
ABSTRACT
Salicylic acid (SA) is an important signaling hormone in plant immunity. It can be synthesized by either the phenylpropanoid pathway or the isochorismate pathway, but mutant studies of this have been scarce in other species than Arabidopsis. Here we identified a mutation that introduced a stop-codon early in the barley gene for isochorismate synthase (ICS). We found that homozygous ics plants wilted if not sprayed with 1,4-dihydroxy-2-naphthoic acid, a precursor of phylloquinone, also synthesized via the isochorismate pathway. Interestingly, ics had unchanged SA, suggesting that the basal level of SA is synthesized via the phenylpropanoid pathway. Previous studies have failed seeing increased SA levels in barley after attack by the powdery mildew fungus, Blumeria graminis f.sp. hordei (Bgh), and indeed, we saw no changes in the interaction of ics with this fungus. Overall, we hope this mutant will be useful for other studies of SA in barley.
Plants have a highly advanced immune system, which efficiently resist most pathogens. After plant receptors have recognized pathogen cues, a sophisticated signaling network activates transcriptional and cellular reprogramming that prevents pathogens from causing disease. This includes expression of many antimicrobial proteins and secondary metabolites, buildup of cellular structures to prevent pathogen ingress and activation of a localized programmed cell death reaction, referred to as the hypersensitive response (HR).Citation1 A central compound in signaling for immunity is salicylic acid (SA). It activates many downstream response genes, e.g. encoding pathogenesis-related proteins, and besides it has a function as a general amplifier of immunity.Citation2 SA accumulates in many plants after pathogen attack, and RNA-seq data suggest that it stimulates immunity through its activation of a large number of key regulator genes.Citation3 The plant ubiquitous protein, NPR1, plays a central role as an SA sensor that regulates transcriptional reprogramming in immunity after it becomes re-localized to the nucleus where it interacts with a number of transcription factors.Citation4
SA is synthesized by two biosynthetic pathways, the isochorismate pathway and the phenylpropanoid pathway.Citation2 In Arabidopsis, basal and pathogen-induced levels of SA are primarily derived from the isochorismate pathway, albeit the phenylpropanoid pathway also contributes. This was shown in plants mutated in the genes encoding isochorismate synthase 1 (ICS1, also named SA induction-deficient 2, SID2), ICS2 and four genes encoding phenylalanine ammonia-lyase.Citation4–Citation8 However, the use of mutants to study the two pathway’s contribution to SA biosynthesis has been very sparse in other plants. Here this question instead has been addressed using RNA interference.Citation9 For instance, virus-induced gene silencing of ICS in Nicotiana benthamiana suggested that pathogen-induced SA is derived from isochorismate.Citation10
The importance of SA in Poaceae species is less clear than in dicot species, and the contribution of the two biosynthesis pathway is not well studied. Even though SA appears involved in Arabidopsis resistance to bacterial and powdery mildew fungal pathogens,Citation5,Citation11,Citation12 it does not accumulate in barley in response to such pathogens.Citation13,Citation14 Yet, a recent study showed that barley (Hordeum vulgare) leaves inoculated with Fusarium graminearum had a 30-fold increase in the SA level, which, in an ICS RNAi transgenic line, was reduced to basal level.Citation15 Interestingly, the same study showed that the ICS RNAi line has decreased post-invasive immunity to Bgh, which is unexpected according to Hückelhoven et al.Citation13 A study of maize attacked by Ustilago maydis has revealed that a fungal chorismate mutase effector contributes to fungal virulence after transfer to cells of the plant, where it reduces the SA level.Citation16 This strongly advocates for the importance of SA, synthesized by the isochorismate pathway, in immunity in maize.
In the present work, we aimed at mutant studies of SA in barley. This species has a single gene encoding isochorismate synthase (ICS).Citation15 To search for mutations in this gene, a cultivar Golden Promise mutant population was obtained after treatment with fast neutrons in a 12–18 Gy dose. The resulting M2 seeds were mutagenized again by steeping in 1.5 mM sodium azide for 2½ h before soaking in tap water for 2 h. The seeds were subsequently dried and sown in the field. M2 seed samples were harvested separately from 2,132 single plants, and one seed from each sample was germinated for DNA extraction. The resulting DNA samples were screened by TILLING according to de Bang et al.Citation17 This led to the identification of a heterozygous line, ICS ics, where codon 153 (TGG, encoding tryptophan) was changed into the stop-codon, TGA. ICS has several exons. However, transcript sequence data indicate that only one splice variant exits, and it contains codon 153 (https://ics.hutton.ac.uk/barleyrtd/blast_page.html). One-fourth of the offspring resulting from the selfing of ICS ics started wilting ~10 days after emergence (), and genotyping of 84 wilting and 238 non-wilting plants demonstrated complete co-segregation of ics homozygosity and wilting.
