ABSTRACT
Components of the Pacific transported landscape have been used as proxies to trace the prehistoric movement of humans across the Pacific for almost two decades. Analyses of archaeological remains and DNA sequences of plants, animals, and microorganisms moved by or with humans have contributed to understanding prehistoric migration, trade, exchange, and sometimes revealed the geographic origins of particular plants and animals. This paper presents the basic elements of a DNA-based commensal model and discusses the phylogenetic and population genetic approaches these models employ. A clear delineation of the underlying assumptions of these models and the background information required to construct them have yet to appear in the literature. This not only provides a framework with which to construct a commensal model but also highlights gaps in current knowledge. The ways in which commensal models have enriched archaeological reconstructions will be highlighted, as will their current limitations. With these limitations in mind, options will be outlined for augmenting commensal models through the application of established techniques and new technologies in order to provide the best tools for reconstructing ancient human mobility and behavior in the Pacific and beyond.
INTRODUCTION
A “commensal model” is focused on the examination of biological variation in species that have been transported by or with humans. This variation reflects the translocation of specific organisms and so is a suitable proxy for tracing the routes by which each individual species was moved by human agents in the past (Matisoo-Smith Citation1996; Matisoo-Smith and Robins Citation2004). The term commensal derives from the Latin commensalis, com meaning together and mensalis of the table; interpreted as sharing a table. A commensal, in the context of archaeological/anthropological investigation, has become a broadly encompassing term for any species that has a close association with, and is translocated by, humans (Matisoo-Smith Citation2009). Commensal species are often plants and animals that are used for food, although they also include species with utilitarian and other uses, as well as pathogenic and non-pathogenic microbial species. Consistent with the anthropological literature, this broad definition of commensal is the one adopted in this paper. It should be clear, however, that this definition differs from the ecological definition of commensalism as a biological interaction, where one species benefits while the other remains unaffected (Martin and Hine Citation2008). While the ecological definition of commensalism may explain the relationship for some species that traveled on Pacific canoes (e.g., skinks), other relationships such as mutualism and parasitism also feature. Rather than distinguishing between these different types of biological interaction—which is often difficult, especially when dealing with plant and animal species outside of the native distributions of their wild relatives—we use the term commensal to refer to any species that was translocated by humans.
The use of non-human organisms as a proxy for human mobility has several additional strengths. Arguably the most important is to provide a means to examine migration and interaction without using ancient human remains or modern human DNA. Many people have well-founded concerns about how their genetic material may be stored, used, interpreted, or even patented without their knowledge or explicit consent. The destructive nature of most current analytical techniques also means that with very few ancient remains currently available for study, the permanent loss of human skeletal material is a serious concern. Furthermore, the lack of genetic variation observed in human mitochondrial DNA in many Pacific populations (Lum and Cann Citation2000; Murray-McIntosh et al. Citation1998) and the issues of DNA preservation in tropical environments (Hagelberg and Clegg Citation1993; Robins et al. Citation2001) make the use of abundant and widely dispersed animal and plant remains an attractive alternative for studying human mobility.
The Pacific is not the only region in which the biomolecules and distributions of domestic or commensal plants and animals has been used as a means of understanding prehistoric human behavior, but it does possess a number of unique features. The Pacific is the region in which both the oldest human translocations of animals in island environments have been documented, dating to as early as 20,000 years ago (White Citation2004), and the region encompassing some of the most recent, large-scale human migrations that involved the movement of plants, animals, and microorganisms. The Pacific, specifically Remote Oceania, also differs from other island systems for which plant and animal translocation are used to understand human mobility in being relatively isolated from continental land masses, the Americas to the east and Asia to the west. However, commensal models can also be applied in other regions of the world and have provided an alternative means of understanding prehistoric human migration and interaction in the Mediterranean (for examples, see Cheylan et al. Citation1998; Reese Citation1996; Vigne and Valladas Citation1996), Sweden (Fraser et al. Citation2012), the Channel Islands of California (Collins Citation1991; Rick et al. Citation2008; Rick et al. Citation2009), the islands of Mocha and Chiloe off the Chilean coast (Becker Citation1997), and in the Caribbean (de France et al. Citation1996; Giovas et al. Citation2012; Wing Citation1993).
Pacific commensal species are of two major types: those that were purposefully moved as part of a transported landscape, and those that were unintentionally transported, including infectious organisms or stowaways. Transported landscapes are those that contain all or most of the plant and animal stocks that recreate the range of subsistence items found at colonists’ homeland in new locales (Anderson Citation1952; Kirch Citation1997:217–220). Some commensals, including geckos, skinks, and garden snails (Kirch Citation2001; White Citation2001), are believed to have been inadvertently transported. However, it is not always possible to confidently distinguish unintentional from deliberate introductions. Lee et al. (Citation2007) have argued that some Polynesian snail species were purposefully traded for their shells, which were used in the production of jewellery. In addition, a recent study of cricket translocation suggested that these insects may have been intentionally introduced to some Remote Oceanic islands (Tinghitella et al. Citation2011). A list of all species that have been studied to understand human migration and interaction in the Pacific through DNA analysis can be found in , and the locations of islands and archipelagos is illustrated in Figure 1.
Table 1 Number of individual sequences/genotypes for each commensal organism for specific archipelagos or islands, as shown in Figure 1. Genetic data are further distinguished by whether they were taken from chronologically secure ancient specimens or from a period after European contact, in which case data are considered to be modern. In the case of Polynesian Outliers, only those with published commensal data are included.
The development of a commensal model requires several steps to establish a clear association between genetic data and ancient human behavior. The first step is to understand the characteristics of the organism that is to be used as a proxy. The researcher must understand its natural range and origins, reproductive biology, capacity for self-dispersal, adaptability, and value to humans. As part of the first step the researcher must clearly document the association between specific organisms and humans through a critical examination of the existing archaeological and historical evidence. Once it has been established that the organism can and has been moved with humans, a review of the published genetic data may allow for the identification of genetic markers with sufficient resolution to distinguish individuals from one another. Of particular interest are markers that display phylogeographic signal that may reveal something about the origin of the individuals sampled. Defined sequence types (haplotypes) should be examined for their relationships to archaeological assemblages, chronology, and geography. An additional step is required if the reconstruction of prehistoric human mobility is to be based only on evidence from modern DNA (that is, DNA collected from extant populations). This requires an attempt to document the introductions of specific organisms from the time of their first known association with humans in the region of interest up until the time the modern samples were acquired. These introductions can include overlapping waves of importation derived from multiple potential source populations over extended time periods by a variety of agents. Finally, the results, within their archaeological and historical context, can be used to explore the role of humans in creating the genetic patterns observed in space and through time.
The examination of genetic patterns across time and space has been a major focus of Pacific commensal research over the past 15 years. Although other research fields—especially archaeology, zooarchaeology, linguistics, and ethnobiology—can provide insights into commensal histories, we cannot, for space reasons, review that literature here. Instead, our focus is on how specific elements of these fields, especially research on adaptability and human choice, can be integrated into DNA-based models to understand the translocation of organisms by humans. The aim of this paper is to delineate the underlying assumptions of genetic (DNA-based) models, and describe the steps involved in constructing these paradigms.
BUILDING A COMMENSAL MODEL
Documenting the Characteristics of a Transported Organism
Understanding the natural range and origins of a commensal species.
The first step in building any commensal model is to identify the natural range and habitat of the animal, plant, or microorganism to be used as a proxy. Most of the animals introduced to Pacific islands have Southeast Asian origins and do not naturally occur in the Pacific (Matisoo-Smith Citation2007a). This is not so, however, for many of the economically important plant species, many of which are of Oceanic origin (Whistler Citation1991) nor several endemic New Guinea species such as phalangers, moved in early prehistoric times (White Citation2004). This may complicate issues where native and introduced lineages have overlapping geographic ranges. In such cases it must first be established whether introduced varieties are reproductively isolated from wild ancestral types. The possibility of hybridization events between wild and introduced lineages and what effect this might have on interpretations of results generated from the analysis of genetic markers should also be assessed.
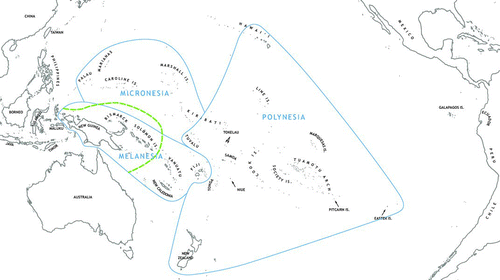
Figure 1 Map of the Pacific (modified from Kirch Citation2000a:6) showing major islands and archipelagos, the traditional geographic divisions of Melanesia, Micronesia, and Polynesia, and the division between Near and Remote Oceania, indicated by the broken line east of the main Solomon Chain (color figure available online).
Documenting dispersal mechanisms and charting distributions
It is essential that the potential for self-dispersal be examined for plants and animals that are to be used as models for human mobility. This can often be accomplished through researching the published literature relating to modes of dispersal, proclivity for migration, and morphological constraints on dispersal. Direct dating of animal bones and investigations of faunal assemblages from eras before human settlement provides lines of evidence to support claims for human-mediated transport of animals. For example, the archaeological distribution of chickens has been documented (Storey et al. Citation2008) and faunal assemblages recovered from Pacific islands archaeological deposits pre-dating colonization do not contain the remains of domestic fowl, supporting their association with humans (Steadman Citation2006). However, one must also be conscious that in some cases human settlement and its associated environmental impacts increases the abundance of some native species, or allows for the self-introduction of exotic species (for avian examples, see Trewick and Gibb Citation2010). Either of these events could be misinterpreted, when examining the archaeological record alone, as the introduction of a commensal species by humans.
Plants present a more difficult case for assessment of the timing and mode of introduction because they are rarely preserved in the archaeological record, and have not always been the target of systematic collection from archaeological excavations. Pacific plant remains recovered from archaeological sites, both as macro-remains and microfossils, include coconut (Kirch and Hunt Citation1988; Lepofsky et al. Citation1992; Spriggs Citation1984), sweet potato (Coil and Kirch Citation2005; Hather and Kirch Citation1991; Ladefoged et al. Citation2005; Orliac and Orliac Citation1998; Skjølsvold Citation1961), bottle gourd (Burney et al. Citation2001; Irwin Citation2004), breadfruit (McCoy et al. Citation2010), and pandanus nuts (Summerhayes et al. Citation2010). The presence of some plants may be attested through residue studies and associated technological innovations and tools (Green Citation2005), as well as by linguistic evidence (Green Citation2000). Secondary evidence, such as the building of garden alignments or the use of specific agricultural techniques, can also be used to identify the prehistoric presence of specific plants, as has been done with sweet potato (e.g., Coil and Kirch Citation2005; Ladefoged et al. Citation2011). However, in most cases, the presence of plant species in specific prehistoric contexts must be established through inference using early historic accounts that mention particular species (e.g., Beaglehole Citation1962; Ladefoged et al. Citation2009; McNab Citation1914). Further complicating plant commensal models is the fact that some species can disperse naturally via wind, water (by drifting on currents or in rafts of floating debris) or by animals (e.g., in bird faeces). Arguments exist both for (Ward and Brookfield Citation1992) and against (Dennis and Gunn Citation1971) the ability of coconuts to survive at sea and successfully germinate upon washing up on a new beachfront. However, wild and domestic coconuts differ in their morphology which, in turn, affects their ability to disperse naturally. It is likely that humans preferentially selected coconut varieties that had a high water content, which is thought to inhibit long-distance water dispersal (Zizumbo-Villarreal and Quero 1998). This property may increase their suitability for use in developing commensal models.
