
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Covering fig trees with color nets changed the quality of light that reach trees. This study was carried out to assess the effects of shade provided by blue and yellow protection nets on leaf attributes and fruit quality of fig trees under rain-fed conditions. The experiment lasted for three months and was a split-plot treatment arrangement in a randomized complete block design with three replications. ‘Sabz’ and ‘Siah’ fig trees were covered with color nets or left uncovered (as the control group). The results showed that yellow nets reduced the net photosynthetic activity by 15.86 µmol CO2 m−2 s−1, but increased leaf chlorophyll and carotenoid content in both cultivars. Leaf temperature decreased in both cultivars when covered with yellow nets. The shade provided by blue nets increased stomatal conductance in the leaves of ‘Sabz’ fig trees. When covered with yellow shade nets, ‘Sabz’ fig trees had the highest relative water content and the lowest ion leakage in both cultivars. The color nets reduced dry fruit diameter in ‘Sabz’ figs and reduced total soluble solids in both cultivars. Also, titratable acidity decreased in fruits as a result of using yellow nets on both cultivars. Yellow nets increased antioxidant activity and anthocyanin content in fruits of the ‘Siah’ cultivar significantly, as compared with the ‘Sabz’ cultivar. The photo-selective, light-dispersive shade nets proved to be interesting tools that can reduce drought stress on trees under rain-fed conditions.
KEYWORDS:
Introduction
Photo-selective netting can be considered as a new agro-technological approach which has been steadily implemented worldwide. Its use has been evaluated in different varieties of crops, climatic regions and cultivation methods (Shahak et al., Citation2008). The color nets provide physical protection and cause partial resistance against climate change impacts (e.g. high temperature, extreme weather events, lack of irrigation water, etc.), together with differential filtration of the solar radiation where particular physiological responses are expected through the regulation of light intensity and quality (Abul-Soud et al., Citation2014). Photo-selective netting can also alter the growth and developmental cycle of fruits, including fruit-set, harvest time, fruit yield, size and color, as well as internal and external qualities of the fruits, whereby physiological disorders are relatively avoided (Racsko and Schrader, Citation2012; Rajapakse and Shahak, Citation2007; Shahak et al., Citation2008, Citation2004). According to the available literature, protective nets can markedly affect gas exchange through the leaves of trees (Szymańska et al., Citation2017). They can cause enhanced levels of relative humidity (Hunsche et al., Citation2010; Kalcsits et al., Citation2017), water use efficiency (Hunsche et al., Citation2010; McCaskill et al., Citation2016), vegetative growth and leaf morphology (Folta and Carvalho, Citation2015; Solomakhin and Blanke, Citation2008). Using color nets can lead to pronounced benefits in fruit set, flower induction and blooming (Do Amarante et al., Citation2011; Zibordi et al., Citation2009).
So far, extensive experiments have tested the effects of photo-selective nets on agricultural plants including ornamentals, vegetables and fruits crops. Different colors of such nets modify desirable plant growth characteristics. For instance, they stimulate the vegetative growth rate, vigor of foliage and cut flower crops, especially when red and yellow nets are used. Whereas blue nets have reportedly caused dwarfing, gray nets have increased branching and bushiness but reduced leaf area (Alkalia-Tuvia et al., Citation2014; CitationGoren et al., ; CitationKong et al., ; Shahak et al., Citation2008). Regarding ‘Pinova apple’, using color nets 12% white, 14% red-white, 18% red-black and 23% green-black nets led to the increase in several vegetative growth indices, such as the average count of one-year-old shoots per tree, mean length of one-year-old shoots and total length of one-year-old shoots, as compared to an uncovered control group (Solomakhin and Blanke, Citation2008) . In grape production, cover materials are used in order to encourage early- or late-harvesting, to protect against rain, hail, snow and storms, and to avoid the negative effects of pests and diseases (Sen et al., Citation2016).
The fig (Ficus carica L.) belongs to the Moraceae family and is one of the first plants to have been cultivated and consumed by human beings (Dueñas et al., Citation2008). Figs are harvested from a total of 365,350 hectares of cultivated land around the world, with an annual production of over one million tons (Fao and UNICEF, Citation2017) . Iran is the third largest producer of dried figs, as well as the fifth largest fresh fig producer in the world with a cultivated area of 53,101 hectares and a production of 70,730 tons per year (Fao and UNICEF, Citation2017). The majority of fig production in Iran is cultivated in Estahban, Fars, under rain-fed conditions. The cultivars ‘Sabz’ and ‘Siah’ are the most desirable and economic cultivars in Iran and they are consumed as dry and fresh fruits, respectively. Since in this region the temperature increases up to 40°C in mid-August and water consumption enhances during fruit ripening, at a time when fig trees are not irrigated, damage to trees and fruits including drought stress, red bees and birds’ damages is inevitable under rain-fed conditions. Meanwhile, using colored nets seems to be a good way to alleviate these problems. So far, no research has been conducted on the effects of color nets on physiochemical changes in fig trees and fruits under rain-fed conditions. Thus, the aim of this study was to investigate the influence of color net and the impacts of light manipulation by photo-selective nets on physiological, biochemical and qualitative properties of two main fig cultivars. The treatments were expected to reduce solar radiation on trees, to maintain water content and fruit quality in the two commercial fig cultivars by the time the harvest season arrives.
