
ABSTRACT
slPHB3 was cloned from Salix linearistipularis, the amino acid sequence blast and phylogenetic tree analysis showed that slPHB3 has the most similarity with PHB3 from Populus trichocarpa using DNAMAN software and MEGA7 software. RT-qPCR results confirmed that the expression of slPHB3 was induced obviously under stress treatments. The growth of recombinant yeast cells was better than that of the control group under the stress treatment, indicating that slPHB3 may be involved in the stress response of yeast cells. The transgenic tobacco was treated with different concentrations of NaCl, NaHCO3 and H2O2, fresh weigh of overexpression tobacco were heavier than wild-types. The results showed that transgenic tobacco was more tolerant to salt and oxidation than wild-type tobacco. Expression of important genes including NHX1 and SOS1 in salt stress response pathways are steadily higher in overexpression tobacco than that in wild-types. We identified 17 proteins interacting with slPHB3 by yeast two-hybrid technique, most of these proteins were relation to the stresses. The salt tolerance of slPHB3 expressing yeast and slPHB3 overexpressing plants were better than that of the control. Ten stress-related proteins may interact with slPHB3, which preliminarily indicated that slPHB3 had a certain response relationship with salt stress. The study of slPHB3 under abiotic stress can improve our understanding of PHB3 gene function.
Introduction
Plants have developed specific mechanisms that allow them to detect precise environmental changes and respond to complex stress conditions.Citation1 Salix linearistipularis (syn. S. mongolica) is a woody plant that is found in Songnen plain, Heilongjiang, China.Citation2 Salix linearistipularis resistant to saline and plays an important role in maintaining ecological balance and improving saline soil.Citation3
PHB was first discovered as a tumor suppressor gene in mammalian cells.Citation4 PHB genes are conserved during evolution.Citation5,Citation6 PHB protein regulates plant growth, regulate membrane protein degradation, control cell proliferation and apoptosis.Citation7,Citation8 PHB3 impacts SA accumulation.Citation9 PHB3 knockout mutants show severe growth defects and have decreased cell division and expansion in the root apex.Citation10 PHB3 regulates stem cell niche maintenance and cell proliferation during root development in Arabidopsis.Citation11 Germination of atPHB3 KO seeds was delayed by 1–2 days compared with wild-type.Citation12 Methylo bacterium has better survival ability under osmotic stress, which is related to the accumulation of PHB in the strain.Citation13In Arabidopsis thaliana, PHB3 coordinates cell division and differentiation in root tip meristems by restricting the ethylene reactive factor transcription factor.Citation14 The growth of wildtype was different from that of the PHB3 mutant under NaCl or H2O2 treatment.Citation10
Salinity stress effect on plant can be reduced by maintaining the appropriate Na+ ions concentration in the cytosol, NHX1 (vacuolar Na+/H+ antiporter) and SOS1 (Salt Overly-Sensitive-1, plasma membrane Na+/H+ antiporter 1) are a set of genes that function in ion homeostasis in plants by regulating Na+ ions in the cytosol. NHX1 are present in tonoplasts and decrease the accumulation of Na+ ions in the cytosol by pumping Na+ in the vacuole to sequester Na+ in vacuoles, SOS1 is located at the plasma membrane and is responsible for efflux Na+ from cytosol to apoplasts,Citation15 NHX1and SOS1 might be used as biomarkers to further investigate the interaction between gene and stress tolerance.Citation16 SpSOS1 transgenic plants might have a high capacity for removing ROS under salt stress.Citation17 TdSOS1 transgenic plants have higher ROS scavenging enzymes activity than that in control plants under oxidative stress.Citation18 Co-expression of Na+/H+ antiporter NHX1 and DEAD-box RNA helicase eIF4A1 from Arabidopsis positively regulates drought stress tolerance by improving ROS scavenging capacity.Citation19
In this study, slPHB3 was cloned from Salix linearistipularis, then transferred into yeast and tobacco genome. The results showed that transgenic tobacco plants had stronger salt tolerance and oxidation resistance than wild-type tobacco. Seventeen positive clones were obtained using yeast two-hybrid assay to further explore the interaction protein with slPHB3. slPHB3 enhanced the stress resistance of transgenic tobacco plants and provided a basis for the study of PHB3 introduction in other valuable plants.
Result
Cloning and bioinformatics analysis of slPHB3 gene
The open reading frame sequence of the slPHB3 gene was cloned from Salix linearistipularis and sequenced. PHB3 protein of other plants was compared with slPHB3 by DNAMan software. The amino acid sequence of slPHB3 has a high degree similarity with the amino acid sequence of other plants PHB3 (). Therefore, the cDNA cloned from Salix linearistipularis is named as slPHB3.
Figure 1. Alignment of the slPHB3 amino acid sequence with PHB3 proteins from other plant species. The amino acid sequence is similar to that of CsPHB3 Protein (XP_006469979.1), PtPHB3 Protein (XP_002323792.1), PePHB3 Protein (XP_011045196.1), PaPHB3 protein (TKR74705.1), HbPHB3 Protein (XP_021677719.1), RcPHB3 protein(XP_002509571.1).
