
ABSTRACT
Strawberry flowers are hermaphroditic and most cultivars are self-compatible, but strawberry plants can produce a mixture of fruit that have arisen from self-pollination and cross-pollination. We aimed to determine whether size, shape, color, taste attributes and shelf life differ between self-pollinated and cross-pollinated fruit competing on the same Redlands Joy strawberry plant. Experiments were repeated at three fertilizer levels to determine the consistency of responses across different levels of crop nutrition. Cross-pollinated fruits were 4.2%–7.5% heavier and 2.7%–5.0% longer than self-pollinated fruits, with the differences being greater at lower levels of nutrient supply. They were also darker or redder than self-pollinated fruits. Cross-pollinated fruits sometimes had lower °Brix (sugar) or lower acidity than self-pollinated fruits, with the effect on sugar:acid ratio depending on the nutrient level. The shelf life of cross-pollinated fruits was approximately 4 days longer than self-pollinated fruits on plants that received medium or high levels of fertilizer. The results suggest that pollen-parent effects on fruit size become evident when self- and cross-pollinated strawberry fruits compete for resources on the same plant, and that the expression of pollen-parent effects can be affected by the level of crop nutrition.
Introduction
Flowers of strawberry (Fragaria × ananassa Duch.) contain hundreds of simple carpels, each of which possesses a single stigma, style and ovule (Ariza et al., Citation2015; Hancock, Citation2020; Petran and Hoover, Citation2018). Successful fertilization of an ovule leads to formation of a filled achene, i.e. the true fruit or strawberry “seed” (Perkins-Veazie, Citation1995). The individual achenes are embedded within an aggregated receptacle, and auxin production by multiple achenes causes enlargement of the surrounding receptacle tissue to form the pseudocarp, i.e. the strawberry “fruit” (Ariza et al., Citation2015; Hancock, Citation2020; Petran and Hoover, Citation2018). Strawberry flowers are hermaphroditic and most strawberry cultivars are self-compatible, but they may not be completely self-fertilizing because the stigmas become receptive before anthers of the same flower release pollen, thus promoting cross-pollination (Albano et al., Citation2009; Kämper et al., Citation2022; Tuohimetsä et al., Citation2014). Either self-pollen from the same cultivar or cross-pollen from a different cultivar may be deposited on the stigmas through the action of insects (MacInnis and Forrest, Citation2019). As a result, a mixture of self-pollinated and cross-pollinated fruits can be present on the same strawberry plant (Yanagi et al., Citation2017).
Pollen parentage can influence the yield and fruit quality of many horticultural crops (Herbert et al., Citation2019a, b; Seal et al., Citation2013; Tuohimetsä et al., Citation2014; Wallace and Lee, Citation1999; Wallace et al., Citation2002). The effects of different pollen parents on characteristics of the seed, fruit and accessory structures such as the receptacle, including size, shape, color, developmental timing and chemical composition, are termed xenia (Denney, Citation1992). Cross-pollination, when compared with self-pollination, has major impacts on fruit yield, mass and quality in, for example, almond (Kämper et al., Citation2021a), avocado (Degani et al., Citation1990), guava (Usman et al., Citation2013), kiwifruit (Seal et al., Citation2013), lychee (Stern et al., Citation1993), macadamia (Kämper et al., Citation2021b) and mandarin (Schneider et al., Citation2009; Wallace and Lee, Citation1999; Wallace et al., Citation2002). Cross-pollination increases yields by up to 30%–50% in avocado (Degani et al., Citation1990) and lychee (Stern et al., Citation1993) and increases fruit retention and kernel size in macadamia (Kämper et al., Citation2021b; Trueman et al., Citation2022). Cross-pollination can sometimes increase fruit mass in strawberry (Tuohimetsä et al., Citation2014), but little is known about the effects of cross-pollination on other fruit characteristics such as flavor, color, firmness and shelf life.
We have shown recently that cross-pollination does not affect yield or individual fruit mass in the self-compatible strawberry cultivar, Redlands Joy, although it does affect fruit color, taste and mineral nutrient concentrations (Dung et al., Citation2021). In that study, all of the flowers on each plant received the same pollination treatment, and so self-pollinated fruits did not compete with cross-pollinated fruits for maternal resources. The presence of competing cross-pollinated fruits can, however, decrease the mass, length and diameter of self-pollinated feijoa fruits (Patterson, Citation1990), decrease the mass and length of self-pollinated almond kernels (Oukabli et al., Citation2002), and decrease the mass of self-pollinated Banksia spinulosa seeds (Vaughton and Carthew, Citation1993) and Phormium tenax seeds (Becerra and Lloyd, Citation1992). These effects may be the result of cross-pollinated seeds producing more auxin, commencing growth earlier, and accruing maternal resources at the expense of self-pollinated seeds (Ozga and Reinecke, Citation2003; Quinet et al., Citation2019; Wietzke et al., Citation2018). If that were the case, then xenia effects on fruit size may become more evident when individual plants possess a mixture of self-pollinated and cross-pollinated fruit. We, therefore, hypothesized that self-pollinated fruits would be smaller than cross-pollinated fruits when both types of fruits were growing on the same strawberry plant.
