
ABSTRACT
Free polyamine (PA) titers in plants may be regulated through reversible conjugate formation and/or through modulation of their synthesis, transport and degradation. PA signaling involves the well-acknowledged signaling molecule, nitric oxide (NO), which functions in diverse biological processes. Present investigations demonstrate the influence of salt stress (120 mM NaCl) and exogenous NO donor (250 µM Diethylenetriamine, DETA) on PA homeostasis of 2 d old, etiolated sunflower (Helianthus annuus L.) seedling cotyledons as a long-distance signaling response. Significantly enhanced intracellular spermine (Spm) accumulation was observed in seedling cotyledons under salt stress and in response to NO donor, the increase being more pronounced in seedlings treated with NO, evidently as a result of upregulation of the PA biosynthetic enzymes – arginine decarboxylase (ADC) and S-adenosylmethionine decarboxylase (SAMDC) – as revealed by Western blot and confocal imaging (CLSM). Moreover, salt stress induced the activity of polyamine oxidase (PAO), a PA catabolic enzyme, while NO lowered its activity in salt-stressed seedling cotyledons. NO, thus, appears to assist the seedlings in adapting to salt stress by positively regulating PA homeostasis through regulation of PA distribution between free, conjugated and bound forms, increased accumulation of PA biosynthetic enzymes and lowering the rate of PA catabolism.
Polyamines (PAs) are key regulators of an array of biological processes crucial for growth and development as well as for survival and defense against stressful environmental conditions.Citation1−Citation4 PAs constitute an integral component of plant defense responses to both biotic and abiotic stress conditions.Citation1,Citation3-Citation7 PA metabolic pathways are carefully regulated in plants and the dynamic balance between their synthesis and degradation is crucial in conferring these molecules the ability to combat stress-induced adversities. PAs are basic in nature and exist in polycationic forms at physiological pH due to the presence of two (diamine) or more (polyamine) amino groups, a property by virtue of which they are able to bind to and exert stabilizing effects on the negatively charged biological macromolecules, such as nucleic acids, phospholipids in the membranes, and proteins. Free PA titers in plants may be regulated through reversible conjugate formation and/or through modulation of their synthesis, transport and degradation. Out of several PAs present in plants, diamine putrescine [Put, NH2(CH2)4NH2], triamine spermidine [Spd, NH2(CH2)3NH(CH2)4NH2 and tetraamine spermine [Spm, NH2(CH2)3NH(CH2)4NH(CH2)3NH2] account for the major components of cellular PA pool, and they can be present in free, conjugated and bound forms. Cytoplasmic levels of PAs are also modulated by compartmentalization within different organelles, conjugation and/or interaction with cell wall. The conjugated forms are mostly found associated with hydroxycinnamic acids. PAs bound to various biomolecules, such as nucleic acids and proteins (bound forms), constitute a significant portion of cellular PA pool in plants and may serve as storage forms of PAs, from which free forms may be released when required.Citation8 Therefore, an equilibrium is maintained between the free and bound PAs. A growing body of evidence suggests that conjugated PAs are essential for the development and biotic interactions and in stress responses, regulating free PA titers and detoxification of phenolic compounds.Citation8-Citation10 Two main pathways of PA biosynthesis exist in plants wherein Put serves as the primer moiety for the production of higher PAs (Spd and Spm). Put can be produced either from ornithine in a reaction catalyzed by ornithine decarboxylase (ODC; EC 4.1.1.17) or from arginine via agmantine in an alternate multi-step pathway involving arginine decarboxylase (ADC; EC 4.1.1.19). Spd and Spm are synthesized from Put by sequential transfer of aminopropyl moieties from decarboxylated S-adenosylmethionine (dcSAM) produced through the action of S-adenosylmethionine decarboxylase (SAMDC; EC 4.1.1.50) on S-adenosylmethionine (SAM).Citation11,Citation12 PA degradation is mediated by a heterogeneous class of enzymes – amine oxidases (AOs). These include diamine oxidases/copper-containing amine oxidases (DAOs/CuAOs) and Flavin-containing (FAD-dependent) polyamine oxidases (PAOs) which catalyze the oxidative deamination of diamines and polyamines at the primary and secondary amino groups, respectively, to generate H2O2, pyrrolidone and respective aminoaldehyde. A precise regulation of PA biosynthesis and PA catabolism is important in the maintenance of cellular