
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Disruption of the Arabidopsis mitogen-activated protein kinase pathway, MEKK1–MKK1/MKK2–MPK4 (hereafter designated as MEKK1 pathway), leads to the activation of distinct NLRs (nucleotide-binding and leucine-rich repeat receptors), TNL (TIR-type NLR) SMN1, and CNL (CC-type NLR) SUMM2, resulting in dwarf and autoimmune phenotypes. Unlike mekk1 and mkk1mkk2 mutants, the dwarf and autoimmune phenotypes of mpk4 are only partially suppressed by the summ2 mutation, suggesting a significant contribution of SMN1 to the mpk4 phenotypes. However, full suppression of mpk4 by the smn1summ2 double mutation remains to be elucidated. To address this key question, we generated a mpk4smn1summ2 triple mutant and analyzed the dwarf and constitutive cell death phenotypes. The mpk4smn1summ2 triple mutant showed restoration of plant size with no detectable cell death, indicating full suppression of the dwarf and autoimmune phenotypes. These results suggest that SMN1 and SUMM2 constitute a robust surveillance system for the MEKK1 pathway against pathogen infection.
The Arabidopsis thaliana MEKK1 pathway functions downstream of the pattern-recognition receptors and has a positive role in resistance against virulent oomycetes and bacterial pathogens.Citation1,Citation2 Disruption of the MEKK1 pathway leads to dwarf and autoimmune phenotypes due to constitutive defense responses such as spontaneous cell death and accumulation of reactive oxygen species.Citation1,Citation3–6 SUMM2, a CNL protein-encoding gene involved in this phenotype, was identified via a suppressor screen of the mkk1 mkk2 background.Citation2 The summ2 mutation suppressed the dwarf and autoimmune phenotypes of mkk1 mkk2 and mekk1 mutants but did not fully suppress these phenotypes in the mpk4 mutant.Citation2 In addition, the mutation in EDS1 required for TNL function also partially does, suggesting that additional component(s) such as TNL protein are likely involved in the case of mpk4.Citation7 In our recent study, we performed novel suppressor screening to identify the components involved in dwarf and autoimmune phenotypes of the mekk1 mutants by using a dwarf autoimmune line that overexpressed the N-terminal regulatory domain of MEKK1, and we identified SMN1, also known as RPS6, encoding a TNL protein that detects the HopA1 effector.Citation8 This suggests that TNL SMN1 and CNL SUMM2 proteins monitor the integrity of the MEKK1 pathway. In the present study, we hypothesized that SMN1 is the additional TNL protein involved in the dwarf and autoimmune phenotypes of mpk4. Along this line, we focused on testing the key hypothesis concerning the suppression of mpk4 phenotypes by simultaneous mutations of smn1 and summ2. MPK4 is also the component of the ANPs–MKK6–MPK4 pathway. It is to be noted that disruption of MPK4 results in defects in cytokinesis leading to developmental dwarfism.Citation9 However, this is the case with Col-0 mpk4-2 mutant, which is reported to be a null allele.Citation10 In this study, we used Ler (Landsberg erecta) background mpk4-1 mutant, which is reported to be a weak allele with no defect in cytokinesis.Citation9 The dwarf phenotype of mpk4-1 is temperature dependent. At 28°C, morphology of mpk4-1 plant seems to be normal ( Suppl.).
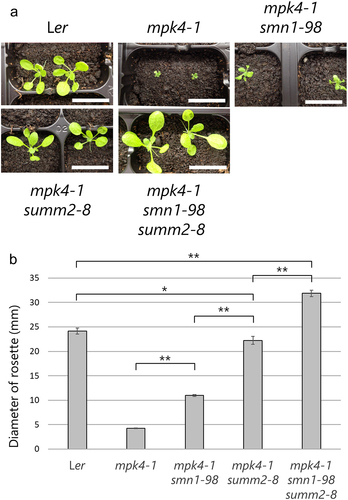
Figure 1. Shoot morphology of Ler, mpk4-1, mpk4-1smn1-98, mpk4-1summ2-8, and mpk4-1smn1-98summ2-8 mutants. Plants were germinated and grown on GM for 10 d, and then grown in soil at 24°C until 1 month of age. Bar = 5 cm. Data is representative of three biological replicates.