Xenia effects on fruit and nut quality can also be influenced by mineral nutrient or carbohydrate supply (Brittain et al., Citation2014; Cunningham et al., Citation2020; Herbert et al., Citation2019b; Klein et al., Citation2014). Calcium fertilizer applications have the potential to affect the number of seeds set because calcium is required for pollen germination and pollen tube growth (Steinhorst and Kudla, Citation2013). Calcium applications also affect the quality of many horticultural crops, such as increasing the firmness and disease resistance of apple fruit (Fallahi et al., Citation1997; Marcelle, Citation1995; Martín-Diana et al., Citation2007), increasing the shelf life of pear fruit (Khala et al., Citation2017) and increasing the firmness of strawberry fruit (Cheng et al., Citation2011). Calcium increases fruit firmness and shelf life by strengthening fruit cell walls (Cheng et al., Citation2011). Calcium applications can also increase the mass (Kaya et al., Citation2002; Singh et al., Citation2007) and the total soluble solid (TSS) concentration, titratable acidity, firmness and shelf life of strawberry fruit (Singh et al., Citation2007). We hypothesized that pollen-parent effects on fruit quality would be influenced by the level of crop nutrition, with the expression of xenia effects being greatest when nutrients were in shortest supply.
In this study, we aimed to determine: 1) how xenia effects become apparent on fruit mass, size, shape, TSS concentration, acidity, color, firmness and shelf life when self-pollinated and cross-pollinated strawberry fruit are growing on the same plant; and 2) how consistent is the expression of xenia effects across different levels of crop nutrition.
Materials and Methods
Plant Material
Two strawberry cultivars, Redlands Joy and Rubygem (Herrington et al., Citation2007; Morrison and Herrington, Citation2002), that have been selected and grown commercially under subtropical conditions in eastern Australia (Menzel and Toldi, Citation2010; Menzel et al., Citation2014; Menzel, Citation2019) were used in this study. Bare-rooted runners were obtained from Sweets Strawberry Runners (Stanthorpe, Australia) in May 2018 and each runner was transplanted into a 4.5 L cylindrical pot filled with coco-peat (EC <1 mS/cm, pH = 5.5–7.0) and perlite (4:1, v:v), with 2.5 g of Osmocote fertilizer (N:P:K = 19.6:16.0:5.0% w/w, plus trace elements) (Scotts International, Heerlen, The Netherlands). The pot dimensions were 20 cm, 20 cm and 15 cm for the depth, top diameter and base diameter, respectively. The potted plants were placed in a glasshouse at the University of the Sunshine Coast, Sippy Downs, Australia (26°43‘S 153°03‘E), with plant spacing of 20 cm from pot to pot (Lieten, Citation2002). Fertilizer regime, irrigation, cultivation practices, and the temperature and irradiance in the glasshouse during the experimental period have been described previously (Dung et al., Citation2021).
Experimental Design
Ten plants of cultivar, Redlands Joy, were arranged into each of three experiments, with the three experiments undertaken under different levels of crop nutrition. The plants received eight sprays of calcium as Grotek Cal-Max (Ca : Mg : Fe = 3.0 : 2.0 : 0.1% w/w) (GS Distribution, Langley, Canada) at 1, 2 or 4 kg elemental Ca ha−1 spray−1 in experiments (a), (b) and (c), respectively. Each plant received approximately 5 mL of solution at each spray. The first spray was performed at first-flower petal-fall. The second spray was performed 7 d later, and subsequent sprays were applied at 14-d intervals. The experiments were conducted from June 2018 until the end of flowering in October 2018.
Each flower on each experimental plant was allocated to one of two pollination treatments: (1) pollination by Redlands Joy (Self) or (2) pollination by Rubygem (Cross). Successive flowers were either cross-pollinated or self-pollinated, creating a 50/50 mixture of self-pollinated and cross-pollinated flowers on each plant. The pollination treatment applied to the first flower on each plant (i.e. Self or Cross) was rotated successively from one experimental plant to the next, with the first flower of five plants in each experiment being self-pollinated while the first flower of the other five plants was cross-pollinated. Each flower had its anthers removed before the flower opened and was then covered with a fine-mesh bag until the stigma was receptive. We collected and stored pollen, and conducted pollinations, using methods described previously (Dung et al., Citation2021).
Fruit Measurements
Fruit set was 100% for each pollination type and so fruits from all flowers on each plant were harvested at commercial maturity when they became fully red (). Each fruit was numbered consecutively within each pollination treatment. We measured time to maturity of fruit, and fruit mass, length, diameter, color and firmness, using methods described previously (Dung et al., Citation2021). Briefly, time to maturity for each fruit was calculated as the number of days from first hand-pollination to harvest. The fresh mass, length and diameter of each fruit, excluding the pedicel and sepals, were recorded. Color was assessed for each fruit using a CR-10 colorimeter (Konica Minolta, Chiyoda, Japan) in the L*a*b* color space. Firmness of each fruit was determined using a Bareiss Hpe-II fruit penetrometer (Bareiss Prüfgerätebau, Oberdischingen, Germany) with a 3-mm diameter probe (Dung et al., Citation2021). The fresh mass, length and diameter of each fruit were recorded for all fruits per treatment in each calcium experiment. Firmness of each fruit, with the exception of the tenth to twelfth fruits harvested from each plant for each pollination treatment (i.e. fruit numbers 10–12), was determined. The tenth to twelfth fruits were instead stored at 4°C to determine shelf life. Fruit shelf life was measured using our previously described methods (Dung et al., Citation2021). All fruits, with the exception of fruits used to measure shelf life, were then frozen at −20°C prior to further quality analyses.
