
ABSTRACT
Rainfall, wind and touch, as mechanical forces, were mimicked on 6-week-old soil-grown tomato and potato under controlled conditions. Expression level changes of xyloglucan endotransglucosylase/hydrolase genes (XTHs) of tomato (Solanum lycopersicum L. cv. Micro Tom; SlXTHs) and potato (Solanum tuberosum L. cv. Desirée; StXTHs) were analyzed in response to these mechanical forces. Transcription intensity of every SlXTHs of tomato was altered in response to rainfall, while the expression intensity of 72% and 64% of SlXTHs was modified by wind and touch, respectively. Ninety-one percent of StXTHs (32 out of 35) in potato responded to the rainfall, while 49% and 66% of the StXTHs were responsive to the wind and touch treatments, respectively. As previously demonstrated, all StXTHs were responsive to ultrasound treatment, and all were sensitive to one or more of the environmental mechanical factors examined in the current study. To our best knowledge, this is the first study to demonstrate that these ubiquitous mechanical environmental cues, such as rainfall, wind and touch, influence the transcription of most XTHs examined in both species.
1. Introduction
Plants are constantly exposed to the effects and stresses of their natural environment. As being sessile organisms with limited spatial mobility, they cannot escape but have had to develop advanced strategies to cope with and adapt to them.Citation1–4
While growing and developing, plants adapt continuously to environmental changes in order to survive, better utilize the resources from nature and mitigate the environmental stresses. External environment-borne mechanical forces have been neglected for a long time, but their importance has been rediscovered in recent decades. Perceiving and responding to them, their evolutionary importance, moreover, their role in the plant growth, development, thigmomorphogenesis and stress mitigation has now begun intensively studied.Citation2,Citation3,Citation5–10 The most obvious and known mechanical forces and perturbations affecting the everyday life of plants include wind pressure, rainfall, water flow, injuries caused by animals and humans, sound, ultrasound and touch.Citation2,Citation6–8,Citation10,Citation11
After applying wind, touch, rain, wounding and dark treatments to Arabidopsis thaliana ecotype Colombia plants, five touch-inducible genes (TCH1–5) were identified by Braam and Davis.Citation12 TCH1 encodes CAM2, a calmodulin, TCH2–3 encodes calmodulin-like proteins (CML24, CML12), while TCH4 encodes XTH22, which is a xyloglucan endotransglucosylase/hydrolase. Later it was proved that TCH4 could be induced also by sound waves (50 Hz, for 30 min) in Arabidopsis thaliana.Citation13 Afterward, a series of genes sensitive to mechano-stimuli was identified including genes encoding different calmodulins, protein kinases and other proteins (summarized in Lee et al.Citation14), among them XTHs (xyloglucan endotransglucosylase/hydrolase), as well.
XTHs play role in the cell growth due to break down and even rejoin crosslinks of hemicellulose polymers in the cell wall, thereby causing loosening, extension and restructuring of the cell wall.Citation15–18 Some XTH genes were recently proved to be mechano-inducible not only in Arabidopsis thaliana Columbia (Col-0)Citation11,Citation14 but in cucumber (Cucumis sativus L. cv. Borszczagowski)Citation19 and potato (Solanum tuberosum L. cv. Desirée),Citation20 as well. In Arabidopsis thaliana, 589 genes (above 2.5% of the total genes of Arabidopsis) were proven to be inducible by touch, while 171 genes responded with down-regulation.Citation14,Citation21 Studying the 33 XTH genes of Arabidopsis thaliana (AtXTH1–33), the expression level of four AtXTHs (AtXTH17, AtXTH22, AtXTH25 and AtXTH31) increased but that of 3 AtXTHs decreased after touch stimulus,Citation14 while only one AtXTH (AtXTH22) was up-regulated in response to sound stimulus.Citation11 Two cucumber XTH genes (CsXTH1 and CsXTH3), which are up-regulated during somatic embryogenesis, were investigated for evaluating the change of their expression intensities in response to mechanical stimuli, such as touch and wounding. After applying mechanical stimuli the promoters of those XTHs were activated.Citation19 These investigations indicated that the studied CsXTHs were regulated not only developmentally but by mechanical stimuli, as well, due to the correlation between the developmental processes and mechanical stresses occur during growth, differentiation and development.Citation21 In tomato, 25 XTH genes (SlXTH1–25) were identified,Citation22 but their reply to mechano-stimulus was not examined. With the corresponding 13 sequences, 11 XTH genes (StXTH1, StXTH2, StXTH3, StXTH5, StXTH6, StXTH7, StXTH9, StXTH10, StXTH12, StXTH16 and StXTH25), which are putatively homologous to AtXTHs and/or SlXTHs, were identified. For identification of these 11 StXTHs, an XTH homology search was used based on Gene Ontology and functional annotation, and they were proven to be responsive to ultrasound in potato either by up- or down-regulation.Citation20
In the present experimental study, we have examined the XTH genes described in tomato (SlXTH)Citation22 and the XTH genes of potato (StXTH) identified earlier by our group and some of them were proven to be responsive to ultrasound.Citation20 We have studied if their expression intensities can be changed by different other mechanical stimuli, like wind, rain and touch; moreover, what is the relationship between the responses to the various mechano-stimuli and in each species.