A number of factors influence the survival of commensal remains in the archaeological record, including taphonomy, differential disposal, and consumption by animals such as pigs. In areas with highly acidic soils, faunal remains are rarely recovered and so the initial introduction and persistence of animals may be impossible to reconstruct. Where soil conditions are conducive to preservation, other factors may act to obscure the true density and distribution of food remains. In some regions refuse may have been thrown into the sea (McCoy and Cleghorn Citation1988:110) which generally results in the destruction of organics. In rare instances, however, remains are preserved in anaerobic environments leading to the retrieval of organics rarely found in other Pacific island sites. Archaeological sites such as Talepakemalai in Mussau, New Guinea (Kirch Citation2001; Kirch et al. Citation1991) and the swampy deposits at Huahine in the Society Islands (Lepofsky Citation1988; Sinoto and McCoy Citation1975) show the potential such deposits hold for reconstructing prehistoric faunal and floral communities.
Other factors may also influence disposal behavior and subsequent recovery, such as cultural values attached to specific animals and the effects of scavengers. It has been suggested that superstitions related to dogs may have resulted in differential deposition of their remains and could account for the absence of evidence for canines in some Pacific archaeological contexts (Bulmer 1976; Bulmer Citation2001; Davidson Citation1969). In addition, observations of pigs and dogs scavenging bones from contemporary garbage heaps may be extended into prehistory as a potential agent inhibiting the preservation of faunal and floral remains in archaeological assemblages (Reitz and Wing Citation1999; Spennemann Citation1990; Takayama and Intoh Citation1978; Trotter Citation1979).
Dispersal mechanisms for microorganisms must also be carefully considered before they can be used as proxies for human mobility. Ideally for commensal models, a microbial species will be vertically transmitted. This means that the virus or bacterium is passed only from human parents to their offspring, linking the evolution of these microbes to the genealogies of their hosts. Horizontally transmitted microorganisms can also be used, in some limited circumstances, to trace human migration. Horizontal transmission implies that an organism or virus can be passed by direct or indirect contact between individuals. In the Pacific, microorganisms used to trace the relationships between prehistoric Pacific populations include malarial parasites (Plasmodium falciparum) (Lum et al. Citation2004), the gram negative stomach bacterium Helicobacter pylori (Moodley et al. Citation2009), human herpesvirus-8 (HHV-8) (Rezza et al. Citation2001), human T-lymphotropic virus type 1 (HTLV-1) (Takao et al. Citation2000), JC virus (JCV) (Takasaka et al. Citation2004; Yanagihara et al. Citation2002), and human papilloma virus type 16 (HPV) (Watts et al. Citation2002). However, very few, if any of these have a fully vertical mode of transmission. HTLV-1 is transmitted through breastfeeding, sexual contact, and blood (Carneiro-Proietti et al. Citation2006), yet horizontal transmission through populations is rare (Ohkura et al. Citation1999). The JC Virus may be passed through contact with urine and, thus is horizontally transmitted (but has been observed to be stable within families). Multiple JCV infections in individuals are rare and, thus, recombination is considered unlikely which also increases its suitability as a commensal model (Pavesi Citation2004). In the study of Helicobacter pylori there is observed interfamily clustering of the bacteria, suggesting transmission along family lines. However, the bacteria can also be spread through saliva, vomit, faeces, and contaminated water supplies and is often found at much higher prevalence in crowded institutional conditions (Brown Citation2000). For a review of microorganisms and what they contribute to the reconstruction of Pacific settlement, the reader is referred to Czarnecki et al. (Citation2007).
Assessing adaptability.
In order to survive transport and successfully establish in their new homes, commensal species must either be sufficiently adaptable or have sufficient effort invested in their husbandry. Specific adaptations to life on Pacific islands may be unique. Rats are known to be particularly adaptable and, as scavengers, require no intentional human care once they are introduced to an island. Dogs are highly adaptable omnivores, and it has been suggested that some of the dogs living on Pacific islands were fed only a vegetarian diet and obtained protein from scavenging scraps (Titcomb and Pukui Citation1969), hunting, or eating wild foods (Bulmer Citation2001). In addition, several island dog populations have become skilled at fishing, both singly and as packs, an activity that can still be observed in many Pacific coastal villages (Mills Citation2004; Titcomb and Pukui Citation1969:31).
In Oceania there is a noticeable west-to-east decline in the number of plant species that were successfully introduced to islands and archipelagos and this is likely related to the adaptability, successful transport, and acclimation of cultigens to new environments (Whistler Citation1991). Many of the plant species that were successfully introduced to Eastern and Marginal Polynesia were those that were able to be propagated vegetatively (without seed) (Hinkle Citation2004; Yen Citation1991). These include ti (Cordyline fruticosa), pandanus (Pandanus sp.), breadfruit (Artocarpus altilis, A. camansi, A. mariannensis), taro (Colocasia esculenta), yams (Dioscorea alata, D. esculenta), paper mulberry (Broussonetia papyrifera), and sweet potato (Ipomoea batatas). Of the species that are able to reproduce vegatatively, some, such as taro and paper mulberry, can persist without human intervention and have the ability to form wild or feral populations (Matthews Citation1995). While it might be predicted that a species’ distribution will be correlated with its ability to survive in varied habitats, this is not always the case. For example, in prehistoric New Zealand, sweet potato and bottle gourd were so important that Māori modified their agricultural practices to ensure the success of these crops in a climate where both species would have otherwise struggled to survive (Best Citation1925; Harris et al. Citation2000).
Assessing the role of human choice in the establishment and dispersal of commensal species.
White (Citation2001) asserted that people have a very clear understanding of the requirements for the transportation of domesticated and commensal animals. Therefore, several choice options should be considered to understand the observed density and distribution of commensal species. The first choice made by voyagers is whether or not to intentionally transport and introduce any particular plant or animal to a pristine island environment. Once introduced, populations must be maintained, which may mean providing protection from predators including other domesticates such as pigs and dogs. In the case of introduced animals, populations may also require feeding and thus impact upon the overall food surplus (Bay-Petersen Citation1984; Giovas Citation2006; Kirch Citation2000b). Finally, should an island's population of animals or plants become endangered or extinct, people can later choose to use their established trade networks to reintroduce species or merely to trade to meet their immediate needs for end products. Trade and exchange can account for the presence of some prehistoric faunal remains and is evidenced in the ethnographic record for islands which cannot bear the production or maintenance costs of particular animals, such as pigs (Baldwin Citation1990).
The plants selected by ancient Pacific seafarers were also typically multipurpose plants. Ti (C. fruticosa) plants served as packaging, raw material for sandals and costumes, and had cultural and religious associations (Hinkle Citation2004). Breadfruit is important in eastern Polynesia as a storable resource in the form of ma (the fruit fermented in pits and stored for several years), and the leaves were used to wrap food and to create rope. Timber from breadfruit trees was widely used for house and ship construction, bowls, coffins, and fishing floats (Ragone Citation1991).
Understanding whether a species was transported intentionally or unintentionally (e.g., stowaways) may be important to how commensal data are interpreted. Purposeful behavior provides evidence about choices and perhaps motivations, with respect to colonization and trade. However, stowaways and infections may reveal interactions that did not include institutionalized or ritual exchange of living organisms and potentially point to unintentional drift voyages.
Choosing an Appropriate Genetic Marker to Determine Evolutionary Relationships
Mitochondrial and plastid/chloro- plast DNA.
Mitochondrial DNA is found in high copy number (between 1000 and 10,000 per cell) in the mitochondria of most eukaryotic cells (Clayton Citation1991). Due to its abundance, mtDNA is more commonly the target for study in ancient animal samples because nuclear DNA is rarely preserved in sufficient quantities or long enough fragments for traditional sequencing methods (Clayton Citation1991). Mitochondrial DNA is nearly always maternally inherited and recombination is thought to occur rarely if at all (Ho and Gilbert Citation2010; Pakendorf and Stoneking Citation2005). Because of the mitochondrion's cellular machinery, the DNA mutation rate is higher than the average mutation rate of nuclear DNA (Horai et al. Citation1995; Nei Citation1987; Pakendorf and Stoneking Citation2005). However, the mutation rates for mtDNA vary among species in both mammalian and avian genomes (Nabholz et al. Citation2009). Regions of the mtDNA genome with the greatest diversity, such as the control region (CR)/displacement loop (D-loop), are typically used for comparisons at the population level and are also those most often employed in commensal studies. However, studies of these regions are further complicated by the presence of mutational hotspots, which not only have an accelerated mutation rate but also are more prone to damage both within the cell and after the death of the organism (Pakendorf et al. Citation2006; Threadgold and Brown Citation2003). Much work is yet to be done to fully understand the implications this may have on the application of evidence from these regions and the interpretation of commensal models.
Compared with Y-chromosome or biparentally inherited nuclear DNA markers, it has been argued that the mtDNA of domestic animals is expected to show less localized geographic differentiation and more continuity through time (between ancient and modern populations) (Jobling et al. Citation2004:333). The assumption is that males produce far more offspring than females in managed breeding systems. In theory then, by sampling a large number of individuals it is possible to infer the hypothetical ancestor(s) from which all the observed variation has arisen (Tapio and Grigaliunaite Citation2003). However, these assumptions can often be violated because of human choice or environmental factors and so it is necessary to evaluate these assumptions and their applicability when using genetic data for the identification of domestication centers (e.g., Storey et al. Citation2012).
Although plants also possess mitochondrial DNA, due to its relative complexity in this kingdom it is generally not used in evolutionary studies. Instead, plastid DNA (usually referred to as chloroplast DNA; cpDNA) is often used, and this can be considered broadly equivalent to the mtDNA of animals. This is because cpDNA also has high copy number, is maternally inherited, does not undergo recombination and, in specific regions, has a high mutation rate (Olmstead and Palmer Citation1994; Palmer et al. Citation2012).
Nuclear DNA.
In addition to mitochondrial and chloroplast DNA, commensal studies often employ multilocus genomic fingerprinting techniques. These include Amplified Fragment Length Polymorphisms (AFLPs; Vos et al. Citation1995), Randomly Amplified Polymorphic DNA (RAPDs; Welsh and McClelland Citation1990; Williams et al. Citation1990), and Inter-Simple Sequence Repeats (ISSRs; Ziętkiewicz et al. 1994). These are all Polymerase Chain Reaction (PCR) based techniques that allow for amplification of DNA fragments from organisms for which there is no known reference sequence, and can therefore be used on organisms for which there is no a priori sequence information (Schlötterer Citation2004). Multilocus DNA fingerprinting techniques can be ideal in many of the situations that apply to studies of Pacific commensal species, such as when the organisms’ DNA sequences are not yet known, when genomic heterogeneity is high (i.e., when it is necessary to amplify many loci to ascertain an accurate measure of genomic diversity), when genetic variability is low (i.e., when it is necessary to amplify many loci to locate the few that are polymorphic), and in polyploids (Bensch and Åkesson Citation2005; Meudt and Clarke Citation2007). Previous studies using genomic fingerprinting techniques demonstrate their applicability to Pacific commensal studies (e.g., Clarke et al. Citation2006; Hinkle Citation2007; Zerega et al. Citation2004).