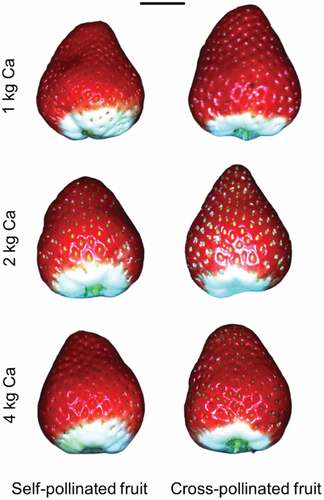
Figure 1. Self-pollinated and cross-pollinated Redlands Joy strawberry fruit under three levels of calcium nutrition (1, 2 or 4 elemental Ca ha −1 spray −1). Scale bar = 1 cm. Photographs: Cao Dinh Dung.
A total of 50 fruits per pollination treatment, being five frozen fruits within each treatment (fruit numbers 1, 4, 8, 14 and 19) from each plant, were used to measure numbers of fertilized and unfertilized seeds, TSS concentration and acidity. The measurement methods were described previously (Dung et al., Citation2021). TSS (°Brix) (i.e. sugar) and acid concentration of the filtered extract were determined using a PAL-BX|ACID4 sugar and acidity meter (Atago, Tokyo, Japan) (Kallio et al., Citation2000). Sugar:acid ratio was calculated for each fruit.
Statistical Analysis
Data were analyzed with IBM SPSS Statistics v.26 using generalized linear models (GLMs) with linear distributions and identity link functions. Each of the fruit quality parameters including length, diameter, mass and color was included as a response variable in separate models for each experiment. Pollination, plant, and fruit number nested within plant were regarded as main factors. Data was square-root transformed, when required, to achieve normality of the distributions. Treatment differences were regarded as significant at P < .05. Means are presented with standard errors.
Results
Fruit Size and Shape
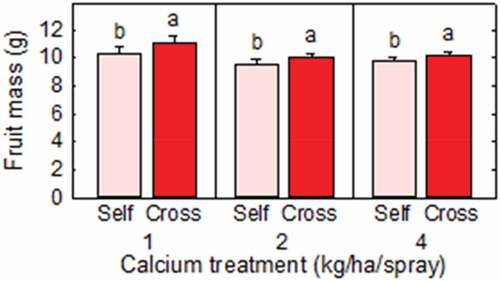
Cross-pollination increased the fresh mass of individual “Redlands Joy” strawberry fruit within all Ca levels (). Cross- and self-pollinated fruit, respectively, weighed 11.09 ± 0.45 g and 10.32 ± 0.45 g at the 1 kg Ca level, 10.07 ± 0.25 g and 9.53 ± 0.38 g at the 2 kg Ca level, and 10.20 ± 0.26 g and 9.79 ± 0.23 g at the 4 kg Ca level. These values represented increases in fresh mass following cross-pollination of 7.5%, 6.7% and 4.2%, respectively.
Figure 2. Individual mass of Redlands Joy strawberry fruit arising from self-pollination (Self) or cross-pollination by cultivar Rubygem (Cross) under three levels of calcium nutrition. Means + SE with different letters within a calcium level are significantly different (GLM; P < .05; n = 236–252 fruit).
Cross-pollination increased fruit length by 5.0%, 2.7% and 2.7% within the 1, 2 and 4 kg Ca levels, respectively, and increased fruit diameter by 2.6% at the 2 kg Ca level (). Cross-pollinated fruit had slightly greater length:diameter ratio than self-pollinated fruit at the 1 kg Ca level, but length:diameter ratio did not differ significantly between cross- and self-pollinated fruit within the other Ca levels (). Cross-pollinated fruit reached maturity approximately 1 d earlier than self-pollinated fruit at the 4 kg Ca level, but time to maturity did not differ significantly between cross- and self-pollinated fruit within the other Ca levels (). The percentage of fertilized seeds on each fruit was high and did not differ significantly between cross- and self-pollinated fruit within any Ca level ().
Table 1. Length, diameter, shape, time to maturity, and percentage of seeds that were fertilized for Redlands Joy strawberry fruit arising from self-pollination (Self) or cross-pollination by cultivar Rubygem (Cross) under three levels of calcium nutrition.
Fruit Color and Taste Attributes
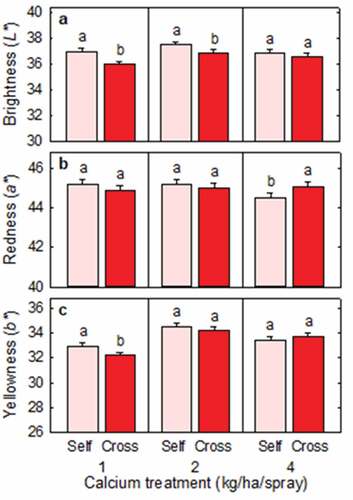
Cross-pollinated fruit were darker than self-pollinated fruit at the 1 kg and 2 kg Ca levels () and redder than self-pollinated fruit at the 4 kg Ca level (). Cross-pollinated fruit were also less yellow than self-pollinated fruit at the 1 kg Ca level ().