We generated a mpk4smn1summ2 triple mutant and compared the phenotypes of mpk4, mpk4smn1, and mpk4summ2. Experiments were performed as described by Takagi et al. (2019, 2020) Citation8,Citation11 except for that shown in . We omitted the smn1 and summ2 single mutant controls because the effects of the single mutations to mpk4 phenotypes with the controls were already analyzed in Takagi et al. (2019) and Zhang et al. (2012), respectively.Citation2,Citation8 First, we analyzed the shoot length. At 24°C, the mpk4smn1summ2 triple mutants showed almost the same length as Ler (), suggesting full suppression of the dwarf and autoimmune phenotypes of mpk4. We further observed suppression of the dwarf phenotype in the mpk4smn1 double mutant, compared to the mpk4 single mutant.Citation8 The mpk4summ2 showed more significant suppression of the mpk4 dwarf phenotype than mpk4smn1. Consistent with previous results,Citation2 mpk4summ2 was slightly shorter than Ler. We also analyzed the rosette sizes of mpk4, mpk4smn1, mpk4summ2, and mpk4smn1summ2. Unexpectedly, mpk4smn1summ2 was larger than Ler, suggesting full suppression of mpk4 dwarfism (). The larger rosette size of mpk4smn1summ2 than Ler may be ascribed to a decreased cost of resistance, which is often associated with a reduction in growth,Citation12 due to simultaneous mutations in two NLR genes: SUMM2 and SMN1. Although mpk4smn1 and mpk4summ2 were larger than mpk4, these plants were smaller than both Ler and mpk4smn1summ2. These results are consistent with the findings of our previous study and those of Zhang et al. (2012).Citation2,Citation8,Citation11
Figure 2. (a) Rosette morphology of Ler, mpk4-1, mpk4-1smn1-98, mpk4-1summ2-8, and mpk4-1smn1-98summ2-8 mutants. The plants were germinated and grown in soil at 22°C for 3 weeks. Bars = 2 cm. (b) Rosette size of the plants. Values represent averages from the following replicates (Ler n = 15; mpk4-1 n = 32; mpk4-1smn1-98 n = 4; mpk4-1summ2-8 n = 17; mpk4-1smn1-98summ2-8 n = 29), and the error bars denote standard deviation. Asterisks indicate significant differences (*p < .05, **p < .001) compared to the indicated two genotypes (in Welch’s t-test).
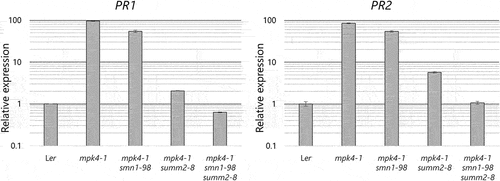
To analyze the autoimmune phenotype, we performed trypan blue staining to observe the cell death of mpk4, mpk4smn1, mpk4summ2, and mpk4smn1summ2 grown at 22°C. Consistent with the previous results, we observed a scattering pattern of stained cells in both the mesophyll and vasculature.Citation8,Citation11 The number of stained cells was significantly lower in mpk4smn1. Stained cells were negligible in mpk4summ2, mpk4smn1summ2, and Ler (). Considering the results of these phenotypic analyses, both SMN1 and SUMM2 contributed to the dwarf autoimmune phenotype of mpk4. The contribution of SUMM2 to the mpk4 phenotype was more significant than that of SMN1. To analyze the suppression of autoimmune phenotypes of mpk4 via gene expression, we also compared the expression levels of defense gene markers PR1 and PR2 with those reported in our previous studyCitation8 in mpk4, mpk4smn1, mpk4summ2, and mpk4smn1summ2 grown at 22°C in soil. Compared to mpk4, PR1 and PR2 expression decreased in a graded manner in mpk4smn1, mpk4summ2, and mpk4smn1summ2 (). The PR1 expression level was 37% lower in mpk4smn1summ2 than Ler. The PR2 expression levels were almost similar in Ler and mpk4smn1summ2. Collectively, our data suggested that mpk4smn1summ2 is comparable to Ler with respect to defense marker gene expression.
