
ABSTRACT
Epigenetic modifications, including DNA methylation, are proposed mechanisms explaining the impact of parental exposures to foetal development and lifelong health. Micronutrients including folate, choline, and vitamin B12 provide methyl groups for the one-carbon metabolism and subsequent DNA methylation processes. Placental DNA methylation changes in response to one-carbon moieties hold potential targets to improve obstetrical care. We conducted a systematic review on the associations between one-carbon metabolism and human placental DNA methylation. We included 22 studies. Findings from clinical studies with minimal ErasmusAGE quality score 5/10 (n = 15) and in vitro studies (n = 3) are summarized for different one-carbon moieties. Next, results are discussed per study approach: (1) global DNA methylation (n = 9), (2) genome-wide analyses (n = 4), and (3) gene specific (n = 14). Generally, one-carbon moieties were not associated with global methylation, although conflicting outcomes were reported specifically for choline. Using genome-wide approaches, few differentially methylated sites associated with S-adenosylmethionine (SAM), S-adenosylhomocysteine (SAH), or dietary patterns. Most studies taking a gene-specific approach indicated site-specific relationships depending on studied moiety and genomic region, specifically in genes involved in growth and development including LEP, NR3C1, CRH, and PlGF; however, overlap between studies was low. Therefore, we recommend to further investigate the impact of an optimized one-carbon metabolism on DNA methylation and lifelong health.
KEYWORDS:
Introduction
Foetal exposures within the intra-uterine environment can affect health outcomes of the offspring during the life course. This is known as the developmental origins of health and disease (DOHaD) paradigm [Citation1,Citation2]. Epigenetic modifications can affect gene-expression and consequently cell function without changing the DNA sequence and are proposed underlying mechanisms connecting parental exposures to gamete maturation, embryonic and foetal development, and long-term health outcomes in the offspring [Citation1,Citation3].
The best characterized epigenetic mechanism is DNA methylation. DNA methylation is regulated by DNA methyltransferases (DNMTs), which transfer methyl groups predominantly to the C-5 position of a cytosine at a cytosine-phosphate-guanine (CpG) site [Citation4,Citation5]. Methylation of CpGs in regulatory genomic regions like promoters typically leads to gene silencing while non-genic, repetitive DNA sequences, such as transposable elements (e.g., LINE-1 and Alu elements), are often heavily methylated to maintain genomic stability and can serve as markers for global methylation [Citation4–6].
During the periconception period, starting 14 weeks before conception till 10 weeks after conception, significant epigenetic modifications with potential effects along the life course take place [Citation7]. Most DNA methylation marks present in parental gametes are removed during the first cell divisions in the zygote and blastocyst stages, followed by the establishment of de novo DNA methylation most prominent after implantation. In utero, proper de novo DNA methylation is essential for key biological processes involved in regulation of gene expression during development and differentiation [Citation4–8]. Therefore, the periconception and pregnancy period is a vulnerable time span for epigenetic disruptions.
The one-carbon metabolism provides methyl groups required amongst others for DNA methylation. Herein, methionine is converted to the major methyl donor s-adenosylmethionine (SAM), which in turn is converted to s-adenosylhomocysteine (SAH) thereby donating its methyl group. Other essential substrates of the one-carbon metabolism, here referred to as one-carbon moieties, include methyl donors such as folate and choline, and co-factors including vitamin B2, B6, and B12, that are all mostly derived from diet () [Citation9].
Figure 1. Simplified overview of the one-carbon metabolism.
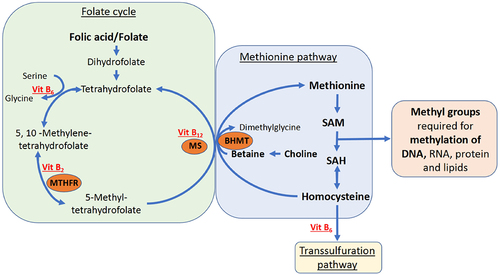
Evidence from predominantly animal studies indicates that differences in one-carbon metabolism during pregnancy can cause lasting DNA methylation changes in the offspring [Citation7,Citation10–17]. A famous example is that a methyl-supplemented diet fed to pregnant dams of the Agouti mouse increases DNA methylation of a regulating locus that results in a lower expression of the Agouti gene in their offspring, resulting in changed adiposity and coat colour [Citation18,Citation19]. In humans conceived during the Dutch Hunger Winter, an increased risk for obesity was observed and methylation differences in whole blood are found related to genes involved in growth and metabolism including lower methylation of insulin-like growth factor 2 (IGF2) gene [Citation20]. Additionally, offspring of women who used periconceptional folic acid supplements had a higher methylation of IGF2 in blood [Citation15]. This illustrates that nutrition, particularly moieties of one-carbon metabolism, plays a crucial role in early life epigenetic programming with potential lifelong health consequences [Citation7].
The placenta is an interesting target organ for DNA methylation studies investigating the relationship between maternal exposures including nutrition, one-carbon metabolism, and foetal programming as it forms the unique and indispensable interface between the mother and the developing embryo and foetus [Citation21]. The changes observed in the placental DNA methylation are suggested to reflect alterations in placental functioning and as such impact foetal programming. The treatment of these adverse variations could provide targets for future preventative or therapeutic interventions () [Citation22]. Additionally, we suggest that the presence of placental-originated cell free DNA (cfDNA) in maternal blood will become an opportunity to non-invasively assess placental DNA methylation profiles during early pregnancy, whereas the non-invasive collection of foetal tissues is not possible. This emphasizes the potential of the placenta as a proxy for foetal health conditions [Citation23].
Figure 2. The one-carbon metabolism as an underlying pathway affecting placental DNA methylation and lifelong health outcomes of the offspring.
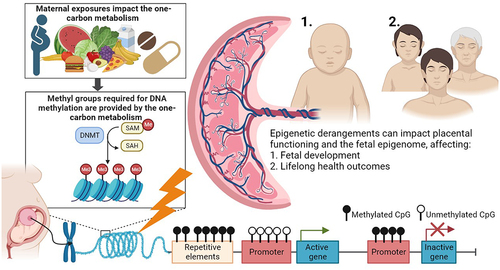
Previous reviews about the impact of the one-carbon metabolism in reproduction have mainly focussed on DNA methylation in tissues other than placenta including umbilical cord blood, focussed on animal studies and/or were limited to one specific one-carbon moiety [Citation7,Citation10–14,Citation24–26]. Additionally, epigenome-wide association studies have specifically emerged over the last years [Citation27]. Therefore, this systematic review aims to provide an overview of the current knowledge obtained from human studies investigating the impact of the one-carbon metabolism on placental DNA methylation. Epigenetic derangements in response to impaired one-carbon metabolism could hold potential targets for future therapeutic or preventative interventions, including prenatal diagnosis and obstetrical care.
Methods
Protocol and registration
The protocol for this systematic review was registered to the PROSPERO registry (PROSPERO 2023 CRD42023393358). The PRISMA Guidelines for systematic reviews and meta-analysis protocols were followed [Citation28].
Search strategy
Medline (on the Ovid platform), Embase, Web of Science Core Collection, Cochrane Central Register of Controlled Trials, and Google Scholar were searched until 12 July 2023. An expert research librarian was involved in setting up the search strategy. The search strategy included but was not limited to terms related to one-carbon metabolism like folate, homocysteine, and nutrition combined with placental related terms and key words related to epigenetics including DNA methylation. The full search is provided in Table S1.