PA homeostasis.Citation12,Citation13 This dynamic regulation of cellular PA levels is important for eliciting responses to specific developmental or environmental cues, including stress, through a complex interplay among signaling molecules such as H2O2, NO, CaCitation2+ and ABA, which have been revealed to be essential components of PA signaling.Citation14 NO, a well-known signaling molecule, implicated in a wide array of biological processes and stress responses, is a component of PA signaling.Citation14,Citation15 The objective of the present investigations was to examine the modulation of the expression/activity of the PA biosynthetic (ODC, ADC and SAMDC) and catabolic enzymes (DAO and PAO) by salt stress (120 mM NaCl), and the impact of NO provided as NO donor (DETA, 250 µM) as long-distance signaling response in dark-grown, 2 dold sunflower (Helianthus annuus L.) seedling cotyledons. The concentrations of NaCl and DETA have been selected on the basis of already established and recently published work from the author’s laboratory.Citation16,Citation17
Salt stress and NO enhance Spm accumulation and abundance of PA biosynthetic enzymes, and differentially regulate PA catabolism
Spm was the major form of PA in cotyledons obtained from 2 d-old dark-grown sunflower seedlings raised in the absence or presence of salt (120 mM NaCl) or 250 µM of NO donor, DETA in the absence or presence of 120 mM NaCl (). Salt stress is reported to induce Spm accumulation with a simultaneous decrease in the lower forms of PAs, Put and Spd or Put alone.Citation18-Citation21 Significant increment in the conjugated as well as bound forms of Spm, represented by supernatant hydrolysate and pellet hydrolysate fractions, respectively, was evident in response to salt stress. NO donor application significantly increased the production of Spm. However, it did not alter the level of free Spm (supernatant fraction) but enhanced the proportion of conjugated and bound Spm, irrespective of salt stress. Hence, a positive regulation of the PA metabolism by NO was revealed. Spm, in particular, has been proposed to play versatile roles in plant responses to stress.Citation22,Citation23 It has been proposed that the presence of two additional primary amino groups (-NH2) in Spm as compared to Put endows it with a greater, more efficient protective capacity.Citation24 Spm has been suggested to act as a signaling molecule in the cascade leading to activation of the antioxidant enzymes. Another important role ascribed to Spm is the protection of DNA from free radical attack since it is present in nucleus in large quantities.Citation22 It has also been implicated in plant defense responses against biotic threats by triggering defense-related genes and causing hypersensitive response (HR). Thus, increased accumulation of Spm in seedlings reported in the present work suggests its predominant protective role in counteracting salt stress in sunflower seedlings. Synergistic interplay between NO and PAs confers tolerance to abiotic as well as biotic stresses.Citation25-Citation28 NO promotes an increase in contents of PAs and protective metabolites, like γ-aminobutyric acid (GABA) and proline.Citation27,Citation29,Citation30 Increase in the conjugated form of Spm in response to NO could possibly strengthen the antioxidant machinery of plant cells. Based on earlier reports, conjugation of PAs to phenolic acids could serve as an effective non-enzymatic antioxidant mechanism and, thus, improve the cellular antioxidant capacity.Citation8,Citation31,Citation32 NO also promoted an increase in the insoluble bound fraction of Spm under both saline and non-saline conditions. However, this increment in insoluble Spm fraction was more pronounced in seedlings subjected to NO donor in the absence of salt. It implies that NO is likely promoting PAs to interact with biological macromolecules. PAs, when bound to these macromolecules, exert stabilizing effects on them, protect them from ROS attack and maintain their structural and functional integrity.Citation4,Citation8 Thus, a possible protective role for NO in the protection of macromolecules is suggested. Bound PAs also serve as storage forms of PAs from where they can be released in a regulated manner whenever needed by the cell or upon encountering specific conditions.Citation8 Altogether, these findings indicate that NO may facilitate de novo synthesis of Spm (by facilitating the conversion from Put) and promote its conjugation to phenolic acids as well as biological macromolecules and is likely to aid the plant to cope with the stressful conditions and minimize the salt-induced damage. These observations, however, need further investigations.