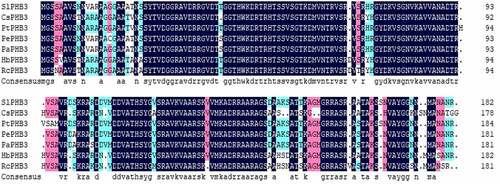
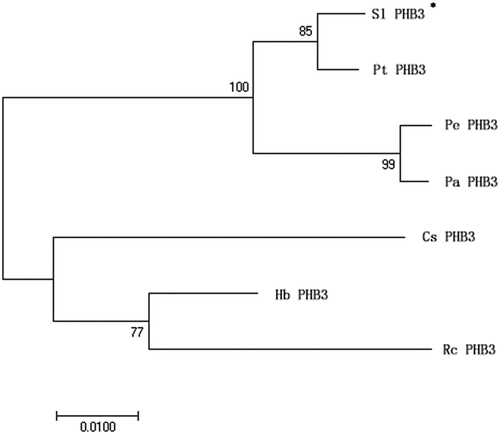
In order to explore the amino acid sequence homology between slPHB3 and other plant PHB3, we constructed a phylogenetic tree (), which showed that slPHB3 is closely related to PHB3 from Populus trichocarpa, Populus euphratica, and Populus alba.
Figure 2. slPHB3 evolutionary tree analysis. The MEGA7 program was used for the construction of phylogenetic trees. Bar represents 0.1 amino acid substitutions per site.
Expression of slPHB3 in salix linearistipularis under abiotic stresses
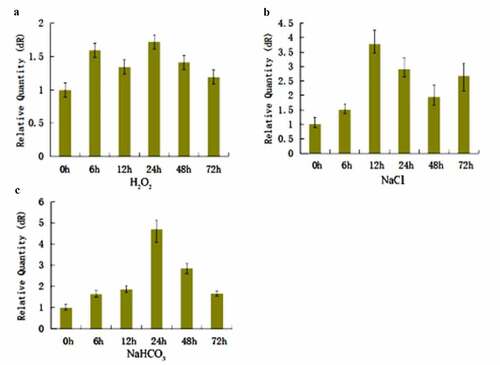
The slPHB3 expression level in the Salix linearistipularis leaves increased until the highest level at 24 h and then decreased gradually under 3 mM H2O2 and 5mM NaHCO3 stress (). The slPHB3 expression level showed the highest value at 12 h under 125 mM NaCl stress (). That means that the stresses influenced the slPHB3 gene expression level in Salix linearistipularis leaves.
Figure 3. Real-time quantitative PCR analysis for slPHB3 expression in Salix linearistipularis under different stresses. (a): Relative expression of slPHB3 at different time under 3 mM H2O2 stress; (b): Relative expression of slPHB3 at different time under 125 mM NaCl stress; (c): Relative expression of slPHB3 at different time under 5 mM NaHCO3 stress.
Expression of slPHB3 gene in transgenic yeast in response to stresses
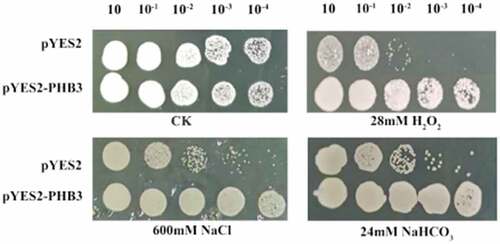
The yeast cells growth under various abiotic stresses were studied (). The slPHB3-transgenic lines grew normally on the medium, while the control plants did not growth in the presence of 1 mM NaCl, 24 mM NaHCO3 or 3.2 mM H2O2. The slPHB3-transgenic lines grew better than the control.
Figure 4. Growth of slPHB3 transgenic yeast cells under salt stress. Ten-fold dilutions of yeast cells containing pYES2 (upper line) and pYES2-slPHB3 vector (lower line) were spotted onto solid YPG media supplemented with the indicated stresses. No treatment is a control (CK).
Stress tolerance of the transgenic tobacco
The plants grew well in the medium without stress. The transgenic plants severely lost water and wilted than wild type after stress treatment (). There was no significant difference in fresh weight under CK conditions. To the opposite, under stress treatments, the fresh weight of over-expressed plants was larger than those of wild type, which generally grew better and suffered less damage ().
Table 1. Fresh weigh (mg) of tobacco under different stress treatments. Results are present as means of 10 plants. CK, control; WT, wild type; #2, #3 and #4 are slPHB3 transgenic plants
Figure 5. The relative stress tolerance of wild-type and transgenic plants (2, # 3, # 4) in reproduction stage was studied. The plants grown on the soil with 1.5 M H2O2, 300 mM NaCl or 300 mM NaHCO3 or without (CK).

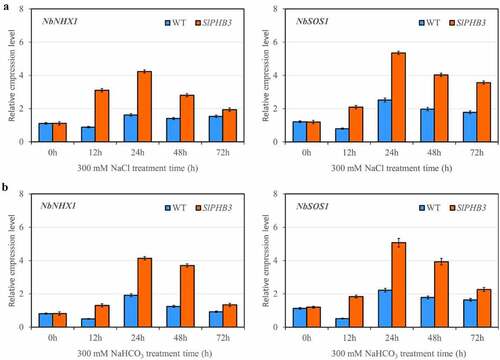
Expression of salt stress-related genes(NHX1 and SOS1) in tobacco
In order to further investigate the mechanisms of slPHB3 overexpression plants enhanced salt tolerance, expression levels of salt-related marker genes (NHX1 and SOS1) in salt response pathways were determined. Under salt condition, NHX1 and SOS1 showed steadily higher expression in overexpression lines than in WT ().
Figure 6. Quantitative qRT-PCR determination of representative gene expression in leaves. Relative expression level of slPHB3 in leaves of wildtype tobacco and transgenic tobacco seedlings exposed to 300 mM NaCl (a) or 300 mM NaHCO3 (b), values are mean ± SD, n = 3. Tublin was used as an internal control.