Figure 3. (a) Brightness (L*), (b) redness (a*), and (c) yellowness (b*) of Redlands Joy strawberry fruit arising from self-pollination (Self) or cross-pollination by cultivar Rubygem (Cross) under three levels of calcium nutrition. Means + SE with different letters within a calcium level are significantly different (GLM; P < .05; n = 236–253 fruit).
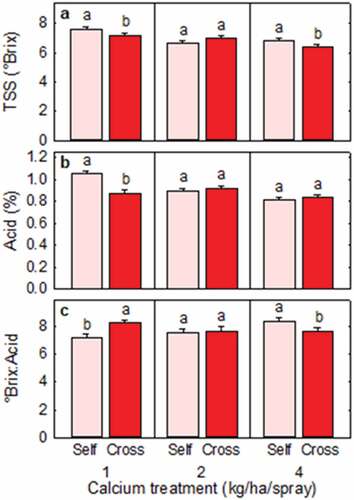
Cross-pollinated fruit had lower TSS concentration (°Brix) than self-pollinated fruit at the 1 kg and 4 kg Ca levels, but TSS concentration did not differ significantly at the 2 kg Ca level (). Cross- and self-pollinated fruit had °Brix values of 7.2 ± 0.2 and 7.6 ± 0.2, respectively, at the 1 kg Ca level and 6.4 ± 0.2 and 6.8 ± 0.2, respectively, at the 4 kg Ca level. Cross-pollinated fruit had lower acid concentration than self-pollinated fruit at the 1 kg Ca level, but acid concentrations did not differ significantly within the other Ca levels (). Cross- and self-pollinated fruit had acid concentrations of 0.87 ± 0.03% and 1.05 ± 0.03%, respectively, at the 1 kg Ca level. As a result, cross-pollinated fruit had higher sugar:acid of 8.28 ± 0.19 compared with 7.24 ± 0.20 for self-pollinated fruit at the 1 kg Ca level, but lower sugar:acid of 7.71 ± 0.19 compared with 8.40 ± 0.26 at the 4 kg Ca level (). Sugar:acid ratio did not differ significantly between cross- and self-pollinated fruit at the 2 kg Ca level ().
Figure 4. (a) Concentration of total soluble solids (TSS; °brix), (b) acid (%), and (c) sugar:acid ratio (°brix:acid) of Redlands Joy strawberry fruit arising from self-pollination (Self) or cross-pollination by cultivar Rubygem (Cross) under three levels of calcium nutrition. Means + SE with different letters within a calcium level are significantly different (GLM; P < .05; n = 50 fruit).
Fruit Firmness and Shelf Life
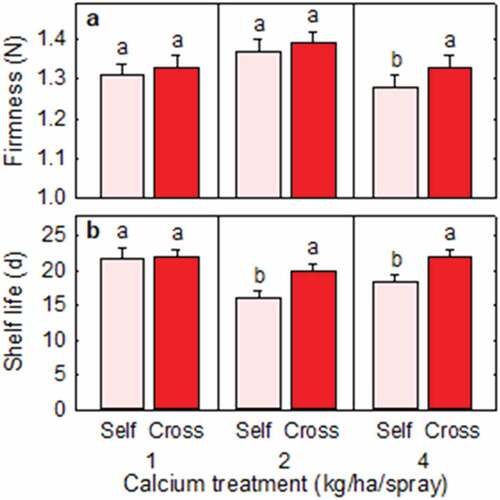
Cross-pollinated fruit were firmer than self-pollinated fruit at the 4 kg Ca level, but firmness did not differ significantly between cross- and self-pollinated fruit within the other Ca levels (). Cross- and self-pollinated fruit had firmness of 1.33 ± 0.03 N and 1.28 ± 0.03 N, respectively, at the 4 kg Ca level. Cross-pollinated fruit had longer shelf life than self-pollinated fruit at the 2 kg and 4 kg Ca levels (). Cross- and self-pollinated fruit had shelf lives of 19.9 ± 1.2 d and 16.0 ± 1.1 d, respectively, at the 2 kg Ca level and 22.0 ± 1.1 d and 18.3 ± 1.0 d, respectively, at the 4 kg Ca level.
Figure 5. (a) Firmness and (b) shelf life of Redlands Joy strawberry fruit arising from self-pollination (Self) or cross-pollination by cultivar Rubygem (Cross) under three levels of calcium nutrition. Means + SE with different letters within a calcium level are significantly different (GLM; P < .05; n = 207–223 fruit for firmness; n = 28–30 fruit for shelf life).
Discussion
Xenia effects were found among cross-pollinated and self-pollinated fruits produced on the same plant of Redlands Joy strawberry. Cross-pollinated fruits were longer and sometimes broader, resulting in greater fresh mass, and these effects were amplified when nutrient supply was lowest. Cross-pollination also affected fruit shape, color, flavor, firmness and shelf life, with these effects also often depending on the level of crop nutrition. The results confirm our first hypothesis that self-pollinated fruits would be smaller than cross-pollinated fruits when both types of fruit were growing on the same plant. They also confirm our second hypothesis that xenia effects would be influenced by the level of crop nutrition, with the expression of xenia effects often being greatest when nutrients were in shortest supply.
