
ABSTRACT
Changes in the growth environment can generate imbalances in chloroplast photosynthetic metabolism. Under water deficit, stomatal closure limits CO2 availability such that the production of ATP and NADPH by the thylakoid membrane-localized electron transport chain may not match the consumption of these energy intermediates by the stroma-localized Calvin-Benson cycle, thus challenging energy balance. Alternatively, in an elevated CO2 atmosphere, carbon fixation by the Calvin-Benson cycle may outpace the activity of downstream carbohydrate-utilizing processes, thus challenging carbon balance. Our previous studies have shown that, in both of the above scenarios, a mitochondrial alternative oxidase contributes to maintaining energy or carbon balance, highlighting the importance of photosynthesis-respiration interactions in optimizing photosynthesis in different growth environments. In these previous studies, we observed aberrant amounts of chloroplast ATP synthase protein across the different transgenic plant lines and growth conditions, compared to wild-type. Based on these observations, we develop here the hypothesis that an important determinant of chloroplast ATP synthase protein amount is the stromal concentration of inorganic phosphate. ATP synthase is a master regulator of photosynthesis. Coarse control of ATP synthase protein amount by the stromal inorganic phosphate status could provide a means to coordinate the electron transport and carbon fixation reactions of photosynthesis.
Chloroplast photosynthesis and mitochondrial respiration represent the core of plant primary metabolism, and these pathways share many important carbon and energy intermediates. Hence, it is likely that these organelles and metabolic pathways must act in a coordinated manner to maintain energy and carbon balance.Citation1–Citation4 Such coordination of activities might be particularly important during developmental or environmental changes that can challenge metabolism. Key aspects of energy balance include the ATP/ADP, NAD(P)H/NAD(P)+, and ATP/NAD(P)H ratios of different cell compartments.Citation5 Key aspects of carbon balance include the carbohydrate status of source (photosynthetic) and sink (non-photosynthetic) tissues within the plant. These effects relate to the importance of carbohydrates not only as a source of carbon and energy but also as signaling molecules able to influence growth and development through changes in gene expression.Citation6–Citation8
Photosynthesis involves a light-driven electron transport process [the chloroplast electron transport chain (cETC)] generating NADPH and ATP, and a CO2-dependent carbon fixation pathway [the Calvin-Benson (CB) cycle, which includes the enzyme ribulose bisphosphate carboxylase oxygenase (Rubisco)] that utilizes the NADPH and ATP to produce triose phosphates (TP).Citation9 The effective delivery of CO2 to the CB cycle depends upon stomatal pores on the leaf surface being open. However, if water becomes limiting in the environment, these pores close to conserve water. Hence, changes in the availability of light, CO2 and water each affect parts of the photosynthetic process, and hence have the potential to generate carbon and/or energy imbalances in this metabolism.Citation5 The TP produced by the CB cycle can be used in the chloroplast for starch synthesis, or may be transported to the cytosol for sucrose synthesis or for entry into respiratory and other pathways that support the growth and maintenance of the source tissue. Sucrose synthesized in the source is exported via the phloem to sink tissues, where it is processed by respiratory and other pathways in support of sink growth, maintenance and storage processes.Citation10–Citation13
The respiratory pathways include glycolysis, the oxidative pentose phosphate pathway, the tricarboxylic acid cycle and the mitochondrial electron transport chain (mETC).Citation14,Citation15 Respiration consumes carbohydrate generated by photosynthesis. This produces various carbon intermediates and NAD(P)H used for biosynthetic reactions, but also generates NAD(P)H that is subsequently oxidized by the mETC, and coupled to the production of ATP. A distinctive feature of the plant mETC is that it includes two terminal oxidases able to reduce oxygen to water. These include the usual cytochrome (cyt) oxidase that accepts electrons from the ubiquinone pool via Complex III and cyt c; and an alternative oxidase (AOX) that accepts electrons directly from the ubiquinone pool.Citation15,Citation16 While electron flow through both Complex III and cyt oxidase (the cyt pathway) is coupled to proton translocation from the matrix to inter-membrane space, electron flow through AOX is not. Hence, the rate of ATP synthesis by respiration will depend upon both the absolute rate of electron flow to oxygen and the partitioning of electrons in the ubiquinone pool between the two terminal oxidases. Practically, as the partitioning of electrons to AOX increases, the ability of respiration to support any given rate of ATP turnover will require higher overall rates of NAD(P)H turnover that might also be coupled with higher overall rates of carbohydrate consumption. Said differently, AOX should be most active when the demand for respiration to oxidize reductant or to consume carbohydrate is greater than the demand to produce additional ATP. AOX is subject to tight transcriptional regulation and sophisticated biochemical control, presumably to ensure an appropriate partitioning of electrons to this non-energy conserving pathway.Citation16,Citation17