In addition, nuclear microsatellite markers have been widely used at the population level, and can be applied to Pacific commensal studies. When compared to genomic fingerprinting techniques, microsatellite markers are often slower and more expensive to develop, yet they are also considered more robust, especially when used across different labs (Belaj et al. Citation2003; Bensch and Åkesson Citation2005). A recent study examining the population structure and dispersal of coconuts used this approach in an attempt to better characterize the dispersal history of coconuts and discern episodes of purposeful human translocation of the species (Gunn et al. Citation2011).
Employing genetic methods to identify significant patterns.
Just as a typology allows archaeologists to group sets of like artifacts together, a phylogeny allows geneticists to group similar genetic sequences together based on their evolutionary history. Identical sequences which, by definition, share single nucleotide polymorphisms (SNPs) are designated as haplotypes. As the name implies, SNPs involve variation of a single base pair. While any nucleotide has the potential to mutate through transversion or transition to another nucleotide, SNP studies concentrate on those bases that show an appreciable frequency of variation within the population of interest and thus may differentiate populations through space and time. Phylogenetic analyses of genetic sequences estimate the relationships among them. Closely related haplotypes are called regional chronology has not yet haplogroups and these are usually seen as clades on phylogenetic trees.
Commensal studies that use genetic data typically employ a phylogeographic approach, that is: they aim to understand the relationship between genetic patterns and geographic space. Bradley (Citation2006:275) described phylogeography as having “two orthogonal dimensions—the horizontal one representing the geography of genetic diversity and the vertical one representing the time through which this geography emerges.” Ancient DNA (usually DNA from archaeological material) contributes to phylogeography by providing a means to date the appearance/disappearance of types, and examine continuity of genetic lineages through time. Ancient DNA can be obtained from macro remains of both animals and plants, although remains from the latter are rare, particularly from seedless plants, which are not often preserved in the archaeological record. For plants, increasing attention has been focussed on recovering DNA from microfossils (namely: pollen, starch and phytoliths) but this field is still developing. DNA has been successfully recovered from ancient pollen (e.g., Parducci et al. Citation2005) and modern but not yet ancient starch (Waiblinger Citation2012). Current evidence suggests that DNA may not be present in phytoliths (Elbaum et al. Citation2009). Although past studies have largely focussed on obtaining microfossil pollen DNA from sediments, future research may focus on microfossils trapped in ancient tools and teeth. This would help control for potential leaching/percolation of microfossils, which is a serious concern (Anderson-Carpenter Citation2011; Haile et al. Citation2007). In cases where ancient DNA cannot be obtained from archaeological material, herbarium and museum specimens can be used to extend the temporal range of studies (e.g., Clarke et al. Citation2006).
Documenting Historic Era Dispersals to Interpret Modern DNA Evidence
Ancient DNA is a powerful tool to determine prehistoric patterns of genetic diversity. However, due to a paucity of suitable archaeological material and technical limitations, amplification of ancient DNA is often not possible, and modern (contemporary) DNA sampling is used instead to infer prehistoric genetic diversity. Because of their poor representation in the archaeological record, this reliance on modern DNA alone is especially common for plants and microbes. In order to apply modern sequence data to the construction of commensal models an additional step is needed: in-depth research into historic commensal transfers, both within the Pacific and from outside ports to determine the relationship between prehistoric groups of animals and plants, and those found living on islands today. Without this historical context, the data can provide an extremely limited view of prehistory (Bradley Citation2006; Jobling Citation2012).
The application of modern DNA data to archaeological questions introduces new complexities not encountered when using ancient DNA. Most importantly, inferences using modern DNA data must be tempered with a clear understanding that contemporary processes (e.g., translocation, natural dispersal, selection and genetic drift) may have obscured ancient patterns.
Sampling and DNA sequencing/genotyping of modern individuals within populations provides a snapshot of genetic diversity at the time of sampling. The extent to which modern genetic diversity reflects past genetic diversity can vary. For plant translocations, Whistler (Citation1991:142) defined the time after which translocations could no longer be considered to be free of European influence as AD 1769 (the year of Cook's first voyage into the Pacific) due to the documented impact of Europeans on Pacific floras. Studies of plant genetics are further hindered by the complex interactions that still occur between contemporary Pacific populations and the historic and modern species moved by Europeans. This means that conclusions based on plant DNA studies must be considered in light of known historic and contemporary interaction spheres (Yen Citation1974).
An analogy to this problem is the limited interpretive power provided by a seriated typology developed from collections of conflated assemblages on the surface of a site where the regional chronology has not yet been established. As with surface-collected assemblages, it is unlikely that all types will be represented, nor will the assemblage adequately reflect all time periods of interest. Instead the assemblage reflects more modern processes that have intermixed ancient and modern signatures. This is not to imply studies based only on modern information are uninformative, but they are limited in their ability to reconstruct prehistoric translocations. To maximize these modern results they must be carefully examined within an archaeological and historic context to disentangle the layers of signal that may be observed.
A number of processes can change the frequencies of particular genetic signatures through time, and the influence of these processes will differ between species and locations. At best, contemporary genetic diversity will broadly reflect prehistoric genetic diversity; at worst, contemporary diversity at a particular location may be reflecting a full-scale replacement that occurred within the past decade. This is especially true for crop and livestock species, where the huge selective pressures exerted by humans for more desirable breeds and cultivars can cause complete genetic replacement in short periods. For examples, post-contact introductions and translocations of pigs, dogs, and chickens in the Pacific by Europeans may have changed both the phenotypes and genotypes of existing populations. It is likely that post-contact voyagers also moved domesticated animals in multiple directions, bringing both European breeds west from the Americas and Asian breeds east from the continents. As several of the important commensal species of the early Pacific settlers are of Asian origin, differentiating an ancient Asian phylogenetic signal from a more recent one may be incredibly difficult. The exception is the Pacific rat (Rattus exulans), the only Pacific commensal mammal that has escaped introgression with strains of animals introduced by European sailing ships (Matisoo-Smith Citation1994).
Several animal species, such as pigs and dogs, can swim between islands and may self-disperse from the island to which Europeans introduced them without human intervention or intent. Pigs were observed swimming between islands in the Malay archipelago by Wallace (Citation1869:81). In addition, diseases introduced with foreign stock are likely to have caused the loss of indigenous lineages of commensal plants and animals. The loss of these genetic lines is something that can only be observed using ancient DNA data (Matisoo-Smith Citation2002). Even where selective pressures are weak or dispersal is limited, natural processes such as genetic drift will cause changes if given enough time and sufficient isolation. These changes will occur more rapidly in small populations, such as those often found on islands.
Analysis of the historical literature including shipping records, naturalists’ accounts, missionary journals, and records of animal and plant disease, can provide valuable information about events that have changed genetic diversity through time. This information can be used to critically evaluate the extent to which modern sampling and genetic data can be used to infer prehistoric diversity. For example, the use of historical records helped to demonstrate that the frequency of Asian mtDNA signatures in modern European pig breeds was due to the importation of pigs from Asia to Europe in the eighteenth and nineteenth centuries to improve stocks, and was not a pattern which reflected the initial domestication or dispersal of pigs (Giuffra et al. Citation2000). Similar effects should be expected in the Pacific.
While modern DNA may certainly reveal some clues about prehistoric process, it will require the addition of ancient DNA data to reveal a fuller, temporally stratified picture of prehistoric migrations and interactions. Therefore, modern genetic information for domestic animals will be of limited use until pre-contact Pacific lineages can be reliably identified and separated from the European signatures.
FURTHER CONSIDERATIONS: PROSPECTS FOR ADVANCING COMMENSAL STUDIES
Commensal DNA studies have focused on the density, distribution, and chronology of faunal and floral introductions. This increased scrutiny on previously neglected faunal assemblages (see Nagaoka Citation1988), in particular, has led to a much deeper understanding of human-animal relationships that extends beyond documenting routes of transport. The development of commensal models has led to a greater appreciation for the nuances of specific interactions between prehistoric people and their traveling companions, both chosen and stowaways. Studies focused on commensal specimens have provided new perspectives on taphonomy, the cultural and religious significance of animals, differential disposal behavior, and the choices humans face in extirpation of species from their transported landscapes.
Due to the focus on plants and animals and their archaeological context, the timing of introductions of many of the major animal species has been more firmly resolved. It has become clear that the idea of singular packages of transported landscapes has limited utility. Animals and plants were moved to different places at different times to meet a variety of needs. The development of commensal models has highlighted the areas in which animals were introduced (e.g., the temporal delay in the introduction of Pacific rats to Micronesia) or not at all (e.g., New Caledonia) (Matisoo-Smith Citation2007a). Commensal models have also provided evidence for multiple, temporally distinct, introductions of some species, such as chickens (Storey et al. Citation2012).
Each commensal species has its own individual advantages and limitations for the construction of models. The different commensal species provide opportunities to explore different aspects of human settlement behavior and subsequent interaction. Comparison of the patterns of the various commensal species allows for a much more complete understanding of the complexities of prehistoric human behavior. As commensal studies progress, coverage may become sufficiently even through time and space to compare the patterns among species. Examining the similarities and differences in the translocation of domesticates and stowaways may reveal unique aspects of human behavior and mobility. A comparative approach may also prove fruitful in the application of several biomolecular methods to address the movement of specific plants and animals. DNA has proven useful in some situations, where it is available. Unfortunately it has been observed that the preservation of DNA in mammalian bones from open archaeological sites on tropical islands is the exception rather than the rule (Robins et al. Citation2001). It is also becoming increasingly clear that mtDNA markers may not always provide the best dataset with which to reconstruct prehistoric migration and interaction (Storey et al. Citation2012). In these situations other techniques may be used in conjunction with, or substituted for, DNA analyses in order to address questions related to the translocation of commensals in prehistory.