Xenia effects on fruit characteristics are sometimes confounded by pollen-parent effects on the percentage of fertilized ovules in multi-seeded fruit (de Jong and Scott, Citation2007). Cross-pollinated strawberry fruits in the current study had a similar percentage of fertilized seeds to self-pollinated fruits, but they were heavier and longer than self-pollinated fruits. In our previous study (Dung et al., Citation2021), the number of fertilized seeds did not differ significantly between cross-pollinated fruits and self-pollinated when these fruits developed on different plants; however, cross-pollinated fruits were darker red, sweeter tasting, and had higher concentrations of some mineral nutrients. Therefore, differences in strawberry fruit size and quality were true xenia effects (Denney, Citation1992), related to the paternity of developing seeds rather than differences in fertilization success.
Cross-pollinated and self-pollinated Redlands Joy fruits did not differ significantly in mass, length or length:diameter in our previous study in which the two types of fruits were growing on different plants (Dung et al., Citation2021). The same cross-pollen source, Rubygem, was used in both the current and previous studies. However, cross-pollinated fruits had higher mass and length, and sometimes higher length:diameter ratio, than self-pollinated fruits in the current study in which the two types of fruits were growing on the same plant. The relative difference in fruit mass declined from 7.5% to 4.2%, and the relative difference in fruit length declined from 5.0% to 2.7%, as the calcium application rate increased from 1 to 4 kg calcium ha−1 spray−1. Furthermore, a significant xenia effect on length:diameter ratio, i.e. fruit shape, was only evident under the lowest nutrient supply. These results indicate that cross-pollinated fruits out-competed self-pollinated fruits on the same strawberry plant, exhibiting stronger xenia effects on fruit size and shape when resource supply was low. Cross-pollinated pear (Quinet et al., Citation2019), feijoa (Patterson, Citation1990) and almond fruit (Oukabli et al., Citation2002) are also larger than self-pollinated fruit when they develop on the same plant.
Cross-pollinated Redlands Joy fruits were darker but not redder than self-pollinated fruits on different plants (Dung et al., Citation2021). They also had higher sugar:acid ratio than self-pollinated fruits, solely because of reduced acid concentrations (Dung et al., Citation2021). Pollen parentage often affects fruit color and sweetness (Denney, Citation1992). For example, cross-pollination increases the darkness and redness of kiwifruit and pear fruit (Dussi and Sugar, Citation1995; Seal et al., Citation2013), and increases the sweetness of mandarin (Citrus reticulata Blanco) and sweet cherry (Prunus avium L.) fruit (Bhat et al., Citation2015; Wallace and Lee, Citation1999). In the current study, cross-pollinated fruits were darker than self-pollinated fruits at the 1 kg and 2 kg ha−1 spray−1 calcium levels and they were redder than self-pollinated fruits at the 4 kg ha−1 spray−1 calcium level. They also had lower acidity at the 1 kg ha−1 spray−1 calcium level. However, cross-pollination decreased TSS concentration at the 1 kg and 4 kg ha−1 spray−1 calcium level. As a result, cross-pollination increased sugar:acid at the low nutrient level but decreased sugar:acid at the high nutrient level. These results again demonstrate xenia effects on the appearance and taste attributes of strawberry fruit, and highlight that xenia effects on color, TSS concentration and acidity can be affected by the level of crop nutrition (Dung et al., Citation2022). Red color development in strawberry fruit is associated with the accumulation of anthocyanin pigments, which are beneficial for human health (Given et al., Citation1998; Lila, Citation2004). Consumer acceptance of fruit can rely heavily on the color of the product, and consumers prefer strawberry fruit with darker red color (Bhat et al., Citation2015; Westland and Shin, Citation2015). Consumers also prefer sweet fruit over sour fruit (Bhat et al., Citation2015). Fruit TSS and acid concentrations determine strawberry fruit flavor (Petran and Hoover, Citation2018; Wietzke et al., Citation2018), and so growers might increase the consumer appeal of their crop by planting multiple varieties in close proximity to each other and managing bee populations to maximize cross-pollen transfer.
Cross-pollinated fruits were sometimes firmer or had longer shelf life than self-pollinated fruits. In contrast to the results for fruit size, shape, color and taste attributes, significant xenia effects on firmness were only evident at the 4 kg ha−1 spray−1 calcium level and significant effects on shelf life were only evident at the 2 kg and 4 kg ha−1 spray−1 calcium levels. Self-pollinated fruit had short shelf life at higher calcium levels, and cross-pollination raised shelf life toward the duration obtained from both self- and cross-pollinated fruit at the low calcium level. Cross-pollinated fruits at the higher calcium levels were possibly able to accumulate more calcium than self-pollinated fruit, thus making them firmer (Cheng et al., Citation2011; Jiménez-Bermúdez et al., Citation2002; Martín-Diana et al., 207). Higher fruit firmness is often associated with longer shelf life of strawberry fruit (Jiménez-Bermúdez et al., Citation2002), and longer shelf life plays an important role in reducing postharvest losses of strawberry fruit (Vu et al., Citation2011).