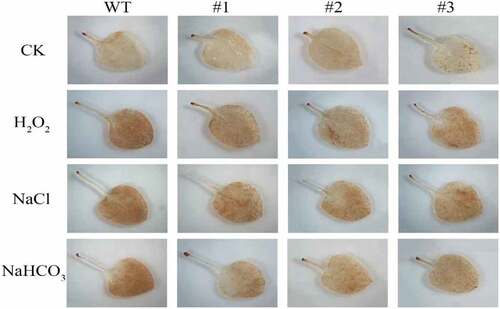
Effects of treatments on H2O2 content in the transgenic tobacco
Hydrogen peroxide in tobacco leaves was detected in situ using 3, 3′-Diaminobenzidine (DAB) histochemical staining method. The color of the leaf showed no difference between WT and slPHB3-transgenic plants without stresses. The color of the WT leaf was darker than that of the transgenic line leaves under different stress (). H2O2 accumulation in wildtype or transgenic plants was detected under stress conditions. The H2O2 content in the transgenic line was lower than that of the WT plant after 48 h treatment () which indicated that slPHB3 increased the H2O2 scavenging function of the transgenic plants.
Table 2. H2O2 content in leaves of wild type and transgenic Arabidopsis under different stresses was measured with Plant H2O2 Kit. Data are means of three replicates. WT, wild type; #2, #3 and #4 are slPHB3 transgenic plants
Figure 7. H2O2 accumulation in leaves was detected by DAB staining. DAB staining in leaves in wild type and transgenic plants (2, # 3, # 4) under 1.5 M H2O2, 300 mM NaCl or 300 mM NaHCO3 stresses.
Screening of slPHB3 interacting protein by yeast two-hybrid
The total of 17 positive gene were obtained from colonies grown on SD/-Trp-Leu-His-Ade+X-α-gal+AbA solid medium turned blue. BLAST these colonies in NCBI, 17 genes were successfully compared (), ATP-dependent protease La regulates protein stability, Lon hydrolyzed protein impaired seedling production. The glycosyl hydrolase is related to low temperature stress and osmotic stress, P-loop containing nucleoside triphosphate hydrolases, HSP and PAO4 are related to temperature stress. ATP synthase subunit beta is related to cold stress and oxidative stress. ATP synthase epsilon chain and TIF7 are related to salt stress, NAC13 is related to salt and drought stress. Glycine decarboxyla plays a major role in photorespiration. Non-intrinsic ABC protein family functions are related to accumulation of metal ions and stability of chloroplast structure. ATNAP7 is an essential ATP binding site for embryo development. PAO5 is highly responsive to drought stress. Salt induced the expression of SOT12 gene. Tom3 have a specific role in the transport of plant iron carriers. Proline-rich nuclear receptor modulates transcriptional activation of multiple nuclear receptors.
Materials and methods
Plant materials and growth conditions
Salix linearistipularis and tobacco plants were grown in a controlled growth chamber at 24 ± 2°C with a 16 h light/8 h dark cycle.
Cloning and bioinformatics analysis of slPHB3 gene
Total RNA from leaves of Salix linearistipularis was isolated using RNeasy Plant Mini Kit (Qiagen, Hilden, Germany), first-strand cDNA was synthesized by reverse transcribing 500 ng of total RNA with using Prime-Script Reverse Transcriptase (Takara, Tokyo, Japan). The forward slPHB3F and reverse primer slPHB3R (slPHB3F: ATGGGTAGCAGCCAAGCAGC, slPHB3R: TCAACGGTTTGCATTCAGGG) were designed according to the Salix linearistipularis transcriptome data. The PCR products were ligated to pMD18-T vector (Takara, Tokyo, Japan) and sequenced. The homologous amino acid sequence of slPHB3 protein were compared using DNAMAN software, and the phylogenetic tree was constructed using MEGA7 software.
Real-time quantitative PCR (RT-qPCR) analysis for slPHB3 expression
Salix linearistipularis seeds were sown onto 1/2 MS medium. The seedlings of one-month age were exposed to 3 mM H2O2, 150 mM NaCl and 5 mM NaHCO3 treatments for 0, 6, 12, 24, 36 or 72 h, respectively. The slPHB3 expression in leaves under treatments was examined by RT-qPCR analysis. Total RNA was isolated from leaves of Salix linearistipularis and cDNA was synthesized. Subsequently RT-qPCR analyses were carried out by SYBR green (Takara, Tokyo, Japan) and IQ5 real-time PCR equipment (Bio-Rad, Hercules, CA, USA) with the steps: 95°C for 30s, 30 cycles of 95°C for 5s, and 55°C for 30s. The next steps were added to meltcurve analysis: 95°C for 15s, followed by continuously increased from 60°C to 95°C. The SaActin gene expression was used as control. The forward primer sequence was SaActin-F: GGTAACATTGTGCTCAGTGGTGG and reverse primer sequence was SaActin-R: AACGACCTTAATCTTCATGCTGC. All tests were repeated in triplicate.
Construction of yeast expression vectors and transformation
The coding region of the slPHB3 gene was PCR amplified with BamHI sense primer 5’- GGATCCATGGGTAGCAGCCAAGCAGC-3’ and XhoI antisense primer 5’-CTCGAG TCAACGGTTTGCATTCAGGG −3’. The PCR amplified fragments were digested with BamHI and XhoI, then subcloned to the same site of the pYES2 expression vector (Clontech, Tokyo, Japan) resulting in pYES2-slPHB3. Then transformed into the competent yeast strain INVSC1 for protein expression in Yeast. Transgenic yeast cells (OD600 = 0.5) containing pYES2-slPHB3 and pYES2 (control) with serial dilutions (10, 10−1, 10−2, 10−3 and 10−4) were spotted onto YPD agar plates supplemented with 3.2 mM H2O2, 1 M NaCl or 24 mM NaHCO3, respectively.