An important component of chloroplast energy balance is the production ratio of ATP to NADPH. It is widely accepted that chloroplast linear electron transport (LET) alone does not produce enough ATP relative to NADPH to fulfill CB cycle requirements.Citation5 Further, the CB cycle requirement for ATP relative to NADPH is not a constant, but rather decreases as the ratio of carboxylation to oxygenation by Rubisco increases.Citation18 In other words, growth conditions that favor photorespiration (e.g. water deficit) increase the chloroplast ATP/NADPH imbalance associated with LET, while conditions that suppress photorespiration [e.g. growth at elevated atmospheric concentrations of CO2 (ECO2)] decrease this imbalance. Other chloroplast-localized processes such as cyclic electron transport around photosystem I,Citation19 the Mehler reaction,Citation20 or a plastid terminal oxidaseCitation21 could correct the ATP/NADPH imbalance. However, another means to correct the imbalance would be to export some reductant to the cytosol, such as through the malate valve.Citation22 This would allow LET to support additional ATP synthesis in the chloroplast without a buildup of stromal NADPH. In turn, the cytosolic reductant may be oxidized by the mETC.Citation1–Citation4 For this, the partitioning of electrons between cyt oxidase and AOX could be optimized to fulfill the extra-chloroplastic demand for ATP.
Our recent studies using transgenic Nicotiana tabacum (tobacco) plants with increased or decreased amounts of AOX protein indicated that this respiratory pathway aided in maintaining photosynthetic performance under different growth conditions by preserving energy or carbon balance (). One such growth condition was water deficit, as recently reviewed.Citation23 As the severity of water deficit increased, wild-type (WT) tobacco plants increased their capacity for AOX respiration, while their capacity for cyt pathway respiration declined.Citation24 Owing to this, the respiration rate of AOX knockdowns under water deficit was lower than in WT plants,Citation25 while the respiration rate of AOX overexpressors was higher than in the WT.Citation26 These differences in respiration had a strong impact on photosynthetic performance across the plant lines. In the cETC, the fraction of closed (reduced) photosystem II reaction centers was higher in the AOX knockdowns than WT, and there was a strong engagement in the knockdowns of feedback controls on the cETC that ultimately compromised carbon fixation relative to WT plants.Citation25 On the other hand, AOX overexpressors maintained higher rates of carbon fixation than WT under the water deficit conditions.Citation26 During long-term water deficit, these differences in photosynthetic rate across plant lines translated into differences in plant growth.Citation27 Overall, the differences in photosynthesis across plant lines appeared to be primarily due to differences in chloroplast energy balance, which as the severity of water deficit increased, translated into differences in the amount of key photosynthetic proteins and hence photosynthetic capacity. In particular, the knockdowns displayed lower amounts and the overexpressors displayed higher amounts (relative to WT) of chloroplast ATP synthase (AtpB subunit) and Rubisco (RbcS subunit) protein (see more below).
Figure 1. A working model for photosynthesis-respiration interactions in tobacco under different growth conditions. The “ATP/NADPH imbalance” in the chloroplast refers to the shortage in production rate of ATP relative to NADPH, assuming that only linear electron transport is supplying these to the Calvin-Benson cycle. The Calvin-Benson cycle demand for ATP relative to NADPH decreases as the ratio of carboxylation to oxygenation by ribulose bisphosphate carboxylase oxygenase increases. Compared to normal growth conditions, this demand is low during growth at elevated atmospheric concentrations of CO2 and high during growth under water deficit. One means to correct the ATP/NADPH imbalance is to export NADPH to the cytosol via a metabolite shuttle such as the malate valve. Hence, to correct the chloroplast ATP/NADPH imbalance, and relative to normal growth conditions, growth at elevated CO2 may reduce NADPH export to the cytosol while water deficit may increase NADPH export to the cytosol. Compared to normal growth conditions, growth at elevated CO2 increases the Calvin-Benson cycle supply of triose phosphates to the cytosol, while water deficit decreases this supply. The mitochondrial electron transport chain oxidizes NAD(P)H that is being supplied either directly from the chloroplast via the malate valve, or indirectly via the metabolism of chloroplast-derived triose phosphates by cytosolic glycolysis and the mitochondrial tricarboxylic acid cycle. Under normal growth conditions, it is likely that most electrons are consumed by cytochrome oxidase, allowing the mitochondrion to supply sufficient ATP to the cytosol for sucrose synthesis. Under water deficit, cytochrome oxidase protein amount declines while alternative oxidase protein amount increases; and alternative oxidase becomes a necessary electron sink in the photosynthetic cell. This reduces the mitochondrial supply of ATP to the cytosol, but ATP demand for sucrose synthesis is likely low regardless, due to the relatively low rates of chloroplast triose phosphate production. Growth at elevated CO2 increases both cytochrome oxidase and alternative oxidase protein amounts, but little information is available regarding the partitioning of electrons between the two pathways under these conditions. Nonetheless, AOX does become a necessary electron sink, in this case to prevent a buildup of excess carbohydrate in the photosynthetic cell due to the relatively high rates of chloroplast triose phosphate production. See text for additional details. AOX, alternative oxidase; COX, cytochrome oxidase; TP, triose phosphate.
