
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
The effects of NaCl salinity on the seed germination, growth, and some physiological characteristics in ‘Akbari,’ ‘Akbari’ hybrid (P. integrrima × ‘Akbari’), ‘Qazvini,’ and ‘Qazvini’ hybrid (P. integrrima × ‘Qazvini’) were investigated. The experiment was conducted with three salinity levels (0, 100, and 200 mM) as NaCl. The results indicated that the highest germination percent in salt-treated seeds was observed in ‘Akbari.’ Prolin and Na concentrations in the leaves were significantly (P ≤ 0.01) increase by salinity regardless of the various cultivars. Vegetative growth in terms of rootlet, stem and leaf number, fresh weight, dry weight were reduced by salinity. However, the reduction of growth was varied with cultivars. Leaf number, fresh and dry weight of ‘Akbari’ and ‘Akbari’ hybrid in salinity treatments was higher than that of treated two other cultivars. Finally results were showed that although ‘Akbari’ showed higher tolerance to salinity but significant differences were not observed between ‘Akbari’ and ‘Akbari’ hybrid rootstocks. Hence ‘Akbari’ hybrids could be appropriate candidates for researches and cultivation of pistachio in the saline areas.
Introduction
The genus Pistacia L. belongs to the Anacardiaceae family that comprises about 83 genera and 860 species. This genus involves at least 11 trees and shrub species. Among these species, P. vera L. is the only economically cultivated species, whereas the P. atlantica, P. integerrima, and P. terebinthus are used as rootstocks for P. vera (Sheikhi et al., Citation2019). P. vera is cultivated in the arid and semi-arid areas of Iran and is referred to as the “green gold” due to its abundant health-benefiting nutrients and commercial importance (Akbari et al., Citation2018a; Aliakbarkhani et al., Citation2017; Talebi et al., Citation2016). Being the most economically important horticultural crop, pistachio has the most cultivation area among all horticultural crops in Iran (Aliakbarkhani et al., Citation2015; Taghizadeh-Alisaraei et al., Citation2017).
Salinity and water deficit in pistachio orchards of Iran reduce the yield and productivity and causes billions of economic losses per annum (Goharrizi et al., Citation2020; Khoyerdi et al., Citation2016). Soil salinization is a severe menace for crop productivity with devastating global effects, estimated at 50% of the land loss by 2050 (Akbari et al., Citation2020a). Saline soils, commonly known with electrical conductivity of more than 4 dS/m and pH < 8.2, are encompassing about 41 million ha (25%) of Iran’s lands (Mesgaran et al., Citation2017). One of the most important characteristics of the pistachio trees is the ability to store large amounts of sodium in the roots and stems (Picchioni et al., Citation1990). Although the species used as pistachio rootstock are considered semi-tolerant to saline and poor-quality soils, the appropriate commercial production has been occurred in deep, well-drained, sandy loam soils. (Akbari et al., Citation2018b; Jamshidi Goharrizi et al., Citation2019). It has been reported that salinity causes the reduction in fruit tree biomass, canopy height, trunk diameter, leaf area, leaf number, and overall yield (Ferguson et al., Citation2002; Bastam et al., Citation2013).
Roots are considered as an anchor for the plants in the soil, absorbing water and nutrients; storage organ, and are the primary zone of contact with the soil elements and organisms. These organs are a fundamental part of coordinating the plant response to biotic and abiotic stresses. However, their systems differ significantly in construction and function, both within and between species (Warschefsky et al., Citation2016). In the long-lived woody plants, such as pistachios, grafting is the common way to clonally propagate desirable scions (cultivars), and for its cultivation, choosing an appropriate rootstock is an indispensable necessity (Del Carmen Gijón et al., Citation2010). It has been indicated that leaves are more sensitive to salinity than stems (Karimi et al., Citation2009; Mohamad Khani, Citation1997). Moreover, it has been demonstrated that the sensitivity of pistachio leaves and stems to salinity are more than its underground organs (Adish et al., Citation2010), which indicates the importance of rootstocks in alleviating the adverse effects of salinity stress on pistachio growth.
Tavallali et al. (Citation2008) demonstrated that NaCl salinity, 150 mM, g with different concentrations of calcium sulfate in P. vera (cv. ‘Qazvini’ and ‘Badami zarand’) rootstocks reduced the relative water content of leaves in ‘Badami Zarand’ while has no significant decrease in ‘Qazvini’ rootstock. In addition, 150 mM NaCl significantly increased the electrolyte leakage percentage in ‘Badami Zarand,’ also salt treatments significantly increased the concentration of sodium in the leaves and roots of ‘Badami Zarand,’ reducing potassium content in the shoots. The tolerance of different P. vera rootstocks to saline and drought conditions have been demonstrated in several studies (Khoyerdi et al., Citation2016; Akbari et al., Citation2018a; Akbari et al., Citation2020a) and was shown to be significantly varying among the domesticated pistachio rootstocks. However, some literature now exists concerning the superiority of ‘Qazvini” (Hokmabadi et al., Citation2005), ‘Akbari’ (Adish et al., Citation2010; Akbari et al., Citation2018a; Goharrizi et al., Citation2020), and UCB-1 cultivar (Raoufi et al., Citation2020a) to salinity stress over the other P. vera rootstocks. However, in new researches, it has been determined that ‘Akbari’ rootstock is more tolerant to salinity than other rootstocks (Raoufi et al., Citation2019).
