
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Leon Chua, the discoverer of a memristor, theoretically predicted that voltage gated ion channels can be memristors. We recently found memristors in different plants such as the Venus flytrap, Mimosa pudica, Aloe vera, apple fruits, and in potato tubers. There are no publications in literature about the existence of memristors in seeds. The goal of this work was to discover if pumpkin seeds might have memristors. We selected Cucurbita pepo L., cv. Cinderella, Cucurbita maxima L. cv Warty Goblin, and Cucurbita maxima L., cv. Jarrahdale seeds for this analysis. In these seeds, we found the presence of resistors with memory. The analysis was based on cyclic voltammetry where a memristor should manifest itself as a nonlinear two-terminal electrical element, which exhibits a pinched hysteresis loop on a current-voltage plane for any bipolar cyclic voltage input signal. Dry dormant pumpkin seeds have very high electrical resistance without memristive properties. The electrostimulation by bipolar sinusoidal or triangular periodic waves induces electrical responses in imbibed pumpkin seeds with fingerprints of memristors. Tetraethylammonium chloride, an inhibitor of voltage gated K+ channels, transforms a memristor to a resistor in pumpkin seeds. NPPB (5-Nitro-2-(3-phenylpropylamino)benzoic acid) inhibits the memristive properties of imbibed pumpkin seeds. The discovery of memristors in pumpkin seeds creates a new direction in the understanding of electrophysiological phenomena in seeds.
Abbreviations:
| 9-AC | = | 9-Anthracenecarboxylic acid |
| C | = | capacitance |
| DAQ | = | data acquisition |
| PXI | = | PCI eXtensions for Instrumentation |
| I | = | electrical current |
| M | = | memristance |
| NPPB | = | 5-Nitro-2-(3-phenylpropylamino)benzoic acid |
| q | = | charge |
| R | = | resistance |
| TEACl | = | tetraethylammonium chloride |
| VFG | = | voltage of an function generator |
| VP | = | voltage between electrodes in plants |
| VR | = | voltage on resistor R |
| φ | = | magnetic flux |
Introduction
Chua postulated the existence of the fourth basic electrical circuit element, memristor, a resistor with memory, which is a non-linear passive two-terminal electrical element relating magnetic flux linkage and electrical charge.Citation1 A memristor is a nonlinear element because its current-voltage characteristic is similar to that of a Lissajous pattern.Citation2 Mathematically memristance M can be described by equation:(1)
(1) where φ and q denote the magnetic flux and charge, respectively.
Different memristors were developed as solid state semiconductor devices,Citation3 polymersCitation4 and modified electrodes.Citation5,6 Biological tissue in many organisms exhibits memristive behaviors.Citation2,7 Gale et al. found memristive properties of protoplasmic tubes in a cellular slime mold Physarum polycephalum.Citation8 Hota et al. created transparent memristors from natural regenerated silk fibroin protein obtained from cocoons of Bombyx mori silkworm.Citation9 Johnsen et al. found that the sweat ducts in the skin are memristors.Citation10 We have found memristors in the electrical circuitry of the Venus flytrap,Citation11,12 potato tubers,Citation13 apple fruits, Aloe veraCitation14 and Mimosa pudica.Citation15 Pharmaceutical analysis showed that the tetraethylammonium chloride (TEACl) blocker of K+ channels inhibits memristance of plants.Citation11-16 Uncouplers carbonylcyanide-4-trifluoromethoxyphenyl hydrazone (FCCP) and carbonylcyanide-3-chlorophenylhydrazone (CCCP) decreased the amplitude of hysteresis in I-V plane.Citation14 Markin et al. proposed analytical models of a generic memristor and a memristor connected in parallel to a capacitor for analysis of plant memristors, which describe experimental data in plants.Citation16
There are no publications in literature about the existence of memristors in seeds. We selected pumpkin seeds for our project because their size is large enough for insertion of electrodes. There are a few publications about enzymes in germinating pumpkin seedsCitation16-19 but there is no information about voltage gated ion channels in dormant or germinating pumpkin seeds. Electrophysiology and electrical signaling in pumpkin plants was analyzed by Vodeneev et al.Citation20,21
The main goal of this article is to find out if dormant or imbibed pumpkin seeds have memristing properties.
Results
Dormant seeds
Experimental setup for cyclic voltammetry is shown in . Bipolar sinusoidal or triangular periodic waves with varying amplitude VFG was applied from a function generator. To measure electrical current, we included an additional resistor R in the circuit, so that the electrical current through the pumpkin seeds was found as I = VR/R.
