
Abstract
Elevated CO2 can protect plants from heat stress (HS); however, the underlying mechanisms are largely unknown. Here, we used a set of Arabidopsis mutants such as salicylic acid (SA) signaling mutants nonexpressor of pathogenesis-related gene 1 (npr1-1 and npr1-5) and heat-shock proteins (HSPs) mutants (hsp21 and hsp70-1) to understand the requirement of SA signaling and HSPs in elevated CO2-induced HS tolerance. Under ambient CO2 (380 µmol mol−1) conditions, HS (42°C, 24 h) drastically decreased maximum photochemical efficiency of PSII (Fv/Fm) in all studied plant groups. Enrichment of CO2 (800 µmol mol−1) with HS remarkably increased the Fv/Fm value in all plant groups except hsp70-1, indicating that NPR1-dependent SA signaling is not involved in the elevated CO2-induced HS tolerance. These results also suggest an essentiality of HSP70-1, but not HSP21 in elevated CO2-induced HS mitigation.
Abbreviations
| ABA | = | abscisic acid |
| Fv/Fm | = | maximum photochemical efficiency of PSII |
| HS | = | heat stress |
| HSPs | = | heat-shock proteins |
| MAPK | = | mitogen activated protein kinase |
| npr1 | = | nonexpressor of pathogenesis-related gene 1 |
| PSII | = | photosystems II |
| SA | = | salicylic acid |
High temperature, one of the major abiotic factors affecting plant growth is consistently threatening global food security due to ongoing climate change.Citation1 Heat stress (HS) primarily targets photosynthesis for its high sensitivity and thus minimizing plant productivity. It has been reported that HS could inhibit or inactivate photosystem II (PSII), by degrading the reaction center-binding protein D1 of PSII.Citation2 Plant responses to HS often involve production and accumulation of heat-shock proteins (HSPs) which are required for heat acclimation and subseque7t tolerance.Citation1,3 Small HSPs could combine with thylakoid and protect oxygen evolving complex proteins of PSII against HS.Citation1 It has also been suggested that HSPs mainly function to prevent the damage by serving as molecular chaperons, but do not participate to reverse protein denaturation and aggregation.Citation1,4 In addition, as a phytohormone, salicylic acid (SA) has been implicated in basal thermotolerance, exogenous application of SA in Arabidopsis could induce HSPs gene expression in vivo.Citation3,5 In wheat, SA pre-treatment increases the protein kinase activity and retards the degradation of D1 protein under heat and high light stress. SA accelerates the recovery of D1 protein after termination of stress.Citation6
Besides a threatening rise in global temperature, climate change is also attributed to increases in atmospheric CO2 concentration, which may offer interactive effects of HS and elevated CO2 concentration on plant growth and productivity.Citation7 Previous studies indicate that elevated CO2 could minimize HS-induced deleterious effects on plants.Citation7,8 Nonetheless, the mechanisms of elevated CO2-induced HS mitigation are unknown. Previously, we have demonstrated that elevated CO2-induced HS mitigation does not involve abscisic acid-dependent process in tomato.Citation7 In the present study, we used a set of Arabidopsis mutants such as nonexpressor of pathogenesis-related gene 1 (npr1-1, npr1-5; SA signaling blockage mutant) and HSPs gene mutants (hsp21 and hsp70-1) to understand the requirement of SA signaling and HSPs in elevated CO2-induced HS tolerance. The performance of the photosynthetic machinery was monitored by chlorophyll a fluorescence through determining the photochemical efficiency of PSII (Fv/Fm) which is well recognized as a sensitive HS indicator.Citation7
Under normal temperature conditions, no obvious differences were noticed among the mutants and wild-type (Col-0) Arabidopsis plants in terms of Fv/Fm value (). After the heat shock (42°C, 24 h) in ambient CO2 (380 µmol mol−1) conditions, all groups of seedlings displayed a significant decline in Fv/Fm. HS decreased Fv/Fm by 29.11, 38.12, 37.80, 39.74 and 42.35% in Col-0, npr1-1, npr1-5, hsp21 and hsp70-1, respectively compared to their respective non-stress controls. Among all studied plant groups, hsp21 showed characteristically dwarf phenotype; however, it demonstrated similar heat sensitivity like other mutants. Administration of CO2 enrichment along with HS remarkably increased the Fv/Fm value in all plant groups except hsp70-1. CO2 enrichment increased Fv/Fm value by 33.61, 35.20, 34.16 and 38.87 % in Col-0, npr1-1, npr1-5 and hsp21, respectively compared with their respective only heat treatment counterparts. It can be explained that NPR1-1, NPR1-5 and HSP21 genes might not be involved in the elevated CO2-induced improvement in photosynthetic efficiency. By contrast, the data indicate that HSP70-1 is the most important component required for HS tolerance under both ambient and elevated CO2 concentration in Arabidopsis.