In the salinity challenges coupled with poor drainage condition, sustainable long-term productivity is requiring improved new rootstocks with more salt tolerance (Akbari et al., 2018). Strong vigor and rapid growth along with tolerance to abiotic stress (salinity, drought, cold) are the most important objectives of pistachio rootstock breeding (Sheikhi et al., Citation2019). Efforts toward producing vigorous rootstocks with acceptable tolerance to salinity are continuing. Among the species commonly used as rootstocks (P. atlantica, P. integerrima P. terebinthus, and P. vera) the P. integerrima is the most vigorous rootstock, by the way, the tolerance of this rootstock to abiotic stress is not acceptable (Ferguson et al., Citation2005).
University of California hybrid pistachio rootstock (UCB-1) has been developed in university of californa pistachio hybrid rootstock breeding program. This rootstock is the result of a specific P. atlantica female pollinized by a specific P. integerrima male (Sheikhi et al., Citation2019). UCB-1 has shown to have more growth and salinity tolerance than its parents do, by the way, this rootstock is not commonly used in Iran due to its susceptibility to winter cold and cannot tolerate the winter cold as the P. vera (Ferguson and Haviland, Citation2016). Besides, in the cold areas the cultivars grafted on UCB-1 usually do not fall asleep compared to those grafted on P. vera, P. atlantica, and P. terebinthus, and are very susceptible to early cold weather from October through early December, causing winter juvenile tree dieback.
In this research, the ‘Qazvini’ and ‘Akbari’ were used as maternal parents and the P. integerrima species used as paternal. However, the salinity tolerance of ‘Qazvini’ and ‘Akbari’ and their hybrids with P. integerrima were investigated.
Materials and Methods
Plant Materials and Study Plan
This experiment was carried out in Buin Zahra, Qazvin, Iran (50º 4‘E and 35 º46‘ N 1210 m). The climate of this region is temperate tends hot and dry. Also, Laboratory works were carried out in Shahed University. Tehran, Iran. To produce the hybrid rootstocks seeds, pollination were carried out in two phases during the full bloom period. The. ‘Qazvini’ and ‘Akbari’ were used as maternal parents and P. integerrima species used as paternal.
In order to produce hybrid seeds, the pollens of P. integerrima were collected and stored in a freezer at −20°C and used for the female flowers protected in the pollination bags (). Salinity tolerance of hybrids and female parents was studied prior to pollination, the viability test of P. integerrima pollen was investigated using the IKI method (Gunver-Dalkilic and Dayl-Dogru, Citation2011). In each crossing, many seeds were obtained and 12 seeds were planted in each replication to decrease hetroziogosity effects.
Figure 1. A protected female flower in pollination bag

The study was designed as two separated parts; first at the germination stage and the second in the seedling stage of the plant growth. To investigate the tolerance of hybrids and female parents in the germination stage, a factorial experiment was designed based on a completely randomized design with three replications with 12 seeds in each of them to decrease heteroziogosity effects. For the seedling stage, the same study was designed with four replications by 12 plants in each of them. The first factor comprised four rootstocks (‘Qazvini,’ hybrid of P. integrrima × ‘Qazvini,’ ‘Akbari,’ and P. integrrima × ‘Akbari’); the second factor involved three different NaCl salinity concentrations (0, 100, and 200 mM in irrigation water). After harvesting, the pistachio hybrid seeds and the parent’s seeds were immersed in water for 24 hours before planting and disinfected.
Germination Experiment
First, the seeds were sterilized with Benomyl fungicide at a concentration of 3000 ppm for 20 minutes and sodium hypochlorite at a dose of 10% for 10 minutes to prevent fungal attack. Germination experiment was carried out in 10 cm diameter petri plates. A filter paper was placed in the bottom of each petri, 12 seeds were placed on every petri, and with a filter paper was covered (,). Salinity treatments were performed by adding 10 mL of NaCl solution every with concentrations of 0, 100, and 200 mM, and each weak filter papers were replaced. The petri plates were sealed by parafilm in order to protect the humidity and avoid the seeds to face drought stress.
Figure 2. The pistachio germinated seeds

The seeds were immediately transferred to the growth chamber with a constant temperature of 25°C and sufficient light. The germination experiment continued for 30 days and on day 30, the number of germinated seeds were recorded and the rootlets length were measured.
Cultivation Situation
The germinated seeds were planted in polyethylene pots containing a mixture of 50% medium grain perlite and 50% washed pumice. Seedlings were irrigated by half-strength Hoagland nutritional solution for 2 months. Salinity treatments were started 2 months after planting and continued for 2 months. Plants were irrigated every 3 days (each time 1 l each pot), in each irrigation time, approximately 200 ml exited at the bottom of the pots and remained in the pot trays. After salinity treatments, the following growth and physiological parameters were measured:
Growth Characteristics
In order to evaluate the effects of different salinities on growth parameters, seedling height, number of leaflets, and seedlings diameter were measured in the pots in two stages, 15 days and 50 days after the beginning of salinity treatments (). In addition, at the end of the experiment, the root, stem and leaf fresh weight, and leaf area were measured using the leaf area meter, and after drying the samples in an oven (70°C after 24 hours) dry weight were measured using a digital scale.