DNA is not the only biomolecular marker available to the commensal researcher. There are other techniques, such as stable isotope studies, that can and should be employed to understand the origins and mobility of animals using archaeologically recovered bone. Such methods will also need to be carefully considered with reference to potential dispersal mechanisms, adaptability, human choice, and post-contact movements and much of the material in this paper is also relevant to their use. Stable isotopes can and should be used whenever possible to supplement, and even perhaps replace studies of DNA. This is particularly true in cases where the genetic markers currently available are not sufficiently diverse to separate populations from different islands from one another. Several stable isotopes have been employed in the past to reconstruct the origins and migrations of humans and can be extended to the translocation of animals. Strontium isotopes are linked to local geology and can separate individuals who were born in regions with distinct geological histories (Bentley Citation2006) and thus may differentiate animals that were born and raised on high islands from those reared on coral atolls. Strontium is prone to contamination by the introduction of exogenous strontium through groundwater percolation, and sampling of enamel is preferential to the use of bone to avoid inaccurate results (Bentley Citation2006). Stable isotopes of sulphur may be used to differentiate local from foreign individuals (Richards et al. Citation2003) but can be complicated on small islands where the sea is always proximate, or on large islands were several sulphur profiles can occur. This may mean the questions about introductions or translocations need to be more narrowly defined in a Pacific context. In addition the sulphur content of rainwater is altered as sea spray contributes most of the atmospheric sulphur in the Remote Ocean (Thode Citation1991). Thus the sulphur profiles of small islands, such as atolls may be very similar to each other and therefore unsuitable as a means to discriminate origins. Oxygen isotopes were once considered a reliable means to differentiate the origins of individuals in a single burial environment, but are less reliable in Oceanic environments due to the dependence of oxygen fractionation on latitude, altitude, and distance from the sea (Schwarcz Citation2002). Similarly deuterium, the heavier stable isotope of hydrogen, has been successfully applied to tracking migrating birds and in forensic investigations (e.g., Brown et al. Citation2005) but it has not yet been successfully applied in archaeological contexts. Finally, in terms of oxygen and hydrogen isotopes it is not yet clear if the dynamic history of hydrology in a particular region and changes to the water cycle through time may influence the distribution of stable isotopes and thus skew results in ways not yet fully understood. Regardless of the isotope chosen, like DNA testing, isotopic determinations require the destruction of material and only in large and complete samples would one have the opportunity to obtain a DNA sequence, a radiocarbon date, and a full spectrum of isotopic information from the same sample. At present decisions must be made on a case-by-case basis as to which method is best suited to answering the archaeological question of interest given the particular circumstances of the assemblage under investigation.
In the case of understanding the ultimate origins, and breeding histories of animals, DNA analysis may still be the best method currently available. In fact with the advent of Next Generation Sequencing it may provide a whole new perspective on these fascinating aspects of human-animal relationships. It will also be interesting, once a large enough database is assembled, to compare and contrast the DNA patterns seen on island types. It has been noted that in the Mediterranean the genetic diversity in the nuclear DNA of black rats may be related to factors such as island size and relative isolation (Cheylen et al. Citation1998). In the case of interactions and attempts to reconstruct the more proximate sources of animals through time to establish trade and exchange relationships isotopes may provide more informative. For example, if animals were part of the sewai exchange between the Carolinian atolls and the larger continental island of Yap (Intoh Citation1992) then the study of stable isotopes of domestic animals is likely to be more fruitful to examine the source of individual animals.
THE COMMENSAL DATABASE
Several comprehensive reviews of the contributions commensal studies have made to Pacific prehistory have been written in the last five years (Czarnecki et al. Citation2007; Matisoo-Smith Citation2007a, 2007b, 2009) and it is not our intention to repeat that information in this paper. Instead we aim to highlight both the progress of, and gaps in, our current knowledge of commensal species. To do this a survey of published literature was undertaken and a summary of the available DNA data was compiled in . Individual sequences as reported in the papers are presented in aggregate by archipelago. This highlights both the sheer number of commensal species identified and studied in the past 15 years as well as the substantial geographic gaps in coverage. It is clear that the geographic coverage for commensal organisms using DNA from both modern and ancient sources is uneven and, in its current form, difficult to accurately compare across time and space. It is unlikely that there is a representative sample even of the potential diversity of modern commensals in Oceania. The existing database for examining ancient genetic signatures is even more sparsely populated.
It seems very likely that as the geographic coverage of commensals is expanded and larger sequence datasets are assembled that new haplotypes will be discovered. For example new dog haplotypes have recently been reported in New Guinea (Oskarsson et al. Citation2011). In addition the ancestral gene pool of the Pacific commensal species is often less well understood than their Oceanic descendants. The amount of genetic and archaeological work that remains to be done in Southeast Asian domestication centers is staggering (for examples see the summaries presented in Glover and Bellwood Citation2004; O’Reilly Citation2007). As a result the use of the term ‘Pacific clade’ to describe any haplogroup, for chickens (Dancause et al. Citation2011), dogs (Oskarsson et al. Citation2012), or pigs (Larson et al. Citation2005) is likely to be premature and will require much larger ancient DNA-based datasets. A larger sample from the Pacific as well as potential domestication centers will be necessary to ensure that extant Pacific haplogroups are truly unique as a result of genetic isolation and not due to inadequate sampling in other parts of the world.
The availability of samples is affected by a myriad of factors, including accessibility of islands and the survival of DNA in animal and plant remains during excavation, analysis, storage and/or transport to laboratories (for a discussion see Hinkle Citation2004; Robins et al. Citation2001). The unreliable preservation not only of DNA in archaeological remains but also of the remains themselves will always be a hindrance to developing a complete picture of prehistory. DNA studies are also destructive, and where only a few remains are available their use in commensal studies is not always the best way to answer the archaeological question of interest. Studies based on pathogenic commensals will largely be restricted to modern DNA evidence and are inhibited by modern rates of infection. For example in a study of 138 individuals from Easter Island, only one was infected with HTLV-1 (Ohkura et al. Citation1999). However, the numbers assembled in do provide a means of identifying gaps in the data for existing commensal coverage. Future researchers can endeavor to provide a more even coverage of all the islands and archipelagos of the Pacific.
As can be seen from , Polynesia has the best coverage of commensal organisms in the Pacific, with fewer sequences available from Near Oceania and Micronesia, and very few from the Polynesian outliers. Of 18 Polynesian outliers, only 4 have had samples included in commensal studies. Polynesian outliers may provide important corridors for the spread of commensals between Island Melanesia and Polynesia, especially in the period after the initial colonization of Polynesia. This hypothesis could be tested by the inclusion of commensal samples from Polynesian outliers in future research projects. For example Kirch (Citation2000a) has indicated that subtle dietary shifts are evident in the archaeological record of Tikopia after the arrival of Polynesians on the island. This included the introduction of Canarium almonds from Vanuatu, mosaic-tailed rats from the main Solomon Islands, and furthermore that changes in anthropophilic land snail populations may be linked to the movement of plants (Kirch Citation1986). In addition, it has been speculated that Tikopians may have obtained pigs or pig meat from Vanikoro in later prehistory. These sorts of questions are difficult to document using traditional archaeological techniques and are most suited to investigations using chronologically controlled analyses of commensal DNA or stable isotopes.
At present the bulk of commensal sampling appears to have occurred in the more easily accessed regions, such as Fiji and the Cook Islands. This is likely due to issues of convenience, affordability, and safety. In addition, samples for the study of commensal bacteria and viruses were often collected in opportune situations such as during the South Pacific Games where people from around the Pacific gathered (for example, see Yanagihara et al. Citation2002). These studies represent important first steps in developing a model based around specific organisms but it is clear that much work remains to be done. In some cases the gaps in plant and animal commensal studies can be filled by using specimens from more remote locales gathered for herbarium collections or during archaeological excavations. Greater collaboration among commensal researchers could result in targeted collecting expeditions to more remote regions to obtain samples of several species for use in multiple studies. Such an approach would not only increase the depth of current studies but would also broaden discussions and allow wider interpretations of the resulting genetic signatures of commensal species, both purposefully and accidentally transported.
PROSPECTS WITH NEW TECHNOLOGY
Projects such as Google Books and the Internet Archive (archive.org), among others, provide researchers with unparalleled access to historical documents that can help with examining both ethnographic accounts of human/animal interactions in the Pacific, and also records for European dispersals of plants and animals into and around the Pacific. In addition, free online translators are vastly improved from only five years ago and applications such as Google Translate provide easy access to information in several languages. This provides new opportunities for background research when constructing a commensal model, particularly with respect to understanding the origins, biology, translocation, effects of human choice, and historic transport on the organism of interest.
The development of next generation sequencing technologies means that researchers can start to target whole viral and mitochondrial genomes and specific nuclear genes, including those most likely to be the target of human selection. Identifying and targeting these genes, such as those that control yield, breeding, time to maturity, and important morphological traits, may provide unprecedented insights into the specific choices made by groups of humans in the selection of domesticated lineages of plants and animals. Questions about the tempo of evolutionary change, especially with reference to domesticated animals, will be enhanced by studies of ancient DNA as these add a necessary temporal component (Matisoo-Smith Citation2002; Ramakrishnan et al. Citation2005). Analysis of nuclear genes may also reveal how individual species have adapted to different island environments (e.g., high islands versus atolls). The identification of New Guinea as an independent center of domestication (Denham et al. Citation2003) suggests such studies are likely to have implications far beyond the Pacific. Commensal studies remain in their adolescence but the application of these models has grown considerably. The future for commensal studies is bright and will continue to provide an excellent avenue for the investigation of aspects of ancient human behavior, migration, interaction, and beyond.
ACKNOWLEDGEMENTS
The authors would like to thank Mike Carson for offering us the opportunity to articulate the basis for models centered on the analysis of the DNA of plants and animals to understand human mobility and to publish them for the first time in this issue. We are also grateful to Luc Baudouin and Gerard Ward for information and discussions related to coconuts. Last but not least, we very much appreciate the insightful comments and questions of three anonymous reviewers, which provided more fodder for discussion amongst the co-authors and in the final submission.
REFERENCES
- Agostini , H. , Yanagihara , R. , Davis , V. , Ryschkewitsch , C. F. and Stoner , G. L. 1997 . Asian genotypes of JC virus in native Americans and in a Pacific island population: Markers of viral evolution and human migration . Proceedings of the National Academy of Sciences , 94 : 14542 – 14546 .
- Anderson , E. 1952 . Plants, Man, and Life , Berkeley : University of California Press .
- Anderson-Carpenter , L. L. , McLachlan , J. S. , Jackson , S. T. , Kuch , M. , Lumibao , C. Y. and Poinar , H. N. 2011 . Ancient DNA from lake sediments: Bridging the gap between paleoecology and genetics . BMC Evolutionary Biology , 11 : 30
- Austin , C. C. 1999 . Lizards took express train to Polynesia . Nature , 397 : 113 – 114 .
- Austin , C. C. and Zug , G. R. 1999 . Molecular and morphological evolution in the south-central Pacific skink Emoia tongana (reptilia: Squamata): Uniformity and human-mediated dispersal . Australian Journal of Zoology , 47 : 425 – 437 .
- Baldwin , J. A. 1990 . “ Muruk, dok, pik, kakaruk: Prehistoric implication of geographical distributions in the southwest Pacific ” . In Pacific Production Systems: Approaches to Economic Prehistory , Edited by: Yen , D. E. and Mummery , J. M. J. 231 – 257 . Canberra : Department of Prehistory, Research School of Pacific Studies, The Australian National University .
- Barnes , S. S. , Matisoo-Smith , E. and Hunt , T. L. 2006 . Ancient DNA of the Pacific rat (Rattus exulans) from Rapa Nui (Easter Island) . Journal of Archaeological Science , 33 : 1536 – 1540 .
- Baudouin , L. and Lebrun , P. 2008 . Coconut (Cocos nucifera L.) DNA studies support the hypothesis of an ancient Austronesian migration from southeast Asia to America . Genetic Resources and Crop Development , 56 ( 2 ) : 257 – 262 .
- Bay-Petersen , J. 1984 . Competition for resources: The role of pig and dog in the Polynesian agricultural economy . Journale de la Societe des Oceanistes , 39 : 121 – 129 .