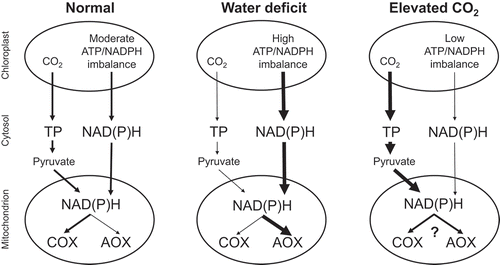
In another study, photosynthesis was compared between the WT and AOX transgenic lines following growth of the plants, since germination, at ECO2.Citation28 Under these growth conditions, photosynthesis was again compromised in the AOX knockdowns relative to WT, but this was not due to lower amounts of ATP synthase or Rubisco protein (in fact these were elevated compared to WT, see more below). The knockdown plants accumulated significantly higher amounts of starch, sucrose and hexose phosphates in the source leaf relative to WT, indicating that AOX respiration was necessary to maintain carbon balance under ECO2 growth conditions (). This carbohydrate accumulation lowered the capacity of these plants to utilize the TP being generated by the CB cycle (i.e. they displayed a TPU limitationCitation13). This was accompanied by a lower ATP synthase activity that resulted in a higher proton motive force (pmf) across the thylakoid membrane, hence engaging pH-dependent feedback controls on the cETC that then limited carbon fixation.Citation28
The above studies (summarized in ) illustrate that AOX respiration is necessary to optimize photosynthesis in tobacco during water deficit or during growth at ECO2. Interestingly, for both of these growth conditions, the perturbations of photosynthesis in the AOX transgenic plants related to the activity or protein amount of chloroplast ATP synthase. Hence, the remainder of this report focuses on this critical component of photosynthesis.
Light-driven chloroplast electron transport results in the pumping of protons from the stroma to thylakoid lumen and the resulting pmf is used by the chloroplast ATP synthase to drive the synthesis of ATP from ADP and inorganic phosphate (Pi) (). Effective photosynthesis depends upon an optimal chloroplast ATP synthase activity.Citation29,Citation30 If activity were too low, an insufficient rate of ATP supply could limit CB cycle activity. Too low ATP synthase activity could also decrease lumen pH to the extent that pH-dependent feedback controls on the cETC become engaged.Citation31,Citation32 These photoprotective feedback controls include energy-dependent non-photochemical quenching of excitation energy at photosystem II, and “photosynthetic control” that slows plastoquinol oxidation by the cyt b6f complex.Citation18,Citation33,Citation34 On the other hand, if ATP synthase activity were too high, this might compromise the plant’s ability to engage these photoprotective controls when needed. Too high ATP synthase activity might also promote the engagement of futile ATP-consuming cycles.Citation35–Citation37
Figure 2. Chloroplast ATP synthase is a master regulator of photosynthesis. Light-driven electron transport generates NADPH and a proton gradient between the stroma and lumen. The proton gradient provides the proton motive force for energy (ATP) generation, but the lumen protons can also engage feedback biochemical controls that slow electron transport. The activity of the chloroplast ATP synthase will determine the extent to which the proton gradient is either dissipated to provide ATP for the Calvin-Benson cycle, or allowed to build up and hence engage the feedback controls on electron transport. Stromal Pi and ADP are the substrates for ATP synthase. Stromal Pi has also been hypothesized to act as an allosteric effector of ATP synthase activity (see text) and/or as a signal molecule able to influence ATP synthase protein amount (this work). NPQ, non-photochemical quenching; pmf, proton motive force.