Materials and Methods
A trial was conducted in Fig Research Station, Estahban, Iran (29°08´ N, 54°02´ E, 1760 m altitude). Cultivars ‘Sabz’ and ‘Siah’ (50-year-old plants) were selected at 10 m × 10 m spacing on a sandy loam soil. The trees were covered with photo-selective shading nets provided by Exsirsaz e Shomal Co. (Iran). Two colors (blue and yellow) and a control (full sun) were used. Shading percentage of experimental treatments are shown in . The nets were installed horizontally at 3.5 m height on each tree on August 7, 2019, and remained on the trees for three months during fruit ripening (i.e. before commercial maturity of the fruits). They were removed after the last fruit harvesting (November 7). The experiment was a randomized complete block design with three replications (blocks) and a split plot treatment arrangement. Main plots were two cultivars and subplots were three net colors. Each block consisted of two adjacent rows with three trees each (). At harvest time (mid-September), healthy and fully developed leaves were collected from a particular part of each tree, i.e. adjacent to the fruit of the third node, the southern part of the canopy surface. The fruits were selected from the middle nodes after they had reached commercial maturity. Some of the leaves were picked and immediately used for leaf analysis, while some of the sampling leaves and fruits were sorted, cleaned and packed in bags of polyethylene film and stored at 4°C for future physical and chemical analyses.
Table 1. The influence of color nets on illumination and shading level
Figure 1. Layout of experiment: (C1 = Sabz cultivar, C2 = Siah cultivar, N1 = control, N2 = blue net, N3 = yellow net)

Measurements and Observations of Leaf Sample
Leaf Dimensions
Width and length (cm) of leaf were measured in four leaves that were similar to other tree leaves and had been collected from different sides of each tree for each replication. The results were described as a mean value of 12 leaves.
Leaf Temperature
Leaf temperature (°C) was recorded in four sun-exposed leaves that had been collected from different sides of each tree for each replication by Infra-Red thermometer (Testo 830- T2, Germany). The measurement was taken between 13:00 and 14:00 PM that the air temperature, wind speed, humidity, and light intensity for uncovered trees at time of measurement were 33°C, 0, 12%, and 830 W.m−2, respectively.
Light Intensity
Light intensity was measured at a distance of 20 cm, bellow the color nets and around the tree canopy using a solar power meter (TES, 1333) from 13:00 to 14:00 PM. Light intensity was expressed as Watts per square meter (Wm−2).
Net Photosynthesis Measurement
Net CO2 assimilation (µmol CO2 m−2 s−1) of a fully developed leaf, exposed to sunlight, was evaluated. Net photosynthesis of two leaves on the south side of each tree was measured three times. One leaf of each tree for each replication was placed inside a device clamp and the measurements were carried out from 13:00 to 14:00 PM. Photosynthetic parameters were measured using an open gas exchange system with a 4 cm2 leaf cuvette (LCi, ADC, UK). The results were described as a mean value of three times of measurements.
Stomatal Conductance
Stomatal conductance (mol H2O) m−2 s−1) was measured in six leaves for each replication using a leaf porometer (LCi, ADC, UK) from 13 to 14 PM.
Electrolyte Leakage
Ionic leakage was measured as described by Bartolozzi and Fontanazza (Citation1999). Five discs with 10 mm diameter were collected from leaves of each treatment and placed in a test tube containing 25 ml distilled water and shacked for 4 h. Electrolytic conductivity (EC1) was measured using digital conductivity meter (Model HI8633, USA). Solutions and samples were then autoclaved to kill the cells. Once the solution was cooled, conductance was again measured (EC2). Ionic leakage (EC %) was calculated based on the equation.
EC% = EC1/EC2 × 100
Relative Water Content (RWC)
Five fully developed young leaves were selected for RWC determination immediately after removing the nets. Fifteen uniform disks (diameter 10 mm) were separated from sample leaf that there was no vein. After measuring the fresh weight of the discs, using a digital balance (A&D EJ-303, Korea), the disks were put into double deionized water for 24 h under dim light to get turgid weight (TW). Then the leaf was dried in an oven at 70◦C for 48 h to get dry weight (DW). RWC (%) was calculated by the following equation:
Leaf Water Content (LW)
Leaf water content (%) in two leaves of each tree for each replication was calculated with following equation.
Where FW is fresh weight of leaf and DW is dry weight of the leaf after drying in the oven at 70°C for 72 h.
Chlorophyll and Carotenoid Determination
Chlorophyll a and b were measured, along with total chlorophyll and carotenoid contents. One leaf was considered for each tree per replication, as determined according to a method described by Arnon (Citation1949). DMSO was used as the extraction solvent for pigments (Hiscox and Israelstam, Citation1979). The absorbance of the extractions was recorded at 663, 645 and 470 nm by a spectrophotometer (Epoch, Bio Tek©, United States). Chlorophyll and carotenoid concentrations (mg.g−1FW) were calculated by Arnon equations (Citation1949):
Chl. a = (19.3 × A663 – 0.86A645) V/100 W
Chl. b = (19.3 × A645 – 3.6A663) V/100 W
Chl. T = Chl. a+ Chl. b
Car = (1000A470-1.8 Chl.a-85.2 Chl.b)/198
Where Chl.a is chlorophyll a, Chl. b is chlorophyll b, Chl.T is total chlorophyll, Car. is carotenoid, V is the volume made and W is weight of the sample.
Measurements of Fruit Samples
Fruit Diameter
Fruit diameter of 6 fruits in each replication was measured by a digital caliper and the results were expressed as millimeter.
Fresh and Dry Weight of Fruits
The fresh weight (FW) of six fruits were measured using a digital balance and to determine the fruit dry weight, they were placed in envelopes and dried in an oven at 70°C for 72 h and then the dry weight (DW) was measured using a digital balance with 0.01 g accuracy.
Total Soluble Solids (TSS)
Total soluble solid contents of six fruits were determined using a hand held refractometer and the results were expressed as °Brix.
Titratable Acidity (TA)
The titratable acidity was expressed as % citric acid after titration with 0.1 N alkaline solution (0.1Msodium hydroxide) up to an end point of pH 8.1. TA(%) was calculated by the following equation (Ranganna, Citation1986).