Figure 1. Wilting of ics is overcome by spraying with 1,4-dihydroxy-2-naphthoic acid (NA). Four 18-day-old ics plants sprayed with 0.1 mM NA or buffer (0.1% dimethyl sulfoxide, 0.1% Tween-20) every other day from day 7 after sowing. WT, Golden Promise. Homozygous ics plants were selected by genotyping using the allele-specific primers, ICS-F 5ʹ-TGGCCACAAGGAGCAGTAG-3ʹ, ics-F 5ʹ-GTGGCCACAAGGAGCAGTAA-3ʹ, and reverse primer, ICS-R 5ʹ-TCACTGAACTCAACCTGCAAA-3ʹ, in a touch-down PCR reaction in which the annealing temperature was reduced from 74ºC to 66ºC over the first eight cycles.

Garcion et al.Citation7 previously found the Arabidopsis double mutant ics1 ics2 to be completely deficient of phylloquinone, which is essential for electron transfer in photosystem I. Isochorismate is an early precursor for phylloquinone, synthesized via 1,4-dihydroxy-2-naphthoic acid (NA) in a multistep pathway.Citation18,Citation19 Thus, we were able to rescue the barley ics plants to almost wild-type size by spraying with NA (). ICS is located on barley chromosome 5, whereas the genes, encoding the enzymes mediating the biosynthetic steps between isochorismate and NA,Citation19 are located on barley chromosomes 3, 4, 6 and 7. Thus, the rescuing by NA, together with the co-segregation of ics and wilting, confirms that the ics mutation prevents isochorismate synthesis.
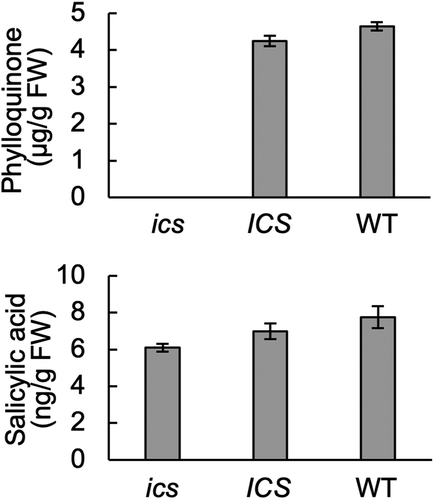
Next, we analyzed the content of phylloquinone and SA in homozygous ics and ICS, segregating from ICS ics, as well as in wild-type Golden Promise. Phylloquinone was undetectable in ics and indifferent in ICS and wild-type, while all three samples had approximately 7 ng SA/g fresh weight (). This indicated that basal SA is not derived from the isochorismate pathway in barley.
Figure 2. Phylloquinone, but not SA, is reduced in ics. Analyses were made by UHPLC-MS/MS according to Glauser et al.Citation20 Leaves from homozygous ICS and ics plants were selected by analyzing DNA from leaf tips as in . Each sample consisted of two first leaves from 10-day-old barley plants. Error bars, SE; n, 6 (phylloquinone) and 8 (SA).
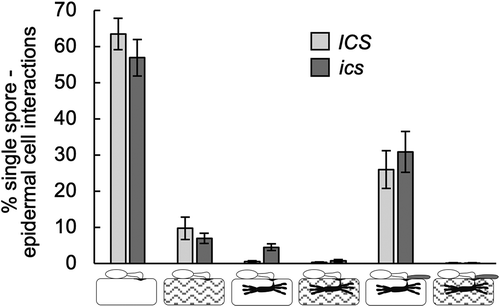
Despite no induction of the SA-level has been observed after inoculation with Bgh,Citation13 Hao et al.Citation15 observed that ICS RNAi lines had decreased immunity to this pathogen. This spurred us to re-test this observation, as it cannot be ruled out that a minor localized SA increase, not detectable in whole leaf extracts, can have significance. Therefore, genotyped ics and ICS plants, segregating from ICS ics, were inoculated with Bgh and the rate of penetration, secondary hyphal growth and host cell death were estimated (). However, no significant differences between ics and ICS plants were revealed.
Figure 3. ics has a normal interaction with the powdery mildew fungus. Two days after inoculation with the virulent Bgh isolate, DH14, leaf tips were cut for genotyping (see ), and the remaining leaf parts were trypan blue stained for microscopy. Five leaves (repeats) were assessed from each genotype, and the outcome of 110 to 210 single spore interactions was scored from each leaf according to Zhang et al.Citation21 Error bars, SE.
