
ABSTRACT
Dichogamy is generally thought to be a mechanism that prevents self-fertilization in flowering plants. This study aims to investigate the relationships between floral age and stigma receptivity, style length and pollen viability, and define how floral characters avoid self-pollination in a gynodioecious Chinese plant, Elsholtzia rugulosa. We assessed the relationships between flower age and style length, stigma receptivity, and pollen viability in E. rugulosa. This species produces 2 forms with plants bearing either hermaphrodite flowers (H) or female flowers (F). Corolla length in F flowers was shorter than the corolla length of H flowers and produced no pollen. H flowers were protandrous, pollen release of H flowers occurred before stigma receptivity. Stigma receptivity was significantly positively correlated with style length in both F flowers and H flowers. Pollen viability in H flowers declined significantly with floral age. Our results suggest that self-pollination in H flowers is likely reduced by dichogamy because stigma receptivity and pollen viability were effectively separated in time. However, because H inflorescences typically have multiple flowers open at the same time means that geitonogamous selfing is not avoided.
Introduction
The various breeding system exhibited by flowering plants are thought to favor outcrossing and reduce pollen-stigma interference.Citation1-7 Dichogamy is defined as the temporal separation of male and female functions.Citation2 In protandrous species, anther dehiscence occurs before the stigma is receptive, while in protogyny the reverse arrangement occurs. These arrangements have generally been thought to be adaptive mechanisms to prevent self-fertilization and reduce the physical interference between pollen dispersal and receipt within flowers.Citation8 Mechanisms preventing self-fertilization primarily increase female fitness, whereas mechanisms avoiding self-interference function to promote male fitness.Citation2
However, the effectiveness of dichogamy as a barrier to self-fertilization has been questioned.Citation9-12 Navarro (1997) concluded that the occurrence of self-pollination in the protandrous species Salvia verbenaca species probably depends on pollen germinability patterns and proposed that further study on stigma receptivity and pollen germination may be necessary to reveal mechanisms of self-compatibility. Munoz and Devesa (1987) have suggested that protandry can effectively prevent self-fertilization in Lavandula stoechas, but not in the genus Nepeta because pollen dispersal and stigma receptivity are effectively separated in time while self-fertilization is not eliminated within-inflorescence because of geitonogamy.Citation13,14 Outcrossing is obligate in female (F) flowers but selfing can occur in hermaphrodite (H) flowers assuming they are self-compatible. In addition, geitonogamous pollination is not prevented within or between inflorescences on a plant, when flowers are at different sexual phases.
Elsholtzia rugulosa is an important medicinal plant in China and as a result much is known about its chemistry.Citation15,16 In contrast, little is known about the expression of male and female traits during floral development in this gynodioecious species (Personal observation). The objective of the present study was to describe the pattern of male and female phases over the lifespan of E. rugulosa flowers. Specifically we examined the temporal relationships between flower age and style length, stigma receptivity, and pollen viability.
Materials and methods
Study species and site
Elsholtzia rugulosa is an herb to subshrub distributed in Yunnan, Guangxi, Sichuang, and Guizhou provinces in China. It grows to 150 cm in height in hilly grasslands and ruderal areas at altitudes of 1300-2800m. The small, white flowers are arranged in densely packed inflorescences and flower from October to December. The pollinator of Elsholtzia rugulosa is Apis cerana Fabricius (Apidae; Asian honey bee).Citation17,18 The study was conducted October 2013 to December 2014 on Ailao Mountain, Yunnan Province, China (N 25°02'29," E 99°51'27").
Floral characters
Sixty floral buds from 6 plants (10 buds from each of 3 H and 3 F plants) were observed for 6 days to record the timing of flower developmental events including stamen and style elongation, and anther dehiscence. The floral tube length of H flowers and F flowers (30 flowers from an additional 3 plants) was measured from the base of the flower to the top of the corolla (we used flowers that had been open for 6 hours). An additional 30 flowers were collected from 3 plants (30 flowers from each of 3 H and 3 F plants) and style length was measured at the following times from the start of anthesis: 6-, 12-, 24, 48-, 72-h.
Stigma receptivity in H and F flowers and pollen viability (H flowers) were assessed via the MTT (thiazolyl blue) method.Citation19 Stigmas of 30 H and 30 F flowers from 6 plants (10 flowers each per plant) per floral stage (6-, 12-, 24, 48-, 72-h-old flowers) were examined and pollen from the 10 flowers each time (repeated 3 times, total 30 flowers) was mixed and 15 samples from the mixture were measured at each floral stage (6-, 12-, 24, 48-, 72-h-old flowers from 6 plants). Stigma receptivity and pollen grains were scored as 0 when there was no stain and 1 when stained blue.
Data analysis
Student's t-test was used to compare the size of H and F flowers and post hoc (LSD) were used to analyze differences in style length, stigma receptivity, and pollen viability. Relationships among variables were evaluated using Pearson correlation coefficient. All analyses were performed using SPSS 16 (SPSS Company, Chicago, Illinois, USA), with measured variables presented as mean ± SE.