Figure 3. Measuring the diameter and root length

Relative Water Content
In order to measure the relative water content of the leaves, the fresh weight (FW) of well-expanded leaves were immediately determined. The samples were then placed in the water at room temperature and 24 hours light and turgid weight (TW) was measured. The samples were dried in the oven at 70°C for 24 hours and their dry weight (DW) were determined (Barrs and Weatherley, Citation1962). At the end, the relative water content of leaves was calculated using the following formula:
Electrolyte Leakage
To measure electrolyte leakage, two fresh leaves were taken as samples. About 0.2 grams of each leaves were weighted and washed by distilled water and placed into covered test tubes, 10 ml of distilled water was added to it and placed on a 120 rpm shaker for 24 hours at laboratory temperature. After 24 hours, their initial electro conductivity (EC1) were measured by a digital EC meter. The samples were then placed in a 90–100°C hot water bath for 20 minutes and after cooling, their secondary electro conductivity (EC2) was measured (Lutts et al., Citation1996). Finally, Electrolyte leakage percentage (EL) was obtained by the formula (No 2):
Sodium and Potassium Content
The concentration of Na and K was measured in the leaves. About 0.1 g milled leaves were weighed from each sample and transferred to the Kjeldahl digestive tubes. Then, 10 ml of concentrated nitric acid (65%) was added to each tube. Two tubes without plant samples and only nitric acid were used as control. Samples were stored overnight (12 hours) at room temperature. Then, the samples were kept in the digested oven for 3–4 hours at 110°C until the sample's color was clarified (yellow amber). About 20 ml of distilled water were added to each tube and filtered by a filter paper and the final volume reached 100 ml in the balloon. The obtained extract was used to measure Sodium and Potassium amounts. The amount of Potassium and Sodium in the samples was read by a Flame Photometer apparatus. Potassium to Sodium ratio was calculated. The ratio of potassium to sodium can be considered as resistance to salinity criterion
The Proline Content
The leaves’ free proline content was measured according to the method described by Bates et al. (Citation1973). The amount of 0.5 g fresh weight, including fresh leaf texture, was mixed and grinded with 10 ml of 3% sulfosalicylic acid and centrifuged at 3,000 rounds for 5 minutes. Then, 2 ml of supernatant, 2 ml Ninhydrin acid, and 2 ml acetic acid were added to the test tube. The test tubes were placed in a hot water bath at a temperature of 90–100°C for 1 hour and after cooling, 10 ml of Toluene was added to each test tube and were shaken for 15–20 seconds. After cooling at room temperature for 30 minutes, two separate layers were formed. The wavelength of the red phase formed at the top of the tubes was recorded at 520 nm by a spectrophotometer (Bates et al., Citation1973)
Data Analysis
The experiment was carried out based on a completely randomized factorial design with three replicates in germination and four replicates in the seedling stage respectively. The rootstocks have been chosen as the first factor and three levels of NaCl concentrations (0, 100, and 200 mM) as the second factor. The data are presented as mean ± SD and the means were compared by Duncan’s multiple range test to discover the values and differences. The SAS software v. 9.01 was used to test the Normality and analyzing the data and the graphs were generated by Excel software (2016).
Results and Discussion
Germination Percentage and Root Length
The effects of salinity on germination percentage of rootstocks were significant (P < .01); however, their interaction (salinity levels and rootstocks) was not significant. Comparing rootstocks () indicated that the ‘Akbari’ hybrid (P. integrrima × ‘Akbari’) had a lower germination percentage than the other studied rootstocks. Moreover, in all rootstocks a significant decrease in germination percentage was observed by increasing salinity level (). On average, the germination percentage changed from 55% in the control group to 31.67% in 100 mM and 13.33% in 200 mM. The reduction in germination percentage in 100 mM (equivalent to EC about 7.3 dS/m) was about 42.5% and in 200 mM (equivalent to EC about 14.6 dS/m) was 75.8%. The results are in accordance with the results of Hokmabadi et al. (Citation2005) indicating P. vera cv. ‘Badami zarand’ 50% reduction in germination percentage at salinity level of 96 mM NaCl.
Table 1. The mean comparison of variety effect on physiological and morphological characteristics
Salinity significantly affected the rootlet length of plants so that the increased salinity decreased the rootlet length (). The rootlet length changed from 2.61 cm in the control treatment to 0.74 cm in 100 mM and 0.14 cm in 200 mM salinity treatments. The rootlet length of ‘Akbari’ and ‘Qazvini’ is significantly superior in comparison with two hybrid rootstocks. This clearly shows that domesticated pistachio rootstocks have most likely better rootlet growth than nondomesticated ones. Rootlet growth decreased in the second and third salinity level by 71.6% and 94.6%, respectively.
Growth Characteristics
Vegetative growth characteristics were affected by the salinity treatments (). Stem height and diameter, the number of leaves after 15 and 50 days, leaf area, fresh and dry weight of roots, were significantly affected by salinity (). Moreover, the interaction of rootstock and salinity affected the final stem height, leaf number, leaf area, leaf fresh weight, stem, and leaf dry weight. Among the studied rootstocks, ‘Akbari’ and ‘Akbari’ hybrid showed maximum final stem diameter, the final number of leaves, stem fresh weight, root, and stem dry weight compared to other rootstocks ().
