
ABSTRACT
Fifteen newly-produced cacao hybrids were evaluated in the field with an objective of understanding their genetic diversity using morphological descriptors. The field trials were conducted in Ibadan (7° 13ʹN, 3° 51ʹE), and Owena (7° 11ʹN, 5° 1ʹE), Nigeria. Juvenile performance data were subjected to ANOVA, Principal Component and Cluster analyses. The first three principal axes accounted for 34.41%, 16.71%, and 14.09% of variation among the plant characters, respectively. Numerical taxonomy showed P2 x P11 as the most distinct. The FASTCLUS technique grouped the genotypes into four clusters, P2 x P11 being the only member of its group. There is significant genetic diversity among the genotypes, while plant height, stem diameter, Cherelle wilt, fruiting, and tree size among others are important discriminatory traits among cacao genotypes.
Introduction
Cacao is a commodity tree crop of immense scientific and economic importance grown predominantly in the tropics and sub-tropics. It is the source of the cacao bean which is a major source of income to the growers since the cacao bean is the raw material for the production of chocolate, which is consumed either directly or in confectionery mainly in the temperate countries. The economic and nutritional importance of the crop makes the choice of better performing genotypes useful for optimum field performance so as to make them desirable. More so, breeding for sustainable optimal performance is dependent on the availability and accessibility of intra-specific diversity of species. Adequate variability within a species can provide a genetic buffer for their survival in the face of unfavorable conditions (Adewale and Kehinde, Citation2014).
Germplasm characterization must necessarily precede selection for improvement purposes. Characterization of germplasm accessions is centered on the description of accession composition and morphological characteristics or phenotypic expression, while morphological characterization is an account of the plant morphology either through its lifespan or only at maturity (Adewale, Citation2012). Phenotypic traits with high heritability are very useful for cacao germplasm characterization (Bekele et al., Citation2006; Lachenaud and Oliver, Citation2005). Tezara et al. (Citation2015) stated that cacao genotypes showed variation in the expression of morphological traits, suggesting significant genetic variability resulting from a large number of crosses done to develop cacao clone in Ecuador.
The adoption of improved hybrids with high yield and bean quality potentials in Nigeria is very low, as Eskes (Citation2000) reported that farmers have continued to use their own trees on-farm as sources of materials for new plantings and rehabilitation of old plantations. The genetic base of cacao material on-farm in Nigeria is thus perceived to be narrow (Aikpokpodion, Citation2012). The search for resistance to the cocoa swollen shoot virus disease in Ghana and Nigeria led to the introduction of several materials belonging to the Trinitario (T-clones) and Upper Amazon Forastero populations into West Africa in 1940s (Aikpokpodion, Citation2007). The F3 Amazon, a derivative of the Upper Amazon Forastero material, found to exhibit a short juvenile stage (Aikpokpodion, Citation2007), is a common variety in Nigeria. The continuous use of these (T-clones and Upper Amazon Forastero) populations in cacao improvement has contributed significantly to disease resistance and adaptability to changing environment in Nigeria (Zhang and Motilar, Citation2016). However, with the predominant continuous use of these populations for new planting, the on-farm genetic diversity of cacao in Nigeria remains low (Zhang and Motilar, Citation2016) thereby requiring improved breeding strategies to provide farmers with enhanced genetic diversity. Therefore, new hybrids are needed to enhance on-farm diversity of cacao and the adoption of improved genetic materials.
In 2011, the Cocoa Research Institute of Nigeria (CRIN) released some new improved cacao varieties. These new varieties were involved in a hybridization programme which yielded 15 new hybrids. These 15 new hybrids were produced from hybridization procedures involving the T-clones discussed earlier. There was therefore the need to evaluate these newly-generated hybrids so as to establish the diversity among them, while also emphasizing the broadening of the genetic base of cacao germplasm in the country. The objectives of the study were to determine the phenotypic diversity in some cacao hybrids in Nigeria and the relative contribution of the qualitative and quantitative economic traits to the diversity observed among the hybrids. The genetic potentials in the genotypes for subsequent selection and improvement can thus be better understood.