Antioxidant Activity
Antioxidant activity of six fig fruits were measured, as described by Ao et al. (Citation2008). The initial absorbance of α,α-diphenyl-1-picryl-hydrazyl (DPPH), through a free radical scavenging assay, was read at 517 nm using a spectrophotometer (HITACHI, U − 1800, Tokyo, Japan) until the absorbance reached a constant value. A total of 10 ml of fruit extract was added to 4 ml of distilled water and 1 ml of 250 mM methanolic DPPH solution. The mixture was incubated at room temperature for 30 min, before a change in absorbance at 517 nm was observed. Percentage reduction of the DPPH was defined by: DPPH (%) = 100 (A0− Ac)/A0, where A0 is the control absorbance without sample concentration and Ac is the value for added sample concentration.
Determination of Anthocyanin
Total anthocyanin contents of six fruits were determined according to the pH by the differential method (Kim et al., Citation2003). Absorbance was measured at 520 and 700 nm and expressed as mg cyanidin-3-glycoside equivalents per 100 g FW of fruit.
Statistical Analysis
Data were analyzed as plot means using SAS software version 9.00 for all analyses. Data analysis was conducted using analysis of parametric variance (ANOVA) procedure. The comparison of mean values was carried out according to the Fisher’s Least Significant Difference (LSD) (P ≤ 0.05).
Results
Leaf Dimensions
Fig trees that received full sun developed enhanced leaf dimensions in both cultivars (). In using the yellow net, the ‘Siah’ cultivar developed the smallest leaf width under rain-fed conditions. The shortest leaf length was observed in Siah cultivar when covered with yellow nets, although the difference with other covered treatments was not significant ().
Table 2. The main influence of net colors on some fig attributes
Table 3. The combined effects of color nets and fig cultivars on some leaf physiological properties
Leaf Temperature
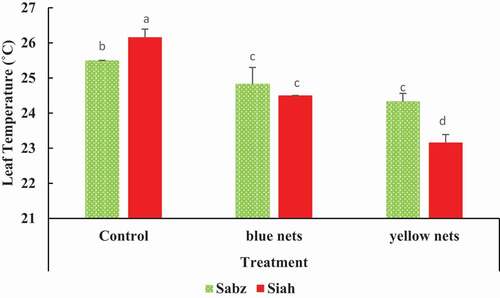
Colored nets changed leaf temperature in both cultivars. As shown in , Siah cultivars that received full sun ultimately obtained the highest leaf temperature. But yellow nets reduced the leaf temperature to almost 23°C in the Siah cultivar (). In general, fig trees that were covered with blue and yellow nets had lower leaf temperature ().
Figure 2. The combined effects of nets and fig cultivars on leaf temperature in two fig cultivars. Means (n = 3) followed by different letters are significantly different at p < .05 (LSD test)
Net Photosynthesis
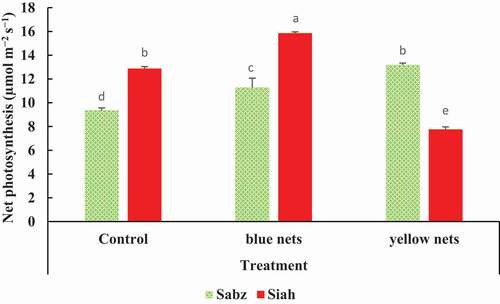
As shown in , CO2 assimilation rate was significantly affected by color nets in both cultivars. The highest net photosynthesis was recorded in Siah cultivar covered with blue nets (15.86 µmol m−2 s−1). Yellow net caused the lowest value of net photosynthesis in Siah cultivar (7.76 µmol m−2 s−1).
Figure 3. The combined effects of nets and fig cultivars on leaf net photosynthesis (CO2 assimilation rate). Means (n = 3) followed by different letters are significantly different at p < .05 (LSD test)
Stomatal Conductance
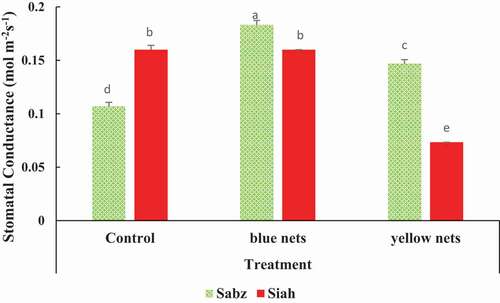
Colored nets influenced the stomatal conductance in both cultivars. The Sabz cultivar with a blue net had an enhanced stomatal conductance of 0.18 mol m−2 s−1 (), whereas yellow nets reduced the stomatal conductance to 0.07 mol m−2 s−1 in Siah cultivar ().
Figure 4. The combined effects of nets and fig cultivars on leaf stomatal conductance. Means (n = 3) followed by different letters are significantly different at p < .05 (LSD test)
Relative Water Content (RWC), Electrolyte Leakage and Leaf Water Content
Covering trees with blue and yellow nets affected the relative water content (). Yellow nets caused the highest RWC in Sabz cultivar, whereas the lowest percentage (68.16%) was observed in Siah fig trees which received full sun ().
A great amount of electrolyte leakage was observed when using blue nets and in uncovered fig trees in both cultivars. With yellow nets, the Sabz and Siah cultivars showed the lowest value of electrolyte leakage under rain-fed conditions, but the electrolyte leakage in the Sabz cultivar was lower than in the Siah ().
Blue nets caused the highest leaf water content in the Sabz cultivar, as compared with other treatments (62.39%). ‘Sabz’ fig trees which received full sun had the lowest leaf water content (56.91%) (). As shown in , the direct effect of color nets on leaf water content was recorded in fig trees covered with blue nets (61.47%).
Chlorophyll a and B, along with Total Chlorophyll and Carotenoid Contents
Fig trees covered with yellow nets had enhanced levels of chlorophyll content (). As shown in , the Sabz cultivar being covered with yellow nets had enhanced levels of chlorophyll a content (15.36 mg/g FW). Uncovered Siah fig trees showed the lowest chlorophyll a content (6.60 mg/g FW). The greatest content of chlorophyll b was recorded in Sabz cultivar covered with yellow nets, whereas Siah fig trees under the full sun had the lowest content of chlorophyll b. Total chlorophyll increased in the ‘Sabz’ cultivar when covered with yellow nets (16.93 mg/g FW) but decreased in the Siah cultivar which received full sun (7.37 mg/g FW) ().