Acquisition of slPHB3 overexpressed tobacco transgenic lines
The slPHB3 PCR amplified fragment with added with BamH I and Xho I restriction enzymes was obtained by slPHB3BamH I-F and reverse primer slPHB3Xho I-R and ligase into pBI121 vector plasmid, the plasmid DNAs of pBI121-slPHB3 was transformed into the Agrobacterium tumefaciens strain EHA105 (Takara, Tokyo, Japan), and the tobacco was infected with agrobacterium. The independent transgenic lines were obtained and verified the expression of slPHB3 by RT-qPCR, all temples were tested in triplicate, then used for further analyses.
Wild type and transgenic seed were planted to pots containing nutrient-rich soil. After growing up for two months, the pots were irrigated with 50 mL solution of 1.5 M H2O2, 300 mM NaCl or 300 mM NaHCO3 every 4 days. The pots were covered with a breathable plastic cover to minimize evaporation and keep the concentration of the solution from changing too much. Images of the plants were taken after 12 days of treatment.
RT-qPCR analysis for NHX1 and SOS1 expression in tobacco
The expression levels of the NHX1 (Accession number: JX987081.1), and SOS1(Accession number: JK739016.1) were analyzed using qRT-PCR, Total RNA was isolated from leaf samples of 300 mM NaCl or 300 mM NaHCO3 treated tobacoo plants. The relative levels of the transcript accumulated for the NHX1 (NbNHX1qPCR -F: 5′ - AGGCACGCTTTTGCAACTTT -3ʹ,NbNHX1qPCR-R:5ʹ-ACACTGCGCCTCTCATCAAA-3ʹ), and SOS1 genes (NbSOS1qPCR-F:5ʹ-Acagctagtttttctcatcaa-3ʹ, NbSOS1qPCR-R: 5ʹ-tgaccagttcgttccacatc-3ʹ) were normalized to actin (NbActin-F: 5ʹ-tggtcgtaccaccggtattgtgtt-3ʹ, NbActin-R:5ʹ- tcacttgcccatcaggaagctcat-3ʹ).Citation20
Reaction to H2O2 stress in transgenic tobacco plants
One-month-old WT and transgenic plants were treated without (control) or with each of 1.5 M H2O2, 300 mM NaCl or 300 mM NaHCO3 respectively for 48 h. H2O2 accumulation in plant leaves was visualized by histochemical staining with 3, 3′-Diaminobenzidine (DAB). The treated leaves were immersed in 1 mg·mL−1 DAB solution (Solarbio, Beijing, China), incubated in the dark at room temperature, after DAB solution incubated, the samples chlorophyll was removed using absolute ethanol, the leaf was photographed with a microscopy (Olympus). The H2O2 content was also measured using Plant H2O2 ELISA Kit (America Rapid Bio).
Screening of slPHB3 interacting protein by yeast two-hybrid
slPHB3 was cloned into pGADT7 vector, the recombinant plasmids pGADT7-slPHB3 were identified by double enzyme digestion, then they were transformed into Y2HGold strain, the proteins interacting with pGADT7-slPHB3 were screened from Salix linearistipularis cDNA library. Yeast DNA was extracted and sequenced to obtain the interaction gene and protein sequences.
Discussion
A slPHB3 gene was cloned from Salix linearistipularis to develop the Salix linearistipularis resistance mechanism. The slPHB3 expression level increased under 3 mM H2O2, 125 mM NaCl and 5 mM NaHCO3 stress (). This means the stresses influenced the slPHB3 gene expression level in Salix linearistipularis leaves.
In the yeast resistance analysis, the growth of the transgenic strain was better than that of the control under 1 mM NaCl, 24 mM NaHCO3 or 3.2 mM H2O2 medium treatment, indicating that slPHB3 gene expression increased yeast resistant to stress. Yeast belongs to Eukaryote, which shows that slPHB3 expression in yeast can improve the saline resistance of eukaryote.
Under the 2 M H2O2, 300 mM NaCl or 300 mM NaHCO3 stress, the fresh weight of over-expressed plants was larger than those of wild type, the wild-type plants died, while the transgenic plants survived (). The results showed that overexpression slPHB3 can improved tobacco stresses resistance.
Molecular mechanisms underlying slPHB3 functioning under salt conditions have not been completely elucidated, NHX1 and SOS1 expression to regulate Na+ ions toxicity that leads to enhanced salinity stress tolerance in tomato plants,Citation21 BvSOS1 and BvNHX1 played vital roles in tetraploid sugar beet cultivar which exhibited more tolerant to salinity than diploid sugar beet,Citation22 two pomegranate cultivars study finding that difference between cultivars in salt tolerance is associated with transcriptional regulation of SOS1 and NHX1 genes.Citation23 In this study, we analyzed the expression of two salt-related marker gene (NHX1 and SOS1) in tobacco, both NHX1 and SOS1 expression higher in overexpression lines than that in WT (), NHX1 and SOS1 have a high capacity for scavenging ROS under salt stress.Citation17–19 The increased transcript level of NHX1 and SOS1 in overexpression plants under salinity stress may increase plant tolerance to salt by improving ROS scavenging capacity.