Study selection
Clinical studies were eligible if they studied one-carbon moieties during the preconception period and/or during pregnancy including birth and studied DNA methylation in human placental tissues. Since in practice, one-carbon moieties are consumed by foods and vitamin supplements, we also included articles investigating associations between dietary patterns reflecting the intake of one-carbon moieties or intake of multivitamin supplements. In vitro studies were eligible if they studied one-carbon moieties and DNA methylation in a human in vitro placental model. Review articles, conference abstracts, and articles not available in English language were excluded. MV and SS performed the title and abstract screening and the full-text review of remaining articles.
Quality assessment of included studies
We used the ErasmusAGE quality score system for systematic reviews to assess the quality of included clinical studies [Citation29]. Quality of included articles was assessed by MV and SS independently. All articles were scored based on five items and each item was scored zero, one or two points, leading to a score between 0–10 for each article. These items include study design (cross-sectional study = 0, longitudinal study = 1, intervention study = 2), study size (<50 patients = 0, 50–100 patients = 1, >100 patients = 2), method of measuring exposure and method of measuring outcome (both: no appropriate measure = 0, moderate quality of measure = 1, adequate measure = 2), and adjustments for confounders (no adjustment = 0, adjustment for key confounders = 1, adjustment for additional covariates or extra confounders = 2) (Table S2) [Citation29]. We discussed all articles with an ErasmusAGE quality score of at least 5.
Data extraction
Data extraction was done using a predefined form. Collected data included: study design, country of origin, year of publication, study population including sample size, method of measuring (exposure to) one-carbon moieties, timing of exposure (e.g., pre-and periconceptional, pregnancy), which part of placental tissue was studied (e.g., foetal or maternal side biopsy), focus of DNA methylation (global, genome-wide, or gene-specific approach), specified methylation technique. Lastly the main outcomes of the study related to DNA methylation were collected.
Results
Study selection
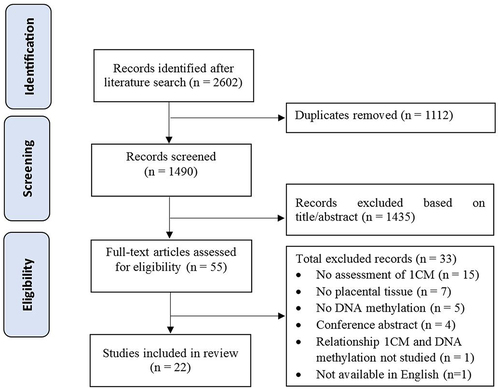
The literature search resulted in 1490 unique records after duplicate removal. After title and abstract screening, 1435 articles were excluded and a total of 55 full-text articles were assessed for eligibility of which 22 were included for the current review (). The reasons for final exclusion included no exposure to or measurements of one-carbon moieties (n = 15), no placental tissue studied (n = 7), no DNA methylation as outcome (n = 5) and other (n = 6) ().
Figure 3. Flowchart of included and excluded articles. 1 CM = one-carbon moieties.
Study characteristics
The study characteristics and main outcomes of included studies are provided in . The included clinical studies were observational (n = 16) or clinical intervention studies (n = 3). Two solely in vitro studies were included [Citation50,Citation51] and one observational study combined clinical and in vitro work [Citation43]. All included in vitro studies used human placental cell lines.
Table 1. Study characteristics and main findings of included studies ordered according to the quality score.
One-carbon moieties that were studied in the clinical studies were folate/folic acid, (n=9), vitamin B12 (n=9), homocysteine (n=4), choline (n=2), SAM and SAH (n=3), combination supplements (n=4), and dietary patterns reflecting differences in intake of one-carbon moieties (n=1). Two in vitro studies investigated the effect of folate and one studied the effect of choline on DNA methylation (). Results are discussed below for the different included moieties.
Most clinical studies used a gene-specific approach (n=14/20). Four used a genome-wide approach and global DNA methylation was investigated in nine studies. Some studies used multiple approaches in the same cohort. summarizes the studies that investigated the associations for all three approaches in clinical studies with an ErasmusAGE quality score of at least 5 points. Included clinical studies scored between 3 and 8 points (median 5) (Table S2).
Table 2. Direct associations between one-carbon moieties and global methylation, genome-wide methylation or gene-specific methylation in included clinical studies with an ErasmusAGE quality score of at least 5 out of 10 points.
Metabolic determinants of one-carbon metabolism and placental DNA methylation outcomes
Folate/Folic acid
Dietary intake/supplement use
One study (ErasmusAGE score 6) assessed dietary folate intake during pregnancy by Food Frequency Questionnaires (FFQ) and found no association with methylation of partially- or highly methylated domains after controlling for the False Discovery Rate (FDR) [Citation38].
Liu et al. (ErasmusAGE score 7) showed lower methylation of 5 out of 9 investigated sites within FAM198A in women who started using folic acid supplements before pregnancy in male offspring (p-values between 0.026 — 0.050) but not in female offspring, indicating a sex-specific impact on DNA methylation. No associations were found between use of folic acid supplements and methylation of 2 ANKRD20B sites [Citation35]. FAM198A and ANKRD20B sites were investigated, after these were formerly identified as top-ranked differentially methylated positions between 3 small for gestational age (SGA) children and 3 appropriate growth for gestational age (AGA) controls. Loke et al. (ErasmusAGE score 6) showed lower methylation of the differentially methylated region (DMR) of the H19 promoter in women using folic acid supplements in first trimester compared to no use of folic acid supplement, but found no methylation differences in the IGF2/H19 imprinted control region (ICR) or two IGF2 DMRs [Citation36].
Blood.
Maternal
In a multivariate model, folate levels associated positively with methylation of 9 out of 13 investigated CpGs at the transcription start site of LEP (range: β = 0.16—0.26, p = 0.003—p = 0.03) but not in univariate analyses (ErasmusAGE score 5) [Citation44]. In another study, folate was positively associated with methylation of the MMP-9 promoter (r = 0.58, p < 0.001) [Citation41] (both ErasmusAGE score 5).
Foetal (umbilical cord)
Van Otterdijk et al. (ErasmusAGE score 5) investigated average methylation of CpG islands, defined as genomic regions between 200 and 500 base pairs with >50% CG content, the 0–2 kb and 2-4kb up- and downstream regions next to CpG-islands, referred to as ‘shores’ and ‘shelves’ respectively, and CpGs outside these regions referred to as ‘open sea.’ Additionally, they took a genome-wide approach investigating single CpGs. The mother-infant dyads in their study were either homozygous for the wild-type MTHFR677 genotype or for the MTHFR C677T variant. The MTHFR C667T polymorphism leads to reduced methylenetetrahydrofolate reductase (MTHFR) activity which is involved in the one-carbon metabolism, leading to higher homocysteine and lower SAM concentrations (). In the total study population, red blood cell (RBC) folate was positively associated with methylation of ‘shelve’ regions and CpG island. In mother-infant dyads homozygous for the MTHFR C677T variant, a positive association was also found between RBC folate and methylation of ‘open sea’ and ‘shore’ regions. No associations between individual CpGs and RBC folate remained significant after correcting for multiple testing [Citation42].
Other
Rahat et al. (2022) (ErasmusAGE score 5) found no association between folate levels within placental villi and global DNA methylation or LINE-1 methylation [Citation43].