Figure 1. Effect of heat stress and elevated CO2 on maximum photochemical efficiency of PSII (Fv/Fm) in Arabidopsis. Chlorophyll a fluorescence imaging was used to determine photosynthetic performance of wild-type Col-0, SA signaling blockage mutants and hsp mutant lines. Four weeks old Arabidopsis plants (wild-type and mutants) were exposed to either ambient CO2 (Amb., 380 µmol mol−1) or elevated CO2 (Ele., 800 µmol mol−1) for 5 d and then challenged with high temperature (42°C) for 24 h under ambient or elevated CO2. Fv/Fm was measured after heat stress as previously described elsewhere.Citation7 (A) Pseudo-color images of Fv/Fm where the false color code depicts ranges vertically from 0.0 (black) to 1.0 (purple). (B) Fv/Fm values averaged over the whole rosette area; values presented are mean ± SD from 4 rosettes. The means denoted by the same letters did not significantly differ at P < 0.05 according to Tukey's test.
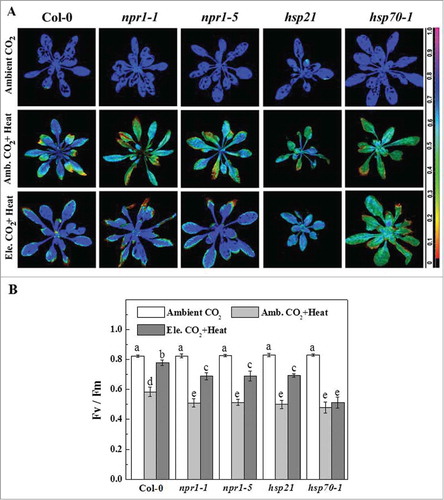
NPR1 is the key transducer of the SA signal. Previous studies showed that NPR1 is required for the basal thermotolerance which support our current observation on npr1-1 and npr1-5 under ambient CO2.Citation9 Interestingly, we found that elevated CO2 can still increase the thermotolerance of npr1 mutants, which indicates NPR1-independent process for HS mitigation in response to elevated CO2 (). Elevated CO2 might stimulate cellular redox homeostasis and MAPK activity and thus ameliorate HS through NPR1-independent process in npr1-1 and npr1-5 mutants.Citation7,8,10,11 HSPs protect various important proteins from irreversible heat-induced damage through preventing denaturation as well as assisting the refolding of damaged protein.Citation1,4,9 The inability of hsp21 and hsp70-1 to produce respective proteins aggravated HS-induced damage to photosynthetic apparatus (). Administration of elevated CO2 could protect photosynthetic apparatus of hsp21 mutants, but not of hsp70-1 mutants. Information on effects of elevated CO2 on HSPs production and accumulation is scanty. A recent study shows that elevated CO2-induced HSPs accumulation varies with the plant species as well as nutrient status (e.g. nitrogen) under HS.Citation12 In corn (Zea mays L.) with high nitrogen supply, elevated CO2-induced photosynthetic thermotolerance might be partly mediated by HSPs (such as HSP60 and HSP70).Citation12 Extreme thermosensitivity of hsp70-1 mutants and its inability to mitigate HS following CO2 enrichment suggest that elevated CO2-induced HS tolerance largely depends on HSP70-1 in Arabidopsis. However, for hsp21 mutants, other protective processes such as efficient reactive oxygen species (ROS) detoxification, improved redox homeostasis and decreased photorespiration as consequences of CO2 enrichment might occur.Citation8 Together, this work indicates that NPR1-mediated SA signaling is important for basal thermotolerance; however, NPR1 is not required for elevated CO2-induced HS mitigation in Arabidopsis. Elevated CO2 could ameliorate HS-induced deleterious effect on photosynthetic apparatus without involvement of HSP21, but basal thermotolerance as well as elevated CO2-induced HS tolerance largely depends on HSP70-1 in Arabidopsis. Further study using advanced physiological, biochemical and molecular-genetic approaches may unveil in-depth mechanisms of elevated CO2-induced HS mitigation and involvement of SA in this process.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We are grateful to Dr. Frantisek Baluska for kindly inviting this manuscript.