Results
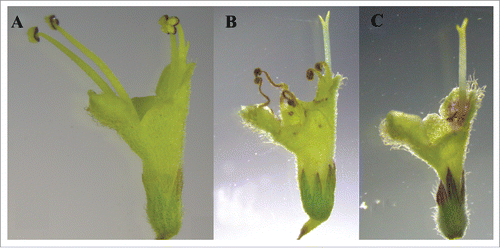
Elsholtzia rugulosa was found to be gynodioecious and produced 2 forms with plants bearing either hermaphrodite (H) or female flowers (F). The ratio of female plants to hermalphrodite plants was almost 1:1. The corolla of F flowers (2.95 ± 0.03 mm (n = 30)) was shorter than those of H flowers (4.20 ± 0.04 mm (n = 30)) (t = -23.16, P < 0.001). H flowers were protandrous and the H flower anthesis period started with the opening of the corolla tube when stamens were completely exserted from the corolla, and the style completely below the level of the anthers, avoiding contact with the anthers. The stigma branches of both H and F flowers were equal and closed. During this flowering phase, H flowers were entirely male and released pollen (). When reaching the female phase, the style elongated and the stigma branches opened with stamen senescence (). The flower lifespan (~96 hours) ended with wilting of the corolla and style. F flowers produced no pollen (). On average there were 614.9 ± 8.99 flowers (n = 30) on an inflorescence of Elsholtzia rugulosa. The lifespan of a single inflorescence was ~one month. Typically, both male- and female-phase flowers occurred simultaneously on a single inflorescence.
Figure 1. Hermaphrodite flowers (H) and female flowers (F) of Elsholtzia rugulosa. Male phase (A), Female phase (B), and F flowers (C).
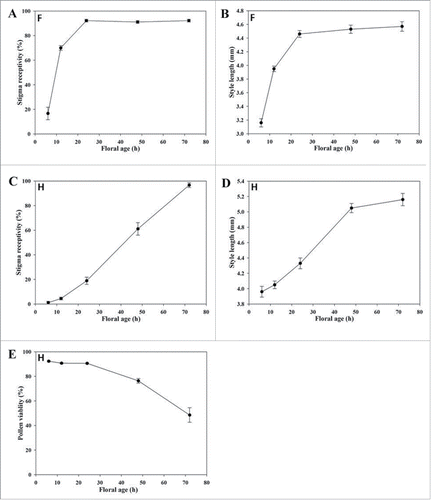
Style length increased with floral age in both H and F flowers, and although the significant increase of the style in F flowers occurred earlier than in H flowers (). Stigma receptivity increased constantly with H flower age following anthesis () before peaking on the third day of the flower' lifetime. Pollen viability declined with floral age. This decline was greater in the first 48h of flower lifetime (). The increase of style length with floral age was positive and significantly correlated with stigma receptivity (H flowers: r = 0.996, P = 0.008; F flowers: r = 0.988, P = 0.002). At the same time, stigma receptivity over the lifetime of the H flower was negative correlated with pollen viability (r = - 0.961, P = 0.009).
Figure 2. Style length, stigma receptivity, and pollen viability over the life span of 2 flower types in Elsholtzia rugulosa. F, female flower without pollen (male sterile); H, Hermaphrodite flower (male fertile).
Discussion
Dichogamy is frequently interpreted as a mechanism that prevents self-fertilization.Citation8,20 This study provides evidences that E. rugulosa flowers (a self-compatible species, seeds were produced by bagged control flowers, unpublished data) are protandrous in nature, and that dichogamy likely plays an important role in avoiding self pollination and pollen-stigma interference.
For protandrous self-compatible species, pollen viability was negatively correlated with stigma receptivity and floral age. This may prevent self-pollination.Citation5 Navarro (1997) showed that the relationship between floral age and pollen germinability was opposite of what would be expected, and in fact the time of greatest pollen germinability coincides with maximum stigma receptivity in protandrous Salvia verbenaca. This may be an adaptive mechanism to allow self-fertilization when cross-pollination fails. Rodriguez-Riano and Dafni (2007) pointed out that stigma receptivity was negatively correlated with pollen viability in Teucrium capitatum and Origaum syriacum. This could be an adaptation to avoid pollen-stigma interference in order to prevent self-fertilization.
In the case of protandrous E. rugulosa there are 2 problems: self-pollination and pollen-stigma interference. When pollen is presented, the stigma is not exposed or receptive, so it cannot receive pollen in an effective way. This may suggest that in order to avoid such interference, the style needs to surpass in length those flower characters likely to interfere with receipt of pollen by the stigma.Citation5 Our result showed that self-pollination in H flowers of E. rugulosa is avoided by dichogamy. Stigma receptivity and pollen viability were effectively separated in time, because anthesis occurred before stigma receptivity, while the occurrence of asynchronous dichogamy can not prevent self-pollination within-inflorescence and on the same plant.Citation2,13,21
In conclusion, the present study suggests that self-pollination in H flowers is avoided by dichogamy. Stigma receptivity and pollen viability were effectively separated in time because anthesis occurred before stigma receptivity. Further research on other members of the genus may provide interesting insights into the function of protandry to decrease selfing and interference.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We are very grateful to Dr. Madeline Carins Murphy for improving the English.