Figure 3. Trypan blue staining of rosette leaves taken from Ler, mpk4-1, mpk4-1smn1-98, mpk4-1summ2-8, and mpk4-1smn1-98summ2-8 mutants. Seeds were germinated on GM for 10 d and then plants were grown in soil at 22°C for 1 month. Detached leaves were stained. Bars = 200 m except in mpk4-1 (100
m). Data are representative of three biological replicates.
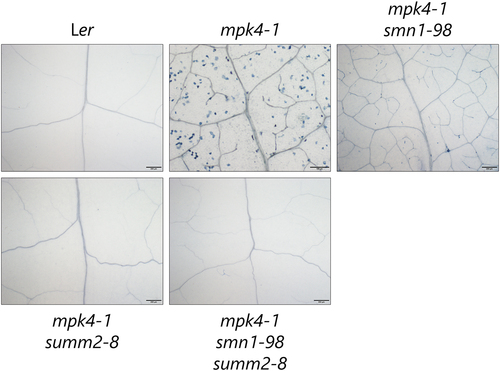
Figure 4. PR1 and PR2 expression levels in Ler, mpk4-1, mpk4-1smn1-98, mpk4-1summ2-8, and mpk4-1smn1-98summ2-8 mutants. Plants were grown in soil at 22°C for 3 weeks. Transcript levels are shown relative to Ler as determined by RT-qPCR using Actin2 as an internal standard. Error bars denote standard deviation (n = 3).
In summary, we generated a triple mutant and showed that simultaneous mutations of smn1 and summ2 fully suppress the dwarf and autoimmune phenotype of mpk4. This supports our hypothesis that the MEKK1 pathway is under the surveillance of distinct types of NLR proteins, SMN1 and SUMM2.Citation8 In addition, disruption of the ANP2/ANP3–MKK6–MPK4 pathway also results in dwarfism and an autoimmune phenotype partially dependent on PAD4 and EDS1, which are required for TNL function.Citation13 This may explain the partial suppression of the mpk4 phenotype by summ2 because MPK4 is shared downstream of both MEKK1–MKK1/MKK2 and ANP2/ANP3–MKK6 pathways. Therefore, it may be hypothesized that SMN1 is activated by the disruption of not only the MEKK1 pathway but also the ANP2/ANP3–MKK6–MPK4 pathway. Generating multiple mutants of anp2anp3 or mkk6 with smn1 would provide further clues to elucidate the intricate functional crosstalk of the distinct MAP kinase pathways.
Supplemental Material
Download TIFF Image (3 MB)Acknowledgments
We thank Susumu Mochizuki for discussions. We also acknowledge the technical expertise of the Gene Research Center of Kagawa University.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data for this article can be accessed on the publisher’s website
Additional information
Funding
References
- Ichimura K, Casais C, Peck SC, Shinozaki K, and Shirasu K. MEKK1 is required for MPK4 activation and regulates tissue-specific and temperature-dependent cell death in Arabidopsis. J Biol Chem. 2006;281:36969–4. doi:10.1074/jbc.M605319200.
- Zhang Z, Wu Y, Gao M, Zhang J, Kong Q, Liu Y, Ba H, Zhou J, Zhang Y. Disruption of PAMP-induced MAP kinase cascade by a Pseudomonas syringae effector activates plant immunity mediated by the NB-LRR protein SUMM2. Cell Host Microbe. 2012;11:253–263. doi:10.1016/j.chom.2012.01.015.