In vitro studies
Investigation of DNA methylation in JEG-3, a placental choricocarcinoma-derived cell line and in HTR-8/SVneo cell line, a model of extra-villous trophoblast, in response to folic acid treatment showed no changes in LINE-1 methylation [Citation43]. In contrast, global methylation decreased in both cell lines in a dose-dependent manner with the largest effect in HTR-8/SVneo cells, where a 4.3% decrease (p < 0.001) was found at 10−7 M folic acid, comparable to the physiological range of 400–600 µg/day, and a 5.7% decrease (p<0.001) was found at a much higher concentration of 10−4 M folic acid. Taking a gene-specific approach, promoter methylation of the tumour suppressor gene APC increased in both cell lines in response to folic acid, while for P16 an increase was observed in JEG-3 cells while methylation decreased in HTR-8/SVneo cells. Methylation of the promoter regions of oncogenes c-myc and c-jun decreased in both cell lines [Citation43]. No changes were found in the MMP-9 promoter region. In a comparable in vitro study, methylation in the promoter regions of SNRPN, PEG10, and MEST was studied [Citation51]. Folic acid suppletion decreased methylation of SNRPN in a dose-dependent manner in JEG-3, HTR-8/SVneo cell lines and in cytotrophoblasts. No changes were found for PEG10 and MEST [Citation51].
Interim conclusions – Folate/folic acid
Mixed results have been reported for the relationship between folate/folic acid and markers of global placental DNA methylation. Only one study took a genome-wide approach and identified no significant associations between methylation of single CpGs and cord blood folate. When taking a gene-specific approach, several – but not all – investigated sites associated with folate/folic acid (). This indicates site-specific relationships between DNA methylation and folate/folic acid.
Vitamin B12
Supplement use
In a randomized trial in Indian women with low vitamin B12 levels, LINE-1 methylation and VEGF promoter methylation did not differ between women who used vitamin B12 supplements from end first trimester till delivery as compared to women without vitamin B12 supplements (ErasmusAGE score 8) [Citation30].
Blood
Maternal
No associations were found between vitamin B12 levels at 28 weeks of gestation and 4 investigated IGF2/H19 DMRs (ErasmusAGE score 6) [Citation36]. Contrary, vitamin B12 levels in first half of pregnancy associated with lower methylation of 2 out of 13 investigated CpGs at the transcription start site of LEP (β = −0.8, p = 0.02; β = −0.02, p = 0.02) [Citation44] and vitamin B12 associated with MMP-9 promoter methylation (r = 0.68, p < 0.001) (both ErasmusAGE score 5) [Citation41]. A negative association was found between plasma vitamin B12 at time of delivery and methylation of 3 out of 7 investigated CpGs in the MTR promoter region (r = −0.208, p = 0.049; r = −0.248, p = 0.017; r = −0.298, p = 0.004) (ErasmusAGE score 5) [Citation40].
Foetal (umbilical cord)
Van Otterdijk et al. found no significant associations between vitamin B12 and methylation of CpG islands, ‘shores,’ ‘shelves,’ or ‘open sea’ regions or with methylation of individual CpGs when taking a genome-wide approach (ErasmusAGE score 5) [Citation42].
Interim conclusions – Vitamin B12
Vitamin B12 did not associate with different markers of global placental DNA methylation or with methylation of individual CpGs taking a genome-wide approach. In a gene-specific approach, several investigated sites were associated with vitamin B12, underlining again the site-specific relationship between DNA methylation and vitamin B12 ().
Homocysteine
Blood
Maternal
No associations were found between homocysteine and 4 studied IGF2/H19 DMRs (ErasmusAGE score 6) [Citation36].
Choline
Supplement use
Jiang et al. randomized women in third trimester to either 480mg or 930mg choline per day for 12 weeks (ErasmusAGE score 6). They studied global DNA methylation and methylation of regions related to cortisol-regulating genes CRH and NR3C1, methylation of DMR0 of IGF2 and methylation of LEP, GNAS-AS1, and IL10 promoter regions. Women in the 930mg group had higher global methylation compared to the 480mg group (4.4 ±0.2% VS 3.6 ±0.2%, p = 0.02) and higher methylation of the CRH promoter region (p=0.05) and the 5= untranslated exon 1F (and flanking regions) of NR3C1 (p=0.002). Of the individual CpGs, 4 out of 15 studied NR3C1 CpGs showed higher methylation while none of the 5 studied individual CpGs for the CRH region reached significance. Although 1 out of 11 studied GNAS-AS1 CpGs was lower methylated in the 930mg group (p = 0.02), average GNAS-AS1 methylation did not differ between groups. No differences were found in methylation of IGF2, IL10, or LEP promoter regions [Citation34].
Blood
Maternal
One study determined maternal plasma choline in first trimester, third trimester, and at term, and investigated methylation levels of 12 genes involved in foetal growth, lipid and energy metabolism, or adipogenesis and DNA methylation of LINE-1 elements (ErasmusAGE score 6). A negative modest association was found between DNA methylation of PPARG1A, NR3C1, HSD11β2, PPARA, and RXRA and maternal choline at all three time points (range: r = −0.188 — r = −0.452). For AdipoQ a negative association with maternal choline levels was observed in first and third trimester (r = −0.307, p = 0.001; r = −0.201, p = 0.030) and for NDUFB6 only in third trimester (r = −0.214, p = 0.021). No associations were found with MEG3, H19, MEST, and LEP methylation. LINE-1 methylation was negatively associated with maternal choline in first and third trimester (r = −0.205, p = 0.022; r = −0.207, p = 0.026) [Citation37].
Foetal (umbilical cord)
The same study showed a negative association between choline in cord blood and methylation of NR3C1, PPARA, and LINE-1 (r=−0.20, p=0.033; r=−0.223, p=0.017; r=−0.248, p=0.008). No associations were found with methylation of MEG3, H19, MEST, LEP, AdipoQ, NDUFB6, PPARG1A, HSD11β2, or RXRA [Citation37].
In vitro studies
In an in vitro study using BeWo cells as a model for first trimester trophoblast, global DNA methylation was not altered by choline treatment [Citation50].
Interim conclusions – Choline
Mixed results have been reported for choline and markers of global placental DNA methylation as well as for choline and NR3C1 methylation. Several other relationships between DNA methylation of specific genes and choline intake or blood levels were found depending on studied site and moment of measuring choline, indicating site-specific relationships between DNA methylation and choline which may differ depending on timing of exposure ().
S-adenosylmethionine (SAM) and S-adenosylhomocysteine (SAH)
Blood
Foetal (umbilical cord)
The SAM:SAH ratio was positively associated with methylation of ‘open sea’ and ‘shore’ regions only in mother-infant dyads homozygous for the MTHFR C677T genotype. There were no significant associations between SAM:SAH and CpG island or ‘shelve’ regions and the observed positive associations with SAM and negative associations with SAH did not reach significance for the different genomic regions. When investigating individual CpGs genome-wide, limited associations were found. The SAM:SAH ratio associated with individual CpGs located in RAD54L2, ETS2, ZNF836, and ABAT, SAM associated with two loci located in LOXL3 and GNL1 gene bodies and SAH associated with methylation of one CpG in a ZC3H12D CpG island. Several other identified associations did not reach significance after correcting for multiple testing (ErasmusAGE score 5) [Citation42].
