvious that balance of plant nutrients is very important for fruit growth. Thus, the occurrence of nutrient imbalance would cause fruit drop in many fruit trees like kinnow (Ashraf et al., Citation2013, Citation2012), mangosteen (Pechkeo et al., Citation2007), and tomato (Zhu et al., Citation2017).
Table 2. leaf N, P, and K concentrations in the retained fruits and the dropped fruits of pummelo cultivar Thong Dee at 1–4 months after fruit set (P > 0.05).
From the results, the preharvest fruit drop in pummelo appears to be due to carbohydrate competition between pummelo fruits during growth and development. This is according to Iglesias et al. (Citation2003) who reported that fruit set in citrus depended on the carbohydrate content of the citrus tree and the dropped fruit has lower starch content than those retained on the tree. Therefore, increasing carbohydrate content in pummelo tree or reducing carbohydrate competition between pummelo fruits such as fruit thinning should reduce the preharvest fruit drop in pummelo cultivar Thong Dee.
Conclusion
The dropped pummelo fruits had an abnormal fruit shape when compared to the retained fruits. This abnormal characteristic could be used to predict which fruits would drop from the tree. Those would benefit the growers in fruit thinning consideration to avoid carbohydrate and plant nutrients competition among the fruitlets which could alleviate the preharvest fruit drop. Fruit and leaf carbohydrate concentrations analysis showed that the retained fruits had higher carbohydrate concentrations than those in the dropped fruits. Moreover, the dropped fruits had statistically lower leaf carbohydrate concentration, fruit N concentration, fruit P concentration, and fruit K concentration than those in the retained fruits. Thus, the causes of early fruit drop in pummelo could be insufficient carbohydrate and plant nutrients supplied for fruit growth and development. From the results, it could suggest that the preharvest fruit drop in pummelo may be alleviated by fruitlet thinning in the suitable stage of fruit growth to ensure good size and fruit quality. Alternatively, orchard management has to be considered carefully including nutrient and irrigation management, ensuring that healthy pummelo trees meet the requirements of fruit growth.
Acknowledgments
The author would like to thank the Thailand Research Fund (TRF) for research funding to complete this study and also to all pummelo growers who have kindly provided material for this research.
Additional information
Funding
Literature cited
- Akao, S., S. Tsukahara, H. Hisada, and S. Ono. 1981. Contribution of photosynthetic assimilates to development of flower and spring flush in Citrus unshiu Marc. J. Japan. Soc. Hort. Sci. 50:1–9. doi: 10.2503/jjshs.50.1.
- Ashraf, M.Y., M. Ashraf, M. Akhtar, K. Mahmood, and M. Saleem. 2013. Improvement in yield quality and reduction in fruit drop in kinnow (Citrus reticulate Blanco) by exogenous application of plant growth regulators, potassium and zinc. Pak. J. Bot 45(51):433–440.
- Ashraf, M.Y., A. Gul, M. Ashraf, F. Hussain, and G. Ebert. 2010. Improvement in yield and quality of kinnow (Citrus deliciosa x Citrus nobilis) by potassium fertilization. J. Plant Nutri. 33:1625–1637. doi: 10.1080/01904167.2010.496887.
- Ashraf, M.Y., M. Yaqub, J. Akhtar, M.A. Khan, M. Ali-Khan, and G. Ebert. 2012. Control of excessive fruit drop and improvement in yield and juice quality of Kinnow (Citrus deliciosa x Citrus nobilis) through nutrient management. Pak. J. Bot. 44:259–265.
- Atkinson, C.J., M.A. Else, A. Stankiewicz, and A.D. Webster. 2002. The effects of phloem girdling on the abscission of Prunus avium L. fruits. J. Hort. Sci. Biotech. 77:22–27. doi: 10.1080/14620316.2002.11511451.
- Berüter, J., and P. Droz. 1991. Studies on locating the signal for fruits abscission in the apple tree. Hort. Sci. 46:3–4. doi: 10.1016/0304-4238(91)90043-X.
- Blanusa, T., M.A. Else, W.J. Davies, and C.J. Atkinson. 2006. Regulation of sweet cherry abscission: The role of photoassimilation, sugars and abscisic acid. J. Hort. Sci. Biotech. 81:613–620. doi: 10.1080/14620316.2006.11512113.
- Davies, F.S., and L.G. Albrigo. 1998. Citrus. CAB International, Wallingford, U.K.
- Garcia-Luis, A., F. Fornes, A. Sanz, and J.L. Guardiola. 1988. The regulation of flowering and fruit set in citrus: Relationship with carbohydrate levels. Israel J. Bot. 37:189–201.
- Goldschmidt, E.E. 1999. Carbohydrate supply as a critical factor for citrus fruit development and productivity. Hort Sci. 34:1020–1024.
- Gonzalez-Carranza, Z.H., E. Lozoya-Gloria, and J.A. Roberts. 1998. Recent developments in abscission: Shedding light on the shedding process. Trends Plant Sci. 3:10–14. doi: 10.1016/S1360-1385(97)01132-1.