Table 4. The combined effects of color nets and fig cultivars on leaf chlorophyll and carotenoid (mg g−1FW)
In the Sabz cultivar, leaf carotenoid content was higher in trees covered with yellow nets and in those which received full sun (7.38, 7.03 mg/g FW, respectively). The lowest carotenoid content was observed in Siah trees with no covering (3.42 mg/g FW) ().
Fresh and Dry Weight of Fruits
Under full sun, fig trees enhanced the fresh weight of fruits in both cultivars (), whereas covering fig trees with blue nets reduced the fresh weight of fruits in both cultivars ().
Table 5. The combined effects of color nets and fig cultivars on some fruit physical attributes
The direct effect of color nets on dry weight of fig fruits revealed that uncovered fig trees grew fruits of heavier dry weight (). The heaviest fruit dry weight was observed in uncovered Sabz cultivar figs (5.93 g) (). Fruit dry weight significantly decreased (3.68 g) in the Sabz cultivar when covered with blue nets ().
Fruit Diameter
Uncovered fig trees in both cultivars developed the highest dry fruit diameter. The Siah cultivar had a greater fruit diameter than the Sabz cultivar under rain-fed conditions (). Blue and yellow nets caused the smallest dry fruit diameter in the Sabz cultivar (22.09, 21.55 mm, respectively).
Fresh fruit diameter increased in fig trees under full sun in both cultivars (), whereas blue nets in the Sabz cultivar caused the smallest diameter of fresh fruits (30.72 mm) ().
Total Soluble Solids (TSS)
According to , the direct effect of color nets on TSS was recorded in fig trees under full sun and in those with yellow and blue nets. Covering the trees with blue and yellow nets reduced TSS in Sabz cultivar (14.6, 13.6 °Brix, respectively) compared with other treatments. The highest TSS was recorded in uncovered Siah cultivar, while having no significant difference with some treatments ().
Table 6. The combined effects of color nets and fig cultivars on some fruit biochemical attributes
Titratable Acidity (TA)
Covering the trees with yellow nets reduced titratable acidity in Sabz and Siah cultivars (0.21% and 0.20%, respectively). ‘Sabz’ and ‘Siah’ fig trees with blue nets had higher titratable acidity (0.24%) ().
Antioxidant Activity and Anthocyanin Content
Antioxidant activity and anthocyanin content was higher in the Siah cultivar compared with Sabz.
Yellow nets and the control group of the Siah fig cultivar caused the highest antioxidant activity (5.57% and 5.43%, respectively). The lowest activity was recorded in Sabz cultivar covered with blue nets, having no significant difference with the other two shade nets in this cultivar ().
The highest anthocyanin content was recorded in Siah cultivar when covered with yellow nets (6.24 mg 100 g FW−1), whereas the lowest content was observed in Sabz cultivar covered with blue nets. Nonetheless, there was no significant difference between the yellow and uncovered treatments in this cultivar ().
Discussion
The influence of net-covering by itself was recorded to alleviate extreme climatic variation, reduce abiotic stress, increase photosynthesis and canopy development and prevent fruit sunburns, as compared to the non-covering practice (Shahak et al., Citation2008, Citation2004; Tanny, Citation2013) . Also, the photo-selective, light-dispersive filtration of sunlight influenced physiological attributes in a wide variety of ways, depending on the chromatic properties of each net.
Shade color nets are less likely to influence vegetative growth such as leaf dimensions in fig trees, which is in line with a previous study on kiwifruit vines with blue nets that showed significantly lower vegetative vigor (Mupambi et al., Citation2018). In contrast, in apple and peach trees, the vegetative growth increased under blue nets (Baraldi et al., Citation1994; Bastías et al., Citation2012). In the current study, since the fig trees were under rain-fed conditions and also, they covered with shading nets, it seemed that the leaf dimensions decreased. The results of this experiment were consistent with the results of the effect of shading on total leaf area in grapevine (Cartechini and Palliotti, Citation1995). It was reported that in pepper, increase in leaf size by increasing shading level, may be due to a decrease in the number of lateral branches in shady conditions, which was not in line with our results (Rylski and Spigelman, Citation1986). Interestingly, photo-selective nets that alter light quality can affect leaf morphology and vegetative growth. Therefore, color nets serve as a useful management technique in the orchard to manage vegetative growth and provide optimum light interception for high-quality fruit production.
The current study revealed that fig trees under full sun receive the highest light intensity. Similar results were found by Stamps (Citation2009) who showed that photo-selective nettings can reduce the amount of radiation reaching the crops beneath. Notably, blocked radiation (%) depends on the degree of shade (Stamps, Citation2009). Also, similar to our results, the analysis of light intensity in tomato fruits showed that solar radiation passes less through color nets in comparison with radiation in the open field (Ilić et al., Citation2012). Additionally, Ilić et al., (Citation2017) reported that color shade nets can reduce light intensity, as compared with the natural environment during the summer months (Ilić et al., Citation2017).
In this study, different net colors reduced leaf temperature under rain-fed conditions. This reduction in temperature was associated with radiation, intercepted by the nets (Iglesias and Alegre, Citation2006). In accordance with our results, a range of 1–3°C reduction in temperature occurred in response to using shade nets, as reported by (Iglesias and Alegre (Citation2006); Solomakhin and Blanke (Citation2010)). The various responses founded in previous research suggest a need to determine whether the temperature effect of protective netting in an orchard can be influenced by shading factor, net porosity and growing environment (Mupambi et al., Citation2018).