Other measures
Heil et al. (ErasmusAGE score 5) studied SAM and SAH levels in placentas and methylation of LINE-1 and 1 CpG in the 5’ flanking region of PlGF. SAM and SAM:SAH ratio were positively associated with PlGF methylation (r=0.31, p=0.006; r=0.24, p=0.04, respectively). LINE-1 methylation associated with PlGF methylation (r=0.40, p<0.001) but no association between LINE-1 and SAM or SAH was studied [Citation39].
Interim conclusions – SAM/SAH
A higher SAM:SAH ratio indicates higher availability of methyl group donors (). Although only two studies investigated SAM and SAH, the SAM:SAH ratio in umbilical cord blood associated positively with markers of global methylation depending on genotype, and SAM, SAH, and SAM:SAH associated with methylation of a few individual CpGs in a genome-wide study. Both placental SAM and SAM:SAH ratio associated positively with PlGF methylation (). This suggests that SAM and SAH can impact placental DNA methylation both at a regional/global level as well as at gene-specific sites.
Combinations
Dietary patterns
Lecorguille et al. (ErasmusAGE score 7) assessed dietary intake in the year before pregnancy and distinguished three dietary patterns [Citation32]. After adjustment for vitamin use, the ‘varied and balanced’ diet which is rich in B vitamins, choline, and methionine but low in betaine showed a positive association with a DMR related to NPDC1 (p=0.02). A ‘vegetarian tendency’ diet which is rich in vitamin B6, folate, and betaine but low in vitamin B12 showed a significant negative association with 2 DMRs related to DLL1 (p = 5.3.10−7, p = 7.0.10−9) and FAR1 (p = 3.7.10−9, p = 7.8.10−3). Lastly, ‘bread and starchy food’ diet which is rich in betaine but low in vitamin B2, B6, and folate associated negatively with DMRs located near AS3MT and GSDMD, and positively with DMRs related to SLC25A46 and ZNF175 (p = 0.02; p = 0.004; p = 0.01; p = 0.006, respectively). No associations were found with individual CpGs or global DNA methylation [Citation32].
Supplement use
An intervention study (Erasmus AGE score 6) randomized women to daily use of small quantity lipid-based micronutrient (SQLNS) starting at least three months before conception until delivery versus no supplements. SQLNS contain essential fatty acids, proteins, and multiple vitamins including folate, vitamin B6, and B12. Methylation of 2 CpGs in the IGF1 promoter region were assessed. No differences were found between study groups [Citation33].
Composition of prenatal vitamins varies but they usually contain folic acid, vitamin B6, and B12 as well as other micronutrients. Lecorguille et al. (ErasmusAGE score 7) found higher methylation of Alu repetitive elements in women taking vitamin supplements before and during pregnancy compared to women not taking any vitamins (β = 0.40, p = 0.005) although no differences in LINE-1 methylation were found. Vitamin use only before or during pregnancy was not associated with Alu or LINE-1 methylation [Citation32].
Two studies investigated placental DNA methylation and self-reported maternal vitamin intake in the first month of pregnancy. Dou et al. (ErasmusAGE score 8) studied DNA methylation in the MARBLES and in the EARLI cohort, both comprise couples at increased risk for offspring with autism spectrum disorder (ASD). Mean array methylation for MARBLES women taking vitamins was −0.60% (CI −1.08, − 0.13) and the same magnitude of effect was seen in the EARLI cohort, although not significant −0.52% (CI −1.04, 0.01). No individual CpGs associated with vitamin intake using the FDR-corrected significance threshold (p < 1.0−7). With a less stringent significance threshold of p < 0.01, they found 9216 and 3442 differentially methylated CpGs in the MARBLES and EARLI cohort, respectively. About 95% of these showed lower methylation in the vitamin group in both cohorts (average −4%). The identified CpGs were enriched in neuronal developmental pathways [Citation31]. Schmidt et al. (ErasmusAGE score 6) found no associations between vitamin use and methylation of predefined partially- or highly methylated domains or methylation of enhancers, active promoters, and bivalent promoters [Citation38].
Interim conclusions – combinations
Global methylation was not associated with dietary patterns reflecting differences in intake of one-carbon moieties and use of vitamin supplements showed both increased and decreased global methylation as well as no differences depending on used technique and/or timing of supplement use. A genome-wide study revealed a few DMRs associated with dietary patterns, but dietary patterns or vitamin supplement use did not associate with individual CpGs after FDR correction. IGF1 promoter methylation was not altered in women taking SQLNS (). In general, dietary patterns and vitamin supplement use did not substantially impact placental DNA methylation in included studies.
Discussion
This review systematically summarizes associations between the one-carbon metabolism and placental DNA methylation. Included studies suggest that one-carbon moieties can impact placental DNA methylation at multiple specific genomic regions rather than affecting global methylation. On the other hand, this review shows that there is a lot of heterogeneity among studies regarding investigated exposures, studied outcomes, sample size, and study design. Additionally, differences in DNA methylation have been found between female and male offspring [Citation35], depending on which part of the placenta was sampled [Citation52], timing of exposure [Citation32,Citation37], and underlying genotype [Citation42], which could also explain parts of observed differences between studies. Based on the use of ErasmusAGE quality score, we discussed studies with a quality score of at least 5 out of 10 points.
Global Methylation
A wide variety of techniques to study global methylation have been used by included studies (). Global DNA methylation was not altered by the use of vitamin B12 supplements, while higher intake of choline did increase global DNA methylation. One study found associations between one-carbon moieties and average DNA methylation to differ between genomic regions (i.e., CpG islands, ‘shores,’ ‘shelves’ and ‘open sea’) [Citation42]. Overall, most studies found no associations between one-carbon moieties and measures of global DNA methylation (). Hence, higher availability of methyl donors provided by the one-carbon metabolism does not just lead to increased global placental methylation.
Genome-wide approach
The number of included genome-wide studies is limited, and only three had a quality score of at least five. All three studies used the Illumina Infinium HumanMethylation450 (450K) BeadChip array, which covers >480,000 CpGs and one partly used the Infinium methylation EPIC array covering >850,000 CpGs. All three studies investigated different exposures (). CpGs associated with maternal vitamin use in the first month of pregnancy could only be identified when a less stringent FDR-corrected p-value was applied and 95% of these were hypomethylated in women who did take vitamins in the first month of pregnancy. Identified CpGs were enriched in neuronal developmental pathways [Citation31]. A few DMRs linked to different genes could be identified based on dietary patterns reflecting differences in intake of one-carbon moieties [Citation32] and methylation of a few CpGs associated with umbilical cord levels of SAM, SAH, and SAM:SAH, but not with RBC folate or vitamin B12 levels in cord blood [Citation42].
Gene-specific approach
Included studies mostly investigated different loci, hampering comparison of results. Overall, most gene-specific studies did find relationships between one-carbon moieties and placental DNA methylation. On the other hand, most studies reported both significant and non-significant associations and there is a lack of uniformity in directionality of identified associations, indicating site-specific relationships ().
Genes involved in growth and metabolism
Genes involved in growth and metabolism have been of particular interest in included studies.