- Hilgeman, R.H., J.A. Dunlap, and G.C. Sharples. 1967. Effect of time of harvest of Valencia oranges on leaf carbohydrate content and subsequent set of fruit. P. Am. Soc. Hortic. Sci. 90:110–116.
- Hodge, J.E., and B.T. Hofreiter. 1962. Determination of reducing sugars and carbohydrates, p. 380–394. In: Whistler R.L. and Wolfron M.L. (eds.). Method in carbohydrate chemistry. Academic Press, New York.
- Ibrahim, M., N.A. Abbasi, Z.H. Hafeez-ur-Rehman, and I.A. Hafiz. 2011. Phenological behaviour and effect of different chemicals on pre-harvest fruit drop of Sweet orange cv. Salustiana. Pakistan J. Bot. 43:453–457.
- Iglesias, D.J., F.R. Tadeo, E. Primo-Millo, and M. Talon. 2003. Fruit set dependence on carbohydrate availability in citrus trees. Tree Physiol. 23:199–204. doi: 10.1093/treephys/23.3.199.
- Jones, W.W., and M.L. Steinacker. 1951. Seasonal changes in concentration of sugars and starch in leaves and twigs of citrus trees. P. Am. Soc. Hortic. Sci. 58:1–4.
- Leghari, S.J., N.A. Wahocho, G.M. Laghari, A. Hafeezlaghari, G. Mustafabhabhan, K. Hussaintalpur, T.A. Bhutto, S.A. Wahocho, and A.A. Lashari. 2016. Role of nitrogen for plant growth and development: A review. Adv Environ. Biol. 10(9):209–218.
- Mirdehghan, S.H., and M. Rahemi. 2007. Seasonal changes of mineral nutrients and phenolics in pomegranate (Punica granatum L.) fruit. Sci. Hortic. 111(2):120–127. doi: 10.1016/j.scienta.2006.10.001.
- Nachtigall, G.R., and A.R. Dechen. 2006. Seasonality of nutrients in leaves and fruits of apple trees. Sci. Agri. 63(5):493–501. doi: 10.1590/S0103-90162006000500012.
- Nartvaranant, P. 2010. A suitable sampling method of pummelo leaf cv. Thongdee for evaluation of plant elements nutrient status. Agric. Sci. . 41(3):373–382.
- Pechkeo, S., S. Sdoodee, and C. Nilnond. 2007. Changes of plant nutrients concentration in soils and trees of mangosteen (Garcinia mangostana L.) during the fruit development. Kasetsart J. (Nat. Sci.) 41:61–71.
- Ruiz, R., A. Garcia-Luis, C. Monerri, and J.L. Guardiola. 2001. Carbohydrate availability in relation to fruitlet abscission in citrus. Ann. Bot. 87(6):805–812. doi: 10.1006/anbo.2001.1415.
- Scholefield, P.B., M. Sedgley, and D. McE. Alexander. 1985. Carbohydrate cycling in relation to shoot growth, floral initiation and development and yield in the avocado. Sci. Hortic. 25:99–110. doi: 10.1016/0304-4238(85)90081-0.
- Sheng, O., X.Y. Shu, A. Peng, X.X. Deng, and Y.W. Fang. 2009. Seasonal changes in nutrient concentrations of ‘Newhall’ and ‘Skagg’s Bonanza’ navel oranges. Commun. Soil Sci. Plan. 40:3061–3076. doi: 10.1080/00103620903261627.
- Shimizu, T., H. Torikata, and S. Torii. 1978. Studies on the effect of crop load on the composition of Satsuma mandarin trees. V. Analysis of production processes of bearing and non-bearing trees on the carbohydrate economy. J. Japan. Soc. Hort. Sci. 46:465–478. doi: 10.2503/jjshs.46.465.
- Singh, J.K., J. Prasad, H.K. Singh, and A. Singh. 2008. Effect of micronutrient and plant growth regulators on plant growth and fruit drop in aonla (Emblica officinalis Gaertn.) fruits cv. ‘Narendra aonla-10ʹ. Plant Archives 8(2):91 1–913.
- Smith, D. 1969. Removing and analyzing total non-structural carbohydrates from plant tissue. Wisconsin: Wisconsin Univ., Leaflet No. 14.
- Spiegel-Roy, P., and E.E. Goldschmidt. 1996. Biology of Citrus. Cambridge University Press, Cambridge, U.K.
- Ward, D.L. 2004. Factors affecting pre-harvest fruit drop of apple. Ph.D thesis, Virginia Polytechnic Institute and State University. 143 p.
- Zhu, Q., M. Ozores-Hampton, Y. Li, K. Morgan, G. Liu, and R.S. Mylavarapu. 2017. Effect of phosphorus rates on growth, yield, and postharvest quality of tomato in a calcareous soil. HortScience 52(10):1406–1412. doi: 10.21273/HORTSCI12192-17.