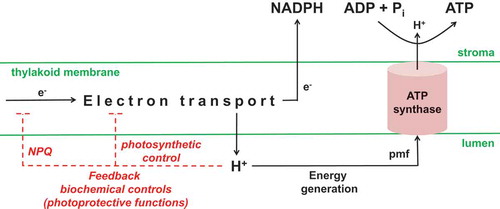
A critical factor determining chloroplast ATP synthase activity is the concentration of its substrate, Pi (). There is a kinetic effect related to the Km of ATP synthase for Pi. There is also a thermodynamic effect, since a lower Pi, at constant ATP/ADP, increases ΔGATP, thus necessitating a greater pmf for ATP synthesis. Hence, recent experiments with intact leaves and isolated chloroplasts of barley, as well as recent model simulations of photosynthetic electron transport in Arabidopsis, each indicate that even minor changes in stromal Pi concentration can have a major impact on chloroplast ATP synthase activity.Citation38,Citation39 Further, when Cucumis melo was grown under Pi-limiting conditions, it displayed a restricted chloroplast ATP synthase activity.Citation40 Also, environmental conditions that favor the accumulation of phosphorylated carbon intermediates deplete stromal Pi,Citation41 and such conditions restrict chloroplast ATP synthase activity.Citation42 The possibility of Pi (or a closely related metabolite) acting as an allosteric effector of ATP synthase has also been speculated.Citation31,Citation32
Based on our previous work, we develop here a hypothesis that the stromal concentration of Pi is also an important determinant of chloroplast ATP synthase protein amount. In other words, stromal Pi provides a coarse control of ATP synthase activity by influencing the amount of ATP synthase protein present. Such coarse control by Pi, presumably acting directly or indirectly as a signal molecule, could complement the finer biochemical control of ATP synthase activity by stromal Pi acting as substrate, as discussed above. Our hypothesis is based on a comparison of the ATP synthase protein amounts measured in the WT and AOX transgenic tobacco lines under different growth conditions, as described above. These comparisons are summarized in . As discussed earlier, under water deficit, increased photorespiration enhances the imbalance of ATP to NADPH production by LET, requiring AOX to act as an additional electron sink. In AOX knockdowns, a slowed stromal NADPH turnover rate will slow stromal ATP synthesis, relative to WT. In overexpressors, an enhanced stromal NADPH turnover rate will enhance stromal ATP synthesis, relative to WT. Hence, the stromal ATP/NADPH ratio is predicted to be lower in the AOX knockdowns and higher in the AOX overexpressors, relative to WT (). Based on this, one can also predict that stromal Pi concentration will be higher in the knockdowns and lower in the overexpressors, relative to WT. High Pi enhances ATP synthase activity. Hence, if stromal Pi acts as a signal controlling ATP synthase protein amount, the abnormally high Pi in the knockdowns relative to WT should be interpreted by the plant as a need to lower ATP synthase protein amount relative to WT, as was indeed observed (). Otherwise, the plant risks too high ATP synthase activity and the potential problems associated with this (see above). On the other hand, the abnormally low Pi in the overexpressors relative to WT should be interpreted as a need to increase ATP synthase protein amount, as a means to bolster ATP synthase activity in the face of low Pi. This was also observed ().
Table 1. Predicted relative concentrations of stromal inorganic phosphate (Pi) correlate with measured changes in chloroplast ATP synthase (AtpB subunit) and Rubisco (RbcS subunit) protein amounts in different growth environments (water deficit or elevated atmospheric concentrations of CO2) and across plant lines with altered respiration (alternative oxidase knockdowns and overexpressors). See text for further details. Based on data from references 25–28.