- Beaglehole , J. C. 1962 . The Endeavour Journal of Joseph Banks 1768–1771 , Sydney, New South Wales , , Australia : Angus & Robertson .
- Becker , C. 1997 . “ Zooarqueología y etnohistoria: Una contrastación en Isla Mocha ” . In La Isla de las Palabras Rotas , Edited by: Quiroz , D. and Sánchez , M. Santiago : Centro de Investigaciones Dego Barros Arana .
- Belaj , A. , Satovic , Z. , Cipriani , G. , Baldoni , L. , Testolin , R. , Rallo , L. and Trujillo , I. 2003 . Comparative study of the discriminating capacity of RAPD, AFLP and SSR markers and of their effectiveness in establishing genetic relationships in Olive . Theoretical and Applied Genetics , 107 ( 4 ) : 736 – 744 .
- Bensch , S. and Åkesson , M. 2005 . Ten years of AFLP in ecology and evolution: Why so few animals? . Molecular Ecology , 14 ( 10 ) : 2899 – 2914 .
- Bentley , A. R. 2006 . Strontium isotopes from the earth to the archaeological skeleton: A review . Journal of Archaeological Method and Theory , 13 ( 3 ) : 135 – 187 .
- Best , E. 1925 . Maori Agriculture: The cultivated food plants of the natives of New Zealand, with some account of native methods of agriculture, its ritual and origin myths , Wellington : Published under the direction of the Board of Maori Ethnological Research for the Dominion Museum by Whitcombe and Tombs .
- Bradley , D. G. 2006 . “ Documenting domestication: Reading animal genetic texts ” . In Documenting Domestication: New Genetic and Archaeological Paradigms , Edited by: Zeder , M.A. , Bradley , D. G. , Emshwiller , E. and Smith , B. D. 273 – 278 . Berkeley , CA : University of California Press .
- Brown , G. , Wassenaar , L. I. and Hobson , K. A. 2005 . Global application of stable hydrogen and oxygen isotopes to wildlife forensics . Oecologia , 143 : 337 – 348 .
- Brown , L. M. 2000 . Helicobacter pylori: Epidemiology and routes of transmission . Epidemiologic Reviews , 22 ( 2 ) : 283 – 297 .
- Bulmer , R. 1976 . “ Selectivity in hunting and in disposal of animal bone by the Kalam of the New Guinea Highlands ” . In Problems in Economic and Social Archaeology , Edited by: Sieveking , G. d. G. , Longworth , I. H. and Wilson , K. E. 169 – 186 . London : Duckworth .
- Bulmer , S. 2001 . “ Lapita dogs and singing dogs and the history of the dog in New Guinea ” . In The Archaeology of Lapita Dispersal in Oceania: Papers from the Fourth Lapita Conference , Edited by: Clark , G. , Anderson , A. and Vunidilo , T. 183 – 201 . Canberra : Pandanus Books .
- Burney , D. A. , James , H. F. , Pigott Burney , L. , Olson , S. L. , Kikuchi , W. , Wagner , W. L. Burney , M. 2001 . Fossil evidence for a diverse biota from Kaua’i and its transformation since human arrival . Ecological Monographs , 71 ( 4 ) : 615 – 641 .
- Caillon , S. , Quero-Garcia , J. , Lescure , J.-P. and Lebot , V. 2006 . Nature of taro (Colocasia esculenta (L.) Schott) genetic diversity prevalent in a Pacific Ocean island, Vanua Lava, Vanuatu . Genetic Resources and Crop Evolution , 53 : 1273 – 1289 .
- Carneiro-Proietti , A. B. F. , Catalan-Soares , B. C. , Castro-Costa , C. M. , Murphy , E. L. , Sabino , E. C. , Hisada , M. Galvão-Castro , B. 2006 . HTLV in the Americas: Challenges and perspectives . Pan American Journal of Public Health , 19 ( 1 ) : 44 – 53 .
- Cassar , O. , Capuano , C. , Bassot , S. , Charavay , F. , Duprez , R. , Afonso , P. V. Abel , M. 2007 . Human T Lymphotropic Virus type 1 subtype C Melanesian genetic variants of the Vanuatu archipelago and Solomon Islands share a common ancestor . Journal of Infectious Diseases , 196 ( 4 ) : 510 – 521 .
- Cheylan , G. , Granjon , L. and Britton-Davidian , J. 1998 . Distribution of genetic diversity within and between Western Mediterranean island population of the black rat Rattus rattus . Biological Journal of the Linnean Society , 63 : 393 – 408 .
- Clarke , A. C. 2009 . Origins and Dispersal of the Sweet Potato and Bottle Gourd in Oceania: Implications for prehistoric human mobility. Ph.D. Dissertation. Palmerston North , New Zealand : Massey University .
- Clarke , A. C. , Burtenshaw , M. K. , McLenachan , P. A. , Erickson , D. L. and Penny , D. 2006 . Reconstructing the origins and dispersal of the Polynesian bottle gourd (Lagenaria siceraria) . Molecular Biology and Evolution , 23 ( 5 ) : 893 – 900 .
- Clayton , D. A. 1991 . Replication and transcription of vertebrate mitochondrial DNA . Annual Review of Cell Biology , 7 : 453 – 478 .
- Coil , J. and Kirch , P. V. 2005 . “ An Ipomoean landscape: Archaeology and the sweet potato in Kahikinui, Maui, Hawaiian islands ” . In The Sweet Potato in Oceania: A Reappraisal , Edited by: Ballard , C. , Brown , P. , Bourke , R. M. and Harwood , T. 71 – 84 . Sydney , , Australia : Centatime .
- Collins , P. W. 1991 . Interaction betwen island foxes (Urocyon littoralis) and Indians on island off the coast of Southern California: I. Morphological and archaeological evidence of human assisted dispersal . Journal of Ethnobiology , 11 : 51 – 81 .
- Czarnecki , J. , Friedlaender , J. S. and Stoner , G. 2007 . “ Viral phylogeny and human migration in the southwest Pacific ” . In Genes, Language and Culture History in the Southwest Pacific , Edited by: Friedlaender , J. S. 171 – 180 . Oxford : Oxford University Press .
- Dancause , K. N. , Vilar , M. G. , Steffy , R. and Lum , J. K. 2011 . Characterizing genetic diversity of contemporary Pacific chickens using mitochondrial DNA analyses . PLoS ONE , 6 ( 2 ) : e16843
- Davidson , J. M. 1969 . “ Excavation of a coastal midden deposit, SU-LO-1 ” . In Archaeology in Western Samoa , Edited by: Green , R. C. and Davidson , J. 224 – 252 . Auckland : Auckland Institute and Museum .
- de France , S. D. , Keegan , W. F. and Newsom , L. A. 1996 . “ The archaeobotanical, bone isotope, and zooarchaeological records from Carribean sites in comparative perspective ” . In Case Studies in Environmental Archaeology , Edited by: Reitz , E. J. , Newson , L. A. and Scudder , S. J. 289 – 304 . New York : Plenum Press .
- Denham , T. , Haberle , S. G. , Lentfer , C. , Fullagar , R. , Field , J. , Therin , M. , Porch , N. and Winsborough , B. 2003 . Origins of agriculture at Kuk swamp in the highlands of New Guinea . Science , 301 : 189 – 193 .
- Dennis , J. V. and Gunn , C. R. 1971 . Case against trans-Pacific dispersal of the coconut by ocean currents . Economic Botany , 25 ( 4 ) : 407 – 413 .
- Elbaum , R. , Melamed-Bessudo , C. , Tuross , N. , Levy , A. A. and Weiner , S. 2009 . New methods to isolate organic materials from silicified phytoliths reveal fragmented glycoproteins but no DNA . Quaternary International , 193 : 11 – 19 .
- Fraser , M. , Sten , S. and Götherström , A. 2012 . Neolithic Hedgehogs (Erinaceus europaeus) from the Island of Gotland show early contacts with the Swedish mainland . Journal of Archaeological Science , 39 ( 2 ) : 229 – 233 .
- Gessain , A. , Pecon-Slattery , J. , Meertens , L. and Mahieux , R. 2000 . Origins of HTLV-1 in South America . Nature Medicine , 6 : 232
- Giovas , C. M. 2006 . No pig atoll: Island biogeography and the extirpation of a Polynesian domesticate . Asian Perspectives , 45 ( 1 ) : 69 – 95 .
- Giovas , C. M. , LeFebvre , M. J. and Fitzpatrick , S. M. 2012 . New records for prehistoric introduction of Neotropical mammals to the West Indies: Evidence from Carriacou, Lesser Antilles . Journal of Biogeography , 39 ( 3 ) : 476 – 487 .
- Giuffra , E. , Kijas , J. M. H. , Amarger , V. , Carlborg , O. , Jeon , J. T. and Andersson , L. 2000 . The origin of the domestic pig: Independent domestication and subsequent introgression . Genetics , 154 ( 4 ) : 1785 – 1791 .
- Glover , I. and Bellwood , P. S. 2004 . Southeast Asia: From prehistory to history , New York : RoutledgeCurzon .
- Gongora , J. , Fleming , P. , Spencer , P. B. S. , Mason , R. , Garkavenko , O. , Meyer , J.-N. , Droegemueller , C. , Lee , J. H. and Moran , C. 2004 . Phylogenetic relationships of Australian and New Zealand feral pigs assessed by mitochondrial control region sequence and nuclear GPIP genotype . Molecular Phylogenetics and Evolution , 33 ( 2 ) : 339 – 348 .
- Green , R. C. 2000 . A range of disciplines support a dual origin for the bottle gourd in the Pacific . Journal of the Polynesian Society , 109 : 191 – 197 .
- Green , R. C. 2005 . “ Sweet potato transfers in Polynesian prehistory ” . In The Sweet Potato in Oceania: A Reappraisal , Edited by: Ballard , C. , Brown , P. , Bourke , R. M. and Harwood , T. 43 – 62 . Sydney , , Australia : Centatime .
- Gunn , B. F. , Baudouin , L. and Olsen , K. M. 2011 . Independent origins of cultivated coconut (Cocus nucifera L.) in the Old World tropics . PLoS ONE , 6 ( 6 ) : e21143
- Hagelberg , E. and Clegg , J. B. 1993 . Genetic polymorphisms in prehistoric Pacific islanders determined by analysis of ancient bone DNA . Proceedings of the Royal Society of London B , 252 : 163 – 170 .
- Haile , J. , Holdaway , R. , Oliver , K. , Bunce , M. , Gilbert , M. T. P. , Nielsen , R. , Munch , K. , Ho , S. Y. W. , Shapiro , B. and Willerslev , E. 2007 . Ancient DNA chronology within sediment deposits: Are paleobiological reconstructions possible and is DNA leaching a factor? . Molecular Biology and Evolution , 24 ( 4 ) : 982 – 989 .
- Hamilton , A. M. , Zug , G. R. and Austin , C. C. 2010 . Biogeographic anomaly or human introduction: A cryptogenic population of tree skink (reptilia: Squamata) from the Cook Islands, Oceania . Biological Journal of the Linnean Society , 100 ( 2 ) : 318 – 328 .