The imprinted IGF2/H19 cluster plays an important role in foetal and placental growth [Citation53]. Methylation of IGF2/H19 has been investigated in multiple studies, with most reporting no significant associations with investigated one-carbon moieties [Citation36,Citation37]. In intervention studies, higher intake of choline did also not alter IGF2 methylation [Citation34] and the use of SQLNS did not affect methylation of IGF1, another major growth factor [Citation33]. This is in contrast with other studies showing that maternal intake of methyl-group donors including folate/folic acid associated negatively with IGF2 methylation in buccal epithelial cells in 6 months old children and positively with IGF2 methylation in infant blood and cord blood suggesting a tissue-specific response [Citation15,Citation54,Citation55].
LEP encodes the hormone Leptin which plays a central role in energy homoeostasis including appetite regulation and mutations in LEP can contribute to the development of obesity and diabetes type 2 [Citation56]. Different results for LEP methylation have been reported depending on studied one-carbon moiety: positive associations with serum folate, negative associations with serum vitamin B12 [Citation44], and no associations with choline have been reported [Citation37] (Both ErasmusAGE score 6).
Cortisol regulating genes could play a role in multiple diseases including cardiovascular complications. Jiang et al. investigated choline intake (480mg/daily versus 930 mg/daily) and found increased methylation in the higher dose group of the CRH promotor and the 5’untranslated exon 1F of NR3C1 [Citation4]. In contrast, NR3C1 methylation associated negatively with choline levels in a study by Nakanishi et al. (ErasmusAge score 6) [Citation37]. A possible explanation is that both studies targeted a different genomic region of NR3C1 or there may be lack of comparability between blood levels of choline and use of choline supplements.
Nakanishi et al. investigated several other gene-specific sites related to growth and metabolism and choline in maternal blood and cord blood. Negative associations with choline levels were also found for AdipoQ, PPARG1A, HSD11β2, PPARA, NDUFB6, and RXA, while no associations were found with methylation of MEG3 and MEST, all involved in foetal growth. Differences have been found for different time points during pregnancy [Citation7]. In an in vitro study, MEST methylation was also not altered in response to folic acid [Citation51].
MMP-9 codes for a matrix metalloproteinase induced during labour and MMP-9 is also increased in multiple cardiovascular diseases including hypertension and myocardial infarction [Citation57]. Serum vitamin B12 and folate associated positively with MMP-9 promoter methylation and with lower MMP-9 RNA and protein levels [Citation41]. Contrary, folic acid treatment did not change MMP-9 methylation in an in vitro model [Citation43].
Genes related to placental development and function
The one-carbon metabolism has also been linked to placental angiogenesis which is crucial for a well-functioning placenta. The placenta itself produces several angiogenic factors involved in endothelial growth and function, including vascular endothelial growth factor (VEGF) and placental growth factor (PlGF), both of which are decreased in preeclampsia (PE) [Citation58,Citation59]. For example, increased maternal choline intake can suppress anti-angiogenic soluble fms-like tyrosine kinase 1 (sFLT-1) expression in human placentas and promotes angiogenesis in mice placentas [Citation10]. Vitamin B12 supplements did not alter VEGF methylation in a high-quality intervention study [Citation30], while another study showed PlGF methylation associated with SAM and SAM:SAH in the placenta [Citation39]. Since a higher SAM:SAH ratio indicates higher availability of methyl group donors (), a deranged one-carbon metabolism may be involved in the pathogenesis of placental-related complications partly through inducing epigenetic changes resulting in impaired placental functioning.
Genes directly related to one-carbon metabolism
Plasma vitamin B12 was negatively associated with MTR promoter methylation. This can have an impact on the one-carbon metabolism, since MTR codes for methionine synthase which is crucial in the conversion of homocysteine to methionine [Citation40] ().
Genes selected based on prior genome-wide analysis
Lastly, Liu et al. selected gene-specific sites of interest based on DMRs identified through genome-wide analysis. Methylation of FAM198A associated negatively with preconceptional start of folic acid only in male offspring indicating a sex-specific impact on DNA methylation. No associations were found between timing of folic acid supplements and methylation of 2 ANKRD20B CpGs [Citation35].
In summary, most studies reported both significant and non-significant associations depending on the studied genes and moieties, indicating a site-specific relationship between the one-carbon metabolism and DNA methylation at various genes in the placenta ().
Previous Reviews
Our review is largely in line with previously published human and animal reviews. James et al. focused predominantly on DNA methylation in cord blood and showed several associations between maternal one-carbon metabolism and methylation of specific genes. Overlapping genes with the current review include IGF2/H19, LEP, NR3C1, RXRA, MEG3, MEST, IL10, and GNAS-AS1 and several other associations were reported. For most genes, differences were found depending on studied tissue, moiety, specific loci, and timing of exposure [Citation12]. In rodents, maternal folic acid supplements [Citation24] and choline intake or methyl group supplemented diets [Citation14] have been shown to impact DNA methylation at several gene-specific sites, leading to both hypo-and hypermethylation depending on studied genomic region. Additionally, these studies found mixed outcomes for global methylation in multiple offspring tissues including brain, mucosa, and liver. For homocysteine, associations with hypomethylation were more prominent in animal models as compared to human studies, where mixed results have been reported [Citation25]. Likewise, multiple studies using livestock showed that dietary restrictions or excesses, or a methyl supplemented diet can affect both gene-specific and global DNA methylation in the offspring, depending on studied tissue, offspring sex, and timing of exposure [Citation26,Citation60]. A systematic review focusing on physical activity and dietary intake of carbohydrates, fats, and proteins during pregnancy showed similar mixed associations with DNA methylation in placental tissues and cord blood [Citation61]. This could indicate a role for a variety of lifestyle behaviours affecting the epigenome, however, the one-carbon metabolism is influenced by lifestyle behaviours which could also be (part of) the underlying biological mechanism for observed associations.
Strengths and Limitations
In the present article, we systematically review studies investigating relationships between different one-carbon moieties and DNA methylation focusing on the human placenta. We included a wide range of exposures and outcomes and were not limited to a specific timing in relation to delivery, to be able to give a comprehensive overview of current literature. Consequently, heterogeneity between studies limits comparability of results and a meta-analysis is therefore not possible. Besides, most included studies face several challenges which should be taken into account. First, dietary patterns and vitamin use are often associated with other (lifestyle) behaviours potentially affecting DNA methylation, including smoking, BMI, and socio-economic status. Most studies are observational and do not adequately adjust for potential confounders. Second, one-carbon moieties are usually not consumed independently of each other and of other nutrients in the diet, creating difficulties when studying the effect of a single moiety. Additionally, most studies were performed in healthy cohorts without major nutritional deficiencies. The effect of differences in one-carbon moieties could be limited within a normal range. Sample size of most included studies is relatively small and self-reported exposures like food questionnaires and vitamin intake could be less reliable. Differences in sampling methods and storage conditions of placental tissues could also lead to differences between studies. Lastly, the timing between exposure and measuring outcome may affect results. Placental DNA methylation is only studied postpartum in all included studies, and might not accurately reflect methylation profiles at time of earlier exposures because of accumulating exposures during gestation also impacting DNA methylation profiles [Citation62].
Clinical Implications and Future Research
Current studies investigating one-carbon metabolism and placental DNA methylation display low overlap in terms of studied exposures and genomic regions, and the number of high-quality intervention studies is limited. Therefore, high-quality intervention studies in combination with a genome-wide readout for DNA methylation and preferably randomized and blinded, are warranted to further investigate the impact of an optimized maternal one-carbon metabolism on DNA methylation profiles and health in the offspring. Ideally, studies will include follow-up after birth to assess the impacts on lifelong health.