Under ECO2 growth conditions, photorespiration is suppressed. This lowers the imbalance between ATP and NADPH production by LET, which should lower the reliance upon alternative electron sinks such as AOX to correct this energy imbalance. Nonetheless, AOX remained essential to optimize photosynthesis under these conditions, but for different reasons. Rather than energy balance, the need for AOX was to maintain carbon balance. Enhanced rates of Rubisco carboxylation necessitated that some carbohydrate be consumed in the photosynthetic cell using AOX respiration. Hence, AOX knockdowns accumulated phosphorylated sugars (as well as starch and sucrose), relative to WT plants (). Since the production of phosphorylated sugars requires stromal Pi, the buildup of phosphorylated sugars is predicted to lower stromal Pi in the knockdowns relative to WT. This is the opposite of the prediction under water deficit. Based on our hypothesis, this low stromal Pi should now signal for an increase in ATP synthase protein amount in the knockdowns, relative to WT, as was indeed observed (). On the other hand, AOX overexpression had no impact on the sugar phosphate pools, relative to WT. Hence, these plants can be predicted to display a similar ATP synthase protein amount as WT, as was again observed ().
The observations summarized in and discussed above are consistent with the hypothesis that the stromal concentration of Pi (or perhaps some closely associated factor) is an important determinant of chloroplast ATP synthase protein amount. These observations involved contrasting growth conditions, multiple plant lines with differing respiratory sink capacities, and plants displaying a wide range of photosynthetic characteristics. Hence, the hypothesis seems robust and worthy of further investigation. A definitive experimental approach would include direct in vivo measurements of the stromal Pi concentration under defined physiological conditions. Such subcellular measurements are becoming more feasible, particularly given advances in the use of genetically encoded Pi sensors.Citation43,Citation44 Similar ATP sensors have also been developed.Citation45,Citation46
Interestingly, all of the patterns of ATP synthase protein amount across plant lines and growth conditions discussed above were accompanied by a similar pattern of Rubisco (RbcS) protein amount (). As discussed before,Citation27 this tight coordination may be part of a more general system acting to harmonize the capacity of the thylakoid and stromal reactions.Citation47–Citation49 Previous studies have also emphasized the importance of balancing the activities of ATP synthase and cyt b6f since cyt b6f is the rate-limiting step in pmf generation across the thylakoid membrane while ATP synthase is the major means to dissipate this pmf.Citation48 However, this coordination was clearly disrupted in the AOX knockdowns and overexpressors in that, whenever they exhibited aberrant amounts of ATP synthase relative to WT, they nonetheless maintained normal amounts of cyt b6f (cyt f subunit) relative to WT.Citation25–Citation28
Other work also provides evidence consistent with the hypothesis that Pi status can act as a signal controlling the abundance of key chloroplast photosynthetic components. Hurry and colleagues, utilizing Arabidopsis pho mutants with decreased and increased shoot Pi amounts, showed that low Pi favored an increase in RbcS transcript and Rubisco activity.Citation50 While no data on chloroplast ATP synthase was reported, the study does nonetheless provide evidence that the abundance of photosynthetic components is responsive to Pi status. Another study in tobacco showed that a sugar-mediated repression of RbcS transcript did not occur if tissue Pi was simultaneously low.Citation51
Relatively little is known about the molecular processes controlling chloroplast ATP synthase protein amount.Citation52 Nonetheless, proteins that facilitate ATP synthase assembly are becoming identifiedCitation53–Citation55 and N-terminal acetylation has been recognized as a factor controlling the degradation rate of a chloroplast ATP synthase subunit in response to water deficit.Citation56,Citation57 It would be interesting to establish whether any of these processes respond to stromal concentrations of Pi.
Disclosure of potential conflicts of interest
The authors report no conflict of interest.
Additional information
Funding
References
- Hoefnagel MHN, Atkin OK, Wiskich JT. Interdependence between chloroplasts and mitochondria in the light and the dark. Biochim Biophys Acta. 1998;1366:1–7. doi:10.1016/S0005-2728(98)00126-1.
- Raghavendra AS, Padmasree K. Beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Trends Plant Sci. 2003;8:546–553. doi:10.1016/j.tplants.2003.09.015.
- Gardeström P, Igamberdiev AU. The origin of cytosolic ATP in photosynthetic cells. Physiol Plant. 2016;157:367–379. doi:10.1111/ppl.12455.
- Shameer S, Ratcliffe G, Sweetlove LJ. Leaf energy balance requires mitochondrial respiration and export of chloroplast NADPH in the light. Plant Physiol. 2019;180:1947–1961. doi:10.1104/pp.19.00624.
- Kramer DM, Evans JR. The importance of energy balance in improving photosynthetic productivity. Plant Physiol. 2011;155:70–78. doi:10.1104/pp.110.166652.
- Lastdrager J, Hanson J, Smeekens S. Sugar signals and the control of plant growth and development. J Exp Bot. 2014;65:799–807. doi:10.1093/jxb/ert474.