Figure 1. Experimental setup.
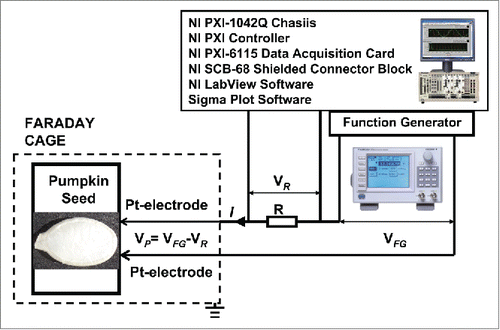
Cyclic voltammetry of dry dormant Cucurbita pepo L., cv. Cinderella seeds at different positions of platinum electrodes inserted in seeds through their coats is shown in . A bipolar sinusoidal wave with amplitude of ± 5.5 V was applied from a function generator to seeds through a resistor. Potential difference, VP, between electrodes in plants is equal to VP = VFG – VR (). The frequency of electrostimulation was 1 mHz (1000 scans/s, 1,000,000 scans). Similar results were obtained using dry dormant Cucurbita maxima L. cv Warty Goblin () and Cucurbita maxima L., cv. Jarrahdale () seeds. Dry seeds show very high resistance without memristive properties between electrodes inserted in pumpkin seeds. There is a strong electrical current increasing and resistance decreasing at voltages about ± 4 V (). This phenomenon is reversible and can be reproduced by multiple cycling of applied voltage. This effect of resistance decreasing can be caused by opening of alternative pathways for electrical current.
Figure 2. Cyclic voltammetry of dry dormant Cucurbita pepo L., cv Cinderella seeds at different orientation of electrodes in seeds. Bipolar sinusoidal wave with amplitude of ± 5.5 V was applied from a function generator. The frequency of electrostimulation was 1 mHz (1000 scans/s, 1,000,000 scans). Position of platinum electrodes inserted in a seed through its coat is shown.
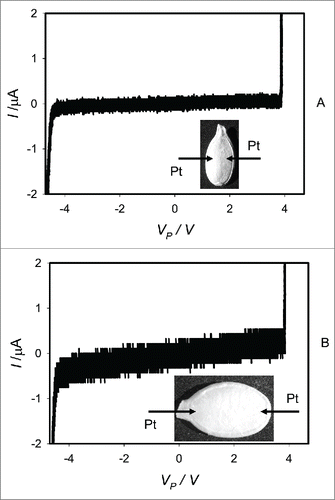
Figure 3. Cyclic voltammetry of dry dormant Cucurbita maxima L. cv. Warty Goblin (A) and Cucurbita maxima L., cv Jarrahdale (B) seeds. Bipolar sinusoidal wave with an amplitude of ± 5.5 V. The frequency of electrostimulation was 1 mHz (1000 scans/s, 1,000,000 scans). Position of platinum electrodes inserted in a seed through its coat is shown.
Imbibed and dormant seeds without coats
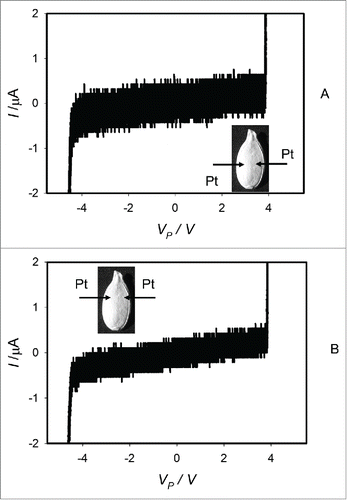
If pumpkin seeds without coats were soaked in water for 15–25 hours, the electrical properties of imbibed seeds undergo drastic changes (). Electrostimulation of imbibed pumpkin seeds by a bipolar sinusoidal or triangular wave induces a pinched hysteresis loop in the voltage-current plane at low frequencies of electrostimulation, which is a typical fingerprint of a memristor (). There is a pinched point without self-crossing in hysteresis loop at low frequency of bipolar sinusoidal wave in the voltage-current plane when VP = 0 V and I = 0 μA ().