Materials and Methods
Fifteen (15) newly generated cacao hybrids were established at two different cacao agro-ecologies: Ibadan (7° 13ʹN, 3° 51ʹE; a derived savanna agro-ecology), Oyo State and Owena (7° 11ʹN, 5° 1ʹE; a humid rain forest agro-ecology), Ondo State, Nigeria in July 2012. Land was selected for planting in a secondary forest and manually cleared. Selective tree felling was done such that trees in the lower storey were felled and the tall trees retained to provide shade for the young cacao seedlings. Plantain suckers were established a year ahead of field establishment of the cacao on the planting site to ensure that shade was sufficient ahead of the cacao crop. Holing was done to a depth of 15 cm. At the planting period, rainfall had stabilized at both locations. Sixty individual seedlings were established per genotype as 10 seedlings per plot in three replications in a Randomized Complete Block Design (RCBD) at each of the two locations. The list of the 15 genotypes and their pedigree is shown in . The spacing used was 2.5 × 2.5 m. The number of plant stands per plot corresponded to 450 stands per location, and 900 plants population that the study was based on. Manual weeding was carried out five times a year at 2-months interval during the rainy seasons. Irrigation water was supplied manually at 3-days interval during the dry seasons (between November and March) to prevent moisture stress on the crop.
Table 1. Cacao genotypes that were used in the study
Data were collected from five randomly sampled individual stands per genotype per replicate. Quantitative juvenile data collected for these genotypes included measurements of plant height and stem diameter and counting of the number of leaves at 3-months interval. As all the genotypes formed fan branches (called jorquette) in less than 12 months of field establishment, which is a pointer toward reproductive maturity, the data collection shifted from 12 months of field establishment to Time to Jorquette (Weeks), 3-monthly measurements of Jorquette Height (cm) and Tree Circumference (cm).
Qualitative data were scored using nominal codes (with modification) following the Short Descriptor List used by Cocoa Research Unit, Trinidad and Tobago (Bekele and Butler, Citation2000) adapted from International Board of Plant Genetic Resources (IBPGR Citation1981) Cacao descriptor. These are presented in . For leaf flush color, an average of four branches with flushes was observed for the intensity of anthocyanin in new terminal flushes. The anthocyanin intensity in the flush color (which if high in intensity indicates the dominance of the Criollo cacao populations, thereby presenting patterns for classification of the genotypes) was scored as indicated in . Data collection was terminated at 30 months of field establishment for all the genotypes. The means from the sampling unit per hybrid were subjected to analysis of variance (ANOVA) in the Statistical Analysis System, SAS-V9.2 (SAS Institute Inc., Citation2007) to estimate the variability among the genotypes.
Table 2. Descriptor list for qualitative traits used in the study
Means were obtained for each genotype across the three replications for the eleven parameters, resulting in a 15 × 11 basic multivariate data matrix. The data were standardized (mean = 0; standard deviation = 1) to eliminate the effects of the different scales of measurements among the parameters. The resulting product was subjected to Principal Component Analysis (PCA) as well as a Single Linkage Cluster Analysis (SLCA) in the Statistical Analysis System, SAS-V9.2 (Institute, Citation2007). The PCA produced an eigenvector for each axis, while a dendrogram was produced for the SLCA. The character loading was used to determine the genotype component scores. The FASTCLUS technique was used to classify the genotypes into distinct groups.
Results
The variance components and the coefficient of variation for 22 characters in the combined (pooled) locations are presented in . Significant (P ≤ 0.05) phenotypic variation existed among the 15 hybrids for time to jorquette, jorquette height (21 months after planting) and tree circumference (21, 24 27, and 30 months after planting), as well as fruiting (at 30 months after planting) (P ≤ 0.01). Hybrids that had fruited by 30 months after planting included P1 x P10, P1 x P11, P2 x P10, P3 x P10, P3 x P11, P3 x P9, P6 x P10, P7 x P8 and P7 x P10.
Table 3. Mean squares of characters of the 15 cacao genotypes in the combined locations
The main effect of the location was highly significant (P ≤ 0.05) on most of the twenty-two agronomic characters except tree circumference (15 and 30 months after planting), fruiting (27 months after planting), and Cherelle wilt (24, 27, and 30 months after planting). The variance component for interaction (i.e. genotype by location, GxL) had a significant (P ≤ 0.01) effect on only two of the twenty-two characters, i.e. tree circumference (21 months after planting) and fruiting (30 months after planting).