In the current study, yellow nets reduced photosynthetic rates and caused lower stomatal conductance in fig trees, which was in agreement with a previous study by Zhou et al. (Citation2018) on orange trees (Zhou et al., Citation2018). These results emanate from the enriched green light that is transmitted through yellow nets, as compared to a blue net that bars wavelengths in this region. Green light is associated with reduced leaf size, lower chloroplast accumulation and reduced stomatal conductance (Folta and Carvalho, Citation2015).
In the present study, fig leaves by yellow nets contained more chlorophyll than leaves exposed to direct sunlight. At high light intensities, the degradation rate of chlorophyll in plant leaves is higher than the synthesis rate. Thus, the concentration of chlorophyll decreases as less chloroplasts are formed (Fu et al., Citation2012; Kosma et al., Citation2013). Also, it seemed that leaves under shading nets have higher chlorophyll contents and are generally thinner than those exposed to full sun, thereby expanding light harvesting efficiency for photosynthesis (Taiz and Zeiger, Citation2012). In line with our study, changes in leaf chlorophyll content were found by the effect of spectral light composition under colored nets in citrus trees: plants under blue nets acquired the greatest amount of leaf chlorophyll a, b, and total chlorophyll content, whereas leaves under red nets had the lowest of these values (Li and Syvertsen, Citation2006). Shaded lettuce leaves generally have larger total chlorophyll content, which is in agreement with our study (Ilić et al., Citation2017).
Carotenoids act as a selective filter in plants that protect chlorophylls from too much light or from inappropriate wave lengths (Díaz-Pérez, Citation2013). Excited chlorophyll molecules can result in singlet oxygen formation. Thus, carotenoids are able to absorb energy to prevent this formation. Furthermore, carotenoids can scavenge any singlet oxygen that forms during photosynthesis and excess radiation (Bergquist, Citation2006). If they are not scavenged, photo inhibition in plants may occur (Bergquist et al., Citation2007).
Because of reduced solar radiation through the shading nets, the use of irrigation water in orchards is reduced. This can be accompanied by a greater relative water content, less electrolyte leakage and more leaf water content in fig trees under netting, leading to lower levels of drought stress under rain-fed conditions. Our results were in line with the findings of Mihailova et al. (Citation2018), who reported that electrolyte leakage decreases in shady plants compared to sun plants. In apple, potential evapotranspiration was reduced by 30% under protective netting and with less solar radiation reaching the orchard floor. Together with a reduction in wind speed, water loss from the soil and evapotranspiration were reduced and a desirable water content in apple trees under netting was achieved (Gindaba and Wand, Citation2007; McCaskill et al., Citation2016). Nicolás et al. (Citation2008) found that shaded lemon trees have lower transpiration and higher water use efficiency when compared with unshaded trees, resulting from the altered microclimate under the protective net. In this study, shading nets reduced total soluble solids of fig fruits in both cultivars under rain-fed conditions, which was in line with previous reports (Do Amarante et al., Citation2011; Dussi et al., Citation2005; Iglesias and Alegre, Citation2006). A lower level of total soluble solids was observed in fig trees covered with nets. This may be a result of the lower photosynthetic activity of plants and, consequently a lower carbohydrate accumulation in fruits. A previous report claimed that light intensity and temperature have significant effects on soluble solids accumulation in tomatoes. Therefore, exposing fruit to higher temperatures, especially during fruit cell division and ripening, can result in an increase of total soluble solids (Ilić et al., Citation2015). Furthermore, an increase in carbohydrate biosynthetic enzyme activity and enhanced levels of transpiration were observed parallel to the increase in total soluble solids (Gindaba and Wand, Citation2007). As the leaves are exposed better to light in different wavelengths, higher amounts of total soluble solids are produced in the fruit (Do Amarante et al., Citation2011; Ilić et al.). Photo-selective nets had significant effects on fruit firmness in rain-fed fig fruits (Jokar et al., Citation2021) () and tomato fruits (Ilić et al., Citation2015). It has been reported that growing tomato plants under color nets and in optimal conditions can make firmer fruits with a thicker pericarp and a better tolerance to transport (Ilić et al., Citation2015). According to Campbell et al. (Citation1992), a softer flesh was observed in apples under shading conditions. They suggested that a decrease in flesh firmness might be the result of poor activity in cell wall formation and a high water influx to the cortex cells of the fruit. The present study revealed that titratable acidity (TA) was affected by different colors of shading nets in fig trees. In line with our results, Fernández-Trujillo et al. (Citation2009) suggested that the increase in TA content of pepper fruits under color nets could be associated with fruit senescence. Covering the trees with white nets did not reduce titratable acidity (TA), as this was assessed at harvest, after cold storage and during shelf life maintenance of two apple cultivars. Nonetheless, several authors did not report a decrease in fruit TA by covering fruit orchards with photo-selective nets (Iglesias and Alegre, Citation2006; Wagenmakers and Tazelaar, Citation1999). A reduced TA content in‘Pinova’ and ‘Fuji’ apple fruits was reported when trees were protected by nets (Solomakhin and Blanke, Citation2010). It must be highlighted that besides the differences between cultivars, several authors attributed a stronger role to orchard management, crop load and climate in defining the internal fig fruit quality, as compared to the effect of different net colors. The accumulation of antioxidant compounds in fruit trees depends on many factors such as temperature, light quantity and quality, cultivars, growing season and metabolic factors. Elevated temperatures and a decrease in plant growth can enhance the antioxidant scavenging activity in countering reactive oxygen species (Miller et al., Citation2010). Blue light especially increased the number of phenolic compounds, e.g. anthocyanins (Johkan et al., Citation2010). As a higher anthocyanin content was observed in the Siah cultivar, a higher antioxidant activity was expected. Thus, more yellow light passes through and the transmittance of blue light through yellow nets (500–600 nm) increased the production of antioxidants and the level of antioxidant scavenging activity in Siah fig trees. In a previous report, oregano leaves had a significantly higher content of total antioxidant activity when grown under yellow nets, which was in accordance with our results (CitationIlić et al.,). In this study, as the skin color of Siah fruits is darker than Sabz fruits, the former is characterized by a higher anthocyanin content, whereas carotenoids are the main pigments in Sabz fruits. Anthocyanin pigments accumulate under light stress conditions and they are more stable under these conditions in comparison with chlorophyll. A higher anthocyanin accumulation was observed in fruits grown under yellow nets which could be the result of a relative enrichment/reduction of blue and yellow spectral bands of transmitted light. Moreover, anthocyanin serves as a defensive pigment and, thus, anthocyanins maintain apple peel against light-induced stress and protect against damage to fruit skin by trapping light in the chlorophyll absorption gap (Mupambi et al., Citation2018). Since environmental conditions changed under protective shade netting, they could potentially affect yield, fruit size and weight. In fig trees, light limitation, shading and protective netting can potentially reduce fruit size as a result of reduced tree photosynthesis. Nevertheless, it was suggested that in regions where trees frequently experience abiotic stress due to extreme solar radiation, netting can have a positive impact on fruit size by alleviating side effects and stress, thereby sustaining higher photosynthetic rates later on in the day, as compared with trees under full sun for which the photo-inhibition phenomenon is inevitable (Mupambi et al., Citation2018). Under color shading nets, fruit weight decreased in both fig cultivars, which was not in agreement with a previous report about the increase in highbush blueberry weight when plants were under color nets (Lobos et al., Citation2013).