Furthermore, DNA methylation is highly cell type specific. Investigating placental DNA methylation per cell type could therefore decrease heterogeneity both within and between studies. Moreover, placental tissue might not be an appropriate model for some research questions and other tissues or cell types could also be incorporated [Citation63]; however, tissues from internal organs are largely inaccessible and the placenta could provide insight in both placental function and foetal epigenetic programming. Generally, placental DNA methylation is only studied at one time point, i.e., after delivery. In the future, the presence of cfDNA originated from placental cell types derived from maternal blood might be an unique in vivo opportunity to non-invasively study placental DNA methylation profiles already during pregnancy [Citation23]. Moreover, cfDNA could potentially provide targets to improve future prenatal diagnosis and obstetrical care by identifying modifiable epigenetic derangements during pregnancy, contributing to improved health outcomes across the life course.
Conclusions
Most included studies indicate relationships between several measures of one-carbon moieties in the prenatal period and site-specific DNA methylation in the placenta rather than changes in global DNA methylation. Several gene-specific associations with DNA methylation were found, especially in genes involved in growth and metabolism. A deranged one-carbon metabolism leading to detrimental changes in the foetal epigenome in genes involved in for example the cardiovascular system may support the DOHaD paradigm. Changes in DNA methylation could not only lead to altered placental functions but could also serve as a proxy for foetal epigenetic programming, with both consequences for foetal development as well as future health outcomes.
On the other hand, conflicting outcomes were reported and there is a lack of uniformity between studies in geographically study populations, investigated loci, exposure measures, and study designs. Moreover, DNA methylation is only studied in postpartum placentas, though this may be overcome by investigating methylation of cfDNA in future studies. High-quality intervention studies tackling common limitations of included studies are needed to elucidate the relationship between one-carbon metabolism and specific differences in epigenetic programming in the offspring, ideally with longitudinal follow-up after birth. This could enable us to assess the corresponding impact on health outcomes across the life course and identify preventative or therapeutic measures contributing to improved health outcomes for future generations.
List of abbreviations
| ABAT | = | 4-aminobutyrate aminotransferase |
| AdipoQ | = | Adiponectin, C1Q and collagen domain containing |
| AGA | = | Appropriate growth for gestational age |
| ANKRD20B | = | Ankyrin repeat domain 20 family member A8, pseudogene |
| APC | = | APC regulator of WNT signaling pathway |
| AS3MT | = | Arsenite methyltransferase |
| ASD | = | Autism spectrum disorder |
| cfDNA | = | Cell free DNA |
| CpG | = | Cytosine-phosphate-Guanine |
| CRH | = | Corticotropin releasing hormone |
| DLL1 | = | Delta like canonical Notch ligand 1 |
| DMR | = | Differentially methylated region |
| DOHaD | = | Developmental origins of health and disease |
| DNMTs | = | DNA Methyltransferases |
| ETS2 | = | ETS proto-oncogene 2, transcription factor |
| FAM198A | = | Family with sequence similarity 198 member A |
| FAR1 | = | Fatty acyl-CoA reductase 1 |
| FDR | = | False Discovery Rate |
| FFQ | = | Food Frequency Questionnaires |
| GNAS-AS1 | = | GNAS antisense RNA 1 |
| GNL1 | = | G protein nucleolar 1 (putative) |
| GSDMD | = | Gasdermin D |
| H19 | = | H19 imprinted maternally expressed transcript |
| HSD11β2 | = | Hydroxysteroid 11-beta dehydrogenase 2 |
| ICR | = | Imprinted control region |
| IGF1 | = | Insulin like growth factor 1 |
| IGF2 | = | Insulin like growth factor 2 |
| IL10 | = | Interleukin 10 |
| LEP | = | Leptin |
| LINE-1 | = | Long interspersed nuclear elements 1 |
| LOXL3 | = | Lysyl oxidase like 3 |
| MEG3 | = | Maternally expressed 3 |
| MEST | = | Mesoderm specific transcript |
| MMP9 | = | Matrix metallopeptidase 9 |
| MTHFR | = | Methylenetetrahydrofolate reductase |
| MTR | = | 5-methyltetrahydrofolate-homocysteine methyltransferase |
| NDUFB6 NADH: | = | ubiquinone oxidoreductase subunit B6 |
| NPDC1 | = | Neural proliferation, differentiation and control 1 |
| NR3C1 | = | Nuclear receptor subfamily 3 group C member 1 |
| P16 or CDKN2Aink4a: | = | cyclin-dependent kinase inhibitor 2A |
| PE | = | Preeclampsia |
| PlGF | = | Placental growth factor |
| PPARA | = | Peroxisome proliferator activated receptor alpha |
| PEG10 | = | Paternally expressed 10 |
| PPARG1A | = | Peroxisome proliferator activated receptor gamma |
| RAD54L2 | = | RAD54 like 2 |
| RBC | = | Red blood cell |
| RXRA | = | Retinoid X receptor alpha |
| SAH | = | S-adenosylhomocysteine |
| SAM | = | S-adenosylmethionine |
| SGA | = | Small for gestational age |
| SLC25A46 | = | Solute carrier family 25 member 46 |
| SNRPN | = | Small nuclear ribonucleoprotein polypeptide N |
| SQLNS | = | Small quantity lipid-based micronutrient |
| VEGF | = | Vascular endothelial growth factor |
| ZC3H12D | = | Zinc finger CCCH-type containing 12D |
| ZNF175 | = | Zinc finger protein 175 |
| ZNF836 | = | Zinc finger protein 836 |
Authors’ roles
MV and SS wrote the review and selected the articles. MV, SS, RS, JB conceptualized the scope of the review. All authors contributed to the writing of this article and approved the final version.
-)Supplement Review 1CM placenta DNAm.docx
Download MS Word (18.8 KB)Acknowledgements
The authors would like to thank Wichor M. Bramer, biomedical information specialist, Erasmus University Medical Centre, for his assistance with the search strategy. was created with BioRender.com.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The authors hereby confirm that the data supporting the findings of this systematic review are available within the article.