Figure 1. (a,b) Modulation of spermine content in various fractions obtained from sunflower seedling (2 d old) cotyledons (as estimated post-dansylation followed by quantification of TLC fractions by spectrofluorometry). Statistical significance of changes was analyzed by one-way ANOVA using SPSS 22.0 and changes were found to be significant (*P < .05, **P < .001, ***P < .0001) in comparison to the control. Data represent mean values from triplicates. (c–f) Modulation of expression of PA biosynthetic enzymes [ADC (Arginine decarboxylase), ODC (Ornithine decarboxylase) and SAMDC (S-adenosylmethionine decarboxylase)] in seedling cotyledons by NaCl (120 mM) and NO donor (DETA, 250 μM) application.
![Figure 1. (a,b) Modulation of spermine content in various fractions obtained from sunflower seedling (2 d old) cotyledons (as estimated post-dansylation followed by quantification of TLC fractions by spectrofluorometry). Statistical significance of changes was analyzed by one-way ANOVA using SPSS 22.0 and changes were found to be significant (*P < .05, **P < .001, ***P < .0001) in comparison to the control. Data represent mean values from triplicates. (c–f) Modulation of expression of PA biosynthetic enzymes [ADC (Arginine decarboxylase), ODC (Ornithine decarboxylase) and SAMDC (S-adenosylmethionine decarboxylase)] in seedling cotyledons by NaCl (120 mM) and NO donor (DETA, 250 μM) application.](/cms/asset/e2cd5ade-0168-4c8b-83d5-36da6fef7e1a/kpsb_a_1667730_f0001_oc.jpg)
Accumulating evidence report upregulation of most of the genes encoding PA biosynthetic enzymes, including Arginine Decarboxylase (ADC), Spermidine Synthase (SPDS), Spermine synthase (SPMS) and S-adenosylmethionine Decarboxylase (SAMDC) under various stress conditions, although the timing and degree of induction may differ in different species.Citation33,Citation34 Salt-induced upregulation of the genes of PA metabolic pathway enzymes has been well-documented.Citation1,Citation20,Citation35,Citation36 Transgenic plants engineered to overexpress or those introduced with genes of enzymes catalyzing PA biosynthesis exhibit enhanced tolerance to both biotic and abiotic stresses and as well as improved yield.Citation6,Citation37-Citation39 Elevated PA contents in the plants are attributed in part to their increased de novo synthesis as a result of enhanced accumulation of PA biosynthetic enzymes ODC, ADC and SAMDC. Same is evident from the present investigations (Western blot analysis and CLSM imaging), both under salt-stress and in response to NO application, with effect of NO being more pronounced ( and ). Salt stress enhanced the abundance of ADC and SAMDC, the key enzymes of PA biosynthesis. No noteworthy influence was observed on ODC. NO significantly increased the accumulation of enzymes ADC and SAMDC both under control and saline conditions. The expression of these enzymes is, thus, positively affected by NaCl stress as well as NO.
Figure 2. Immunolocalization of the three PA biosynthetic enzymes ADC, ODC and SAMDC using CLSM. (a) Immunolocalization of the three PA biosynthetic enzymes was performed by CLSM imaging of 7 µm thick transverse sections of cotyledons harvested from 2 d-old-dark-grown seedlings raised in the absence or presence of salt stress (120 mM NaCl) or those raised in Hoagland nutrient medium supplemented with 250 µM of DETA in the absence or presence of 120 mM NaCl. (b–d) Quantification of fluorescence units for ADC, ODC and SAMDC, respectively. Statistical significance of changes was analyzed by one-way ANOVA using SPSS 22.0 and changes were found to be significant (*P < .05, **P < .001) in comparison to control. Data represent mean values from triplicates [Magnification: 200×; Scale bar: 60 µm].