Figure 4. Cyclic voltammetry of imbibed (A, B) and dry dormant (C) Cucurbita pepo L., cv. Cinderella seeds without coats. Bipolar sinusoidal waves with amplitude of ± 4.5 V (A) and ± 5.5 V (B, C) were applied from a function generator. The frequency of the electrostimulation was 1 mHz (1000 scans/s, 1,000,000 scans). Platinum electrodes were inserted directly in a seed.
Electrostimulation of dry dormant pumpkin seeds without coats by a bipolar sinusoidal or triangular wave does not induce a pinched hysteresis loop in the voltage-current plane at low frequencies of electrostimulation.().
Imbibed seeds with coats
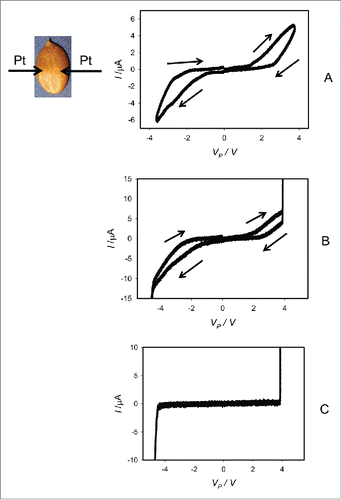
The imbibed seeds in their coats of Cucurbita pepo L., cv Cinderella (), Cucurbita maxima L., cv. Jarrahdale (), and Cucurbita maxima L. cv Warty Goblin show very similar memristive properties. Even after removing the pumpkin seeds' coats, the results remained the same (). These seeds have similar memristive properties as imbibed pumpkin seeds in coats.
Figure 5. Dependencies of electrical current, I, in imbibed Cucurbita pepo L., cv Cinderella seeds on VP induced by bipolar sinusoidal voltage wave with amplitude of ± 5.5 V from a function generator. Frequency of bipolar sinusoidal voltage VFG scanning was 1 mHz (A) or 50 μHz (B). Position of platinum electrodes inserted in a seed through its coat is shown.
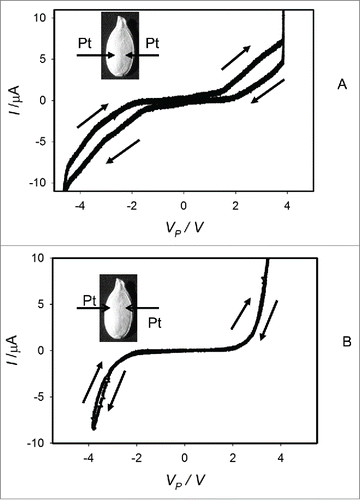
Figure 6. Dependencies of electrical current, I, in an imbibed Cucurbita maxima L., cv. Jarrahdale seed on VP induced by bipolar sinusoidal voltage wave with amplitude of ± 5.5 V from a function generator. Frequency of bipolar sinusoidal voltage VFG scanning was 1 mHz (A) or 100 μHz (B). Position of platinum electrodes inserted in a seed through its coat is shown.
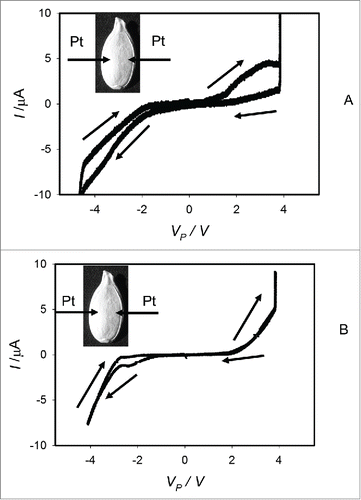
At low frequencies of cyclic voltammetry scanning of imbibed pumpkin seeds, there is a pinched hysteresis loop on a current-voltage plane for bipolar sinusoidal or triangular cyclic voltage input signal, where a memristor should manifest itself as a nonlinear two-terminal electrical element. The pinched hysteresis disappears at high frequencies (). The pinched hysteresis loop transforms to a non-pinched hysteresis loop instead of a single line I = VP//RP at high frequencies of the applied voltage because the amplitude of electrical current depends on capacitance of a seed tissue and electrodes, frequency and direction of scanning:
Figure 7. Dependencies of electrical current, I, in imbibed Cucurbita maxima L. cv Warty Goblin seeds on VP induced by bipolar sinusoidal voltage wave with amplitude of ± 5.5 V from a function generator. Frequency of bipolar sinusoidal voltage scanning was 0.1 mHz (A), 1 mHz (B) and 1 kHz (C). Position of platinum electrodes is shown in .