In important number of ROS is produced under the stresses.Citation24 Upon PHB3 loss-of-function, the ROS contents will be out of homeostasis.Citation11 Compared to WT, H2O2 was accumulated more in the PHB3 mutant root meristem.Citation14 These study showed that PHB3 was relation to ROS contents in plant. In this study, the content of ROS in overexpressing slPHB3 tobacco was significantly higher than that in wild-type tobacco. The results showed that tobacco overexpressing slPHB3 could reduce the content of ROS and eliminate the effect of excessive ROS on plants to improved tobacco stresses resistance.
PHB3 also interacts with a variety of other proteins. In mitochondria, PHB3 forms complexes with other PHB proteins. The formation of the atPHB3-ICS1 complex stabilizes ICS1 to promote SA production.Citation9 In this research, 17 sequences were successfully matched (). The function of P-loop containing nucleoside triphosphate hydrolases is related to temperature changes.Citation25 The function of glycosyl hydrolase family is related to glycosylation.Citation26 The glycosyl hydrolase 18 (GH18) and GH19 families are also related to low temperature stress and osmotic stress,Citation27 ATP-dependent protease La is involved in the degradation of abnormally folded proteins and specific regulatory proteins, and regulates protein stability.Citation28 Lack of Lon hydrolyzed protein causes plant growth retardation and impaired seedling production.Citation29 ATP Synthase subunit Beta family is involved in REDOX reactions, mediates protein interactions, and is associated with cold stress.Citation30,Citation31 Proteome analysis of Date Palm showed that the α and β subunits of ATP synthase changed significantly under salt stress and drought stress.Citation32 Alpha/beta-hydrolases superfamily functions to catalyze the hydrolysis of ester bonds between fatty acids and glycerol.Citation33 It was involved in delaying the senescence of strawberry fruit at low temperature.Citation34 HSP family functions are related to temperature changes.Citation35 However, under NaCl stress, the root-cap length of OSHSP40 transgenic seedlings was significantly shorter than that of wild-type seedlings. The results showed that HSP40 was related to salt stress.Citation36 The TIFY family may play crucial and divergent roles in phytohormone crosstalk and plant defense.Citation37 The TIFY gene responds to abiotic stresses such as jasmonic acid (JA) and salt and drought.Citation38 Glycine decarboxyla plays a major role in photorespiration. Glycine decarboxylase and other proteins increase CO2 assimilation, vegetative biomass, and seed yield in Arabidopsis thaliana. AtGLDP1 is involved in the transition of C-3 through C-2 to C4 photosynthesis.Citation39 Non-intrinsic ABC protein family functions are related to accumulation of metal ions and stability of chloroplast structure.Citation40 ATNAP7 is an essential ATP binding site for Arabidopsis embryo development.Citation41 PAO family functions play a major role in PA catabolism. ScPAO5 is highly responsive to drought stress, while ScPAO1 and CspAO2 are sensitive to changes in nitrogen nutrition.Citation42 Transcription level of PaO4 in tomato leaves increased in response to heat stress and cold stress.Citation43 Glycoprotein effect on rice seedlings salt tolerance.Citation44 loss of NatB function increased plant sensitivity toward osmotic and high-salt stress, indicating that NatB is required for tolerance of these abiotic stress.Citation45 Salt, osmotic stress and hormone treatment strongly induced the expression of Atsot12 gene.Citation46 Tom3 is thought to have a specific role in the transport of plant iron carriers.Citation47 Proline-rich nuclear receptor coactivator participates regulatory protein that modulates transcriptional activation of multiple nuclear receptors.Citation48
Table 3. Information on the putative slPHB3-interacting proteins identified by the Y2H system
Some NAC genes have been identified as candidates for breeding programmer to improve drought resistance in crops.Citation49 In maize, 13 SNAC transcripts in the SNAC subfamily were responsive to drought stress, and almost all of them in roots and 11 in leaves were upregulated under drought stress.Citation50 The NAC-type transcription factor CaNAC46 from Capsicum annuum regulates the salt and drought tolerance of transgenic Arabidopsis thaliana.Citation51
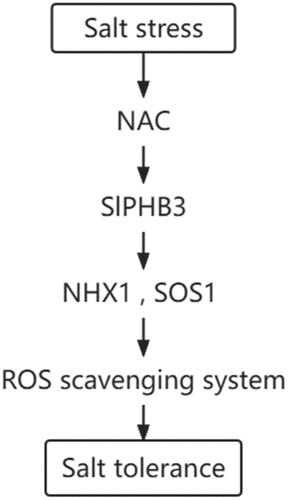
In our study, H2O2 content were significantly lower in slPHB3-overexpressing tobacco lines comparing to those in WT plants under salt stresses, overexpression of slPHB3 likely improved salt tolerance through the ROS scavenging system pathways. We identified NAC proteins interacting with slPHB3 by yeast two-hybrid technique. Consistent with these findings, we hypothesized a molecular pathway that the increase of NAC gene expression under salt treatment. NAC might induce the increase of slPHB3 expression, slPHB3 may increase the expression of NHX1 and SOS1, NHX1 and SOS1 improved ROS scavenging capacity. In this pathway, slPHB3 enhanced the scavenging effect on ROS and improved plant tolerance ().
Figure 8. Model of the regulatory mechanisms of the slPHB3 gene in response to the tolerance to salt stresses. Salt stress increased expression of NAC, NAC might induce the increase of slPHB3 expression, slPHB3 induceed the increase of NHX1 and SOS1 expression, NHX1 and SOS1 genes enhanced the scavenging effect on ROS, which resulted in improved tolerance to salt stresses.