Figure 4. Effect of salinity levels on the pistachio rootstock rootlet length
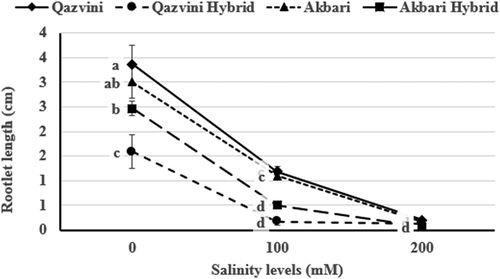
Table 2. The mean comparison of variety Effect on physiological and morphological characteristics
Table 3. The mean comparison of variety effect on physiological and morphological characteristics
Table 4. The mean comparison of salinity level effect on physiological and morphological characteristics
Table 5. The mean comparison of salinity level effect on physiological and morphological characteristics
Raoufi et al. (Citation2019) revealed that ʽAkbari,’ ʽAhmad-Aghai,’ and ʽUCB-1ʹ had the lowest reduction in morphological and growth parameters under salinity stress. Also, they showed that these cultivars had the highest chlorophyll content, SPAD index, and soluble sugar content under salinity conditions. These rootstocks by osmoregulation, protector substances, and balancing of toxic ions had better growth and morphological characteristics in saline condition.
Salinity decreases the leaf area and number of leaves by affecting the amount of cell division or by affecting cell enlargement size (Rahneshan et al., Citation2018). This decrease might be because of the increased costs for the plant’s cells for producing and accumulating the osmotic regulatory materials. The increased energy consumption causes the cells to receive less energy for division and as a result their growth will reduce (Volkmar et al., Citation1998). Literatures have shown that the growth of developing young leaflets depends on cell division, followed by the irreversible increase in the cell size. Growth often depends directly on turgid, and the development is related to biochemical processes that alter the properties of the cell wall (Dubey, Citation1997). The relative salinity tolerance in pistachios is probably due to the growth reduction in the shoots, especially leaves, in order to reduce the evapotranspiration and increase the root resistance against ion displacement (Benmahioul et al., Citation2009). Our results indicate that the fresh and dry weight of the roots was not affected by the interaction between the rootstock and the salinity. It has been demonstrated that the sensitivity of leaves and stems of pistachios to salinity is higher than the plant’s underground organs (Adish et al., Citation2010; Karimi et al., Citation2009; Picchioni et al., Citation1990).
Stem height was not influenced by 100 mM salinity level after 15 days of stress and significantly decreased of 200 mM. However, after 50 days of stress, the effect of 100 mM was also significant. Stem diameter was not affected by salinity after 15 days in all studied rootstocks; however, after 50 days, the rootstocks showed a different response to salinity stress. Regarding the number of leaves, the interaction effect of variety and salinity significantly affected the studied values after 50 days.
Stem height after 50 days, leaf number after 50 days, leaf area, fresh weight of root, stem and leaf, dry weight of root, stem, and leaf have significantly decreased by increasing the salinity levels. As a result, the salinity level of 100 mM has been the threshold of reducing the mentioned growth characteristics. Increasing the concentration of NaCl in irrigation water to more than 75 mM causes a significant reduction in the total biomass of pistachio (Hokmabadi et al, Citation2005). Other studies have also demonstrated that the average salinity of the pistachio root development zone for a 50% reduction in vegetative growth is between 7.9 and 10 dS/m (Sepaskhah and Maftoun, Citation1988). Significant decrease in stem and root growth in electrical conductivity of 8.7 dS/m was reported by Picchioni et al. (Citation1990).
Since the rootstock of ‘Akbari’ and ‘Akbari’ hybrid has significantly exceeded in the growth characteristics, it might be inferred that these rootstocks can be introduced as semi-tolerant rootstocks to saline conditions.
Physiological Characteristics
Relative Water Content of Leaves (RWC)
Results indicate that the rootstocks have different RWC in response to salinity (). Among the studied rootstocks ‘Qazvini’ had the minimum RWC in the leaf than other rootstocks. According to Balaguer et al. (Citation2002), the lower reduction in RWC could be due to the sufficient amount of osmotic regulation in the plant. More the plant could resist under stress conditions to decreasing the RWC of the leaf, it is considered as a plant that is resistant to salinity and drought. Hence, in the present study, it was concluded that pistachios resist to lose water through leaves in the conditions of salinity stress.
Similar to our findings, Hokmabadi et al. (Citation2005) demonstrated that the relative leaf water percentage in different salinity treatments was not significantly different. In another on Pistacia mutica spices, P. vera cv. ‘Sarakhs’ and P. vera cv. ‘Badami’ no significant difference with increasing salinity from 0 to 150 mM was observed in Badami and Sarakhs rootstocks (Panahi, Citation2009).