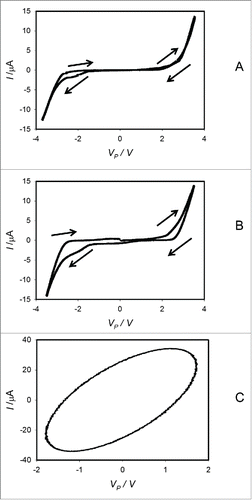
At very low frequencies, amplitude of a hysteresis decreases and disappears (). This effect was predicted in a theoretical analysis by Markin et al.Citation16
There is a strong electrical current increasing and resistance decreasing at voltages about ± 4 V () between platinum electrodes, which is similar to effects in dry seeds with coats () or without coats (). This effect of resistance decreasing can be caused by opening of alternative electrical circuits in wet or dry seeds.
Effect of ion channel inhibitors
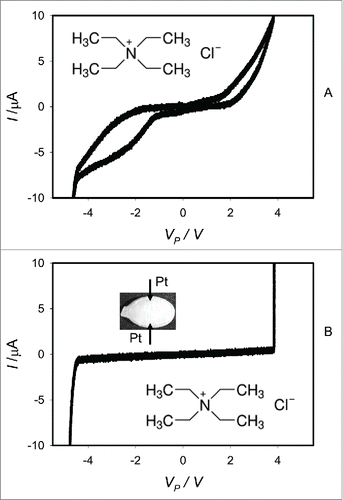
shows the effect of tetraethylammonium chloride (TEACl) on electrical responses induced by periodic bipolar sinusoidal waves from a function generator when platinum electrodes are inserted in a pumpkin seed. Ten μL of 0.3 M TEACl was injected in imbibed Cucurbita pepo L., cv. Cinderella seeds with coats 15 minutes () and 3 hours () before electrical measurements. There is a slow distribution and partition of TEACl in the seed tissue. TEACl decreases the amplitude of electrical current between electrodes in the imbibed pumpkin seed and the pinched hysteresis loop disappears (). Tetraethylammonium chloride, an inhibitor of voltage gated K+ channels, transforms a memristor to a conventional resistor in a seed tissue at low frequencies of a bipolar periodic sinusoidal or triangular wave. A blocker of the voltage gated potassium ion channels, TEACl inhibits the memristive properties of the wet Cucurbita pepo L., cv Cinderella seeds (). It means that the voltage gated potassium ion channels in these seeds can be memristors. However, TEACl is not specific and could also block some non-selective cation channels and aquaporins.
Figure 8. Electrical current I versus voltage VP applied to a imbibed Cucurbita pepo L., cv. Cinderella seeds. Frequency of bipolar sinusoidal voltage scanning was 1 mHz. 10 μL of 0.3 M TEACl was injected by a syringe to a seed (A) 15 min and (B) 3 hours before measurements. Position of platinum electrodes inserted in a seed through its coat is shown.
NPPB, 5-Nitro-2-(3-phenylpropylamino)benzoic acid is known as an inhibitor of chloride channels in plants, but it can also inhibit potassium channels.Citation21 Inhibitory effect of NPPB solution in ethanol on memristive properties of imbibed pumpkin seeds is shown in . This inhibitory effect is associated with injection of NPPB, because injection of ethanol into wet Cucurbita pepo L., cv. Cinderella seeds does not inhibit the memristive properties of imbibed pumpkin seeds ().
Figure 9. Dependencies of electrical current, I, in imbibed Cucurbita pepo L., cv Cinderella seeds on VP induced by bipolar sinusoidal voltage wave from a function generator. Frequency of bipolar sinusoidal voltage scanning was 1 mHz (1000 scans/s, 1,000,000 scans). (A): 5 μL of 20 mM NPPB solution in ethanol was injected to a seed 3 hours before cyclic voltammetry experiments. (B): 10 μL of ethanol was injected to a seed 3 hours before cyclic voltammetry experiments. Position of platinum electrodes inserted in a seed through its coat is shown.
Discussion
Dry dormant pumpkin seed do not have memristive properties (). Dormant state helps pumpkin seeds to conserve energy. Seed dormancy is caused by endogenous characteristics of the embryo.