Conclusion
The growth of recombinant yeast cells was better than that of the control group under the stress treatment. Comparison between slPHB3 transgenic tobacco and wild-type tobacco showed that the transgenic plants had higher salt tolerance and oxidation resistance than the wild-type plants. Expression of NHX1 and SOS1 in transgenic tobacoo are higher in that in wild-types. Through the screening of slPHB3 interaction proteins, 10 of the 17 genes were related to abiotic stress, indicating that PHB3 gene plays a role in plant stress resistance.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Atkinson NJ, Urwin PE. The interaction of plant biotic and abiotic stresses: from genes to the field. J Exp Bot. 2012;63(10):3523–10. doi:10.1093/jxb/ers100.
- Ishida TA, Nara K, Ma SR, Takano T, Liu SK. Ectomycorrhizal fungal community in alkaline-saline soil in northeastern China. Mycorrhiza. 2009;19(5):329–335. doi:10.1007/s00572-008-0219-9.
- Nan GX, Zhang Y, Li S, Lee I, Takano T, Liu SK. NaCl stress-induced transcriptomics analysis of Salix linearistipularis (syn. Salix mongolica). J Biol Res. 2016;23:14. doi:10.1186/s40709-016-0038-7.
- McClung JK, Danner DB, Stewart DA, Smith JR, Schneider EL, Lumpkin CK, Dell’Orco RT, Nuell MJ. Isolation of a cDNA that hybrid selects antiproliferative mRNA from rat liver. Biochem Biophys Res Commun. 1989;164(3):1316–1322. doi:10.1016/0006-291x(89)91813-5.
- Di C, Xu WY, Su Z, Yuan JS. Comparative genome analysis of PHB gene family reveals deep evolutionary origins and diverse gene function. BMC Bioinform. 2010;11(S6):15. doi:10.1186/1471-2105-11-s6-s22.
- Thuaud F, Ribeiro N, Nebigil CG, Desaubry L. Prohibitin ligands in cell death and survival: mode of action and therapeutic potential. Chem Biol. 2013;20(3):316–331. doi:10.1016/j.chembiol.2013.02.006.
- Merkwirth C, Dargazanli S, Tatsuta T, Geimer S, Lower B, Wunderlich FT, von Kleist-retzow JC, Waisman A, Westermann B, Langer T. Prohibitins control cell proliferation and apoptosis by regulating OPA1-dependent cristae morphogenesis in mitochondria. Genes Dev. 2008;22(4):476–488. doi:10.1101/gad.460708.
- Merkwirth C, Langer T. Prohibitin function within mitochondria: essential roles for cell proliferation and cristae morphogenesis. Biochim Biophys Acta-Mol Cell Res. 2009;1793(1):27–32. doi:10.1016/j.bbamcr.2008.05.013.
- Seguel A, Jelenska J, Herrera-Vasquez A, Marr SK, Joyce MB, Gagesch KR, Shakoor N, Jiang SC, Fonseca A, Wildermuth MC, et al. PROHIBITIN3 forms complexes with ISOCHORISMATE SYNTHASE1 to regulate stress-induced salicylic acid biosynthesis in arabidopsis. Plant Physiol. 2018;176(3):2515–2531. doi:10.1104/pp.17.00941.
- Wang Y, Ries A, Wu KT, Yang A, Crawford NM. The arabidopsis prohibitin gene phb3 functions in nitric oxide-mediated responses and in hydrogen peroxide-induced nitric oxide accumulation. Plant Cell. 2010;22(1):249–259. doi:10.1105/tpc.109.072066.
- Huang RH, Yang CW, Zhang SC. The Arabidopsis PHB3 is a pleiotropic regulator for plant development. Plant Signal Behav. 2019;14(11):5. doi:10.1080/15592324.2019.1656036.
- Van Aken O, Pecenkova T, van de Cotte B, De Rycke R, Eeckhout D, Fromm H, De Jaeger G, Witters E, Beemster GTS, Inze D, et al. Mitochondrial type-I prohibitins of Arabidopsis thaliana are required for supporting proficient meristem development. Plant J. 2007;52(5):850–864. doi:10.1111/j.1365-313X.2007.03276.x.
- Woo SP, Ramasamy K, Joe MM, Sa T. EPS production, PHB accumulation and abiotic stress endurance of plant growth promoting methylobacterium strains grown in a high carbon concentration. Korean Journal of Soil Science & Fertilizer. 2012;45(4):572–581. doi:10.7745/kjssf.2012.45.4.572.
- Kong XP, Tian HY, Yu QQ, Zhang F, Wang R, Gao S, Xu WH, Liu JJ, Shani E, Fu CX, et al. PHB3 Maintains Root Stem Cell Niche Identity through ROS-Responsive AP2/ERF Transcription Factors in Arabidopsis. Cell Rep. 2018;22(5):1350–1363. doi:10.1016/j.celrep.2017.12.105.
- Zhang WD, Wang P, Bao ZLT, Ma Q, Duan LJ, Bao AK, Zhang JL, Wang SM. SOS1, HKT1;5, and NHX1 synergistically modulate Na+ homeostasis in the halophytic grass puccinellia tenuiflora. Front Plant Sci. 2017a;8:8. doi:10.3389/fpls.2017.00576.
- Chen CZ, Lv XF, Li JY, Yi HY, Gong JM. Arabidopsis NRT1.5 is another essential component in the regulation of nitrate reallocation and stress tolerance. Plant Physiol. 2012;159(4):1582–1590. doi:10.1104/pp.112.199257.