The eigenvalues of the Principal Component axes are presented in . The first, second, and third principal components accounted for 34.41%, 16.71%, and 14.09%, respectively, of the total variation among the 15 cacao hybrids. These three axes together accounted for 68.21% of the total variation among the eleven characters that described the 15 cacao hybrids in the diversity studies. In addition, these first three principal components had eigenvalues of 4.12, 1.84, and 1.55, respectively.
Table 4. Eigenvalues and factor scores of major characters associated with the first four principal component axes used in ordination of 15 cacao genotypes
The first principal component axis was loaded largely by stem diameter (0.43), tree circumference (0.40), tree size (0.44), and presence of fruits (0.39). The second principal component was loaded mainly by plant height (−0.54), as well as jorquette height (−0.54), while number of leaves (0.38) as well as canopy shape (0.33) were also important here. The third principal component was loaded mainly by canopy shape (0.50) loaded on it. Among other characters, Principal axis IV was loaded mainly by the presence of flush (−0.52), while time to jorquette (0.31), tree circumference (0.35), and the presence of fruits (−0.31) are important characters also that were loaded there.
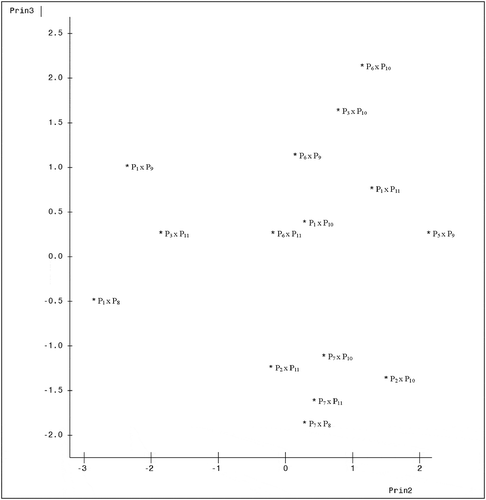
A plot of the genotypes on axes 1 and 2 which accounted for 52.84% of total variation is presented in . The figure showed how widely separated the genotypes were from one another. Genotypes 6 (P2 x P11) and 1 (P1 x P8) appear clearly distant from the rest, while genotypes 3 (P1 x P10) and 13 (P7 x P8) clearly were the most closely related among the 15 genotypes. A plot of axes 1 and 3 which jointly accounted for 48.51% of total variation () still shows genotype 6 (P2 x P11) as well distant from the rest. Genotypes 3 (P1 x P10), 8 (P3 x P11) and 12 (P6 x P11) are the closest here. , which showed the graph of axes 2 and 3, also expresses the distinctness of all the 15 genotypes from one another, and genotypes 5 (P2 x P10), 6 (P2 x P11), 13 (P7 x P8), 14 (P7 x P10), and 15 (P7 x P11) all appeared to have formed a cluster, suggesting similarities among them.