- Ruan Y-L. Sucrose metabolism: gateway to diverse carbon use and sugar signaling. Annu Rev Plant Biol. 2014;65:33–67. doi:10.1146/annurev-arplant-050213-040251.
- Griffiths CA, Paul MJ, Foyer CH. Metabolite transport and associated sugar signalling systems underpinning source/sink interactions. Biochim Biophys Acta. 2016;1857:1715–1725. doi:10.1016/j.bbabio.2016.07.007.
- Stitt M, Lunn J, Usadel B. Arabidopsis and primary photosynthetic metabolism – more than the icing on the cake. Plant J. 2010;61:1067–1091. doi:10.1111/j.1365-313X.2010.04142.x.
- Lewis CE, Noctor G, Causton D, Foyer CH. Regulation of assimilate partitioning in leaves. Aust J Plant Physiol. 2000;27:507–519.
- Paul MJ, Foyer CH. Sink regulation of photosynthesis. J Exp Bot. 2001;52:1383–1400. doi:10.1093/jexbot/52.360.1383.
- Dong S, Beckles DM. Dynamic changes in the starch-sugar interconversion within plant source and sink tissues promote a better abiotic stress response. J Plant Physiol. 2019;234–235:80–93. doi:10.1016/j.jplph.2019.01.007.
- McClain AM, Sharkey TD. Triose phosphate utilization and beyond: from photosynthesis to end product synthesis. J Exp Bot. 2019;70:1755–1766. doi:10.1093/jxb/erz058.
- Plaxton WC, Podesta FE. The functional organization and control of plant respiration. Crit Rev Plant Sci. 2006;25:159–198. doi:10.1080/07352680600563876.
- Millar AH, Whelan J, Soole KL, Day DA. Organization and regulation of mitochondrial respiration in plants. Annu Rev Plant Biol. 2011;62:79–104. doi:10.1146/annurev-arplant-042110-103857.
- Vanlerberghe GC. Alternative oxidase: a mitochondrial respiratory pathway to maintain metabolic and signaling homeostasis during abiotic and biotic stress in plants. Int J Mol Sci. 2013;14:6805–6847. doi:10.3390/ijms14046805.
- Selinski J, Scheibe R, Day DA, Whelan J. Alternative oxidase is positive for plant performance. Trends Plant Sci. 2018;23:588–597. doi:10.1016/j.tplants.2018.03.012.
- Foyer CH, Neukermans J, Queval G, Noctor G, Harbinson J. Photosynthetic control of electron transport and the regulation of gene expression. J Exp Bot. 2012;63:1637–1661. doi:10.1093/jxb/ers013.
- Yamori W, Shikanai T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annu Rev Plant Biol. 2016;67:81–106. doi:10.1146/annurev-arplant-043015-112002.
- Asada K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006;141:391–396. doi:10.1104/pp.106.082040.
- Nawrocki WJ, Tourasse NJ, Taly A, Rappaport F, Wollman F-A. The plastid terminal oxidase: its elusive function points to multiple contributions to plastid physiology. Annu Rev Plant Biol. 2015;66:49–74. doi:10.1146/annurev-arplant-043014-114744.
- Taniguchi M, Miyake H. Redox-shuttling between chloroplast and cytosol: integration of intra-chloroplast and extra-chloroplast metabolism. Curr Opin Plant Biol. 2012;15:252–260. doi:10.1016/j.pbi.2012.01.014.
- Vanlerberghe GC, Martyn GD, Dahal K. Alternative oxidase: a respiratory electron transport chain pathway essential for maintaining photosynthetic performance during drought stress. Physiol Plant. 2016;157:322–337. doi:10.1111/ppl.12451.
- Dahal K, Vanlerberghe GC. Alternative oxidase respiration maintains both mitochondrial and chloroplast function during drought. New Phytol. 2017;213:560–571. doi:10.1111/nph.14169.
- Dahal K, Wang J, Martyn GD, Rahimy F, Vanlerberghe GC. Mitochondrial alternative oxidase maintains respiration and preserves photosynthetic capacity during moderate drought in Nicotiana tabacum. Plant Physiol. 2014;166:1560–1574. doi:10.1104/pp.114.247866.
- Dahal K, Martyn GD, Vanlerberghe GC. Improved photosynthetic performance during severe drought in Nicotiana tabacum overexpressing a nonenergy conserving respiratory electron sink. New Phytol. 2015;208:382–395. doi:10.1111/nph.13479.