- Harris , G. , Burtenshaw , M. , Davidson , J. and Leach , B. F. 2000 . An experimental kumara garden at Robin Hood Bay, near Port Underwood, Marlborough . Archaeology in New Zealand , 43 : 301 – 313 .
- Hather , J. and Kirch , P. V. 1991 . Prehistoric sweet potato (Ipomoea batatas) from Mangaia island, central Polynesia . Antiquity , 65 : 887 – 893 .
- Hinkle , A. E. 2004 . The distribution of a male sterile form of ti (Cordyline fruticosa) in Polynesia: A case of human selection? . Journal of the Polynesian Society , 113 ( 3 ) : 263 – 290 .
- Hinkle , A. E. 2007 . Population structure of Pacific Cordyline fruticosa (laxmanniaceae) with implications for human settlement of Polynesia . American Journal of Botany , 94 ( 5 ) : 828 – 839 .
- Ho , S. Y. W. and Gilbert , M. T. P. 2010 . Ancient mitogenomics . Mitochondrion , 10 ( 1 ) : 1 – 11 .
- Horai , S. , Hayasaka , K. , Kondo , R. , Tsungane , K. and Takahata , N. 1995 . Recent African origin of modern humans revealed by complete sequences of hominoid mitochondrial DNAs . Proceedings of the National Academy of Sciences , 92 : 532 – 536 .
- Intoh , M. 1992 . Why were pots imported to Ngulu atoll? A consideration of subsistence strategy . Journal of the Polynesian Sociery , 101 ( 2 ) : 159 – 168 .
- Irwin , G. J. 2004 . Kohika: The archaeology of a late Maori lake village in the Ngati Awa Rohe, Bay of Plenty, New Zealand , Auckland : Auckland University Press .
- Irwin , S. V. , Kaufusi , P. , Banks , K. , de la Peña , R. and Cho , J. J. 1998 . Molecular characterization of taro (Colocasia esculenta) using RAPD markers . Euphytica , 99 : 183 – 189 .
- Jobes , D. V. , Friedlaender , J. S. , Mgone , C. S. , Agostini , H. T. , Koki , G. , Yanagihara , R. , Ng , T. C. N. , Chima , S. C. , Ryschkewitsch , C. F. and Stoner , G. L. 2001 . New JC virus (JCV) genotypes from Papua New Guinea and Micronesia (type 8 and type 2e) and evolutionary analysis of 32 complete JCV genomes . Archives of Virology , 146 : 2097 – 2113 .
- Jobling , M. A. 2012 . The impact of recent events on human genetic diversity . Philosophical Transactions of the Royal Society of London B , 367 : 793 – 799 .
- Jobling , M. A. , Hurles , M. E. and Tyler-Smith , C. 2004 . Human Evolutionary Genetics: Origins, Peoples and Disease , New York : Garland Science .
- Jokhan , A. D. and McLenachan , P. 2004 . Genetic fingerprinting of Fijian kava . South Pacific Journal of Natural Sciences , 22 ( 1 ) : 41 – 43 .
- Kirch , P. V. 1986 . “ Exchange systems and inter-island contact in the transformation of an island society: The Tikopia case ” . In Island Societies: Archaeological Approaches to Evolution and Transformation , Edited by: Kirch , P. V. 33 – 41 . Cambridge : Cambridge University Press .
- Kirch , P. V. 1997 . The Lapita Peoples: Ancestors of the Oceanic world , Cambridge, , MA : Blackwell .
- Kirch , P. V. 2000a . On the Road of the Winds: An Archaeological History of the Pacific Islands Before European Contact , Berkeley , CA : University of California Press .
- Kirch , P. V. 2000b . “ Pigs, humans, and trophic competition on small Oceanic islands ” . In Australian Archaeologist: Collected Papers in Honour of Jim Allen , Edited by: Anderson , A. and Murray , T. 427 – 439 . Canberra : Coombs Academic Publishing, Australian National University .
- Kirch , P. V. 2001 . Lapita and its Transformations in Near Oceania: Archaeological Investigations in the Mussau Islands, Papua New Guinea, 1985–88 , Berkeley , CA : Archaeological Research Facility University of California .
- Kirch , P. V. and Hunt , T. L. 1988 . The Talepakemalai site and Oceanic prehistory . National Geographic Research , 4 : 328 – 342 .
- Kirch , P. V. , Hunt , T. L. , Weisler , M. , Butler , V. and Allen , M. S. 1991 . “ Mussau islands prehistory: Results of the 1985–86 excavations ” . In Report of the Lapita Homeland Project , Edited by: Allen , J. and Gosden , C. 144 – 163 . Canberra : Department of Prehistory, Research School of Pacific Studies, Australian National University .
- Kreike , C. M. , Van Eck , H. J. and Lebot , V. 2004 . Genetic diversity of taro, Colocasia esculenta (L.) Schott, in Southeast Asia and the Pacific . Theoretical and Applied Genetics , 109 : 761 – 768 .
- Ladefoged , T. N. , Graves , M. W. and Coil , J. H. 2005 . The introduction of sweet potato in Polynesia: Early remains in Hawai‘i . Journal of the Polynesian Society , 114 ( 4 ) : 359 – 373 .
- Ladefoged , T. N. , Kirch , P. V. , Gon Ill , S. O. , Chadwick , O. A. , Hartshorn , A. S. and Vitousek , P. M. 2009 . Opportunities and constraints for intensive agriculture in the Hawaiian archipelago prior to European contact . Journal of Archaeological Science , 36 : 2374 – 2383 .
- Ladefoged , T. N. , McCoy , M. D. , Asner , G. P. , Kirch , P. V. , Puleston , C. O. , Chadwick , O. A. and Vitousek , P. M. 2011 . Agricultural potential and actualized development in Hawai‘i: An airborne LiDAR survey of the leeward Kohala field system (Hawai’i island) . Journal of Archaeological Science , 38 : 3605 – 3619 .
- Larson , G. , Cucchi , T. , Fujita , M. , Matisoo-Smith , E. , Robins , J. , Anderson , A. Rolett , B. 2007 . Phylogeny and ancient DNA of Sus provides insights into Neolithic expansion in Island Southeast Asia and Oceania . Proceedings of the National Academy of Sciences , 104 ( 12 ) : 4834 – 4839 .
- Larson , G. , Dobney , K. , Albarella , U. , Fang , M. Y. , Matisoo-Smith , E. , Robins , J. Lowden , S. 2005 . Worldwide phylogeography of wild boar reveals multiple centers of pig domestication . Science , 307 ( 5715 ) : 1618 – 1621 .
- Lebot , V. , McKenna , D. , Johnston , E. , Zheng , Q. and McKern , D. 1999 . Morphological, phytochemical, and genetic variation in Hawaiian cultivars of ‘awa (kava, Piper methysticum, piperaceae) . Economic Botany , 53 ( 4 ) : 407 – 418 .
- Lebot , V. , Prana , M. S. , Kreike , N. , van Heck , H. , Pardales , J. , Okpul , T. Gendua , T. 2004 . Characterisation of taro (Colocasia esculenta (L.) Schott) genetic resources in Southeast Asia and Oceania . Genetic Resources and Crop Evolution , 51 : 381 – 392 .
- Lee , T. , Burch , J. B. , Coote , T. , Fontaine , B. , Gargominy , O. , Pearce-Kelly , P. and Foighil , D. O. 2007 . Prehistoric inter-archipelago trading of Polynesian tree snails leaves a conservation legacy . Proceedings of the Royal Society B—Biological Sciences , 274 : 2907 – 2914 .
- Lepofsky , D. 1988 . “ The environmental context of Lapita settlement locations ” . In Archaeology of the Lapita Cultural Complex: A Critical Review , Edited by: Kirch , P. V. and Hunt , T. L. 33 – 48 . Seattle : Burke Museum .
- Lepofsky , D. , Harries , H. C. and Kellum , M. 1992 . Early coconuts on Mo‘orea island, French Polynesia . Journal of the Polynesian Society , 101 : 299 – 308 .
- Lum , J. K. and Cann , R. L. 2000 . MtDNA lineage analyses: Origins and migrations of Micronesians and Polynesians . American Journal of Physical Anthropology , 113 ( 2 ) : 151 – 168 .
- Lum , J. K. , Kaneko , A. , Tanabe , K. , Takahashi , N. , Bjorkman , A. and Kobayakawa , T . 2004 . Malaria dispersal among islands: Human mediated Plasmodium falciparum gene flow in Vanuatu, Melanesia . Acta Tropica , 90 ( 2 ) : 181 – 185 .
- Lum , J. K. , McIntyre , J. K. , Greger , D. L. , Huffman , K. W. and Vilar , M. G. 2006 . Recent Southeast Asian domestication and Lapita dispersal of sacred male pseudohermaphroditic “tuskers” and hairless pigs of Vanuatu . Proceedings of the National Academy of Sciences , 103 ( 46 ) : 17190 – 17195 .
- Mace , E. S. and Godwin , I. D. 2002 . Development and characterization of polymorphic microsatellite markers in taro (Colocasia esculenta) . Genome , 45 : 823 – 832 .
- Mace , E. S. , Mathur , P. N. , Izquierdo , L. , Hunter , D. , Taylor , M. B. , Singh , D. , DeLacy , I. H. , Jackson , G. V. H. and Godwin , I. D. 2006 . Rationalization of taro germplasm collections in the Pacific Island region using simple sequence repeat (SSR) markers . Plant Genetic Resources , 4 : 210 – 220 .
- Martin , E. and Hine , R. 2008 . A Dictionary of Biology , Oxford : Oxford University Press . Oxford Reference Online, http://www.oxfordreference.com/views/ENTRY.html?subview=Main&entry=t6.e961(accessed August 27, 2012, University of New England Subscription)
- Matisoo-Smith , E. 1994 . The human colonisation of Polynesia—A novel approach: Genetic analyses of the Polynesian rat (Rattus exulans) . Journal of the Polynesian Society , 103 : 75 – 87 .
- Matisoo-Smith , E. 1996 . No hea te kiore: MtDNA variation in Rattus exulans—A model for human colonisation and contact in prehistoric Polynesia , Auckland : University of Auckland . Ph.D. Dissertation
- Matisoo-Smith , E. 2002 . Something old, something new: Do genetic studies of contemporary populations reliably represent prehistoric populations of Pacific Rattus exulans? . Human Biology , 74 ( 3 ) : 489 – 496 .
- Matisoo-Smith , E. 2007a . “ Animal translocations, genetic variation and the human settlement of the Pacific ” . In Genes, Language and Culture History in the Southwest Pacific , Edited by: Friedlaender , J. S. 157 – 170 . Oxford : Oxford University Press .
- Matisoo-Smith , E. 2007b . “ The peopling of Oceania ” . In Anthropological Genetics: Theory, Methods and Applications , Edited by: Crawford , M. H. 409 – 432 . Cambridge : Cambridge University Press. .
- Matisoo-Smith , E. 2009 . The commensal model for human settlement of the Pacific 10 years on—what can we say and where to now? . Journal of Island and Coastal Archaeology , 4 : 151 – 163 .
- Matisoo-Smith , E. , Allen , J. S. , Ladefoged , T. N. , Roberts , R. M. and Lambert , D. M. 1997 . Ancient DNA from Polynesian rats: Extraction, amplification and sequence from single small bones . Electrophoresis , 18 ( 9 ) : 1534 – 1537 .