- Petersen M, Brodersen P, Naested H, Andreasson E, Lindhart U, Johansen B, Nielsen HB, Lacy M, Austin MJ, and Parker JE, et al. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell. 2000;103():1111–1120. doi:10.1016/S0092-8674(00)00213-0.
- Nakagami H, Soukupová H, Schikora A, Zárský V, Hirt H. A mitogen-activated protein kinase kinase kinase mediates reactive oxygen species homeostasis in Arabidopsis. J Biol Chem. 2006;281:38697–38704. doi:10.1074/jbc.M605293200.
- Qiu JL, Zhou L, Yun BW, Nielsen HB, Fiil BK, Petersen K, Mackinlay J, Loake GJ, Mundy J, Morris PC. Arabidopsis mitogen-activated protein kinase kinases MKK1 and MKK2 have overlapping functions in defense signaling mediated by MEKK1, MPK4, and MKS1. Plant Physiol. 2008;148:212–222. doi:10.1104/pp.108.120006.
- Suarez-Rodriguez MC, Petersen M, Mundy J. Mitogen-activated protein kinase signaling in plants. Annu Rev Plant Biol. 2010;61:621–649. doi:10.1146/annurev-arplant-042809-112252.
- Brodersen P, Petersen M, Bjørn Nielsen HB, Zhu S, Newman MA, Shokat KM, Rietz S, Parker J, Mundy J. Arabidopsis MAP kinase 4 regulates salicylic acid- and jasmonic acid/ethylene-dependent responses via EDS1 and PAD4. Plant J. 2006;47:532–546. doi:10.1111/j.1365-313X.2006.02806.x.
- Takagi M, Hamano K, Takagi H, Morimoto T, Akimitsu K, Terauchi R, Shirasu K, Ichimura K. Disruption of the MAMP-induced MEKK1-MKK1/MKK2-MPK4 pathway activates the TNL immune receptor SMN1/RPS6. Plant Cell Physiol. 2019;60:778–787. doi:10.1093/pcp/pcy243.
- Kosetsu K, Matsunaga S, Nakagami H, Colcombet J, Sasabe M, Soyano T, Takahashi Y, Hirt H, Machida Y. The MAP kinase MPK4 is required for cytokinesis in Arabidopsis thaliana. Plant Cell. 2010;22:3778–3790. doi:10.1105/tpc.110.077164.
- Xing Y, Jia W, Zhang J. AtMKK1 mediates ABA-induced CAT1 expression and H2O2 production via AtMPK6-coupled signaling in Arabidopsis. Plant J. 2008;54:440–451. doi:10.1111/j.1365-313X.2008.03433.x.
- Takagi M, Iwamoto N, Kubo Y, Morimoto T, Takagi H, Takahashi F, Nishiuchi T, Tanaka K, Taji T, Kaminaka H, et al. Arabidopsis SMN2/HEN2, encoding DEAD-box RNA helicase, governs proper expression of the resistance gene SMN1/RPS6 and is involved in dwarf, autoimmune phenotypes of mekk1 and mpk4 mutants. Plant Cell Physiol. 2020;61:1507–1516. doi:10.1093/pcp/pcaa071.
- Richard MMS, Gratias A, Meyers BC, Geffroy V. Molecular mechanisms that limit the costs of NLR-mediated resistance in plants. Mol Plant Pathol. 2018;19:2516–2523. doi:10.1111/mpp.12723.
- Lian K, Gao F, Sun T, van Wersch R, Ao K, Kong Q, Nitta Y, Wu D, Krysan PJ, Zhang Y. MKK6 functions in two parallel MAP kinase cascades in immune signaling. Plant Physiol. 2018;178:1284–1295. doi:10.1104/pp.18.00592.