- Dahal K, Vanlerberghe GC. Improved chloroplast energy balance during water deficit enhances plant growth: more crop per drop. J Expt Bot. 2018;69:1183–1197. doi:10.1093/jxb/erx474.
- Dahal K, Vanlerberghe GC. Growth at elevated CO2 requires acclimation of the respiratory chain to support photosynthesis. Plant Physiol. 2018;178:82–100. doi:10.1104/pp.18.00712.
- Rott M, Martins NF, Thiele W, Lein W, Bock R, Kramer DM, Schöttler MA. ATP synthase repression in tobacco restricts photosynthetic electron transport, CO2 assimilation, and plant growth by overacidification of the thylakoid lumen. Plant Cell. 2011;23:304–321. doi:10.1105/tpc.110.079111.
- Yamori W, Takahashi S, Makino A, Price GD, Badger MR, von Caemmerer S. The roles of ATP synthase and the cytochrome b6/f complexes in limiting chloroplast electron transport and determining photosynthetic capacity. Plant Physiol. 2011;155:956–962. doi:10.1104/pp.110.168435.
- Kanazawa A, Kramer DM. In vivo modulation of nonphotochemical exciton quenching (NPQ) by regulation of ATP synthase. PNAS. 2002;99:12789–12794. doi:10.1073/pnas.182427499.
- Takizawa K, Kanazawa A, Kramer DM. Depletion of stromal Pi induces high “energy dependent” antenna exciton quenching (qE) by decreasing proton conductivity at CF0-CF1 ATP synthase. Plant Cell Environ. 2008;31:235–243. doi:10.1111/j.1365-3040.2007.01753.x.
- Cruz JA, Avenson TJ, Kanazawa A, Takizawa K, Edwards GE, Kramer DM. Plasticity in light reactions of photosynthesis for energy production and photoprotection. J Exp Bot. 2005;56:395–406. doi:10.1093/jxb/eri022.
- Tikhonov AN. pH-dependent regulation of electron transport and ATP synthesis in chloroplasts. Photosynth Res. 2013;116:511–534. doi:10.1007/s11120-013-9845-y.
- Livingston AK, Cruz JA, Kohzuma K, Dhingra A, Kramer DM. An Arabidopsis mutant with high cyclic electron flow around photosystem I (hcef) involving the NADPH dehydrogenase complex. Plant Cell. 2010;22:221–233. doi:10.1105/tpc.109.071084.
- Sharkey TD, Weise SE. The glucose 6-phosphate shunt around the Calvin-Benson cycle. J Exp Bot. 2016;67:4067–4077. doi:10.1093/jxb/erv484.
- Li J, Weraduwage SM, Preiser AL, Tietz S, Weise SE, Strand DD, Froehlich JE, Kramer DM, Hu J, Sharkey TD. A cytosolic bypass and G6P shunt in plants lacking peroxisomal hydroxxypyruvate reductase. Plant Physiol. 2019;180:783–792. doi:10.1104/pp.19.00256.
- Carstensen A, Herdean A, Schmidt SB, Sharma A, Spetea C, Pribil M, Husted S. The impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant Physiol. 2018;177:271–284. doi:10.1104/pp.17.01624.
- Morales A, Yin X, Harbinson J, Driever SM, Molenaar J, Kramer DM, Struik PC. In silico analysis of the regulation of the photosynthetic electron transport chain in C3 plants. Plant Physiol. 2018;176:1247–1261. doi:10.1104/pp.17.00779.
- Li P, Weng J, Zhang Q, Yu L, Yao Q, Chang L, Niu Q. Physiological and biochemicial responses of Cucumis melo L. chloroplasts to low-phosphate stress. Front Plant Sci. 2018;9:Article 1525. doi:10.3389/fpls.2018.01525.
- Sharkey TD, Vanderveer PJ. Stromal phosphate concentration is low during feedback limited photosynthesis. Plant Physiol. 1989;91:679–684. doi:10.1104/pp.91.2.679.
- Kiirats O, Cruz JA, Edwards GE, Kramer DM. Feedback limitation of photosynthesis at high CO2 acts by modulating the activity of the chloroplast ATP synthase. Func Plant Biol. 2009;36:893–901. doi:10.1071/FP09129.
- Mukherjee P, Banerjee S, Wheeler A, Ratliff LA, Irigoyen S, Garcia LR, Lockless SW, Versaw WK. Live imaging of inorganic phosphate in plants with cellular and subcellular resolution. Plant Physiol. 2015;167:628–638. doi:10.1104/pp.114.254003.