![Figure 2. Immunolocalization of the three PA biosynthetic enzymes ADC, ODC and SAMDC using CLSM. (a) Immunolocalization of the three PA biosynthetic enzymes was performed by CLSM imaging of 7 µm thick transverse sections of cotyledons harvested from 2 d-old-dark-grown seedlings raised in the absence or presence of salt stress (120 mM NaCl) or those raised in Hoagland nutrient medium supplemented with 250 µM of DETA in the absence or presence of 120 mM NaCl. (b–d) Quantification of fluorescence units for ADC, ODC and SAMDC, respectively. Statistical significance of changes was analyzed by one-way ANOVA using SPSS 22.0 and changes were found to be significant (*P < .05, **P < .001) in comparison to control. Data represent mean values from triplicates [Magnification: 200×; Scale bar: 60 µm].](/cms/asset/448ec04c-9041-4378-928c-bcb909a12071/kpsb_a_1667730_f0002_oc.jpg)
The method of estimation of PA catabolic enzyme activities was based on the estimation of H2O2 produced.Citation40 No significant modulation of PA catabolic enzyme DAO was observed by 120 mM NaCl stress or NO donor in terms of its activity (). In contrast, significant increase in PAO activity, another PA catabolic enzyme, was revealed both under salt stress and in response to NO treatment. DETA application, however, reduced the salt-induced increase in enzyme activity, which possibly helps in maintaining PA levels in the tissue by retarding their catabolic rate. This suggested the regulatory role played by PAO in PA catabolism and, therefore, overall PA homeostasis, in response to both salt stress and NO. Salt stress-induced PAO activity has been reported to be important in regulating salt tolerance in plants.Citation41,Citation42 Moreover, loss of PAO activity induces tolerance to salt stress.Citation43,Citation44 Irrespective of the pathways involved, oxidative catabolism of PAs by DAO and PAO is crucial in PA-mediated stress tolerance as it leads to the generation of H2O2, a signaling molecule which regulates downstream components in signal transduction pathways leading to triggering of various stress responses and GABA which serves as an osmoprotectant and has important protective roles in plant growth.Citation45,Citation46
Table 1. Alteration in the activity of the two polyamine catabolic enzymes, diamine oxidase (DAO) and polyamine oxidase (PAO) in sunflower seedling cotyledons in response to salt stress and treatment with NO donor (DETA). Data represent mean values from triplicates and the standard errors. Statistical significance of changes was analyzed by one-way ANOVA using SPSS 22.0 and changes were found to be significant (*P < .05, **P < .001) in comparison with control.
Conclusions
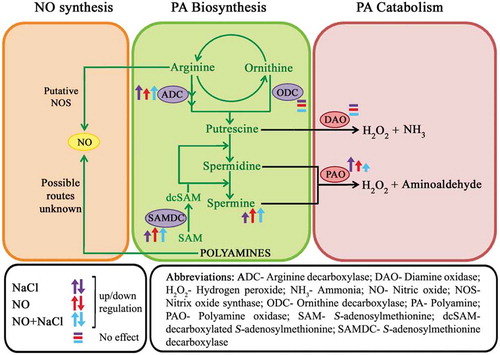
Maintenance of balance between PA biosynthesis and catabolism and their careful regulation by salt stress and NO appear to be crucial in acclimatization to salt stress (). Salt stress leads to elevated PA catabolic enzyme (PAO) activity. NO reduces this salt-induced increase in the enzyme activity and thus seems to assist plants in adapting to salt stress by positively regulating overall intracellular PA homeostasis by increasing the abundance of PA biosynthetic enzymes (especially ADC and SAMDC) and lowering the rate of PA catabolism by PAO. NO also regulates spatial distribution of Spm in affected cells. The exact mechanism underlying the modulation of PA metabolic pathways by NO, however, needs to be further investigated in order to know the precise means by which it participates in PA signaling routes, particularly under salt stress.
Figure 3. Nitric oxide and salt stress-mediated modulation of polyamine biosynthetic and catabolic routes.
Abbreviations
Acknowledgments
AT is grateful to University Grants Commission (UGC) for Research Fellowship awarded to her vide sanction no. 2061530546; Ref. No. 21/06/2015(i)EU-V dated 27 July 27 2016. Funding source for the present work is UGC-ISF Joint Research Grant [Grant No. F. No. 6-9/2017 (IC)].