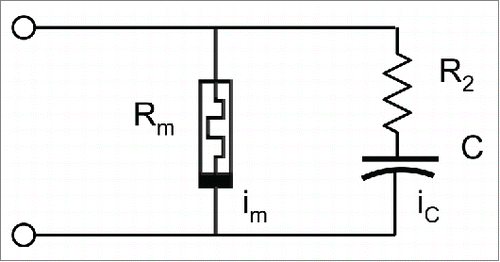
Initial absorption of water by seeds is a primary physical process of hydration and osmosis. The electrostimulation of wet pumpkin seeds by bipolar periodic sinusoidal or triangular waves induces electrical responses with fingerprints of memristors. To elucidate this mechanism, Markin et al.Citation16 proposed a simple analytical model of two types of memristors that can be found within plants. It consists of a memristor and a memristor connected in parallel to a capacitor and a resistor ().
Figure 10. Electrical circuit.
It was found previouslyCitation7,23 that the ideal memristor has 3 characteristic fingerprints:
When driven by a bipolar periodic signal, the device must exhibit a pinched hysteresis loop in the voltage-current plane, assuming the response is periodic.
Starting from some critical frequency, the hysteresis lobe area should decrease monotonically as the excitation frequency increases.
Pinched hysteresis loop should shrink to a single-valued function when the frequency tends to infinity.
However, the pinched hysteresis loop was experimentally observed in imbibed pumpkin seeds (), but it did not shrink to a single-valued function when the frequency is very high (). Similar properties were found earlier in the Venus flytrap, Mimosa pudica and Aloe vera.Citation11-16 It was explained by an obvious fact that the biological membranes with ion channels had capacitance connected in parallel to transmembrane organized ion channels. Such type of memristing devices with a capacitance is called a generic memristor.
The analytical model of biological generic memristors developed by Markin et al.Citation16 predicted additional fingerprint of a memristor connected in parallel to a capacitor and resistor. If frequency of a bipolar periodic signal is very low, the pinched hysteresis loop should shrink to a single-valued function. This prediction explains experimental results shown in.
So, the simple analytical model by Markin et al.Citation16 of generic memristors in plants successfully explained previous observations not only in plants, but also in pumpkin seeds. The basic property of memristor behavior is the fact that its resistance is voltage and time dependent, so when external voltage changes the resistance also changes but follows the voltage with some delay. The model considers a memristor M with resistance RM, which varies with potential difference VP(t) (). At constant VP, the resistance is constant but with varying voltage it follows with a certain delay time that explains the existence of hysteresis. So, this model explains the current observations in pumpkin seeds.
We selected Cucurbita pepo L., cv Cinderella, Cucurbita maxima L. cv. Warty Goblin, and Cucurbita maxima L., cv Jarrahdale seeds for this analysis. In these seeds, we found the presence of resistors with memory. When driven by a bipolar periodic sinusoidal or triangular signal, pumpkin seeds exhibit a pinched hysteresis loop in the voltage-current plane (). Starting from some critical frequency, the hysteresis loop changes shape and a pinched hysteresis loop transforms to a non-pinched hysteresis () as the excitation frequency increases. At very low frequencies, amplitude of a hysteresis decreases and disappears.()
Voltage gated ion channels regulate generation and transduction of electrical signals in plants.Citation24,25 For their analysis there are very efficient tools: blockers of ionic channels. It was intriguing to investigate if these blockers can change characteristics or even the presence of memristors in seeds. Tetraethylammonium chloride (TEACl) is known as a blocker of a voltage gated K+channel. We found that the injection of TEACl in wet pumpkin seeds induces contraction of the amplitude of electrical current and the shrinkage of the hysteresis (). Increasing resistance in wet seeds due to ion channels blocking can cause this result. Injection of distilled water in control experiments does not inhibit memristive properties of wet pumpkin seeds while an inhibitor of voltage gated K+ channels tetraethylammonium chloride transforms a memristor to a resistor.
We have to make a few comments about the shape of curves we have observed. The central part of these voltammograms happened to be somewhat flat. This detail makes it a little different from the predictions of the model in Ref.Citation16 Indeed the model in Ref. Sixteen was intentionally done very simplified to discover basic structures that could produce the very specific pinched shape of voltammograms. This goal was achieved: the model produced the curve that was pinched and there was no crossing between brunches. Of course the real biological tissue of seeds is much more complex. We believe that some additional effective-circuit elements might be present in seeds that can somewhat deform the observed curve. There are a few possibilities. One of them is the presence of some structures that might exhibit inductance. Adhikaru et al.Citation23 theoretically analyzed memristors with a second order RLC filter. Possible existence of inductance in plasma membranes was discussed by ColeCitation27 and by Hodgkin and Huxley.Citation28
There is another possibility for a flat pinched plateau. When the electrostimulation voltage is less than a threshold value, voltage gated channels can be closed and resistance of memristors will be very high. This effect will also generate a flat pinched plateau. Electrostimulation is a very efficient method for evaluation of biologically closed electrical circuits in seeds, fruits and plants. The information gained from electrostimulation can be used to elucidate and to observe electrochemical intracellular and intercellular communication in seeds.