Funding
This work was supported by the National Key Technology R&D Program of China (2013AA102406), the National Natural Science Foundation of China (31372108), the China Postdoctoral Science Foundation (517000-X91414) and the Chinese Academy of Agricultural Sciences Innovation Project.
Related Research Data
References
- Mathur S, Agrawal D, Jajoo A. Photosynthesis: Response to high temperature stress. J Photochem Photobiol B 2014; 137:116-26; PMID:24796250; http://dx.doi.org/10.1016/j.jphotobiol.2014.01.010
- Yoshioka M, Uchida S, Mori H, Komayama K, Ohira S, Morita N, Nakanishi T, Yamamoto Y. Quality control of photosystem II - Cleavage of reaction center D1 protein in spinach thylakoids by FtsH protease under moderate heat stress. J Biol Chem 2006; 281:21660-9; PMID:16735503; http://dx.doi.org/10.1074/jbc.M602896200
- Clarke SM, Mur LAJ, Wood JE, Scott IM. Salicylic acid dependent signaling promotes basal thermotolerance but is not essential for acquired thermotolerance in Arabidopsis thaliana. Plant J 2004; 38:432-47; PMID:15086804; http://dx.doi.org/10.1111/j.1365-313X.2004.02054.x
- Barua D, Downs CA, Heckathorn SA. Variation in chloroplast small heat-shock protein function is a major determinant of variation in thermotolerance of photosynthetic electron transport among ecotypes of Chenopodium album. Funct Plant Biol 2003; 30:1071-9; http://dx.doi.org/10.1071/Fp03106
- Horvath E, Szalai G, Janda T. Induction of abiotic stress tolerance by salicylic acid signaling. J Plant Growth Regul 2007; 26: 290-300; http://dx.doi.org/10.1007/s00344-007-9017-4
- Zhao HJ, Zhao XJ, Ma PF, Wang YX, Hu WW, Li LH, Zhao YD. Effects of salicylic acid on protein kinase activity and chloroplast D1 protein degradation in wheat leaves subjected to heat and high light stress. Acta Ecol Sinica 2011; 31:259-63; http://dx.doi.org/doi.org/10.101/6j.chnaes.2011.06.006
- Li X, Ahammed GJ, Zhang YQ, Zhang GQ, Sun ZH, Zhou J, Zhou YH, Xia XJ, Yu JQ, Shi K. Carbon dioxide enrichment alleviates heat stress by improving cellular redox homeostasis through an ABA-independent process in tomato plants. Plant Biol 2014; 17:81-9; PMID:24985337; http://dx.doi.org/10.1111/plb.12211
- Zinta G, AbdElgawad H, Domagalska MA, Vergauwen L, Knapen D, Nijs I, Janssens IA, Beemster GTS, Asard, H. Physiological, biochemical, and genome-wide transcriptional analysis reveals that elevated CO2 mitigates the impact of combined heat wave and drought stress in Arabidopsis thaliana at multiple organizational levels. Global Change Biol 2014; 20:3670-85; PMID:24802996; http://dx.doi.org/10.1111/Gcb.12626
- Clarke SM, Cristescu SM, Miersch O, Harren FJM, Wasternack C, Mur LAJ. Jasmonates act with salicylic acid to confer basal thermotolerance in Arabidopsis thaliana. New Phytol 2009; 182:175-87; PMID:19140948; http://dx.doi.org/10.1111/j.1469-8137.2008.02735.x
- Kang GZ, Li GZ, Guo TC. Molecular mechanism of salicylic acid-induced abiotic stress tolerance in higher plants. Acta Physiol Plant 2014; 36:2287-97; http://dx.doi.org/10.1007/s11738-014-1603-z
- Niu YF, Jin GL, Chai RS, Wang H, Zhang YS. Responses of root hair development to elevated CO2. Plant Signal Behav 2011; 6:1414-7; PMID:21847031; http://dx.doi.org/10.4161/psb.6.9.17302
- Wang D, Heckathorn SA, Hamilton EW, Frantz J. Effects of CO2 on the tolerance of photosynthesis to heat stress can be affected by photosynthetic pathway and nitrogen. Am J Bot 2014; 101:34-4; PMID:24355208; http://dx.doi.org/10.3732/Ajb.1300267