Figure 1. Configuration of fifteen cacao genotypes under axes 1 and 2
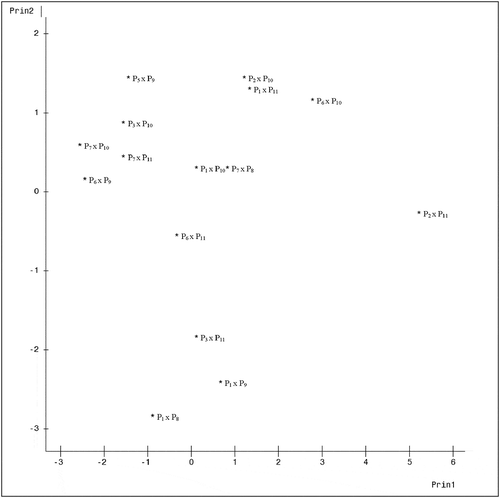
Figure 2. Configuration of fifteen cacao genotypes under axes 1 and 3
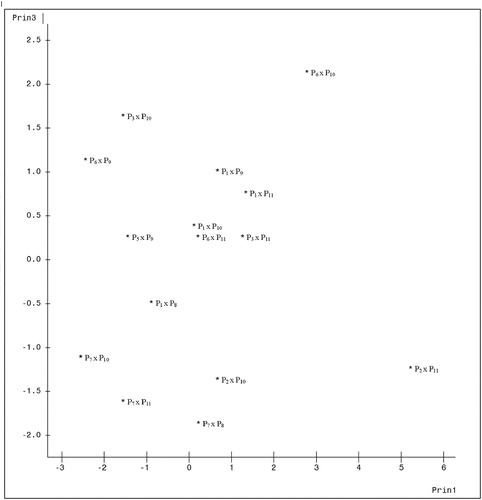
Figure 3. Configuration of fifteen cacao genotypes under axes 2 and 3
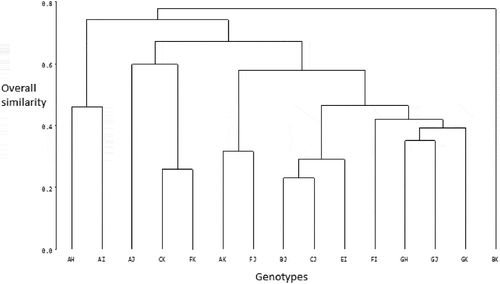
shows the dendrogram drawn from the Single Linkage Cluster Analysis (SLCA) to illustrate the relationship among the 15 cacao hybrids on the basis of overall similarity. At the 0% level of similarity, all the genotypes were distinct from one another. In the expression of similarity among all the genotypes, the first cluster was formed between genotypes P2 x P10 (BJ) and P3 x P10 (CJ) at the 22.95% level of similarity. Cluster 2 was formed between genotypes P3 x P11 (CK) and P6 x P11 (FK) at the 26% level of similarity. Genotype P5 x P9 (EI) was joined to cluster 1 at the 29.17% level of similarity to form cluster 3. Genotypes P1 x P11 (AK) and P6 x P10 (FJ) formed a cluster at the 31.62% level of similarity and this cluster was joined to the cluster among genotypes P2 x P10 (BJ), P3 x P10 (CJ), P5 x P9 (EI), P6 x P9 (FI), P7 x P8 (GH), P7 x P10 (GJ), and P7 x P11 (GK) at the 57.73% level of similarity. Genotypes P1 x P8 (AH) and P1 x P9 (AI) formed a cluster at the 46.09% level of similarity and this cluster was joined to the existing clusters at the 67.06% level of similarity. At the 74% level of similarity, 14 of the genotypes had already been joined in different clusters. Thus, the 14 genotypes in the existing clusters were joined by the most distinct genotype P2 x P11 (BK) at the 77.7% level of similarity to form a single cluster, indicating that the genotypes had at least one neighbor with more than 77% similarity.
Figure 4. Dendrogram resulting from the Single Linkage Cluster Analysis of fifteen cacao genotypes used in the study
The genotypes were classified into four distinct groups, as shown in by the FASTCLUS technique. Group IV had only one genotype (P2 x P11), obviously the most distinct. Group I had two genotypes (P1 x P8 and P1 x P9), group II had seven genotypes, the highest number, while group III had five genotypes. Groups I and IV had tall plants. Group IV consisted of tall, big, and vigorous plants as shown in the highest value for plant height (106.97), stem diameter (2.77), number of leaves (45.70), tree circumference (11.53), and tree size (3.27) and also had a significant presence of flush color (2.20). The hybrid in this group also had the least time to jorquette (31.83), an indication of earliness to reach the reproductive phase, and eventually yielded the highest number of fruits (1.20) at 30 months of field establishment. Group II was composed of the least vigorous hybrids as shown in the least value for plant height, stem diameter, and tree size. The hybrids in this group also took the longest time to jorquette formation and ranked among the least in fruit production. Groups I and III were composed of hybrids with moderate vigor as shown in their plant height, stem diameter, tree circumference, and tree size.