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/15592294.2024.2318516
Additional information
Funding
References
- Gluckman PD, Hanson MA, Cooper C. Effect of in utero and early-life conditions on adult health and disease. N Engl J Med. 2008;359(1):61–26. doi: 10.1056/NEJMra0708473
- Barker DJP. The origins of the developmental origins theory. J Intern Med. 2007 May;261(5):412–7. doi: 10.1111/j.1365-2796.2007.01809.x
- Bianco-Miotto TCJ, Craig JM, Gasser YP, et al. Epigenetics and DOHaD: from basics to birth and beyond. J Dev Orig Health Dis. 2017 Oct;8(5):513–519. doi: 10.1017/S2040174417000733
- Moore LD, Le T, Fan G. DNA methylation and its basic function. Neuropsychopharmacology. 2013;38(1):23–38. doi: 10.1038/npp.2012.112
- Jin B, Li Y, Robertson KD. DNA methylation: superior or subordinate in the epigenetic hierarchy? Genes & Cancer. 2011 Jun;2(6):607–617. doi: 10.1177/1947601910393957
- Greenberg MVC, Bourc’his D. The diverse roles of DNA methylation in mammalian development and disease. Nat Rev Mol Cell Biol. 2019 Oct;20(10):590–607. doi: 10.1038/s41580-019-0159-6
- Steegers-Theunissen RP, Twigt J, Pestinger V. The periconceptional period, reproduction and long-term health of offspring: the importance of one-carbon metabolism. Human Reproduction Update. 2013 Nov-Dec;19(6):640–655. doi: 10.1093/humupd/dmt041
- Zeng Y, Chen T. DNA methylation reprogramming during mammalian development. Genes (Basel). 2019 Mar 29;10(4):257. doi: 10.3390/genes10040257
- Friso S, Udali S, De Santis D. One-carbon metabolism and epigenetics. Molecular Aspects Of Medicine. 2017 Apr;54:28–36. doi: 10.1016/j.mam.2016.11.007
- Korsmo HW, Jiang X. One carbon metabolism and early development: a diet-dependent destiny. Trends In Endocrinology & Metabolism. 2021 Aug;32(8):579–593. doi: 10.1016/j.tem.2021.05.011
- McGee MB, Bainbridge S, Fontaine-Bisson B. A crucial role for maternal dietary methyl donor intake in epigenetic programming and fetal growth outcomes. Nutrition Reviews. 2018; Jun 1;76(6):469–478. doi: 10.1093/nutrit/nuy006
- James JP, Sajjadi AS, Tomar A, et al. EMPHASIS study group. Candidate genes linking maternal nutrient exposure to offspring health via DNA methylation: a review of existing evidence in humans with specific focus on one-carbon metabolism. Int J Epidemiol. 2018 Dec 1;47(6):1910–1937. doi: 10.1093/ije/dyy153
- Anderson OS, Sant KE, Dolinoy DC. Nutrition and epigenetics: an interplay of dietary methyl donors, one-carbon metabolism and DNA methylation. J Nutr Biochem. 2012 Aug;23(8):853–859. doi: 10.1016/j.jnutbio.2012.03.003
- Lee HS. Impact of maternal diet on the epigenome during in utero life and the developmental programming of diseases in childhood and adulthood. Nutrients. 2015 Nov 17;7(11):9492–507. doi: 10.3390/nu7115467
- Steegers-Theunissen S-TR, Obermann-Borst SA, Kremer D, et al. Periconceptional maternal folic acid use of 400 µg per day is related to increased methylation of the IGF2 gene in the very young child. PloS One. 2009 Nov 16;4(11):e7845. doi: 10.1371/journal.pone.0007845
- Sinclair KD, Allegrucci C, Singh R, et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc Natl Acad Sci U S A. 2007 Dec 4;104(49):19351–19356. doi: 10.1073/pnas.0707258104
- Clare CE, Brassington AH, Kwong WY. One-Carbon Metabolism: linking nutritional biochemistry to epigenetic programming of long-term development. Ann Rev Anim Biosci. 2019; Feb 15;7(1):263–287. doi: 10.1146/annurev-animal-020518-115206
- Waterland RA, Travisano M, Tahiliani KG, et al. Methyl donor supplementation prevents transgenerational amplification of obesity. Int J Obes (Lond). 2008 Sep;32(9):1373–1379. doi: 10.1038/ijo.2008.100
- Cooney CA, Dave AA, Wolff GL. Maternal methyl supplements in mice affect epigenetic variation and DNA methylation of offspring. J Nutr. 2002 Aug;132(8):2393S–2400S. doi: 10.1093/jn/132.8.2393S
- Tobi EW, Slieker RC, Stein AD, et al. Early gestation as the critical time-window for changes in the prenatal environment to affect the adult human blood methylome. Int J Epidemiol. 2015 Aug;44(4):1211–1223. doi: 10.1093/ije/dyv043
- Burton GJ, Jauniaux E. What is the placenta? Am J Obstet Gynecol. 2015 Oct;213(4):SS6.e1–S6.e4. doi: 10.1016/j.ajog.2015.07.050
- Nelissen ECM, van Montfoort APA, Dumoulin JLH. Epigenetics and the placenta. Hum Reprod Update. 2011;17(3):397–417. doi: 10.1093/humupd/dmq052
- Wong AIC, Lo YMD. Noninvasive fetal genomic, methylomic, and transcriptomic analyses using maternal plasma and clinical implications. Trends Mol Med. 2015;21(2):98–108. doi: 10.1016/j.molmed.2014.12.006
- Liu H-Y, Liu S-M, Zhang Y-Z. Maternal folic acid supplementation mediates offspring health via DNA methylation. Reprod Sci. 2020 Apr;27(4):963–976. doi: 10.1007/s43032-020-00161-2
- Mandaviya PR, Stolk L, Heil SG. Homocysteine and DNA methylation: a review of animal and human literature. Mol Genet Metab. 2014 Dec;113(4):243–252. doi: 10.1016/j.ymgme.2014.10.006
- Thompson RP, Nilsson E, Skinner MK. Environmental epigenetics and epigenetic inheritance in domestic farm animals. Anim Reprod Sci. 2020 Sep;220:106316. doi: 10.1016/j.anireprosci.2020.106316
- Campagna MP, Xavier A, Lechner-Scott J, et al. Epigenome-wide association studies: current knowledge, strategies and recommendations. Clin Epigenetics. 2021; Dec 4;13(1):214. doi: 10.1186/s13148-021-01200-8
- Shamseer SL, Moher D, Clarke M, et al. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015: elaboration and explanation. BMJ. 2015; Jan 2;349(jan02 1):g7647. doi: 10.1136/bmj.g7647
- Hamilton O. Quality assessment tool for quantitative studies: national collaborating centre for methods and tools 2008 [Updated 2020 Apr13]. Available from: https://dev.nccmt.ca/knowledge-repositories/search/14.