Electrolyte Leakage Percent
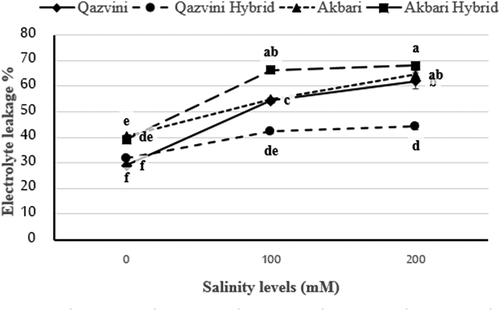
Based on the results of the analysis of variance, the effects of rootstock, salinity levels, and their interaction were significant in changing electrolyte leakage. ‘Qazvini’ hybrid (P. integrrima × ‘Qazvini’) had the lowest percentage of electrolyte leakage with an average of 39.4% among the four rootstocks under testing (). With increasing salinity, electrolyte leakage was significantly increased (). The electrolyte leakage percentage was 35.13% in the control, in the second level of salinity was 54.44% and in the third salinity level was 59.71%. shows the effects of different salinity levels on the percentage of electrolyte leakage in studied pistachio rootstocks. In accordance to our finding, Tavallali et al. (Citation2008) investigated the effects of salinity stress in the ‘Qazvini’ and ‘Badami Zarand’ pistachio rootstocks and demonstrated that 150 mM NaCl treatment significantly increased the electrolyte leakage percentage in Badami rootstock, while ‘Qazvini’ had a lower percentage of electrolyte leakage (Tavallali et al., Citation2008).
Figure 5. Effect of salinity levels on electrolyte leakage percent of pistachio rootstocks
Sodium Content of the Leaves
According to , no significant difference was observed among the studied rootstock; however, salinity stress levels significantly increased the Sodium content of the Leaves. With increasing salinity levels, the amount of leaf sodium increased (). So that the amount of sodium in leaf dry weight was increased from 6.06 mg/g in the control group to 8.56 mg/g in the second salinity level and 11.38 mg/g in the third level of salinity.
Table 6. The mean comparison of salinity level effect on physiological and morphological characteristics
Literatures indicate that with increasing salinity, sodium concentration increases in pistachio above-ground organs (Behboudian et al., Citation1986; Ferguson et al., Citation2002; Tavallali et al., Citation2008). The results of Hokmabadi et al. (Citation2005) on the effect of different levels of salinity on P. vera cv. ‘Badami Zarand,’ P. vera cv. ‘Sarakhs,’ and P. vera cv. ‘Qazvini’ rootstocks showed that ‘Qazvini’ rootstock has the lowest amount of sodium in stem among the studied rootstocks (Hokmabadi et al., Citation2005).
Potassium Amount of the Leaves
The results showed that potassium content in the leaves was different among the studied rootstocks, and effects of salinity levels were not observed to be significant. ‘Qazvini’ rootstock with 23.39 mg/g of dry weight in the leaves had the highest amount of potassium in the leaf ().
The effects of salinity on growth and element uptake in pistachio rootstocks had no effects on leaf potassium content (Picchioni et al., Citation1990). This indicates that the tolerance of pistachio to salinity might be attributed to its ability to preserve the high capacity of sodium in the roots (Picchioni et al., Citation1990). Additionally, the results of Karimi et al. (Citation2015) revealed that potassium content in the shoots was not affected by salinity treatments. The results have shown that the potassium content of stems and roots decreased by increasing NaCl concentration but did not affect the concentration of potassium in the leaf. In consistency with our finding, Tavallali et al. (Citation2008) demonstrated that ‘Qazvini’ had a lower leaf Sodium accumulation and higher potassium content than Badami Zarand (Tavallali et al., Citation2008).
Potassium to Sodium Ratio
The results showed that the rootstock type and salinity stress levels significantly affect the potassium to sodium ratio. This ratio significantly decreased by increasing the salinity levels () ranging from 2.87 in the control group to 1.97 in the second salinity level and to 1.54 in the third salinity level. The highest ratio of potassium to sodium was observed in the ‘Qazvini’ rootstock ().
The ratio of potassium to sodium can be considered as a criteria for tolerance to salinity . With increasing salinity levels, potassium amount, and potassium/sodium ratio have been deceased in pistachio leaf and root (Banakar and Ranjbar, Citation2010). Sodium competes with potassium to absorb root cells in the salty soils. The high sodium/potassium ratio reduces the growth and toxicity of the plants. Sepaskhah and Maftoun (Citation1988) reported that by increasing salinity, the potassium content of almond and hazelnut tended to increase. In addition, the potassium to sodium ratio in the ‘Badami’ rootstock was higher than two other rootstocks (Sepaskhah and Maftoun, Citation1988). Picchioni et al. (Citation1990) showed that increasing the amount of salt due to increased sodium content in the leaf has been decreased potassium to sodium content (Picchioni et al., Citation1990).
Raoufi et al. (Citation2020b) reported that salt stress induces the absorbance of Na and Cl and reduces K concentration. In their study, ion concentration in different organs of pistachio rootstocks increased by increasing salinity levels. ‘Akbari’ kept the low level of Na in the leaves while UCB-1 contained a higher K level in the leaves. Akbari et al. (Citation2018b) reported pistachio rootstock which can keep K in their leaves over sodium can be considered as more tolerant under salt stress.
Proline Concentration of Leaves
The results indicated that the content of proline in the leaves of rootstocks was differently affected (P < .01) by salinity stress. However, the interaction of rootstock and salinity did not show a significant effect (). The 200 mM salinity level significantly increased the proline content (). The highest proline content was observed in the ‘Qazvini’ hybrid with 0.61 mg/g leaf fresh weight ().