Table 5. Major characteristic pattern of four groups of the 15 cacao genotypes (combined data) showing their mean values and coefficient of variation in parenthesis
Discussion
Most of the plant characters morphologically differentiated the 15 cacao hybrids. This was also observed in the performance of the hybrids across the two locations. The occurrence of specific variability in these hybrids assures of evolutionary survival and the possibility of improving the species based on characters with significant variation. The moderate values of the coefficients of variation for some of the quantitative traits further validate the genetic variability of the cacao hybrids. The variation in the performance of the genotypes across both locations suggests the need to emphasize breeding for adaptation to a specific environment. Araque et al. (Citation2012) and Padi et al. (Citation2013) reported variations among cacao families for vigor and seedling survival and other physiological traits across varied weather conditions. Since selection among species is preceded by observed variation, an in-depth understanding of the variation among these hybrids is key to their improvement. As Principal Component Analysis (PCA) highlights similarities and differences among variables, identifies the variation pattern, and assesses the relative contribution of different variables to each principal component (PC) (Adewale, Citation2012), the proportion of total variation among the plant characters accounted for by the first PC axis (37.41%) attests to the strength of the analysis. The relative discriminatory power of the principal axes clearly indicates the strength of the first axis in discriminating among the 15 cacao hybrids. This suggests that traits loaded on it played significant roles and are the most important in describing the variation among the cacao hybrids, since PCA measures axes along which variation between genotypes is maximized (Adewale, Citation2012). In essence, key agronomic traits such as stem diameter, tree circumference and tree size (which describe growth vigor) were significant in describing variation among the hybrids, since they were mainly loaded in the first principal component. The configuration of the genotypes under axes 1 and 3 suggests some degree of closeness among genotypes P1 x P10 and P7 x P8 while genotype P2 x P11 is the most distinct from the rest. In this study, different characters contributed differently to the total variation as indicated by the eigenvectors, weight, and loadings in the four PC axes. The most important characters in the classification of these cacao hybrids were plant height, stem diameter, number of leaves, time to jorquette, jorquette height, tree circumference, tree size, canopy shape, flush color, and fruiting since most of them were significantly loaded on the first Principal Axis. These traits are also believed to be under genetic control (Martinez et al., Citation2015). Among other characters, flush color is a very simple and identifiable trait on the field for genotype differentiations. Pigmentation of plant parts has been reported to be under a genetic factor in Abelmoschus caillei (Kehinde and Adeniji, Citation2003).
The grouping of the 15 genotypes by the Single Linkage Cluster Analysis (SLCA) indicated that each genotype had at least one neighbor with more than 77% level of similarity, but also showing genotype P2 x P11 (BxK) as the most distant from the rest. The FASTCLUS grouping of the genotypes into four distinct clusters further depicts genotype P2 x P11 as the most distinct from the rest as it was the only member of group IV, and had the highest mean for most of the traits that discriminated among the hybrids. It is strongly believed that vigorous juvenile performance can enhance earliness in fruiting, which essentially indicates a reduced juvenile phase, and improved yield in cacao. Therefore, this vigorous hybrid can be considered for selection for further cacao improvement. Padi et al. (Citation2012) recommended juvenile tree growth rate as an index for selecting high yielding cocoa families. One basic assumption in the use of principles of numerical taxonomy is that numerical evaluation of overall similarity between accessions is a measure of genetic similarity (Sneath and Sokal, Citation1973). The SLCA revealed a range of genetic diversity among the cacao hybrids.
The conclusion from this study is that adaptation to specific locations is important for successful cacao hybrid establishment. Furthermore, based on juvenile morphological characterization, there are sufficient variabilities among the cacao hybrids present in Nigeria. Therefore, selection for subsequent cacao improvement can be done among the genotypes in this study.
References
- Adewale, B.D. 2012. Diversity, genetic stability and reproductive biology of African yam bean (Sphenostylis stenocarpa (Hochst. ex A. Rich.) Harms). Federal University of Agriculture, Abeokuta, Nigeria, PhD Thesis.
- Adewale, B.D., and O.B. Kehinde. 2014. Inheritance and stability of some agronomic traits of African Yam Bean (Sphenostylis stenocarpa (Hochst Ex. A. Rich) Harms). p. 349–359. In: K.O. Omokhafe and J.O. Odewale, eds.. Genetics: Public awareness and national transformation. Proceedings of the 38th annual conference of the Genetics Society of Nigeria, 19th-23rd Oct. Genetics Society of Nigeria, Benin City, Nigeria.