Conclusions
Xenia effects on strawberry fruit mass and size were apparent when cross-pollinated and self-pollinated fruits competed for maternal resources on the same plant, with cross-pollination sometimes increasing fruit redness, darkness, sugar:acid ratio, firmness or shelf life. Xenia effects were influenced by the level of crop nutrition, with cross-pollination effects on fruit mass and size being amplified when nutrients were in shortest supply. Strawberry growers could establish farms with multiple cultivars in close proximity to each other, manage bee hives to transfer more pollen between cultivars, and schedule fertilizer applications to optimize fruit size and fruit quality.
Acknowledgments
Thanks to Tracey McMahon, Bruce Randall, Brittany Elliott, Tsvakai Gama, Ian Darby, Darren Morrow and Michael Nielsen for assistance.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Albano, S., E. Salvado, S. Duarte, A. Mexia, and P.A.V. Borges. 2009. Pollination effectiveness of different strawberry floral visitors in Ribatejo, Portugal: Selection of potential pollinators. Part 2. Advanc. Hortic. Sci. 23:246–253.
- Ariza, M.T., C. Soria, and E. Martínez-Ferri. 2015. Developmental stages of cultivated strawberry flowers in relation to chilling sensitivity. AoB Plants 7:lv012. doi: 10.1093/aobpla/plv012.
- Becerra, J.X., and D.G. Lloyd. 1992. Competition-dependent abscission of self-pollinated flowers of Phormium tenax (Agavaceae): A second action of self-incompatibility at the whole flower level? Evol. 42:45–69. doi: 10.2307/2409864.
- Bhat, R., J. Geppert, E. Funken, and R. Stamminger. 2015. Consumers perceptions and preference for strawberries – case study from Germany. Int. J. Fruit. Sci 15:405–424. doi: 10.1080/15538362.2015.1021408.
- Brittain, C., C. Kremen, A. Garber, and A.M. Klein. 2014. Pollination and plant resources change the nutritional quality of almonds for human health. PLOS ONE 9:e90082. doi: 10.1371/journal.pone.0090082.
- Cheng, F., H. Liu, H. Yang, S. Lai, X. Cheng, Y. Xin, B. Yang, H. Hou, Y. Yao, S. Zhang, et al. 2011. Quality attributes and cell wall properties of strawberries (Fragaria × ananassa Duch.) under calcium chloride treatment. Food Chem. 126:450–459. doi: 10.1016/j.foodchem.2010.11.009.
- Cunningham, S.A., M.J. Evans, M. Neave, J. Armstrong, and P.S. Barton. 2020. Pollination and resource limitation as interacting constraints on almond fruit set. Plant Biol. 22:113–119. doi: 10.1111/plb.13045.
- de Jong, T.J., and R.J. Scott. 2007. Parental conflict does not necessarily lead to the evolution of imprinting. Trends Plant Sci. 12:439–443. doi: 10.1016/j.tplants.2007.07.003.
- Degani, C., A. Goldring, I. Adato, and R. El-Batsri. 1990. Pollen parent effect on outcrossing rate, yield, and fruit characteristics of ‘Fuerte’ avocado. HortScience 25:471–473. doi: 10.21273/HORTSCI.25.4.471.
- Denney, J.O. 1992. Xenia includes metaxenia. HortScience 27:722–728. doi: 10.21273/HORTSCI.27.7.722.
- Dung, C.D., H.M. Wallace, S.H. Bai, S.M. Ogbourne, and S.J. Trueman. 2021. Cross-pollination affects fruit colour, acidity, firmness and shelf life of self-compatible strawberry. PLOS ONE 16:e0256964. doi: 10.1371/journal.pone.0256964.
- Dung, C.D., H.M. Wallace, S.H. Bai, S.M. Ogbourne, and S.J. Trueman. 2022. Biomass and mineral nutrient partitioning among self-pollinated and cross-pollinated fruit on the same strawberry plant. PLOS ONE 17:e0269485. doi: 10.1371/journal.pone.0269485.
- Dussi, M.C., and D. Sugar. 1995. Characterizing and quantifying anthocyanins in red pears and the effect of light quality on fruit color. J. Amer. Soc. Hortic. Sci. 120:785–789. doi: 10.21273/JASHS.120.5.785.
- Fallahi, E., W.S. Conway, K.D. Hickey, and C.E. Sams. 1997. The role of calcium and nitrogen in postharvest quality and disease resistance of apples. HortScience 32:831–835. doi: 10.21273/HORTSCI.32.5.831.
- Given, N.K., M.A. Venis, and D. Grierson. 1998. Hormonal regulation of ripening in the strawberry, a non-climacteric fruit. Planta 174:402–406. doi: 10.1007/BF00959527.
- Hancock, J.F. 2020. Strawberries. CAB International, Boston, MA.
- Herbert, S.W., D.A. Walton, and H.W. Wallace. 2019a. Pollen-parent affects fruit, nut and kernel development of Macadamia. Sci. Hortic. 244:406–412. doi: 10.1016/j.scienta.2018.09.027.