Conclusion
We demonstrated how net colors modify the spectrum of light that plants receive. These modifications can influence plant growth performance under rain-fed conditions. Our observations confirmed the significant effect of protective nets on tree physiological performance and on fruit quality by causing enhanced photosynthetic rates, stomatal conductance, relative water content, vegetative growth, antioxidant activity and anthocyanin content in both cultivars under rain-fed conditions. In conclusion, the effects of the blue and yellow nets are manifestations of their relative enrichment/reduction of blue and yellow spectral bands of the transmitted light. Furthermore, color nets can act by maintaining leaf water content in rain-fed fig trees, thereby allowing for a compensation of water deficiency in fruits during ripening and drying on the tree. In summary, the net technique can prove beneficial for agricultural performance and for improvements in fruit yield where fig trees are in semi-dry regions and reside exclusively under rain-fed conditions and used water harvesting technique.
Acknowledgments
The authors wish to extend their thanks and appreciation to the personnel of Estahban Fig Research Station for their collaborations.
References
- Abul-Soud, M.A., M.S.A. Emam, and M.A.A. Abdrabbo. 2014. Intercropping of some Brassica crops with mango trees under different net house color. Res. J. Agric. Biol. Sci. 10:70–79.
- Alkalia-Tuvia, S., A. Goren, Y. Perzelan, T. Weinberg, and E. Fallik. 2014. The influence of colored shade nets on pepper quality after harvest-a possible mode-of-action. Poljopr. I Sumar 60:7.
- Ao, C., A. Li, A.A. Elzaawely, T.D. Xuan, and S. Tawata. 2008. Evaluation of antioxidant and antibacterial activities of Ficus microcarpa L. fil. extract. Food Control 19(10):940–948. doi: https://doi.org/10.1016/j.foodcont.2007.09.007.
- Arnon, D.I. 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris.** Plant Physiol. 24:1.
- Baraldi, R., F. Rossi, O. Facini, F. Fasolo, A. Rotondi, M. Magli, and F. Nerozzi. 1994. Light environment, growth and morphogenesis in a peach tree canopy. Physiol. Plant. 91:339–345. doi: https://doi.org/10.1111/j.1399-3054.1994.tb00442.x.
- Bartolozzi, F., and G. Fontanazza. 1999. Assessment of frost tolerance in olive (Olea europaea L.). Scientia Hortic **81 81(3):309–319. doi: https://doi.org/10.1016/S0304-4238(99)00019-9.
- Bastías, R.M., L. Manfrini, and L.C. Grappadelli. 2012. Exploring the potential use of photo-selective nets for fruit growth regulation in apple. Chil. J. Agric. Res. 72:224. doi: https://doi.org/10.4067/S0718-58392012000200010.
- Bergquist, S. 2006. Bioactive compounds in baby spinach (Spinacia oleracea L.). Agriculturae Sueciae, Acta Univ., SLU, Sveriges lantbruksuniv., Sweden, Doctoral thesis
- Bergquist, S.Å.M., U.E. Gertsson, L.Y.G. Nordmark, and M.E. Olsson. 2007. Ascorbic acid, carotenoids, and visual quality of baby spinach as affected by shade netting and postharvest storage. J. Agric. Food Chem. 55:8444–8451. doi: https://doi.org/10.1021/jf070396z.
- Campbell, R.J., R.P. Marini, and B. JB. 1992. Canopy position affects light response curves for gas exchange characteristics of apple spur leaves. Journal of the Amer.Soc. Hortic.Sci. 1 117(3):467–472. doi: https://doi.org/10.21273/JASHS.117.3.467.
- Cartechini, A., and A. Palliotti. 1995. Effect of shading on vine morphology and productivity and leaf gas exchange characteristics in grapevines in the field. Amer.J. Enol. Vitic 46(2):227–234.
- Díaz-Pérez, J.C. 2013. Bell pepper (Capsicum annum L.) crop as affected by shade level: Microenvironment, plant growth, leaf gas exchange, and leaf mineral nutrient concentration. HortScience. 48:175–182. doi: https://doi.org/10.21273/HORTSCI.48.2.175.
- Do Amarante, C.V.T., C.A. Steffens, and L.C. Argenta. 2011. Yield and fruit quality of ‘Gala’and ‘Fuji’apple trees protected by white anti-hail net. Sci. Hortic. (Amsterdam). 129:79–85. doi: https://doi.org/10.1016/j.scienta.2011.03.010.