Proline is an important osmolyte that maintains the cell osmoregulation under salt stress. Many studies have shown that proline accumulation is a common response of plants under stress (Sheikh-Mohamadi et al., Citation2017; Akbari et al., Citation2018a). The accumulation of proline due to water and salt stress results from proline synthesis in different tissues. It has been demonstrated that salinity stress prevents proline oxidation in mitochondria (Volkmar et al., Citation1998). Proline plays a key role in cellular regulation, such as carbohydrates and organic acids. More osmolyte accumulation during cellular stress is one of the plant's defense mechanisms against water loss or wilt (Akbari et al., Citation2020a). Research indicates the positive effect of Proline accumulation on osmotic regulation of the cell. Therefore, Proline can be introduced as an osmotic regulator in the cytoplasm section of plants (Levitt, 1980). Under the saline condition, typically higher levels of proline content in a plant species is a biological marker for better salinity tolerance (Akbari et al., 2018; Raufi et al., 2020). Hokmabadi et al. (Citation2005) reported that the highest amount of Proline accumulation was related to the ‘Qazvini’ rootstock among the three rootstocks of ‘Badami Zarand,’ ‘Sarakhs,’ and ‘Qazvini.’ So, our results are also confirmed by previous researches. The accumulation of proline under salt stress could be due to the four follow reasons: (1) its biosynthesis, (2) decreased rate of its oxidation to glutamate by P5C dehydrogenase and its dehydrogenase in mitochondria, (3) its decreased utilization because of protein synthesis, and (4) due to the upregulating stress-protective proteins (Akbari et al., Citation2018b).
A mechanism that plants adopted to reduce the negative effects of salt stress is increasing antioxidant enzymes activity to deactivate the free radicals (Grattan and Grieve, Citation1998; Greenway and Munns, Citation1980; Maas and Grattan, Citation1999). Higher activities of CAT, APX, SOD, and POD were observed in pistachio rootstocks forced by salt stress. Raufi et al. (2020) reported that the highest activities of CAT, APX, and SOD were recorded for UCB-1, it was not observed a significant difference between UCB-1 and Akbari rootstocks as well as between Akbari and Ahmad- ghaee rootstocks. They said the most activity of POD was observed in Akbari rootstock which was significantly higher than other cultivars. However, they revealed that there was a highly positive relationship between antioxidant enzyme action and proline, phenol, and Na concentrations in the root, shoot, and the leaf. In their study, Akbari and UCB-1 rootstocks with higher enzyme activity contained upper phenol and proline contents and lower Na content in compared with other rootstocks.
Conclusion
Strong vigor and rapid growth along with higher tolerance to salinity stress are desired traits for pistachio rootstocks breeders and growers. Our findings showed that although ‘Akbari’ showed higher salinity tolerance but significant differences were not observed between ‘Akbari’ and ‘Akbari’ hybrid rootstocks. Hence ‘Akbari’ hybrids could be appropriate candidates for the future researches and cultivation of pistachio in the saline areas.
References
- Adish, M., M. Fekri, and H. Hokmabadi. 2010. Response of Badami-Zarand pistachio rootstock to salinity stress. Int. J. Nuts Related Sci. 1(1):1–11.
- Akbari, M., H. Hokmabadi, M. Heydari, and A. Ghorbani. 2020a. Arota; a new interspecific hybrid pistacio rootstock. Hortscience 55(6):965–966. doi: https://doi.org/10.21273/HORTSCI14845-20.
- Akbari, M., M. Farajpour, M. Aalifar, and M. Sadat Hosseini. 2018a. Gamma irradiation affects the total phenol, anthocyanin and antioxidant properties in three different persian pistachio nuts. Natu. Prod. Res. 32(3):322–326. doi: https://doi.org/10.1080/14786419.2017.1346647.
- Akbari, M., N. Mahna, K. Ramesh, A. Bandehagh, and S. Mazzuca. 2018b. Ion homeostasis, osmoregulation, and physiological changes in the roots and leaves of pistachio rootstocks in response to salinity. Protoplas 255(5):1349–1362. doi: https://doi.org/10.1007/s00709-018-1235-z.
- Akbari, M., R. Katam, R. Husain, M. Farajpour, S. Mazzuca, and N. Mahna. 2020b. Sodium chloride induced stress responses of antioxidative activities in leaves and roots of pistachio rootstock. Biomolecules 10(2):189. doi: https://doi.org/10.3390/biom10020189.
- Aliakbarkhani, S.T., M. Akbari, A. Hassankhah, A. Talaie, and M.F. Moghadam. 2015. Phenotypic and genotypic variation in Iranian Pistachios. J. Genet. Engineer. Biotech. 13(2):235–241. doi: https://doi.org/10.1016/j.jgeb.2015.05.003.
- Aliakbarkhani, S.T., M. Farajpour, A.H. Asadian, M. Aalifar, S. Ahmadi, and M. Akbari. 2017. Variation of nutrients and antioxidant activity in seed and exocarp layer of some persian pistachio genotypes. Annal. Agric. Sci. 62(1):39–44. doi: https://doi.org/10.1016/j.aoas.2017.01.003.