Materials and methods
Pumpkins Cucurbita pepo L., cv. Cinderella, Cucurbita maxima L., cv Jarrahdale and Cucurbita maxima L. cv. Warty Goblin were received from Catbird Seat Garden Center (Madison, Alabama, USA). A few hundred seeds were removed from pumpkins, rinsed, and dried for seven d. All experiments were performed on healthy specimens. The humidity in laboratory was 40 – 43%.
Tetraethylammonium chloride (TEACl), 9-Anthracenecarboxylic acid (9-AC), 5-Nitro-2-(3-phenylpropylamino)benzoic acid (NPPB) and ethanol were obtained from Sigma-Aldrich (New York).
Platinum electrodes were used for electrostimulation of pumpkin seeds by bipolar sinusoidal or triangular periodic waves. Platinum electrodes are stable over a wide range of potentials, both in acidic and alkaline solutions, and in the presence of redox components.Citation26 Platinum electrodes were prepared from Teflon coated platinum wires (A-M Systems, Inc. Sequim, WA, USA) with a diameter of 0.076 mm.
All measurements were conducted in the laboratory at a constant room temperature of 21°С inside a Faraday cage, which was mounted on a vibration-stabilized table (). High speed data acquisition of low-pass filtered signals was performed using microcomputer NI-PXI-1042Q (National Instruments) with simultaneous multifunction I/O plug-in data acquisition board NI-PXI-6115 (National Instruments) interfaced through a NI SCB-68 shielded connector block to electrodes. The function generator FG300 (Yokagawa, Japan) was interfaced to the NI-PXI-1042Q microcomputer and used for electrostimulation of plants. We selected the resistor R = 45 kΩ for measurements of voltage, VR, and for an estimation of electrical current I.
All experimental results were reproduced at least 20 times. Software SigmaPlot 12 (Systat Software, Inc.) was used for statistical analysis of experimental data.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
This article is based on work supported by the Henry C. McBay (UNCF) Research Fellowship and by National Institute of Health under grant number 1R25GM106994-01.
References
- Chua L. Memristor – The missing circuit element. IEE Transactions Circuit Theory 1971; 18:507-19; http://dx.doi.org/10.1109/TCT.1971.1083337
- Chua L. If it's pinched it's a memristor. Semicond Sci Technol 2014; 29:104001-1-42; http://dx.doi.org/10.1088/0268-1242/29/10/104001
- Strukov DB, Snider GS, Stewart DR, Williams RS. The missing memristor found. Nature 2008; 453:80-3; PMID:18451858; http://dx.doi.org/10.1038/nature06932
- Smerieri A, Berzina T, Erokhin V, Fontana MP. Polymeric electrochemical elements for adaptive networks: Pulse mode. J Appl Phys 2008; 104:114513-8; http://dx.doi.org/10.1063/1.3033399
- MacVittie K, Katz E. Electrochemical systems with memimpedance properties. J Phys Chem C 2013; 117:24943-47; http://dx.doi.org/10.1021/jp409257v
- MacVittie K, Katz E. Self-powered electrochemical memristor based on a biofuel cell – towards memristors integrated with biocomputing systems. Chem Commun 2014; 50:4816-19; PMID:24687004; http://dx.doi.org/10.1039/c4cc01540a
- Sah M, Kim H, Chua L. Brains are made of memristors. IEEE Circuits Systems 2014; 14:12-36; http://dx.doi.org/10.1109/MCAS.2013.2296414
- Gale E, Adamatsky A, Costello BDL. Slime mold memristors BioNanoScience 2015; 5:1–8; http://dx.doi.org/10.1007/s12668-014-0156-3
- Hota MK, Bera MK, Kundu B, Kundu SC. A natural silk fibroin protein-based transparent bio-memristor. Adv Funct Mater 2012; 22:4493-99; http://dx.doi.org/10.1002/adfm.201200073