- Matisoo-Smith , E. , Hingston , M. , Summerhayes , G. , Robins , J. , Ross , H. and Hendy , M. 2009 . On the rat trail in Near Oceania . Pacific Science , 63 ( 4 ) : 465 – 475 .
- Matisoo-Smith , E. , Horsburgh , A. , Robins , J. and Anderson , A. 2001 . Genetic variation in archaeological Rattus exulans remains from the Emily Bay settlement site, Norfolk Island . Records of the Australian Museum , 27 : 81 – 84 .
- Matisoo-Smith , E. , Roberts , R. M. , Irwin , G. J. , Allen , J. S. , Penny , D. and Lambert , D. M. 1998 . Patterns of prehistoric human mobility in Polynesia indicated by mtDNA from the Pacific rat . Proceedings of the National Academy of Sciences , 95 ( 25 ) : 15145 – 15150 .
- Matisoo-Smith , E. and Robins , J. H. 2004 . Origins and dispersals of Pacific peoples: Evidence from mtDNA phylogenies of the Pacific rat . Proceedings of the National Academy of Sciences , 101 ( 24 ) : 9167 – 9172 .
- Matisoo-Smith , E. , Sutton , D. G. , Ladefoged , T. N. , Lambert , D. M. and Allen , J. S. 1999 . Prehistoric mobility in Polynesia: MtDNA variation in Rattus exulans from the Chatham and Kermadec Islands . Asian Perspectives , 38 ( 2 ) : 186 – 199 .
- Matthews , P. J. 1995 . Aroids and the Austronesians . Tropics , 4 ( 2 ) : 105 – 126 .
- McCoy , M. D. , Graves , M. W. and Murakami , G. 2010 . Introduction of breadfruit (Artocarpus altilis) to the Hawaiian Islands . Economic Botany , 64 ( 4 ) : 374 – 381 .
- McCoy , P. C. and Cleghorn , P. C. 1988 . Archaeological excavations in Santa Cruz (Nendo), southeast Solomon Islands: Summary report . Archaeology in Oceania , 23 ( 3 ) : 104 – 115 .
- McNab , R. 1914 . Historical Records of New Zealand , Wellington , , New Zealand : Government Printer .
- Meudt , H. M. and Clarke , A. C. 2007 . Almost forgotten or latest practice? AFLP applications, analyses and advances . Trends in Plant Science , 12 ( 3 ) : 106 – 117 .
- Mills , C. 2004 . Dogs of Rarotonga . Discover , 25 ( 6 ) : 70 – 77 .
- Moodley , Y. , Linz , B. , Yamaoka , Y. , Windsor , H. M. , Breurec , S. , Wu , J.-Y Maady , A. 2009 . The peopling of the Pacific from a bacterial perspective . Science , 323 ( 5913 ) : 527 – 530 .
- Murray-McIntosh , R. P. , Scrimshaw , B. J. , Hatfield , P. J. and Penny , D. 1998 . Testing migration patterns and estimating founding population size in Polynesia by using human mtDNA sequences . Proceedings of the National Academy of Sciences , 95 ( 15 ) : 9047 – 9052 .
- Nabholz , B. , Glemin , S. and Galtier , N. 2009 . The erratic mitochondrial clock: Variations of mutation rate, not population size, affect mtDNA diversity across birds and mammals . BMC Evolutionary Biology , 9 ( 1 ) : 54
- Nagaoka , L. 1988 . “ Lapita subsistence: The evidence of non-fish archaeofaunal remains ” . In Archaeology of the Lapita Cultural Complex: A Critical Review , Edited by: Kirch , P. V. and Hunt , T. L. 117 – 134 . Seattle : Burke Museum .
- Nei , M. 1987 . Molecular Evolutionary Genetics , New York : Columbia University Press .
- Ohkura , S. , Yamashita , M. , Cartier , L. , Gutierrez Tanabe , D. , Hayami , M. , Sonoda , S. and Tajima , K. 1999 . Identification and phylogenetic characterization of a human T-cell leukaemia virus type I isolate from a native inhabitant (Rapa Nui) of Easter Island . Journal of General Virology , 80 : 1995 – 2001 .
- Olmstead , R. G. and Palmer , S. A. 1994 . Chloroplast DNA systematics: A review of methods and data analysis . American Journal of Botany , 81 : 1205 – 1224 .
- O’Reilly , D. J. W. 2007 . Early Civilizations of Southeast Asia , Lanham , MD : AltaMira Press .
- Orliac , C. and Orliac , M. 1998 . “ The disappearance of Easter Island's forest: Over-exploitation or climatic catastrophe? ” . In Easter Island in Pacific Context. South Seas Symposium: Proceedings of the Fourth International Conference on Easter Island and East Polynesia , Edited by: Stevenson , C. M. , Lee , G. and Morin , F. J. 129 – 134 . Los Osos , CA : Bearsville and Cloud Mountain Presses, Easter Island Foundation .
- Oskarsson , M. C. R. , Klütsch , C. F. C. , Boonyaprakob , U. , Wilton , A. , Tanabe , Y. and Savolainen , P. 2011 . Mitochondrial DNA data indicate an introduction through mainland Southeast Asia for Australian dingoes and Polynesian domestic dogs . Proceedings of the Royal Society London B , 279 ( 1730 ) : 967 – 974 .
- Pakendorf , B. , Novgorodov , I. N. , Osakovskij , V. L. , Danilova , A. P. , Protod’jakonov , A. P. and Stoneking , M. 2006 . Investigating the effects of prehistoric migrations in Siberia: Genetic variation and the origins of Yakuts . Human Genetics , 120 : 334 – 353 .
- Pakendorf , B. and Stoneking , M. 2005 . Mitochondrial DNA and human evolution . Annual Review of Genomics and Human Genetics , 6 : 165 – 183 .
- Palmer , S. A. , Smith , O. and Allaby , R. G. 2012 . The blossoming of plant archaeogenetics . Annals of Anatomy , 194 : 146 – 156 .
- Parducci , L. , Suyama , Y. , Lascoux , M. and Bennett , K. D. 2005 . Ancient DNA from pollen: A genetic record of population history in Scots pine . Molecular Ecology , 14 : 2873 – 2882 .
- Pavesi , A. 2004 . Detecting traces of prehistoric human migrations by genographic synthetic maps of Polyomavirus JC . Journal of Molecular Evolution , 58 : 304 – 313 .
- Perrier , X. , De Langhe , E. , Donohue , M. , Lentfer , C. , Vrydaghs , L. , Bakry , F. Carreel , F. 2011 . Multidisciplinary perspectives on banana (Musa spp.) domestication . Proceedings of the National Academy of Sciences , 108 ( 28 ) : 11311 – 11318 .
- Ragone , D. 1991 . “ Ethnobotany of breadfruit in Polynesia ” . In Islands, Plants, and Polynesians: An Introduction to Polynesian Ethnobotany , Edited by: Cox , P. A. and Banack , S. A. 203 – 220 . Portland : Dioscorides Press .
- Ramakrishnan , U. , Hadly , E. A. and Mountain , J. L. 2005 . Detecting past population bottlenecks using temporal genetic data . Molecular Ecology , 14 ( 10 ) : 2915 – 2922 .
- Reese , D. S. 1996 . Pleistocene and Holocene Fauna of Crete and its First Settlers , Madison , WI : Monographs in World Archaeology No. 28 .
- Reitz , E. J. and Wing , E. S. 1999 . Zooarchaeology , New York : Cambridge University Press .
- Rezza , G. , Danaya , R. T. , Wagner , T. M. , Sarmati , L. , Owen , I. L. , Monin , P. , Andreoni , M. , Suligoi , B. , Ensoli , B. and Pozio , E. 2001 . Human Herpesvirus-8 and other viral infections, Papua New Guinea . Emerging Infectious Diseases , 7 ( 5 ) : 893 – 895 .
- Richards , M. P. , Fuller , B. T. , Sponheimer , M. , Robinson , T. and Ayliffe , L. 2003 . Sulphur isotopes in palaeodietary studies: A review and results from a controlled feeding experiment . International Journal of Osteoarchaeology , 13 ( 1–2 ) : 37 – 45 .
- Rick , T. C. , Erlandson , J. M. , Vellanoweth , R. L. , Braje , T. J. , Collins , P. W. , Guthrie , D. A. and Stafford , T. W. 2009 . Origins and antiquity of the island fox (Urocyon littoralis) on California's Channel Islands . Quaternary Research , 71 ( 2 ) : 93 – 98 .
- Rick , T. C. , Walker , P. L. , Willis , L. M. , Noah , A. C. , Erlandson , J. M. , Vallanoweth , R. L. , Braje , T. J. and Kennett , D. J. 2008 . Dogs, humans and island ecosystems: The distribution, antiquity and ecology of domestic dogs (Canis familiaris) on California's Channel Islands, USA . The Holocene , 18 ( 7 ) : 1077 – 1087 .
- Robins , J. H. , Hingston , M. , Matisoo-Smith , E. and Ross , H. A. 2007 . Identifying Rattus species using mitochondrial DNA . Molecular Ecology Notes , 7 : 717 – 729 .
- Robins , J. H. , Matisoo-Smith , E. and Furey , L. 2001 . “ Hit or miss? Factors affecting DNA preservation in Pacific archaeological material ” . In Australasian Connections and New Directions: Proceedings of the 7th Australasian Archaeometry Conference , Edited by: Jones , M. and Sheppard , P. J. 295 – 305 . Auckland : The University of Auckland .
- Robins , J. H. , Matisoo-Smith , E. and Ross , H. A. 2003 . The origins of the feral pigs on the Auckland Islands . Journal of The Royal Society of New Zealand , 33 ( 2 ) : 561 – 569 .
- Robins , J. H. , Ross , H. A. , Allen , M. S. and Matisoo-Smith , E. 2006 . Sus bucculentus revisited . Nature , 440 : E7
- Ryschkewitsch , C. F. , Friedlaender , J. S. , Mgone , C. S. , Jobes , D. V. , Agostini , H. T. , Chima , S. C. , Alpers , M. P. , Koki , G. , Yanagihara , R. and Stoner , G. L. 2000 . Human polyomavirus JC variants in Papua New Guinea and Guam reflect ancient population settlement and viral evolution . Microbes and Infection , 2 : 987 – 996 .
- Sardos , J. , Noyer , J.-L. , Malapa , R. , Bouchet , S. and Lebot , V. 2012 . Genetic diversity of taro (Colocasia esculenta (L.) Schott) in Vanuatu (Oceania): An appraisal of the distribution of allelic diversity (DAD) with SSR markers . Genetic Resources and Crop Evolution , 59 : 805 – 820 .
- Satoh , M. , Kaneko , A. , Kokaze , A. , Katano , H. and Sata , T. 2006 . Seroprevalence of human Herpesvirus 8 in the Vanuatu islands in eastern Melanesia . Japanese Journal of Infectious Disease , 59 : 63 – 65 .
- Savolainen , P. , Leitner , T. , Wilton , A. N. , Matisoo-Smith , E. and Lundeberg , J. 2004 . A detailed picture of the origin of the Australian dingo, obtained from the study of mitochondrial DNA . Proceedings of the National Academy of Sciences , 101 ( 33 ) : 12387 – 12390 .