Funding
This work was financially supported by First Class Discipline Construction Project of Forestry in Yunnan Province (51600625).
References
- Charlesworth D. A further study of the problem of the maintenance of females in gynodioecious species. Heredity 1981; 46:27-39; http://dx. doi:10.1038/hdy.1981.3.
- Lloyd DG, Webb CJ. The avoidance of interference between the presentation of pollen and stigmas in angiosperms I. Dichogamy. New Zeal J Bot 1986; 24:135-62; http://dx.doi.org/10.1080/0028825X.1986.10409725.
- Bertin R I, Newman CM. Dichogamy in angiosperms. Bot Rev 1993; 59:112-52; http://dx.doi:10.1007/BF02856676.
- Barrett SCH, Harder LD, Cole WW. Correlated evolution of floral morphology and mating-type frequencies in a sexually polymorphic plant. Evolution 2004; 58:964-75; PMID:15212378; http://dx.doi.org/10.1554/03-417.
- Rodriguez-Riano T, Dafni A. Pollen-stigma interference in two gynodioecious species of Lamiaceae with intermediate individuals. Ann Bot 2007; 100:423-31; PMID:16877457; http://dx. doi.10.1093/aob/mcl168.
- Li XX, Zou Y, Xiao CL, Gituru RW, Guo YH, Yang CF. The differential contributions of herkogamy and dichogamy as mechanisms of avoiding self-interference in four self-incompatible Epimedium species. J Evolution Biol 2013; 26:1949-58; http://dx. doi. 10.1111/jeb.12193.
- Mamut J, Li B, Tan DY. Protogyny and delayed autonomous self-pollination in the desert herb Zygophyllum macropterum (Zygophyllaceae). J Syst Evol 2014; 52:75-83; http://dx.doi. 10.1111/jse.12029.
- Faegri K, Van der Pijl L. The principles of pollination ecology, 3rd ed. Pergamon, New York, New York, USA 1979.
- Muller H. The fertilisation of flowers. London 1883.
- Nyman Y. Pollination mechanisms in six Campanula species (Campanulaceae). P1ant Syst Evol 1992; 181:97-108; http://dx.doi:10.1007/BF00937589.
- Ortega-Olivencia A, Devesa JA. Sexual reproduction in some Scrophularia species (Scrophulariaceae) from the Iberian Peninsula and the Balearic Islands. P1ant Syst Evol 1993; 184:159-74; http://dx. doi.10.1007/BF00937433.
- Navarro L. Fruit-set and seed weight variation in Anthyllis vulneraria subsp. vulgaris (Fabaceae). P1ant Syst Evol 1996; 201:139-48; http://dx. doi:10.1007/BF00989057.
- Ubera JL, Vaiedes B. Revision del genero Nepeta (Labiatae) en la Peninsula Iberica e Islas Baleares. Lagascalia 1983; 12:3-8.
- Bhardwaj M, Eckert CG. Functional analysis of synchronous dichogamy in flowering rush, Butomus umbellatus. Am J Bot 2001; 88:2204-13; PMID:21669653.
- Wei QH, Ruan ZG, Tang ZM, Zheng HY. Studies on pharmacognosy of Bai National herb of Elshotzia rugulisa Helmsl. J Yunnan Univ Tradit Chin Med 2002; 25:14-7.
- Zhao Y, Li QC, Zhao Y, Chen YG. Studies on the constituents from the herb of Elshotzia rugulosa. Zhongguo Zhong Yao Za Zhi 2004; 29:1144-6.
- Zhang FP, Yang QY, Zhang SB. Dual Effect of Phenolic Nectar on Three Floral Visitors of Elsholtzia rugulosa (Lamiaceae) in SW China. PLoS ONE 2016a; 11:e0154381; http://dx.doi:10.1371/journal.pone.0154381.
- Zhang FP, Yang QY, Wang G, Zhang SB. Multiple functions of volatiles in flowers and leaves of Elsholtzia rugulosa (Lamiaceae) from southwestern China. Scientific Reports 2016b; 6:276161; http://dx.doi: 10.1038/srep27616.
- Rodriguez-Riano T, Dafni A. 2000 A new procedure to assess pollen viability. Sex Plant Repro 1996; 12:241-4; http://dx.doi:10.1007/s004970050008.
- Navarro L. Is the dichogamy of Salvia verbenace (Lamiaceae) an effective barrier to self-fertiliaztion? P1ant Syst Evol 1997; 207:111-7; http://dx.doi:10.1007/BF00985212.
- Proctor M, Yeo P, Lack A. The natural history of pollination. London: Harper Collins 1996.