Additional information
Funding
References
- Alcázar R, Altabella T, Marco F, Bortolotti C, Reymond M, Knocz C, Carrasco P, Tiburcio AF. Polyamines: molecules with regulatory functions in plant abiotic stress tolerance. Planta. 2010;231:1–6. PMID:20221631. doi:10.1007/s00425-010-1130-0.
- Tiburcio A, Altabella T, Bitrián M, Alcázar R. The roles of polyamines during the lifespan of plants: from development to stress. Planta. 2014;240:1–18. PMID:24659098. doi:10.1007/s00425-014-2055-9.
- Alcázar R, Tiburcio AF. Polyamine metabolism and abiotic stress tolerance in plants. In: Ramakrishna A, Gill SS, editors. Metabolic adaptations in plants during abiotic stress. CRC Press; 2018. p. 203–212. doi:10.1201/b22206.
- Chen D, Shao Q, Yin L, Younis A, Zheng B. Polyamine function in plants: metabolism, regulation on development, and roles in abiotic stress responses. Front Plant Sci. 2019;9:1945. PMID:30687350. doi:10.3389/fpls.2018.01945.
- Groppa MD, Benavides MP. Polyamines and abiotic stress: recent advances. Amino Acids. 2008;34:35–45. PMID:17356805. doi:10.1007/s00726-007-0501-8.
- Hussain SS, Ali M, Ahmad M, Siddique KH. Polyamines: natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol Adv. 2011;29:300–311. PMID:21241790. doi:10.1016/j.biotechadv.2011.01.003.
- Minocha R, Majumdar R, Minocha SC. Polyamines and abiotic stress in plants: a complex relationship. Front Plant Sci. 2014;5:175. PMID:24847338. doi:10.3389/fpls.2014.00175.
- Martin-Tanguy J. Metabolism and function of polyamines in plants: recent development (new approaches). Plant Growth Regul. 2001;34:135–148. doi:10.1023/A:1013343106574.
- Edreva AM, Velikova VB, Tsonev TD. Phenylamides in plants. Russ J Plant Physiol. 2007;54:287–301. doi:10.1134/S1021443707030016.
- Bassard J-E, Ullmann P, Bernier F, Werck-Reichhart D. Phenolamides: bridging polyamines to the phenolic metabolism. Phytochemistry. 2010;71:1808–1824. PMID:20800856. doi:10.1016/j.phytochem.2010.08.003.
- Kusano T, Berberich T, Tateda C, Takahashi Y. Polyamines: essential factors for growth and survival. Planta. 2008;228:367–381. PMID:18594857. doi:10.1007/s00425-008-0772-7.
- Wuddineh W, Minocha R, Minocha SC. Polyamines in the context of metabolic networks. Methods Mol Biol. 2018;1694:1–23. PMID:29080151. doi:10.1007/978-1-4939-7398-9_1.
- Pál M, Szalai G, Janda T. Speculation: polyamines are important in abiotic stress signaling. Plant Sci. 2015;237:16–23. PMID:26089148. doi:10.1016/j.plantsci.2015.05.003.
- Kusano T, Sagor GHM, Berberich T. Molecules for sensing polyamines and transducing their action in plants. Methods Mol Biol. 2018;1694:25–35. PMID:29080152. doi:10.1007/978-1-4939-7398-9_2.
- Nabi RBS, Tayade R, Hussain A, Kulkarni KP, Imran QM, Mun B-G, Yun B-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ Exp Bot. 2019;161:120–133. doi:10.1016/j.envexpbot.2019.02.003.
- David A, Yadav S, Bhatla SC. Sodium chloride stress induces nitric oxide accumulation in root tips and oil body surface accompanying slower oleosin degradation in sunflower seedlings. Physiol Plant. 2010;140:342–354. doi:10.1111/j.1399-3054.2010.01408.x.
- Singh N, Bhatla SC. Signaling through reactive oxygen and nitrogen species is differentially modulated in sunflower seedling root and cotyledon in response to various nitric oxide donors and scavengers. Plant Signal Behav. 2017;12:e1365214. PMID:28862537. doi:10.1080/15592324.2017.1365214.