- Dueñas, M., J.J. Pérez-Alonso, C. Santos-Buelga, and T. Escribano-Bailón. 2008. Anthocyanin composition in fig (Ficus carica L.). J. Food Compos. Anal. 21:107–115. doi: https://doi.org/10.1016/j.jfca.2007.09.002.
- Dussi, M.C., G. Giardina, D. Sosa, R.G. Junyent, A. Zecca, and P.R. Reeb. 2005. Shade nets effect on canopy light distribution and quality of fruit and spur leaf on apple cv. Fuji. Spanish J. Agric. Res. 3:253–260. doi: https://doi.org/10.5424/sjar/2005032-144.
- Fao, I., and W.W. Unicef. 2017. The state of food security and nutrition in the world. Italy Food Agric. Organ. United Nations, Rome.
- Fernández-Trujillo, J.P., J.M. Serrano, and J.A. Martínez. 2009. Quality of red sweet pepper fruit treated with 1-MCP during a simulated post-harvest handling chain. Food Sci.Tech. Inter. 15(1):23–30. doi: https://doi.org/10.1177/1082013208100464.
- Folta, K.M., and S.D. Carvalho. 2015. Photoreceptors and control of horticultural plant traits. HortScience. 50:1274–1280. doi: https://doi.org/10.21273/HORTSCI.50.9.1274.
- Fu, W., P. Li, and Y. Wu. 2012. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. (Amsterdam). 135:45–51. doi: https://doi.org/10.1016/j.scienta.2011.12.004.
- Gindaba, J., and S.J.E. Wand. 2007. Climate‐ameliorating measures influence photosynthetic gas exchange of apple leaves. Ann. Appl. Biol. 150:75–80. doi: https://doi.org/10.1111/j.1744-7348.2006.00110.x.
- Goren, A., S. Alkalia-Tuvia, Y. Perzelan, Z. Aharon, and E. Fallik. 2011. Photoselective shade nets reduce postharvest decay development in pepper fruits. Adv. Hortic. Sci. 25:26–31.
- Hiscox, J.D., and G.F. Israelstam. 1979. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 57:1332–1334. Pollution. 131: 453-459. doi: https://doi.org/10.1139/b79-163.
- Hunsche, M., M.M. Blanke, and G. Noga. 2010. Does the microclimate under hail nets influence micromorphological characteristics of apple leaves and cuticles? J. Plant Physiol. 167:974–980. doi: https://doi.org/10.1016/j.jplph.2010.02.007.
- Iglesias, I., and S. Alegre. 2006. The effect of anti-hail nets on fruit protection, radiation, temperature, quality and profitability of ‘Mondial Gala’apples. J. Appl. Hortic. 8:91–100. doi: https://doi.org/10.37855/jah.2006.v08i02.22.
- Ilić, Z.S., L. Milenković, L. Stanojević, D. Cvetković, and E. Fallik. 2012. Effects of the modification of light intensity by color shade nets on yield and quality of tomato fruits. Sci. Hortic. (Amsterdam). 139:90–95. doi: https://doi.org/10.1016/j.scienta.2012.03.009.
- Ilić, Z.S., L. Milenković, L. Šunić, and E. Fallik. 2015. Effect of coloured shade‐nets on plant leaf parameters and tomato fruit quality. J. Sci. Food Agric. 95:2660–2667. doi: https://doi.org/10.1002/jsfa.7000.
- Ilić, Z.S., L. Milenković, L. Šunić, S. Barać, J. Mastilović, Ž. Kevrešan, and E. Fallik. 2017. Effect of shading by coloured nets on yield and fruit quality of sweet pepper. Zemdirbyste-Agriculture. 104: 53- 62. doi: https://doi.org/10.13080/z-a.2017.104.008.
- Johkan, M., K. Shoji, F. Goto, S. Hashida, and T. Yoshihara. 2010. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience. 45:1809–1814. doi: https://doi.org/10.21273/HORTSCI.45.12.1809.
- Jokar, A., H. Zare, A. Zakerin, and A. Aboutalebi Jahromi. 2021. Effects of shade net colors on mineral elements and postharvest shelf life and quality of fresh fig (Ficus carica L.) under rain-fed condition. horticulturae. 7(93): 1–12. doi:https://doi.org/10.3390/horticulturae7050093
- Kalcsits, L., S. Musacchi, D.R. Layne, T. Schmidt, G. Mupambi, S. Serra, M. Mendoza, L. Asteggiano, S. Jarolmasjed, and S. Sankaran. 2017. Above and below-ground environmental changes associated with the use of photoselective protective netting to reduce sunburn in apple. Agric. For. Meteorol. 237:9–17. doi: https://doi.org/10.1016/j.agrformet.2017.01.016.
- Kim, D.O., S.W. Jeong, and C.Y. Lee. 2003. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 81(3):321–326. doi: https://doi.org/10.1016/S0308-8146(02)00423-5.
- Kong, Y., L. Avraham, Y. Perzelan, S. Alkalai-Tuvia, K. Ratner, Y. Shahak, and E. Fallik. 2013. Pearl netting affects postharvest fruit quality in ‘Vergasa’sweet pepper via light environment manipulation. Sci. Hortic. (Amsterdam). 150:290–298. doi: https://doi.org/10.1016/j.scienta.2012.11.029.
- Kosma, C., V. Triantafyllidis, A. Papasavvas, G. Salahas, and A. Patakas. 2013. Yield and nutritional quality of greenhouse lettuce as affected by shading and cultivation season. Emirates J. Food Agric. 25:974–979. doi: https://doi.org/10.9755/ejfa.v25i12.16738.
- Li, K.-T., and J. Syvertsen. 2006. Young tree growth and leaf function of citrus seedlings under colored shade netting. HortScience. 41:1022C– 1022. doi: https://doi.org/10.21273/HORTSCI.41.4.1022C.