- Balaguer, L., F.I. Pugnaire, E. Martínez-Ferri, C. Armas, F. Valladares, and E. Manrique. 2002. Ecophysiological significance of chlorophyll loss and reduced photochemical efficiency under extreme aridity in Stipa tenacissima L. Plant Soil 240(2):343–352. doi: https://doi.org/10.1023/A:1015745118689.
- Banakar, M.H., and G.H. Ranjbar. 2010. Evaluation of salt tolerance of pistachio cultivars at seedling stage. Amer.-Eur. J. Agric. Environ. Sci. 9:115–120.
- Barrs, H.D., and P.E. Weatherley. 1962. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 15(3):413–428. doi: https://doi.org/10.1071/BI9620413.
- Bastam, N., Baninasab, B and C, Ghobadi. 2013.Interactive effects of ascorbic acid and salinity stress on the growth and photosynthetic capacity of pistachio (Pistacia vera L.) seedlings. J. Hort. Sci and Biotech. 88(5): 610–616.
- Bates, L.S., R.P. Waldren, and I.D. Teare. 1973. Rapid determination of free proline for water-stress studies.Plant Soil 39(1):205–207. doi: https://doi.org/10.1007/BF00018060.
- Behboudian, M.H., R.R. Walker, and E. Törökfalvy. 1986. Effects of water stress and salinity on photosynthesis of pistachio. Sci. Hort. 29(3):251–261. doi: https://doi.org/10.1016/0304-4238(86)90068-3.
- Benmahioul, B., Daguin, and M. Kaid-Harche. 2009. Effet du stress salin sur la germination et la croissance in vitro du pistachier (Pistacia vera L.). Compt. Rendus Biol. 332(8):752–758. doi: https://doi.org/10.1016/j.crvi.2009.03.008.
- Del Carmen Gijón, M., C. Gimenez, D. Perez-López, J. Guerrero, J.F. Couceiro, and A. Moriana. 2010. Rootstock influences the response of pistachio (Pistacia vera L. cv. Kerman) to water stress and rehydration. Sci. Hort. 125(4):666–671. doi: https://doi.org/10.1016/j.scienta.2010.05.026.
- Dubey, R.S. 1997. Photosynthesis in plants under stressful conditions, p. 859–875. In: M. Pessarakli (ed.). Handbook of photosynthesis. Marcel Dekker publ, NewYork.
- Ferguson, L., and D. Haviland. 2016. Pistachio production manual (3545). University of California Agriculture and Natural Resources Press: UCANR Publications.
- Ferguson, L., H. Reyes, B. Sanden, S. Grattan, L. Epstein, and B. Krueger. 2005. Pistachio rootstocks. Pistach. Product. Manual. 125 (4):67–73.
- Ferguson, L., J.A. Poss, S.R. Grattan, C.M. Grieve, D. Wang, C. Wilson, T.J. Donovan, and C.T. Chao. 2002. Pistachio rootstocks influence scion growth and ion relations under salinity and boron stress. J. Ame. Soci. Hort. Sci. 127(2):194–199. doi: https://doi.org/10.21273/JASHS.127.2.194.
- Goharrizi, K.J., A. Baghizadeh, M. Kalantar, and F. Fateh. 2020. Combined effects of salinity and drought on physiological and biochemical characteristics of pistachio rootstocks. Sci. Hort. 261:108970. doi: https://doi.org/10.1016/j.scienta.2019.108970.
- Grattan, S.R., and C.M. Grieve. 1998. Salinity–mineral nutrient relations in horticultural crops. Sci. Hort. 78(1):127–157. doi: https://doi.org/10.1016/S0304-4238(98)00192-7.
- Greenway, H., and R. Munns. 1980. Mechanisms of salt tolerance in nonhalophytes. Annual Rev. Plant Physiol. 31(1):149–190. doi: https://doi.org/10.1146/annurev.pp.31.060180.001053.
- Gunver-Dalkilic, G., and O. Dayl-Dogru. 2011. Determination of pollen grain viability and germination levels for pistachio and terebinth in Aydin/Turkey Ecology. Pak. J. Bot. 43(2):841–848.
- Hokmabadi, H., K. Arzani, and P.F. Grierson. 2005. Growth, chemical composition, and carbon isotope discrimination of pistachio (Pistacia vera L.) rootstock seedlings in response to salinity. Crop. Past. Sci 56(2):135–144. doi: https://doi.org/10.1071/AR04088.
- Jamshidi Goharrizi, K., A. Baghizadeh, M. Kalantar, and F. Fattahi. 2019. Assessment of changes in some biochemical traits and proteomic profile of UCB-1 pistachio rootstock leaf under salinity stress. J. Plant. Growth. Regul. 39:608–630. doi: https://doi.org/10.1007/s00344-019-10004-3.
- Karimi, H.R., A.M. Kuhbanani, H. Sajadian, H. Hokmabadi, F. Kamiab, M.H. Salehabad, and E. Zamanibahramabadi. 2015. The evaluation of inter specific hybrid of P. atlantica× P. vera cv. ‘Badami Zarand’as a pistachio rootstock to salinity stress. J. Nut. 6(2):113–122.
- Karimi, S., M. Rahemi, M. Maftoun, and V. Tavallali. 2009. Effects of long-term salinity on growth and performance of two pistachio (Pistacia vera L.) rootstocks. Aust. J. Basic App. Sci. 3(3):1630–1639.