- Herbert, S.W., D.A. Walton, and H.W. Wallace. 2019b. The influence of pollen-parent and carbohydrate availability on macadamia yield and nut size. Sci. Hortic. 251:241–246. doi: 10.1016/j.scienta.2019.03.006.
- Herrington, M.E., C.K. Chandler, J.A. Moisander, and C.E. Reid. 2007. ‘Rubygem’ strawberry. HortScience 42:1482–1483. doi: 10.21273/HORTSCI.42.6.1482.
- Jiménez-Bermúdez, S., J. Redondo-Nevado, J. Muñoz-Blanco, J.L. Caballero, J.M. López-Aranda, V. Valpuesta, F. Pliego-Alfaro, M.A. Quesada, and J.A. Mercado. 2002. Manipulation of strawberry fruit softening by antisense expression of a pectate lyase gene. Plant Physiol. 128:751–759. doi: 10.1104/pp.010671.
- Kallio, H., M. Hakala, A.M. Pelkkikangas, and A. Lapveteläinen. 2000. Sugars and acids of strawberry varieties. Eur. Food Res. Technol. 212:81–85. doi: 10.1007/s002170000244.
- Kämper, W., C.D. Dung, S.M. Ogbourne, H.M. Wallace, and S.J. Trueman. 2022. High self-paternity levels and effects of fertilised-seed number on size of strawberry fruit. PLOS ONE 17:e0273457. doi: 10.1371/journal.pone.0273457.
- Kämper, W., G. Thorp, M. Wirthensohn, P. Brooks, and S.J. Trueman. 2021a. Pollen paternity can affect kernel size and nutritional composition of self-incompatible and new self-compatible almond cultivars. Agron 11:326. doi: 10.3390/agronomy11020326.
- Kämper, W., S.J. Trueman, S.M. Ogbourne, and H.M. Wallace. 2021b. Pollination services in a macadamia cultivar depend on across-orchard transport of cross-pollen. J. Appl. Ecol. 58:2529–2539. doi: 10.1111/1365-2664.14002.
- Kaya, C., H. Kirnak, D. Higgs, and K. Saltali. 2002. Supplementary calcium enhances plant growth and fruit yield in strawberry cultivars grown at high (NaCl) salinity. Sci. Hortic. 93:65–74. doi: 10.1016/S0304-4238(01)00313-2.
- Khala, K., N. Ahmadi, and M.K. Souri. 2017. Improvement of postharvest quality of Asian pear fruits by foliar application of boron and calcium. Hortic 3:15. doi: 10.3390/horticulturae3010015.
- Klein, A.M., S.D. Hendrix, Y. Clough, A. Scofield, and C. Kremen. 2014. Interacting effects of pollination, water and nutrients on fruit tree performance. Plant Biol. 17:1–8. doi: 10.1111/plb.12180.
- Lieten, F. 2002. The effect of humidity on the performance of greenhouse grown strawberry. Acta Hortic. 567:479–482. doi: 10.17660/ActaHortic.2002.567.101.
- Lila, M.A. 2004. Anthocyanins and human health: An in vitro investigative approach. J. Biomed. Biotechnol. 5:306–313. doi: 10.1155/S111072430440401X.
- MacInnis, G., and J.R.K. Forrest. 2019. Pollination by wild bees yields larger strawberries than pollination by honey bees. J. Appl. Ecol. 56:824–832. doi: 10.1111/1365-2664.13344.
- Marcelle, R.D. 1995. Mineral nutrition and fruit quality. Acta Hortic. 383:219–226. doi: 10.17660/ActaHortic.1995.383.22.
- Martín-Diana, A.B., C. Rico, J.M. Frías, J.M. Barat, G.T.M. Henehan, and C. Barry-Ryan. 2007. Calcium for extending the shelf life of fresh whole and minimally processed fruits and vegetables: A review. Trends Food. Sci. Technol. 18:210–218. doi: 10.1016/j.tifs.2006.11.027.
- Menzel, C.M. 2019. No evidence of excessive leaf production by strawberries grown in the subtropics. Agric 9:197. doi: 10.3390/agriculture9090197.
- Menzel, C.M., L.A. Smith, and J.A. Moisander. 2014. The productivity of strawberry plants growing under plastic high tunnels in a wet subtropical environment. Hort Tech. 24:334–342. doi: 10.21273/HORTTECH.24.3.334.
- Menzel, C.M., and A. Toldi. 2010. An evaluation of containerized plants for strawberries growing in a subtropical environment. Hort Tech. 20:786–793. doi: 10.21273/HORTTECH.20.4.786.
- Morrison, B., and M. Herrington. 2002. Strawberry breeding in Australia. Acta Hortic. 567:125–128. doi: 10.17660/ActaHortic.2002.567.18.
- Oukabli, A., A. Lansari, D.E. Walali-Loudiyi, and A. Abousalim. 2002. Effects of controlled self-pollination and cross-pollination on fruit set, embryo viability and pomological traits in the self-compatible almond cv ‘Tuono’. Acta Hortic. 591:429–435. doi: 10.17660/ActaHortic.2002.591.66.
- Ozga, J.A., and D.M. Reinecke. 2003. Hormonal interactions in fruit development. J. Plant Growth Regul. 22:73–81. doi: 10.1007/s00344-003-0024-9.