- Aikpokpodion, P.O. 2007. Genetic diversity in Nigerian cacao (Theobroma cacao L.) collections as revealed by phenotypic and simple sequence repeats marker. University of Ibadan, Nigeria, PhD Thesis, p. 131
- Aikpokpodion, P.O. 2012. Defining genetic diversity in the chocolate tree, Theobroma cacao L. grown in West and Central Africa. In: M.C. Alis-Kan, ed. Genetic diversity in plants, Chap.10. InTech. doi: 10.5772/33101.
- Araque, O., R.E. Jaimez, W. Tezara, I. Coronel, R. Urich, and W. Espinoza. 2012. Comparative photosynthesis, water relations, growth and survival rates in juvenile criollo cacao cultivars (Theobroma cacao) during dry and wet seasons. Exp. Agric. 48(4):513–522. doi: 10.1017/S0014479712000427.
- Bekele, F.L., and D.R. Butler. 2000. Proposed list of cocoa descriptors for characterization. In: A.B. Eskes, J.M.M. Engels, and R.A. Lass, eds.. Proceedings of the CFC/ICCO/IPGRI Project Workshop on Working Procedures for Cocoa Germplasm Evaluation and Selection 1-6th February 1998, IPGRI Montpellier, France. 41–44
- Bekele, F.L., I. Bekele, D.R. Butler, and G.G. Bidaisee. 2006. Patterns of morphological variation in a sample of cacao (Theobroma cacao L.) germplasm from the International Cocoa Genebank, Trinidad. Genet. Res. Crop Evol. 53:933–948. doi: 10.1007/s10722-004-6692-x.
- Eskes, A.B. 2000. International cocoa breeding and selection initiatives. INGENIC Newsl. 5:2–3.
- IBPGR Secretariat. 1981. Report: International Board for Plant Genetic Resources (IBPGR) working group on genetic resources of cacao. ACP, IBPGR/80/56, Rome, Mar.
- Kehinde, O.B., and O.T. Adeniji. 2003. Inheritance of pigmentation in West African Okra (Abelmoschus caillei) (A. Chev.) Stevels. ASSET 3:1–6.
- Lachenaud, P.H., and G. Oliver. 2005. Variability & selection for morphological bean traits in wild cacao trees (Theobroma cacao L.) from French Guiana. Genet. Res. Crop Evol. 52:225–231. doi: 10.1007/s10722-004-4005-z.
- Martinez, I.B., M.V. de la Cruz, M.R. Nelson, and P. Bertin. 2015. Morphological characterization of traditional cacao (Theobroma cacao L.) plants in Cuba. Genet. Res. Crop Evol. 62:8.
- Padi, F.K., P. Adu-Gyamfi, A. Akpertey, A. Arthur, and A. Ofori. 2013. Differential response of cacao (Theobroma cacao) families to field establishment stress. Plant Breed. 132(2):229–236. doi: 10.1111/pbr.12039.
- Padi, F.K., S.Y. Opoku, B. Adomako, and Y. Adu-Ampomah. 2012. Effectiveness of juvenile tree growth rate as an index for selecting high yielding cocoa families. Sci. Hortic. 139:14–40. doi: 10.1016/j.scienta.2012.02.039.
- SAS Institute. 2007. The SAS system for Windows. Release 9.2. SAS Inst., Gary, NC.
- Sneath, P.H.A., and R. R. Sokal. 1973. Numerical taxonomy: The Principles and Practice of Numerical Classification. San Francisco: Freeman, 573 pp.
- Tezara, W., J. De Almeida, E. Valencia, J. Cortez, and M. Bolaños. 2015. Photochemical activity of cocoa elite clones Theobroma cacao L, Ecuadorian in the north of the province. Emeralds Invest Knowl. IV(3):37–52.
- Zhang, D., and L. Motilar. 2016. Origin, dispersal and current global distribution of cacao genetic diversity, p. 3–31. In: B.A. Bailey and L.W. Meinhardt, eds.. Cacao diseases: A history of old enemies and new encounters. Springer, Switzerland.