- Devi S, Mukhopadhyay A, Dwarkanath P, et al. Combined vitamin B-12 and balanced protein-energy supplementation affect homocysteine remethylation in the methionine cycle in pregnant south Indian women of low vitamin B-12 status. J Nutr. 2017;147(6):1094–103. doi: 10.3945/jn.116.241042
- Dou JF, Middleton LYM, Zhu Y, et al. Prenatal vitamin intake in first month of pregnancy and DNA methylation in cord blood and placenta in two prospective cohorts. Epigenet Chromatin. 2022;15(1):28. doi: 10.1186/s13072-022-00460-9
- Lecorguille M, Charles MA, Lepeule J, et al. Association between dietary patterns reflecting one-carbon metabolism nutrients intake before pregnancy and placental DNA methylation. Epigenetics. 2022;17(7):715–30. doi: 10.1080/15592294.2021.1957575
- Castillo-Castrejon M, Yang IV, Davidson EJ, et al. Preconceptional lipid-based nutrient supplementation in 2 low-resource countries results in distinctly different IGF-1/mTOR placental responses. J Nutr. 2021;151(3):556–569. doi: 10.1093/jn/nxaa354
- Jiang X, Yan J, West AA, et al. Maternal choline intake alters the epigenetic state of fetal cortisol-regulating genes in humans. FASEB J. 2012;26(8):3563–74. doi: 10.1096/fj.12-207894
- Liu J, Zhang Z, and Xu J, et al. Genome-wide DNA methylation changes in placenta tissues associated with small for gestational age newborns; cohort study in the Chinese population. Epigenomics. 2019;11(12):1399–412. doi: 10.2217/epi-2019-0004
- Loke YJ, Galati JC, Morley R, et al. Association of maternal and nutrient supply line factors with DNA methylation at the imprinted IGF2/H19 locus in multiple tissues of newborn twins. Epigenetics. 2013;8(10):1069–1079. doi: 10.4161/epi.25908
- Nakanishi M, Funahashi N, Fukuoka H, et al. Effects of maternal and fetal choline concentrations on the fetal growth and placental DNA methylation of 12 target genes related to fetal growth, adipogenesis, and energy metabolism. J Obstet Gynaecol Res. 2021;47(2):734–44. doi: 10.1111/jog.14599
- Schmidt RJ, Schroeder DI, Crary-Dooley FK, et al. Self-reported pregnancy exposures and placental DNA methylation in the MARBLES prospective autism sibling study. Environ Epigenet. 2016;2(4):dvw024. doi: 10.1093/eep/dvw024
- Heil SG, Herzog EM, Griffioen PH, et al. Lower S-adenosylmethionine levels and DNA hypomethylation of placental growth factor (PlGF) in placental tissue of early-onset preeclampsia-complicated pregnancies. PloS One. 2019;14(12):e0226969. doi: 10.1371/journal.pone.0226969
- Khot VV, Yadav DK, Shrestha S, et al. Hypermethylated CpG sites in the MTR gene promoter in preterm placenta. Epigenomics. 2017;9(7):985–996. doi: 10.2217/epi-2016-0173
- Moeini N, Momeni AM, Zargar M, et al. The Effect of B9 and B12 Vitamins Deficiency on Hypomethylation of MMP-9 gene Promoter Among Women With Preterm Parturition. Biochem Genet. 2022;60(1):336–350. doi: 10.1007/s10528-021-10099-y
- van Otterdijk SD, Klett H, Boerries, M, et al. The impact of pre-pregnancy folic acid intake on placental DNA methylation in a fortified cohort. FASEB J. 2023;37(1):e22698. doi: 10.1096/fj.202200476RR
- Rahat B, Hamid A, Bagga R, et al. Folic acid levels during pregnancy regulate trophoblast invasive behavior and the possible development of preeclampsia. Front Nutr. 2022;9:847136. doi: 10.3389/fnut.2022.847136
- Sletner L, Moen AEF, Yajnik CS, et al. Maternal glucose and LDL-Cholesterol levels are related to placental leptin gene methylation, and, together with nutritional factors, largely explain a higher methylation level among ethnic south asians. Front Endocrinol. 2021;12:809916. doi: 10.3389/fendo.2021.809916
- Ge J, Wang J, Zhang F, et al. Correlation between MTHFR gene methylation and pre-eclampsia, and its clinical significance. Genet Mol Res. 2015;14:8021–8. doi: 10.4238/2015.July.17.10
- Pinunuri R, Castano-Moreno E, Llanos MN, et al. Epigenetic regulation of folate receptor-α (FOLR1) in human placenta of preterm newborns. Placenta. 2020;94:202–25. doi: 10.1016/j.placenta.2020.03.009
- Tserga A, Binder AM, Michels KB. Impact of folic acid intake during pregnancy on genomic imprinting of IGF2/H19 and 1-carbon metabolism. FASEB J. 2017;31(12):5149–58. doi: 10.1096/fj.201601214RR
- Zhu Y, Mordaunt CE, Yasui DH, et al. Placental DNA methylation levels at CYP2E1 and IRS2 are associated with child outcome in a prospective autism study. Hum Mol Genet. 2019;28(16):2659–74. doi: 10.1093/hmg/ddz084
- Kulkarni A, Chavan-Gautam P, Mehendale S, et al. Global DNA methylation patterns in placenta and its association with maternal hypertension in pre-eclampsia. DNA Cell Biol. 2011;30(2):79–84. doi: 10.1089/dna.2010.1084
- Jiang X, Greenwald E, Jack-Roberts C. Effects of Choline on DNA Methylation and Macronutrient Metabolic Gene Expression in in Vitro Models of Hyperglycemia. Nutr Metab Insights. 2016;9:NMI.S29465. doi: 10.4137/NMI.S29465
- Rahat B, Mahajan A, Bagga R, et al. Epigenetic modifications at DMRs of placental genes are subjected to variations in normal gestation, pathological conditions and folate supplementation. Sci Rep. 2017;7(1):40774. doi: 10.1038/srep40774
- Pinunuri R, Castano-Moreno E, Llanos MN, et al. Epigenetic regulation of folate receptor-α (FOLR1) in human placenta of preterm newborns. Placenta. 2020;94:20–25. doi: 10.1016/j.placenta.2020.03.009
- Nordin NM, Bergman D, Halje M, et al. Epigenetic regulation of the Igf2/H19 gene cluster. Cell Proliferation. 2014 Jun;47(3):189–199. doi: 10.1111/cpr.12106
- Pauwels PS, Ghosh M, Duca RC, et al. Godderis L maternal intake of methyl-group donors affects DNA methylation of metabolic genes in infants. Clin Epigenetics. 2017 Feb 7;9(1):16. doi: 10.1186/s13148-017-0321-y
- Haggarty HP, Hoad G, Campbell DM, et al. Folate in pregnancy and imprinted gene and repeat element methylation in the offspring. Am J Clin Nutr. 2013 Jan;97(1):94–99. Epub Nov 14. PMID: 23151531. doi: 10.3945/ajcn.112.042572
- Qadir MI, Ahmed Z. Lep expression and its role in obesity and type-2 diabetes. Crit Rev Eukaryot Gene Expr. 2017;27(1):47–51. doi: 10.1615/CritRevEukaryotGeneExpr.2017019386
- Yabluchanskiy YA, Ma Y, Iyer RP, et al. Matrix metalloproteinase-9: many shades of function in cardiovascular disease. Physiology. 2013 Nov;28(6):391–403. doi: 10.1152/physiol.00029.2013
- Tjwa TM, Luttun A, Autiero M. VEGF and PlGF: two pleiotropic growth factors with distinct roles in development and homeostasis. Cell Tissue Res. 2003 Oct;314(1):5–14. doi: 10.1007/s00441-003-0776-3
- Maynard SE, Karumanchi SA. Angiogenic factors and preeclampsia. Semin Nephrol. 2011 Jan;31(1):33–46. doi: 10.1016/j.semnephrol.2010.10.004
- Chavatte-Palmer C-P-P, Velazquez MA, Jammes H. Review: Epigenetics, developmental programming and nutrition in herbivores. Animal. 2018 Dec;12(s2):s363–s371. doi: 10.1017/S1751731118001337
- Rasmussen L, Knorr S, Antoniussen CS, et al. The impact of lifestyle, diet and physical activity on epigenetic changes in the offspring—A systematic review. Nutrients. 2021;13(8):2821. doi: 10.3390/nu13082821
- Lapehn S, Paquette AG. The Placental Epigenome as a Molecular Link Between Prenatal Exposures and Fetal Health Outcomes Through the DOHaD Hypothesis. Curr Envir Health Rpt. 2022;9(3):490–501. doi: 10.1007/s40572-022-00354-8
- Herzog EM, Eggink AJ, Willemsen SP, et al. The tissue-specific aspect of genome-wide DNA methylation in newborn and placental tissues: implications for epigenetic epidemiologic studies. J Dev Orig Health Dis. 2021 Feb;12(1):113–123. doi: 10.1017/S2040174420000136