2. Materials and methods
2.1. Plant material and growth
Two plant species, tomato (Solanum lycopersicum L. cv. Micro Tom) and potato (Solanum tuberosum L. cv. Desirée) were used in the experiments. The permission was obtained for purchase of tomato seeds and was obtained for collection of potato plants from in vitro gene bank of Centre for Agricultural Genomics and Biotechnology. Tomato seeds were each sown into pots of 10 cm in diameter. Potato plants were derived from in vitro culture; in vitro 4-week-old potato plantlets were planted individually into pots of 10 cm in diameter and acclimated for 2 weeks. The soil in pots consisted of a 1:1 mixture of perlite and forest soil.
Culture of plants was conducted under 16-h-light and 8-h-dark regime, at 22 ± 2°C daytime and 16 ± 2°C night time temperature, with 60% humidity in the culturing room. Plants were grown for 6 weeks before performing mechanical force treatments.
2.2. Mechanical force treatments and collection of samples
Rainfall, wind and touch were imitated by performing mechanical force treatments on 6-week-old tomato and potato plants.
Rainfall. A flower sprayer bottle (Cortex 2 l HY007–3) completely filled with distilled water was used to simulate the rainfall. Each treated plant was sprayed by two pressures of the spray bottle 3-times one after another, and delivering a total of 20 ml of distilled water onto the top of the plants from a distance of 15 cm.
Wind. A closed box with transparent walls was used to mimic wind pressure. A computer fan (FoxconnⓇ DC Brushless Fan, Model: PV802512L1SF 2E, DC12 V, 0.20 A) was installed into the top wall of the box. Plants were placed into this box, 10 cm from both the top wall and the computer fan, and they were blown with the airflow from the computer fan for 5 min. Each control plant was placed in a similar box but without a computer fan for 5 min.
Touch. Touch stimulation was similar to that described by Johnson et al.Citation13 Each shoot was touched lengthwise from above, with both hands, and bent 10-times to the right and left.
Each force treated and control plant was cultured under the same conditions as described above. Sampling was made immediately (at 0 min), at 15 min, 30 min and 24 h after mechanical force treatments. Three-three independent samples were collected from each treatment and their controls, in each time point from both species. Whole plants of both treated and control plants were collected at each time point and immediately frozen in liquid nitrogen. All plant samples were stored and kept at −80°C until RNA expression analysis.
2.3. Quantification of XTH gene expression by real-time PCR
Total RNA was isolated using Direct-zol™ (Zymo Research, Irvine, CA, USA) with TRIzol reagent based on the manufacturer’s manual. After total RNA extraction, three quality control methods were applied: 1) microcapillary electrophoresis with Implen n50 spectrophotometer (Implen, Munich, Germany); 2) agarose gel electrophoresis; 3) Agilent Bioanalyzer 2100 system to check the quality and quantity of total RNA. Total RNA of 120 ng was used for first-strand cDNA amplification with FIREScript RT cDNA Synthesis MIX (Solis BioDyne, Tartu, Estonia). The second-strand cDNA synthesis was performed by 5 × HOT FIREPol EvaGreen qPCR Synthesis MIX (Solis BioDyne, Tartu, Estonia) on ABI 7300 real-time PCR system (ThermoFischer Scientific, Waltham, MA, USA). Specific primers for SlXTH genes were used from Saladié et al.Citation22
The UniPro UGENE v39.0 programCitation23 was used to design specific primers for StXTHs based on the previous study about the identified homologous StXTH genes.Citation20 We selected five commonly used reference genes (EF1α, elongination factor-1alpha; actin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; sec3, exocyst complex component sec3) as normalizing genes for RT-qPCR based on the Tang et al.Citation24 results on validated reference genes in potato under abiotic stress. To compare the stability of expression intensity among the candidate reference genes, we used several statistic methods: geNorm,Citation25 NormFinder,Citation26 and BestKeeperCitation27 based on the cycle quantification value (Cq). The results were compared from the geNorm, NormFinder, and BestKeeper with the comprehensive ranking platform RefFinderCitation28 which based on the geometric mean of the rankings of every single gene calculated by each statistical program. In the RT-qPCR analysis, we used 2−∆∆Ct method to quantify the relative changes in gene expression.Citation29 Gene expression logarithmic fold change (log2FC) was calculated for comparing the intensity of SlXTH and StXTH gene expression between the control and treatment. The Student’s t test (Independent-Samples T test) was performed on ∆∆Ct values pairwise, by SPSS for Windows (SPSSⓇ, version 25.0) and p-value less than 0.05 was considered to be significant. Log2FC values were used to present the results.