- Johnsen GK, Lutken CA, Martinsen OG, Grimnes S. Memristive model of electro-osmosis in skin. Phys Rev E Stat Nonlin Soft Matter Phys 2011; 83:031916; PMID:21517534; http://dx.doi.org/10.1103/PhysRevE.83.031916
- Volkov AG, Forde-Tucket V, Reedus J, Mitchell CM, Volkova MI, Markin VS, Chua L. Memristor in the Venus flytrap. Plant Signal Behav 2014; 9:e29204-1-12; PMID:25482796; http://dx.doi.org/10.4161/psb.29204
- Volkov AG, Reedus J, Mitchell CM, Tucket C, Forde-Tucket V, Volkova M, Markin VS, Chua L. Memristor in the electrical network of Aloe vera L. Plant Signal Behav 2014; 9:e29056-1-7; PMID: 25763487; http://dx.doi.org/10.4161/psb.29056
- Volkov AG, Nyasani RK, Blockmon AL, Volkova MI. Memristors: Memory elements in potato tubers. Plant Signal Behav 2015; 10:e1071750-1-7; PMID:26237427; http://dx.doi.org/10.1080/15592324.2015.1071750
- Volkov AG, Reedus J, Mitchell CM, Tucket C, Volkova M, Markin VS, Chua L. Memory elements in the electrical network of Mimosa pudica L. Plant Signal Behav 2014; 9:e982029-1-9; PMID:25482796; http://dx.doi.org/10.4161/15592324.2014.982029
- Volkov AG, Tucket C, Reedus J, Volkova M, Markin VS, Chua L. Memristors in plants. Plant Signal Behav 2014; 9:e28152-1-8; PMID:24556876; http://dx.doi.org/10.4161/psb.28152
- Markin VS, Volkov AG, Chua L. An analytical model of memristors in plants. Plant Signal Behav 2014; 9:e972887-1-9; PMID:25482769; http://dx.doi.org/10.4161/15592316.2014.972887
- Benech-Arnold RL, Sanchez RA (Eds). Handbook of Seed Physiology. Applications to Agriculture. New York: Food Products Press and the Haworth Reference Press; 2004
- Bewley JD, Black M. Seeds: Physiology of Development and Germination. New York: Plenum Press; 2012
- Maeshima M, Hara-Nishimura I, Takeuchi Y, Nishimura M. Accumulation of vacuolar H+-pyrophosphatase and H+-ATPase during reformation of the central vacuole in germinating pumpkin seeds. Plant Physiol 1994; 106:61-9; PMID:12232303; http://dx.doi.org/10.1104/pp.106.1.61
- Vodeneev VA, Akinchits EK, Orlova LA, Sukhov VS. The role of Ca2+, H+, and Cl− ions in generation of variation potential in pumpkin plants. Russ J Plant Physiol 2011; 58:974-81; http://dx.doi.org/10.1134/S1021443711050256
- Vodeneev VA, Opritov VA, Pyatygin SS. Reversible changes of extracellular pH during action potential generation in a higher plant Cucurbito pepo. Russ J Plant Physiol 2006; 53:481-7; http://dx.doi.org/10.1134/S102144370604008X
- Graichen F, Giles KR, Abell A, Garill A. Removal of the nitro and phenyl groups from NPPB decreases its inhibitory effect on cytoplasmic streaming in the alga Nitella hookeri. Biochem Cell Biol 2005; 83:133-9; PMID:15864322; http://dx.doi.org/10.1139/o05-019
- Adhikaru AP, Sah MPd, Kim H, Chua L. The fingerprints of memristor. IEEE Trans Circuits Systems 2013; 60:3008-21; http://dx.doi.org/10.1109/TCSI.2013.2256171
- Volkov AG, editor. Plant Electrophysiology. Methods and Cell Electrophysiology. Berlin: Springer; 2012
- Volkov AG, editor. Plant Electrophysiology. Signaling and Responses. Berlin: Springer; 2012
- Volkov AG, Deamer DW, Tanelian DI, Markin VS. Liquid Interfaces in Chemistry and Biology. New York: J. Wiley; 1998
- Cole KS. Membranes, Ions and Impulses. Berkeley: University of California Press; 1972
- Hodgkin AL, Huxley AF. A quantitative description of membrane current and its application to conduction and excitation in nerve. J Physiol 1952; 117:500-44; PMID:12991237; http://dx.doi.org/10.1113/jphysiol.1952.sp004764