- Maiale S, Sánchez DH, Guirado A, Vidal A, Ruiz OA. Spermine accumulation under salt stress. J Plant Physiol. 2004;161:35–42. PMID:15002662. doi:10.1078/0176-1617-01167.
- Roychoudhary A, Basu S, Sengupta DN. Amelioration of salinity stress by exogenously applied spermidine or spermine in three varities of indica rice differing in their level of salt tolerance. J Plant Physiol. 2011;168:317–328. PMID:20728960. doi:10.1016/j.jplph.2010.07.009.
- Do PT, Drechsel O, Heyer AG, Hincha DK, Zuther E. Changes in free polyamine levels, expression of polyamine biosynthesis genes, and performance of rice cultivars under salt stress: a comparison with responses to drought. Front Plant Sci. 2014;5:182. PMID:24847340. doi:10.3389/fpls.2014.00182.
- Li S, Jin H, Zhang Q. The effect of exogenous spermidine concentration on polyamine metabolism and salt tolerance in Zoysiagrass (Zoysia japonica Steud) subjected to short-term salinity stress. Front Plant Sci. 2016;7:1221. PMID:27582752. doi:10.3389/fpls.2016.01221.
- Takahashi T, Kakehi J. Polyamines: ubiquitous polycations with unique roles in growth and stress responses. Ann Bot. 2010;105:1–6. PMID:19828463. doi:10.1093/aob/mcp259.
- Seifi HS, Shelp BJ. Spermine differentially refines plant defense responses against biotic and abiotic stresses. Front Plant Sci. 2019;10:117. PMID:30800140. doi:10.3389/fpls.2019.00117.
- Liu JH, Wang W, Wu H, Gong X, Moriguchi T. Polyamines function in stress tolerance: from synthesis to regulation. Front Plant Sci. 2015;6:827. PMID:26528300. doi:10.3389/fpls.2015.00827.
- Tun NN, Santa-Catarina C, Begum T, Silveria V, Handro W, Floh EI, Scherer GFE. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006;47:346–354. PMID:16415068. doi:10.1093/pcp/pci252.
- Wimalasekera R, Tebartz F, Scherer GFE. Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011;181:593–603. PMID:21893256. doi:10.1016/j.plantsci.2011.04.002.
- Fan H-F, Dua C-X, Guo S-R. Nitric oxide enhances salt tolerance in cucumber seedlings by regulating free polyamine content. Environ Exper Bot. 2017;86:52–59. doi:10.1016/j.envexpbot.2010.09.007.
- Agurla S, Gayatri G, Raghavendra AS. Polyamines increase nitric oxide and reactive oxygen species in guard cells of Arabidopsis thaliana during stomatal closure. Protoplasma. 2018;255:153–162. PMID:28699025. doi:10.1007/s00709-017-1139-3.
- Wang H, Huang J, Liang W, Liang X, Bi Y. Involvement of putrescine and nitric oxide in aluminum tolerance by modulating citrate secretion from roots of red kidney bean. Plant Soil. 2013;366:479–490. doi:10.1007/s11104-012-1447-5.
- Wang Y, Luo Z, Maoa L, Ying T. Contribution of polyamines metabolism and GABA shunt to chilling tolerance induced by nitric oxide in cold-stored banana fruit. Food Chem. 2016;197:333–339. doi:10.1016/j.foodchem.2015.10.118.
- Yamasaki H, Cohen MF. NO signal at the crossroads: polyamine-induced nitric oxide synthesis in plants? Trends Plant Sci. 2006;11:522–524. PMID:17035070. doi:10.1016/j.tplants.2006.09.009.
- Choi SW, Lee SK, Kim EO, Oh JH, Yoon KS, Parris N, Hicks KB, Moreau RA. Antioxidant and antimelanogenic activities of polyamine conjugates from corn bran and related hydroxycinnamic acids. J Agric Food Chem. 2007;55:3920−5. PMID:17397179. doi:10.1021/jf0635154.
- Liu JH, Inoue H, Moriguchi T. Salt stress-mediated changes in free polyamine titers and expression of genes responsible for polyamine biosynthesis of apple in vitro shoots. Environ Exp Bot. 2008;62:28–35. doi:10.1016/j.envexpbot.2007.07.002.