- Khoyerdi, F.F., M.H. Shamshiri, and A. Estaji. 2016. Changes in some physiological and osmotic parameters of several pistachio genotypes under drought stress. Sci. Hort. 198:44–51. doi: https://doi.org/10.1016/j.scienta.2015.11.028.
- Lutts, S., J.M. Kinet, and J. Bouharmont. 1996. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Anua. Bot. 78(3):389–398. doi: https://doi.org/10.1006/anbo.1996.0134.
- Maas, E.V., and S.R. Grattan. 1999. Crop yields as affected by salinity. Agronomy 38:55–110.
- Mesgaran, M.B., K. Madani, H. Hashemi, and P. Azadi. 2017. Iran’s land suitability for agriculture. Sci. Rep. 7(1):1–12. doi: https://doi.org/10.1038/s41598-017-08066-y.
- Mohamad Khani, A. 1997. Appointment of relative resistance in salt condition (NaCl) in pistachio (Alteration of stomata respiration, absorption and element transition), M. Sc. Thesis. Faculty of Agriculture. Tehran University. Karaj. Iran.
- Panahi, B. 2009. Effects of osmotic and salt stresses on water relation parameters of pistachio seedlings. J. Plant Ecophysiol. 1(1):1–8.
- Picchioni, G.A., S. Miyamoto, and J.B. Storey. 1990. Salt effects on growth and ion uptake of pistachio rootstock seedlings. J. Ame. Soci. Hort. Sci. 115(4):647–653. doi: https://doi.org/10.21273/JASHS.115.4.647.
- Rahneshan, Z., F. Nasibi, and A.A. Moghadam. 2018. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant. Interact. 13(1):73–82. doi: https://doi.org/10.1080/17429145.2018.1424355.
- Raoufi, A., M. Rahemi, H. Salehi, and A. Javanshah. 2019. Selecting high-performance rootstocks for pistachio cultivars under salinity stress based on their morpho-physiological characteristics. Inter. J. Fruit Sci. 4(1):1–19.
- Raoufi, A., M. Rahemi, H. Salehi, and M. Pessarakli. 2020b. Pistacia vera L. genotypes; a potential rival for UCB-1 rootstock for cultivating under salt stress conditions. Biocat. Agri. Biotec. 25(3):1–9.
- Raoufi, A., M. Rahemi, and M. Akbar. 2020a. Glycerol foliar application improves salt tolerance in three pistachio rootstocks. J. Saudi Soci. Agri. Sci. 6(19):426–437.
- Sepaskhah, A.R., and M. Maftoun. 1988. Relative salt tolerance of pistachio cultivars. J. Hort. Sci. 63(1):157–162. doi: https://doi.org/10.1080/14620316.1988.11515841.
- Sheikhi A., Arab M.M., Brown P.J., Ferguson L., Akbari M. 2019. Pistachio (Pistacia spp.) Breeding. In: Al-Khayri J., Jain S., Johnson D. (eds.), Advances in Plant Breeding Strategies: Nut and Beverage Crops. Springer, Cham. https://doi.org/http://doi.org/10.1007/978-3-030-23112-5_10
- Sheikh-Mohamadi, M.H., N. Etemadi, A. Nikbakht, M. Arab, M.M. Majidi, and M. Pessarakli. 2017. Antioxidant defense system and physiological responses of Iranian crested wheatgrass (Agropyron cristatum L.) to drought and salinity stress. Acta Physiol. Planta. 39(11):245–253. doi: https://doi.org/10.1007/s11738-017-2543-1.
- Taghizadeh-Alisaraei, A., H.A. Assar, B. Ghobadian, and A. Moteval. 2017. Potential of biofuel production from pistachio waste in Iran. Ren. Sust. Ene. Rev. 72:510–522. doi: https://doi.org/10.1016/j.rser.2017.01.111.
- Talebi, M., M. Akbari, M. Zamani, and B.E. Sayed-Tabatabaei. 2016. Molecular polymorphism in Pistacia vera L. using non-coding regions of chloroplast DNA. J. Genet. Engin. Biotec. 14(1):31–37. doi: https://doi.org/10.1016/j.jgeb.2016.04.002.
- Tavallali, V., M. Rahemi, and B. Panahi. 2008. Calcium induces salinity tolerance in pistachio rootstocks. Fruits 63(5):285–296. doi: https://doi.org/10.1051/fruits:2008024.
- Volkmar, K.M., Y. Hu, and H.H. Steppuhn. 1998. Physiological responses of plants to salinity: A review. Can. J. Plant Sci. 78(1):19–27. doi: https://doi.org/10.4141/P97-020.
- Walker, R.R., E. Torokfalvy, and M.H. Behboudian. 1988. Photosynthetic rates and solute partitioning in relation to growth of salt-treated pistachio plants. Func. Plant Biol. 15(6):787–798. doi: https://doi.org/10.1071/PP9880787.
- Warschefsky, E.J., Laura, L. K., Margaret, H. F., Daniel H. C., Jason, P. L., Eric J.B and J.M, Allison. 2016. Rootstocks: Diversity, Domestication, and Impacts on Shoot Phenotypes. Trend. Plant. Sci. 21(5): 418–437.