- Kanno S, Cuyas L, Javot H, Bligny R, Gout E, Dartevelle T, Hanchi M, Nakanishi TM, Thibaud M-C, Nussaume L. Performance and limitations of phosphate quantification: guidelines for plant biologists. Plant Cell Physiol. 2016;57:690–706. doi:10.1093/pcp/pcv208.
- Yoshida T, Kakizuka A, Imamura H. BTeam, a novel BRET-based biosensor for the accurate quantification of ATP concentration within living cells. Sci Rep. 2016;6:39618. doi:10.1038/srep39618.
- Voon CP, Guan X, Sun Y, Sahu A, Chan MN, Gardeström P, Wagner S, Fuchs P, Nietzel T, Versaw WK, et al. ATP compartmentation in plastids and cytosol of Arabidopsis thaliana revealed by fluorescent protein sensing. PNAS. 2018;115:E10778–E10787. doi:10.1073/pnas.1711497115.
- Yamori W, Evans JR, Von Caemmerer S. Effects of growth and measurement light intensities on temperature dependence of CO2 assimilation in tobacco leaves. Plant Cell Environ. 2010;33:332–343. doi:10.1111/j.1365-3040.2009.02067.x.
- Schӧttler MA, Tόth SZ. Photosynthetic complex stoichiometry dynamics in higher plants: environmental acclimation and photosynthetic flux control. Front Plant Sci. 2014;5:Article 188.
- Yang JT, Preiser AL, Li Z, Weise SE, Sharkey TD. Triose phosphate use limitation of photosynthesis: short-term and long-term effects. Planta. 2016;243:687–698. doi:10.1007/s00425-015-2436-8.
- Hurry V, Å S, Furbank R, Stitt M. The role of inorganic phosphate in the development of freezing tolerance and the acclimatization of photosynthesis to low temperature is revealed by the pho mutants of Arabidopsis thaliana. Plant J. 2000;24:383–396. doi:10.1046/j.1365-313x.2000.00888.x.
- Nielsen TH, Krapp A, Röper-Schwarz U, Stitt M. The sugar-mediated regulation of genes encoding the small subunit of Rubisco and the regulatory subunit of ADP glucose pyrophosphorylase is modified by phosphate and nitrogen. Plant Cell Environ. 1998;21:443–454. doi:10.1046/j.1365-3040.1998.00295.x.
- Schöttler MA, Tóth SZ, Boulouis A, Kahlau S. Photosynthetic complex stoichiometry dynamics in higher plants: biogenesis, function, and turnover of ATP synthase and cytochrome b6f complex. J Exp Bot. 2015;66:2373–2400. doi:10.1093/jxb/eru495.
- Benz M, Bals T, Gügel IL, Piotrowski M, Kuhn A, Schünemann D, Soll J, Ankele E. Alb4 of Arabidopsis promotes assembly and stabilization of a non chlorophyll-binding photosynthetic complex, the CF1CF0-ATP synthase. Mol Plant. 2009;2:1410–1424. doi:10.1093/mp/ssp095.
- Rühle T, Razeghi JA, Vamvaka E, Viola S, Gandini C, Kleine T, Schünemann D, Barbato R, Jahns P, Leister D. The Arabidopsis protein CONSERVED ONLY IN THE GREEN LINEAGE160 promotes the assembly of the membranous part of the chloroplast ATP synthase. Plant Physiol. 2014;165:207–226. doi:10.1104/pp.114.237883.
- Fristedt R, Martins NF, Strenkert D, Clarke CA, Suchoszek M, Thiele W, Schöttler MA, Merchant SS. The thylakoid membrane protein CGL160 supports CF1CF0 ATP synthase accumulation in Arabidopsis thaliana. PLoS One. 2015;10:e0121658. doi:10.1371/journal.pone.0121658.
- Hoshiyasu S, Kohzuma K, Yoshida K, Fujiwara M, Fukao Y, Yokota A, Akashi K. Potential involvement of N-terminal acetylation in the quantitative regulation of the Ε subunit of chloroplast ATP synthase under drought stress. Biosci Biotechnol Biochem. 2013;77:998–1007. doi:10.1271/bbb.120945.
- Linster E, Wirtz M. N-terminal acetylation: an essential protein modification emerges as an important regulator of stress responses. J Exp Bot. 2018;69:4555–4568.