- Liu JH, Nakajima I, Moriguchi T. Effects of salt and osmotic stresses on free polyamine content and expression of polyamine biosynthetic genes in Vitis vinifera. Biol Plant. 2011;55:340–344. doi:10.1007/s10535-011-0050-6.
- Romero FM, Maiale AJ, Rossi FR, Marina M, Ruíz OA, Gárriz A. Polyamine metabolism responses to biotic and abiotic stress. Methods Mol Biol. 2018;1694:37–49. PMID:29080153. doi:10.1007/978-1-4939-7398-9_3.
- Singh P, Basu S, Kumar G. Polyamines metabolism: a way ahead for abiotic atress tolerance in crop plants. In: SH W, editor. Biochemical, physiological and molecular avenues for combating abiotic stress in plants. 1st ed. Academic Press; 2018. p. 39–55. doi:10.1016/B978-0-12-813066-7.00003-6.
- Gill SS, Tuteja N. Polyamines and abiotic stress tolerance in plants. Plant Signal Behav. 2010;5:26–33. PMID:20592804. doi:10.4161/psb.5.1.10291.
- Marco F, Bitrián M, Carrasco P, Alcázar R, Tiburcio AF. Polyamine biosynthesis engineering as a tool to improve plant resistance to abiotic stress. In: Jaiwal PK, Singh RP, Dhankher OP, editors. Genetic manipulation in plants for mitigation of climate change. Cham: Springer; 2015. p. 103–116. doi:10.1007/978-81-322-2662-8_5.
- Khare T, Srivastav A, Shaikh S, Kumar V. Polyamines and their metabolic engineering for plant salinity stress tolerance. In: Kumar V, Wani S, Suprasanna P, Tran LS, editors. Salinity responses and tolerance in plants. Vol. 1. Cham: Springer; 2018. p. 339–358. doi:10.1007/978-3-319-75671-4_13.
- Matsumoto T, Nimura Y, Furuta T, Hayakawa N, Asai M, Kurokawa Y, Hattori T, Iyomasa Y. Some properties of amine oxidase from soybean seedlings. Nagoya J Med Sci. 1984;46:87–94. PMID: 6539854.
- Quinet M, Ndayiragije A, Lefèvre I, Lambillotte B, Dupont-Gillain CC, Lutts S. Putrescine differently influences the effect of salt stress on polyamine metabolism and ethylene synthesis in rice cultivars differing in salt resistance. J Exp Bot. 2010;61:2719–2733. PMID:20472577. doi:10.1093/jxb/erq118.
- Islam MA, Pang J-H, Meng F-W, Li Y-W, Xu N, Yang C, Liu J. Putrescine, spermidine, and spermine play distinct roles in rice salt tolerance. JIA. 2019;18:2–14. doi:10.1016/S2095-3119(19)62705-X.
- Sagor GHM, Zhang S, Kojima S, Simm S, Berberich T, Kusano T. Reducing cytoplasmic polyamine oxidase activity in Arabidopsis increases salt and drought tolerance by reducing reactive oxygen species production and increasing defense gene expression. Front Plant Sci. 2016;7:214. PMID:26973665. doi:10.3389/fpls.2016.00214.
- Zarza X, Atanasov KE, Marco F, Arbona V, Carrasco P, Kopka J, Fotopoulos V, Munnik T, Gomez-Cadenas A, Tiburcio AF, et al. Polyamine oxidase 5 loss-of-function mutations in Arabidopsis thaliana trigger metabolic and transcriptional reprogramming and promote salt stress tolerance. Plant Cell Environ. 2017;40:527–542. PMID:26791972. doi:10.1111/pce.12714.
- Tavladoraki P, Cona A, Angelini R. Copper-containing amine oxidases and FAD-dependent polyamine oxidases are key players in plant tissue differentiation and organ development. Front Plant Sci. 2016;7:824. PMID:27446096. doi:10.3389/fpls.2016.00824.
- Wang W, Paschalidis K, Feng J-C, Song J, Liu J-H. Polyamine catabolism in plants: a universal process with diverse functions. Front Plant Sci. 2019;10:561. PMID:31134113. doi:10.3389/fpls.2019.00561.