- Zhou Y, Yin XC, Wan SM, Hu YP, Xie Q, Li RM, Zhu BB, Fu SP, Guo JC, Jiang XY. The sesuvium portulacastrum plasma membrane Na+/H+ Antiporter SpSOS1 complemented the salt sensitivity of transgenic arabidopsis sos1 mutant plants. Plant Mol Biol Rep. 2018;36(4):553–563. doi:10.1007/s11105-018-1099-6.
- Feki K, Tounsi S, Masmoudi K, Brini F. The durum wheat plasma membrane Na+/H+ antiporter SOS1 is involved in oxidative stress response. Protoplasma. 2017;254(4):1725–1734. doi:10.1007/s00709-016-1066-8.
- Zhang YD, Deng GF, Fan WJ, Yuan L, Wang HX, Zhang P. NHX1 and eIF4A1-stacked transgenic sweetpotato shows enhanced tolerance to drought stress. Plant Cell Rep. 2019;38(11):1427–1438. doi:10.1007/s00299-019-02454-6.
- Ha JH, Jang HA, Moon KB, Baek KH, Choi GJ, Choi D, Cho HS, Kwon SY, Jeon JH, Oh SK, et al. Nicotiana benthamiana matrix metalloprotease 1 (NMMP1) gene confers disease resistance to Phytophthora infestans in tobacco and potato plants. J Plant Physiol. 2017;218:189–195. doi:10.1016/j.jplph.2017.08.010.
- Rao YR, Ansari MW, Sahoo RK, Wattal RK, Tuteja N, Kumar VR. Salicylic acid modulates ACS, NHX1, sos1 and HKT1;2 expression to regulate ethylene overproduction and Na+ ions toxicity that leads to improved physiological status and enhanced salinity stress tolerance in tomato plants cv. Pusa Ruby Plant Signal Behav. 2021;16(11):9. doi:10.1080/15592324.2021.1950888.
- Li SJ, Wu GQ, Lin LY. AKT1, HAK5, SKOR, HKT1;5, SOS1 and NHX1 synergistically control Na+ and K+ homeostasis in sugar beet (Beta vulgaris L.) seedlings under saline conditions. J Plant Biochem Biotechnol. 2021;14. doi:10.1007/s13562-021-00656-2.
- Calzone A, Cotrozzi L, Pellegrini E, Lorenzini G, Nali C, Maathuis F. Can the transcriptional regulation of NHX1, SOS1 and HKT1 genes handle the response of two pomegranate cultivars to moderate salt stress? Salt-tolerance of two pomegranate cultivars. Sci Hortic. 2021;288:8. doi:10.1016/j.scienta.2021.110309.
- Jubany-Mari T, Munne-Bosch S, Alegre L. Redox regulation of water stress responses in field-grown plants. Role of hydrogen peroxide and ascorbate. Plant Physiol Biochem. 2010;48(5):351–358. doi:10.1016/j.plaphy.2010.01.021.
- Zhao L, Vecchi G, Vendruscolo M, Korner R, Hayer-Hartl M, Hartl FU. The Hsp70 chaperone system stabilizes a thermo-sensitive subproteome in E. coli. Cell Rep. 2019;28(5):1335-+. doi:10.1016/j.celrep.2019.06.081.
- Kotik M, Brodsky K, Halada P, Javurkova H, Pelantova H, Konvalinkova D, Bojarova P, Kren V. Access to both anomers of rutinosyl azide using wild-type rutinosidase and its catalytic nucleophile mutant. Catal Commun. 2021;149:5. doi:10.1016/j.catcom.2020.106193.
- Chen JJ, Piao YL, Liu YM, Li XN, Piao ZY. Genome-wide identification and expression analysis of chitinase gene family in Brassica rapa reveals its role in clubroot resistance. Plant Sci. 2018;270:257–267. doi:10.1016/j.plantsci.2018.02.017.
- Tsitsekian D, Daras G, Alatzas A, Templalexis D, Hatzopoulos P, Rigas S. Comprehensive analysis of Lon proteases in plants highlights independent gene duplication events. J Exp Bot. 2019;70(7):2185–2197. doi:10.1093/jxb/ery440.
- Rigas S, Daras G, Tsitsekian D, Hatzopoulos P. The multifaceted role of Lon proteolysis in seedling establishment and maintenance of plant organelle function: living from protein destruction. Physiol Plant. 2012;145(1):215–223. doi:10.1111/j.1399-3054.2011.01537.x.
- Yang JH, Williams D, Kandiah E, Fromme P, Chiu PL. Structural basis of redox modulation on chloroplast ATP synthase. Commun Biol. 2020;3(1):12. doi:10.1038/s42003-020-01221-8.
- Zhang ZY, Li JJ, Pan YH, Li JL, Zhou L, Shi HL, Zeng YW, Guo HF, Yang SM, Zheng WW, et al. Natural variation in CTB4a enhances rice adaptation to cold habitats. Nat Commun. 2017b;8:13. doi:10.1038/ncomms14788.
- El Rabey HA, Al-Malki AL, Abulnaja KO, Rohde W. Proteome analysis for understanding abiotic stress (salinity and drought) tolerance in date palm (Phoenix dactylifera L.). Int J Genomics. 2015;2015:11. doi:10.1155/2015/407165.
- Zan XY, Cui FJ, Sun JN, Zhou S, Song YD. Novel dual-functional enzyme lip10 catalyzes lipase and acyltransferase activities in the oleaginous fungus Mucor circinelloides. J Agric Food Chem. 2019;67(47):13176–13184. doi:10.1021/acs.jafc.9b05617.
- Xu XB, Ma XY, Lei HH, Yin LL, Shi XQ, Song HM. MicroRNAs play an important role in the regulation of strawberry fruit senescence in low temperature. Postharvest Biol Technol. 2015;108:39–47. doi:10.1016/j.postharvbio.2015.05.006.