- Lobos, G.A., J.B. Retamales, J.F. Hancock, J.A. Flore, S. Romero-Bravo, and A. Del Pozo. 2013. Productivity and fruit quality of Vaccinium corymbosum cv. Elliott under Photo-selective Shading Nets ** Sci. Hortic. (Amsterdam). 153:143–149. doi: https://doi.org/10.1016/j.scienta.2013.02.012.
- McCaskill, M.R., L. McClymont, I. Goodwin, S. Green, and D.L. Partington. 2016. How hail netting reduces apple fruit surface temperature: A microclimate and modelling study. Agric. For. Meteorol. 226:148–160. doi: https://doi.org/10.1016/j.agrformet.2016.05.017.
- Mihailova, G., K. Kocheva, V. Goltsev, H.M. Kalaji, and K. Georgieva. 2018. Application of a diffusion model to measure ion leakage of resurrection plant leaves undergoing desiccation. Plant Physiol. Biochem. 125:185–192. doi: https://doi.org/10.1016/j.plaphy.2018.02.008.
- Miller, G.A.D., N. Suzuki, S. Ciftci‐Yilmaz, and R.O.N. Mittler. 2010. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant. Cell Environ. 33:453–467. doi: https://doi.org/10.1111/j.1365-3040.2009.02041.x.
- Mupambi, G., B.M. Anthony, D.R. Layne, S. Musacchi, S. Serra, T. Schmidt, and L.A. Kalcsits. 2018. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. (Amsterdam). 236:60–72. doi: https://doi.org/10.1016/j.scienta.2018.03.014.
- Nicolás, E., V.L. Barradas, M.F. Ortuño, A. Navarro, A. Torrecillas, and J.J. Alarcón. 2008. Environmental and stomatal control of transpiration, canopy conductance and decoupling coefficient in young lemon trees under shading net. Environ. Exper.Bot.1. 63:200–206. doi: https://doi.org/10.1016/j.envexpbot.2007.11.007.
- Racsko, J., and L.E. Schrader. 2012. Sunburn of apple fruit: Historical background, recent advances and future perspectives. CRC. Crit. Rev. Plant Sci. 31:455–504.
- Rajapakse, N.C., and Y. Shahak. 2007. 12 Light-quality manipulation by horticulture industry. Annu. Plant Rev. Light Plant Dev. 30:290.
- Ranganna, S. 1986. Handbook of analysis and quality control for fruit and vegetable products. Tata McGraw-Hill Education.
- Rylski, I., and M. Spigelman. 1986. Effect of shading on plant development, yield and fruit quality of sweet pepper grown under conditions of high temperature and radlation. Sci. Hortic. 29:31–35. doi: https://doi.org/10.1016/0304-4238(86)90028-2.
- Sen, F., R. Oksar, and M. Kesgin. 2016. Effects of shading and covering on ‘Sultana Seedless’ grape quality and storability. J. Agric. Sci. Technol. 18:245–254.
- Shahak, Y., E. Gal, Y. Offir, and D. Ben-Yakir. 2008. Photoselective shade netting integrated with greenhouse technologies for improved performance of vegetable and ornamental crops. **international Workshop on Greenhouse Environmental Control and Crop Production in Semi-Arid Regions 797:75–80.
- Shahak, Y., E.E. Gussakovsky, E. Gal, and R. Ganelevin. 2004. ColorNets: Crop protection and light-quality manipulation in one technology, in: VII International Symposium on Protected Cultivation in Mild Winter Climates: Production, Pest Management and Global Competition 659. pp. 143–151.
- Solomakhin, A., and M.M. Blanke. 2010. Can coloured hailnets improve taste (sugar, sugar: Acid ratio), consumer appeal (colouration) and nutritional value (anthocyanin, vitamin C) of apple fruit? LWT-Food Sci. Technol. 43:1277–1284. doi: https://doi.org/10.1016/j.lwt.2010.02.020.
- Solomakhin, A., and M.M. Blanke. 2008. Coloured hailnets alter light transmission, spectra and phytochrome, as well as vegetative growth, leaf chlorophyll and photosynthesis and reduce flower induction of apple. Plant Growth Regul. 56:211–218. doi: https://doi.org/10.1007/s10725-008-9302-7.
- Stamps, R.H. 2009. Use of colored shade netting in horticulture. HortScience. 44:239–241. doi: https://doi.org/10.21273/HORTSCI.44.2.239.
- Szymańska, R., I. Ślesak, A. Orzechowska, and J. Kruk. 2017. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 139:165–177. doi: https://doi.org/10.1016/j.envexpbot.2017.05.002.
- Taiz, L., and E. Zeiger. 2012. Text book of plant physiology, p. 546–580. Published by sinauer associate Inc publishers., Sunderland. Massachusetts. USA.
- Tanny, J. 2013. Microclimate and evapotranspiration of crops covered by agricultural screens: A review. Biosyst. Eng. 114:26–43. doi: https://doi.org/10.1016/j.biosystemseng.2012.10.008.
- Wagenmakers, P., and M. Tazelaar. 1999. Resulting light reduction determines the future of hail nets in the Netherlands. Fruitt. (Den Haag) 89:10–11.
- Zhou, K., D. Jerszurki, A. Sadka, L. Shlizerman, S. Rachmilevitch, and J. Ephrath. 2018. Effects of photoselective netting on root growth and development of young grafted orange trees under semi-arid climate. Sci. Hortic. (Amsterdam). 238:272–280. doi: https://doi.org/10.1016/j.scienta.2018.04.054.
- Zibordi, M., S. Domingos, and L. Corelli Grappadelli. 2009. Thinning apples via shading: An appraisal under field conditions. J. Hortic. Sci. Biotechnol. 84:138–144. doi: https://doi.org/10.1080/14620316.2009.11512611.