- Patterson, K.J. 1990. Effects of pollination on fruit set, size, and quality in feijoa (Acca sellowiana (berg) Burret). N. Z. J. Crop Hortic. Sci. 18:127–131. doi: 10.1080/01140671.1990.10428082.
- Perkins-Veazie, P. 1995. Growth and ripening of strawberry fruit. Hortic. Rev. 17:267–297.
- Petran, A., and E. Hoover. 2018. The flowers of Fragaria × ananassa: Morphology, response to photoperiod, and genetics of induction. Hortic. Rev. 45:1–31.
- Quinet, M., C. Buyens, P.I. Dobrev, V. Motyka, and A.L. Jacquemart. 2019. Hormonal regulation of early fruit development in European pear (Pyrus communis cv. ‘Conference’). Hortic 5:9. doi: 10.3390/horticulturae5010009.
- Schneider, D., M. Goldway, N. Rotman, I. Adato, and R.A. Stern. 2009. Cross-pollination improves ‘Orri’ mandarin fruit yield. Sci. Hortic. 122:380–384. doi: 10.1016/j.scienta.2009.06.009.
- Seal, A.G., J.K. Dunn, H.N. De Silva, T.K. McGhie, and R.C.M. Lunken. 2013. Choice of pollen parent affects red flesh colour in seedlings of diploid Actinidia chinensis (kiwifruit). N. Z. J. Crop Hortic. Sci 41:207–218. doi: 10.1080/01140671.2013.803129.
- Singh, R., R.R. Sharma, and S.K. Tyagi. 2007. Pre-harvest foliar application of calcium and boron influences physiological disorders, fruit yield and quality of strawberry (Fragaria × ananassa Duch.). Sci. Hortic. 112:215–220. doi: 10.1016/j.scienta.2006.12.019.
- Steinhorst, L., and J. Kudla. 2013. Calcium – a central regulator of pollen germination and tube growth. Biochim. Biophys. Acta. 1833:1573–1581. doi: 10.1016/j.bbamcr.2012.10.009.
- Stern, R.A., S. Gazit, R. El-Batsri, and C. Degani. 1993. Pollen parent effect on outcrossing rate, yield, and fruit characteristics of ‘Floridian’ and ‘Mauritius’ lychee. J. Amer. Soc. Hortic. Sci 118:109–114. doi: 10.21273/JASHS.118.1.109.
- Trueman, S.J., W. Kämper, J. Nichols, S.M. Ogbourne, D. Hawkes, T. Peters, S. Hosseini Bai, and H.M. Wallace. 2022. Pollen limitation and xenia effects in a cultivated mass-flowering tree Macadamia integrifolia (Proteaceae). Ann. Bot. 129:135–146. doi: 10.1093/aob/mcab112.
- Tuohimetsä, S., T. Hietaranta, M. Uosukainen, S. Kukkonen, and S. Karhu. 2014. Fruit development in artificially self- and cross-pollinated strawberries (Fragaria × ananassa) and raspberries (Rubus idaeus). Acta Agric. Scand. B Soil Plant Sci 64:408–415. doi: 10.1080/09064710.2014.919348.
- Usman, M., W.A. Samad, B. Fatima, and M.H. Shah. 2013. Pollen parent enhances fruit size and quality in intervarietal crosses in guava (Psidium guajava). Int. J. Agric. Biol. 15:125–129.
- Vaughton, G., and S.M. Carthew. 1993. Evidence for selective fruit abortion in Banksia spinulosa (Proteaceae). Biol. J. Linn. Soc. 50:35–46. doi: 10.1111/j.1095-8312.1993.tb00917.x.
- Vu, K.D., R.G. Hollingsworth, E. Leroux, S. Salmieri, and M. Lacroix. 2011. Development of edible bioactive coating based on modified chitosan for increasing the shelf life of strawberries. Food Res. Inter. 44:198–203. doi: 10.1016/j.foodres.2010.10.037.
- Wallace, H.M., B.J. King, and L.S. Lee. 2002. Pollen flow and the effect on fruit size in an ‘Imperial’ mandarin orchard. HortScience 37:84–86. doi: 10.21273/HORTSCI.37.1.84.
- Wallace, H.M., and L.S. Lee. 1999. Pollen source, fruit set and xenia in mandarins. J. Hortic. Sci. Biotechnol. 74:82–86. doi: 10.1080/14620316.1999.11511077.
- Westland, S., and M.J. Shin. 2015. The relationship between consumer colour preferences and product-colour choices. J. Inter. Colour Assoc. 14:47–56.
- Wietzke, A., C. Westphal, P. Gras, M. Kraft, P. Pfohl, P. Karlovsky, E. Pawelzik, T. Tscharntke, and I. Smit. 2018. Insect pollination as a key factor for strawberry physiology and marketable fruit quality. Agric. Ecosyst Environ. 258:197–204. doi: 10.1016/j.agee.2018.01.036.
- Yanagi, T., H. Miura, S. Isobe, N. Okuda, and Y. Yoshida. 2017. Effect of insect pollinator on inbreeding versus outbreeding in open pollinated strawberry seeds. Sci. Hortic. 215:112–116. doi: 10.1016/j.scienta.2016.12.015.