In summary, we have identified a barley ics knock-out allele, which when homozygous confers plant wilting. This phenotype could be rescued by spraying with NA, confirming that the mutant is affected in the isochorismate pathway. Yet, we were unable to observe a change in the SA content of ics, suggesting that basal SA in barley is produced by the phenylpropanoid pathway. As anticipated from previous failures to see the induction of SA levels in barley in response to Bgh,Citation13 we observed no altered interaction of ics with this pathogen. Yet, we hope this mutant will become useful for studies of signaling in immunity in barley.
Acknowledgments
We are grateful for fruitful discussions with Assoc. Prof. Mathias Pribil. Financial support to YQ was obtained from The China Scholarship Council.
Additional information
Funding
References
- Jones JD, Dangl JL. The plant immune system. Nature. 2006;444:1–3.
- Zhang Y, Li X. Salicylic acid: biosynthesis, perception, and contributions to plant immunity. Curr Opin Plant Biol. 2019;50:29–36.
- Ding Y, Sun T, Ao K, Peng Y, Zhang Y, Li X, Zhang Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell. 2018;173:1454–1467.
- Withers J, Dong X. Posttranslational modifications of NPR1: a single protein playing multiple roles in plant immunity and physiology. PLoS Pathog. 2016;12:e1005707.
- Nawrath C, Métraux J-P. Salicylic acid induction–deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell. 1999;11:1393–1404.
- Wildermuth MC, Dewdney J, Wu G, Ausubel FM. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature. 2001;414:562–565.
- Garcion C, Lohmann A, Lamodière E, Catinot J, Buchala A, Doermann P, Métraux JP. Characterization and biological function of the ISOCHORISMATE SYNTHASE2 gene of Arabidopsis. Plant Physiol. 2008;147:1279–1287.
- Huang J, Gu M, Lai Z, Fan B, Shi K, Zhou YH, Yu JQ, Chen Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010;153:1526–1538.
- Berens ML, Berry HM, Mine A, Argueso CT, Tsuda K. Evolution of hormone signaling networks in plant defense. Ann Rev Phytopathol. 2017;55:401–425.
- Catinot J, Buchala A, Abou-Mansour E, Métraux JP. Salicylic acid production in response to biotic and abiotic stress depends on isochorismate in Nicotiana benthamiana. FEBS Lett. 2008;582:473–478.
- Reuber TL, Plotnikova JM, Dewdney J, Rogers EE, Wood W, Ausubel FM. Correlation of defense gene induction defects with powdery mildew susceptibility in Arabidopsis enhanced disease susceptibility mutants. Plant J. 1998;16:473–485.
- Zhang -L-L, Ma X-F, Zhou -B-B, Zhao J-Q, Fan J, Huang F, Li Y, Wang W-M. EDS1-mediated basal defense and SA-signaling contribute to post-invasion resistance against tobacco powdery mildew in Arabidopsis. Physiol Mol Plant Pathol. 2015;91:120–130.
- Hückelhoven R, Fodor J, Preis C, Kogel KH. Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation. Plant Physiol. 1999;119:1251–1260.
- Dey S, Wenig M, Langen G, Sharma S, Kugler KG, Knappe C, Hause B, Bichlmeier M, Babaeizad V, Imani J, et al. Bacteria-triggered systemic immunity in barley is associated with WRKY and ETHYLENE RESPONSIVE FACTORs but not with salicylic acid. Plant Physiol. 2014;166:2133–2151.
- Hao Q, Wang W, Han X, Wu J, Lyu B, Chen F, Caplan A, Li C, Wu J, Wang W, et al. Isochorismate-based salicylic acid biosynthesis confers basal resistance to Fusarium graminearum in barley. Mol Plant Pathol. 2018;19:1995–2010.
- Djamei A, Schipper K, Rabe F, Ghosh A, Vincon V, Kahnt J, Osorio S, Tohge T, Fernie AR, Feussner I, et al. Metabolic priming by a secreted fungal effector. Nature. 2011;478:395–398.
- de Bang L, Torp AM, Rasmussen SK. TILLING in Brachypodium distachyon. Meth Mol Biol. 2018;1667:173–186.
- Gross J, Cho WK, Lezhneva L, Falk J, Krupinska K, Shinozaki K, Seki M, Herrmann RG, Meurer J. A plant locus essential for phylloquinone (vitamin K1) biosynthesis originated from a fusion of four eubacterial genes. J Biol Chem. 2006;281:17189–17196.
- van Oostende C, Widhalm JR, Furt F, Ducluzeau A-L, Basset GJ. Vitamin K1 (phylloquinone): Function, enzymes and genes. Adv Bot Res. 2011;59:229–261.
- Glauser G, Vallat A, Balmer D. Hormone profiling. Meth Mol Biol. 2014;1062:597–608.
- Zhang Z, Feechan A, Pedersen C, Newman M-A, Qiu JL, Olesen KL, Thordal-Christensen HA. SNARE-protein has opposing functions in penetration resistance and defence signalling pathways. Plant J. 2007;49:302–312.