- Li GL, Zhao H, Guo HB, Wang Y, Cui XP, Li H, Xu BH, Guo XQ. Analyses of the function of DnaJ family proteins reveal an underlying regulatory mechanism of heat tolerance in honeybee. Sci Total Environ. 2020a;716:15. doi:10.1016/j.scitotenv.2020.137036.
- Wang X, Zhang H, Shao LY, Yan X, Peng H, Ouyang JX, Li SB. Expression and function analysis of a rice OsHSP40 gene under salt stress. Genes Genom. 2019;41(2):175–182. doi:10.1007/s13258-018-0749-2.
- Liu X, Zhao CB, Yang LM, Zhang YY, Wang Y, Fang ZY, Lv HH. Genome-wide identification, expression profile of the TIFY gene family in brassica oleracea var. capitata, and their divergent response to various pathogen infections and phytohormone treatments. Genes. 2020;11(2):19. doi:10.3390/genes11020127.
- Yang YX, Ahammed GJ, Wan CP, Liu HJ, Chen RR, Zhou Y. Comprehensive analysis of TIFY transcription factors and their expression profiles under jasmonic acid and abiotic stresses in watermelon. Int J Genomics. 2019;2019:13. doi:10.1155/2019/6813086.
- Adwy W, Laxa M, Peterhansel C. A simple mechanism for the establishment of C-2-specific gene expression in Brassicaceae. Plant J. 2015;84(6):1231–1238. doi:10.1111/tpj.13084.
- Shimoni-Shor E, Hassidim M, Yuval-Naeh N, Keren N. Disruption of Nap14, a plastid-localized non-intrinsic ABC protein in Arabidopsis thaliana results in the over-accumulation of transition metals and in aberrant chloroplast structures. Plant Cell Environ. 2010;33(6):1029–1038. doi:10.1111/j.1365-3040.2010.02124.x.
- Xu XM, Moller SG. AtNAP7 is a plastidic SufC-like ATP-binding cassette/ATPase essential for Arabidopsis embryogenesis. Proc Natl Acad Sci U S A. 2004;101(24):9143–9148. doi:10.1073/pnas.0400799101.
- Li MS, Lu J, Tao MM, Li MR, Yang H, Xia EH, Chen Q, Wan XC. Genome-Wide Identification of Seven Polyamine Oxidase Genes inCamellia sinensis(L.) and Their Expression Patterns Under Various Abiotic Stresses. Front Plant Sci. 2020b;11:12. doi:10.3389/fpls.2020.544933.
- Upadhyay RK, Fatima T, Handa AK, Mattoo AK. Polyamines and their biosynthesis/catabolism genes are differentially modulated in response to heat versus cold stress in tomato leaves (Solanum lycopersicumL.). Cells. 2020;9(8):20. doi:10.3390/cells9081749.
- Fang LJ, Fu XO, Ye QS, Wu T, Wang ZC, Chen X, Zhu ZK, Zhang XF, Fu CK. Effects of P-glycoprotein inhibitor and elicitor on the salt tolerance of rice seedlings. J Integr Plant Biol. 2007;49(5):588–597. doi:10.1111/j.1744-7909.2007.00440.x.
- Huber M, Bienvenut WV, Linster E, Stephan I, Armbruster L, Sticht C, Layer D, Lapouge K, Meinnel T, Sinning I, et al. NatB-Mediated N-terminal acetylation affects growth and biotic stress responses. Plant Physiol. 2020;182(2):792–806. doi:10.1104/pp.19.00792.
- Baek D, Pathange P, Chung JS, Jiang JF, Gao LQ, Oikawa A, Hirai MY, Saito K, Pare PW, Shi HZ. A stress-inducible sulphotransferase sulphonates salicylic acid and confers pathogen resistance in Arabidopsis. Plant Cell Environ. 2010;33(8):1383–1392. doi:10.1111/j.1365-3040.2010.02156.x.
- Nozoye T, Nagasaka S, Kobayashi T, Sato Y, Uozumi N, Nakanishi H, Nishizawa NK. The Phytosiderophore Efflux Transporter TOM2 Is Involved in Metal Transport in Rice. J Biol Chem. 2015;290(46):27688–27699. doi:10.1074/jbc.M114.635193.
- Zhou D, Quach KM, Yang C, Lee SY, Pohajdak B, Chen S. PNRC: a proline-rich nuclear receptor coregulatory protein that modulates transcriptional activation of multiple nuclear receptors including orphan receptors SF1 (steroidogenic factor 1) and ERRalpha1 (estrogen related receptor alpha-1). Molecular Endocrinology (Baltimore, Md). 2000;14(7):986–998. doi:10.1210/me.14.7.986.
- Sanjari S, Shirzadian-Khorramabad R, Shobbar ZS, Shahbazi M. Systematic analysis of NAC transcription factors’ gene family and identification of post-flowering drought stress responsive members in sorghum. Plant Cell Rep. 2019;38(3):361–376. doi:10.1007/s00299-019-02371-8.
- Li L, Ma YW, Zhang SH, Hao ZF, Li XH. Zea mays NAC transcription factor family members: their genomic characteristics and relationship with drought stress. Res J Biotechnol. 2015;10:63–77.
- Ma J, Wang LY, Dai JX, Wang Y, Lin D. The NAC-type transcription factor CaNAC46 regulates the salt and drought tolerance of transgenic Arabidopsis thaliana. BMC Plant Biol. 2021;21(1):11. doi:10.1186/s12870-020-02764-y.