- Schlötterer , C. 2004 . The evolution of molecular markers—just a matter of fashion? . Nature Reviews Genetics , 5 ( 1 ) : 63 – 69 .
- Schwarcz , H. P. 2002 . “ Tracing human migration with stable isotopes ” . In Human Mate Choice and Prehistoric Marital Networks , Edited by: Kenichi , A. and Takeru , A. 183 – 201 . Tokyo : International Research Center for Japanese Studies .
- Seelenfreund , D. , Piña , R. , Ho , K.-Y. , Lobos , S. , Moncada , X. and Seelenfreund , A. 2011 . Molecular analysis of Broussonetia papyrifera (L.) vent. (magnoliophyta: Urticales) from the Pacific, based on ribosomal sequences of nuclear DNA . New Zealand Journal of Botany , 49 : 413 – 420 .
- Singh , D. , Mace , E. S. , Godwin , I. D. , Mathur , P. N. , Okpul , T. , Taylor , M. , Hunter , D. , Kambuou , R. , Ramanatha Rao , V. and Jackson , G. 2008 . Assessment and rationalization of genetic diversity of Papua New Guinea taro (Colocasia esculenta) using SSR DNA fingerprinting . Genetic Resources and Crop Evolution , 55 : 811 – 822 .
- Sinoto , Y. H. and McCoy , P. C. 1975 . Report on the preliminary excavation of an early habitation site on Huahine, Society Islands . Journal de la Societe des Oceanistes , 47 : 143 – 186 .
- Skjølsvold , A. 1961 . “ Site E-2, a circular stone dwelling, Anakena ” . In Archaeology of Easter Island , Edited by: Heyerdahl , T. and Ferdon , E. N. 295 – 303 . Santa Fe : Monograph of the School of American Research and the Museum of New Mexico .
- Spennemann , D. R. 1990 . “ The role of pigs and dogs in the taphonomy of modern and archaeological assemblages from Tonga ” . In Problem Solving in Taphonomy: Archaeological and Palaeontological Studies from Europe, Africa and Oceania , Edited by: Solomon , S. , Davidson , I. and Watson , D. 101 – 105 . St. Lucia, Qld : Anthropology Museum, University of Queensland .
- Spriggs , M. 1984 . Early coconut remains from the South Pacific . Journal of the Polynesian Society , 93 : 71 – 76 .
- Steadman , D. W. 2006 . Extinction and Biogeography of Tropical Pacific Birds , Chicago : University of Chicago Press .
- Storey , A. A. , Athens , J. S. , Bryant , D. , Carson , M. , Emery , K. , deFrance , S. Higham , C. 2012 . Investigating the global dispersal of chickens in prehistory using ancient mitochondrial DNA signatures . PLoS ONE , 7 ( 7 ) : e39171
- Storey , A. A. , Ladefoged , T. N. and Matisoo-Smith , E. A. 2008 . Counting your chickens: Density and distribution of chicken remains in archaeological sites of Oceania . International Journal of Osteoarchaeology , 18 ( 3 ) : 240 – 261 .
- Storey , A. A. , Ramirez , J. M. , Quiroz , D. , Burley , D. V. , Addison , D. J. , Walter , R. Anderson , A. J. 2007 . Radiocarbon and DNA evidence for a pre-Columbian introduction of Polynesian chickens to Chile . Proceedings of the National Academy of Sciences , 104 ( 25 ) : 10335 – 10339 .
- Storey , A. A. , Spriggs , M. , Bedford , S. , Hawkins , S. C. , Robins , J. H. , Huynen , L. and Matisoo-Smith , E. 2010 . Mitochondrial DNA from 3000-year old chickens at the Teouma site, Vanuatu . Journal of Archaeological Science , 37 ( 10 ) : 2459 – 2468 .
- Summerhayes , G. , Leavesley , M. G. , Fairbairn , A. , Mandui , H. , Field , J. , Ford , A. and Fullagar , R. 2010 . Human adaptation and plant use in Highland New Guinea 49,000 to 44,000 years ago . Science , 330 : 78 – 81 .
- Takao , S. , Ishida , T. , Bhatia , K. , Saha , N. , Soemantri , A. and Kayame , O. W. 2000 . Seroprevalence of human T-lymphotropic virus type 1 in Papua New Guinea and Irian Jaya measured using different western blot criteria . Journal of Clinical Virology , 16 ( 2 ) : 129 – 133 .
- Takasaka , T. , Miranda , J. J. , Sugimoto , C. , Paraguison , R. , Zheng , H. Y. , Kitamura , T. and Yogo , Y. 2004 . Genotypes of JC virus in Southeast Asia and the western Pacific: Implications for human migrations from Asia to the Pacific . Anthropological Science , 112 ( 1 ) : 53 – 59 .
- Takayama , J. and Intoh , M. 1978 . Archaeological Excavation at Chukienu Shell Midden on Tol, Truk , Japan : Tezukayama University . Reports of Pacific Archaeological Survey, No. 5. Nara City
- Tapio , M. and Grigaliunaite , I. 2003 . Use of mitochondrial DNA as a genetic marker in domesticated mammals . Ekologija , 1 : 31 – 33 .
- Thode , H. G. 1991 . “ Sulphur isotopes in nature and the environment: An overview ” . In Stable Isotopes in the Assessment of Natural and Anthropogenic Sulphur in the Enrvironment , Edited by: Krouse , H. R. and Grinenko , V. A. 1 – 26 . New York : John Wiley & Sons Ltd .
- Threadgold , J. and Brown , T. A. 2003 . Degradation of DNA in artificially charred wheat seed . Journal of Archaeological Science , 30 : 1067 – 1076 .
- Tinghitella , R. M. , Zuk , M. , Beveridge , M. and Simmons , L. W. 2011 . Island hopping introduces Polynesian field crickets to novel environments, genetic bottlenecks and rapid evolution . Journal of Evolutionary Biology , 24 ( 6 ) : 1199 – 1211 .
- Titcomb , M. and Pukui , M. K. 1969 . Dog and Man in the Ancient Pacific, with Special Attention to Hawai‘i , Honolulu : Star-Bulletin Print Co .
- Trewick , S. A. and Gibb , G. C. 2010 . Vicars, tramps and assembly of the New Zealand avifauna: A review of molecular phylogenetic evidence . Ibis , 152 : 226 – 253 .
- Trotter , M. M. 1979 . Niue Island Archaeological Survey , Christchurch : Canterbury Museum .
- Vigne , J. D. and Valladas , H. 1996 . Small mammal fossil assemblages as indicators of enfirnomental change in Northern Corsica during the last 2500 years . Journal of Archaeological Science , 23 : 199 – 215 .
- Vos , P. , Hogers , R. , Bleeker , M. , Reijans , M. , van de Lee , T. , Hornes , M. Frijters , A. 1995 . AFLP: A new technique for DNA fingerprinting . Nucleic Acids Research , 23 ( 21 ) : 4407 – 4414 .
- Waiblinger , H.-U. , Schillinger , M. and Hess , N. 2012 . In-house and interlaboratory validation of a method for the extraction of DNA from maize starch . Journal für Verbraucherschutz und Lebensmittelsicherheit , 7 : 163 – 170 .
- Wallace , A. R. 1869 . The Malay Archipelago: The Land of the Orang-utan, and the Bird of Paradise. A Narrative of Travel, with Studies of Man and Nature , London : Macmillan and Co . Vol. II, 2nd ed
- Ward , G. and Brookfield , M. 1992 . The dispersal of the coconut: Did it float or was it carried to Panama? . Journal of Biogeography , 19 : 467 – 480 .
- Watts , K. J. , Thompson , C. H. , Cossart , Y. E. and Rose , B. R. 2002 . Sequence variation and physical state of human Papilloma Virus type 16 cervical cancer isolates from Australia and New Caledonia . International Journal of Cancer , 97 : 868 – 874 .
- Welsh , J. and McClelland , M. 1990 . Fingerprinting genomes using PCR with arbitrary primers . Nucleic Acids Research , 18 ( 24 ) : 7213 – 7218 .
- Whistler , W. A. 1991 . “ Polynesian plant introductions ” . In Islands, Plants, and Polynesians: An Introduction to Polynesian Ethnobotany , Edited by: Cox , P. A. and Banack , S. A. 41 – 66 . Portland : Dioscorides Press .
- White , J. P. 2001 . “ Fauna: More than just food ” . In Pacific 2000: Proceedings of the Fifth International Conference on Easter Island and the Pacific , Edited by: Stevenson , C. M. , Lee , G. and Morin , F. J. 37 – 40 . Los Osos : Bearsville Press .
- White , J. P. 2004 . “ Where the wild things are: Prehistoric aminal translocation in the circum New Guinea Archipelago ” . In Voyages of Discovery: The Archaeology of Islands , Edited by: Ftizpatrick , S. M. 147 – 164 . Westport : Praeger .
- Williams , J. G. K. , Kubelik , A. R. , Livak , K. J. , Rafalski , J. A. and Tingey , S. V. 1990 . DNA polymorphisms amplified by arbitrary primers are useful as genetic markers . Nucleic Acids Research , 18 ( 22 ) : 6531 – 6535 .
- Wing , E. S. 1993 . “ The realm between wild and doemstic ” . In Skeletons in Her Cupboard: Festschrift for Juliet Clutton-Brock , Edited by: Clason , A. , Payne , S. and Uerpmann , H. P. 243 – 250 . Oxford : Oxbow Books .
- Yanagihara , R. , Nerurkar , V. R. , Scheirich , I. , Agostini , H. T. , Mgone , C. S. , Cui , X. H. Jobes , D. V. 2002 . JC Virus genotypes in the western Pacific suggest Asian mainland relationships and virus association with early population movements . Human Biology , 74 ( 3 ) : 473 – 488 .
- Yen , D. E. 1974 . The Sweet Potato in Oceania: An Essay in Ethnobotany. , Honolulu : Bishop Museum Press .
- Yen , D. E. 1991 . “ Domestication: The lessons from New Guinea ” . In Man and a Half: Essays in Pacific Anthropology and Ethnobiology in Honour of Ralph Bulmer , Edited by: Pawley , A. 558 – 569 . Auckland : Polynesian Society .
- Zerega , N. J. C. , Ragone , D. and Motley , T. J. 2004 . Complex origins of breadfruit (Artocarpus altilis, moraceae): Implications for human migrations in Oceania . American Journal of Botany , 91 ( 5 ) : 760 – 766 .
- Zhang , D. P. , Rossel , G. , Kriegner , A. and Hijmans , R. 2004 . AFLP assessment of diversity in sweet potato from Latin America and the Pacific region: Its implications on the dispersal of the crop . Genetic Resources and Crop Evolution , 51 ( 2 ) : 115 – 120 .
- Ziętkiewicz , E. , Rafalski , A. and Labuda , D. 1994 . Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification . Genomics , 20 ( 2 ) : 176 – 183 .
- Zizumbo-Villarreal , D. and Quero , H. J. 1998 . Re-evaluation of early observations on coconut in the New World . Economic Botany , 52 ( 1 ) : 68 – 77 .