The TBtools v1.09852Citation30 was used for visualizing the significantly differently expressed XTHs on the whole genome of potato and tomato based on the Solanum tuberosum SolTub 3.0 (https://plants.ensembl.org/Solanum_tuberosum/Info/Index) and Solanum lycopersicum SL3.0 (https://plants.ensembl.org/Solanum_lycopersicum/Info/Index) genome databases.
The study complies with local and national guidelines.
3. Results
Replies of 25 SlXTHs identified by Saladié et al.Citation22 to three mechanical forces were identified. Changes of expression intensities of 35 potato sequences originated from RNAseq data sets,Citation31,Citation32 which are putatively homologous to 20 AtXTHs and 21 SlXTHs, and thereby putatively StXTHs,Citation20 were studied in response to the rainfall, wind and touch. Specific primers designed for StXTHs are presented in .
Table 1. Sequences of primers designed for StXTH genes.
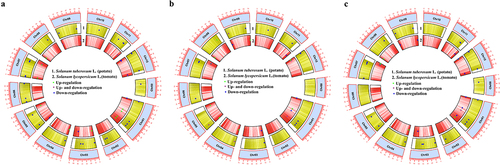
XTH genes of potato and tomato, respectively, which expression level changed at any of the time points (0 h, 15 min, 30 min or 24 h) examined after mechanical forces applied in this study are presented in . Each XTHs, which showed altered gene expression in response to rainfall (), wind (), or touch (), was plotted based on its position on the chromosome.
Figure 1. Circular view of the XTH gene density on potato and tomato chromosomes (Chr) under rainfall (a), wind (b) and touch (c) treatments based on the TBtools30.
3.1. The effect of rainfall treatment on the gene expression of XTHs
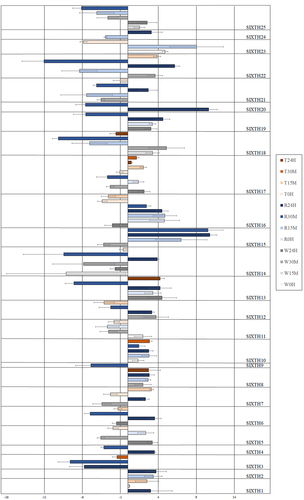
Tomato. All 25 SlXTHs responded to the rainfall treatment and significantly altered their expression levels, at least for one of the time points studied (). The expression level of SlXTH16 showed a significant increase in response to rainfall at all time points. The expression level of SlXTH5, and SlXTH11 changed only immediately after rainfall treatment (at 0 h), but in the opposite way, i.e. expression of SlXTH5 decreased, but that of SlXTH11 increased at 0 h. Expression of SlXTH1 and SlXTH7 increased significantly in response to the rainfall but only at 24 h. Similarly, an increased gene expression of SlXTH23 was detected at 15 min. Gene expression of SlXTH9 was repressed at 30 min. Other SlXTHs responded at least two time points to this mechanical cue. SlXTH2, SlXTH8, SlXTH10, SlXTH15 were induced after rainfall treatment, while SlXTH3, SlXTH18 and SlXTH25 were repressed. In the case of 8 SlXTHs (SlXTH4, SlXTH6, SlXTH12, SlXTH14, SlXTH20, SlXTH21, SlXTH22, SlXTH24) the gene expression was repressed at the first time point, while it was increased later. Just the opposite response was observed in the case of SlXTH17, i.e. it was induced at 0 h and repressed at 30 min. Gene expression was induced at 0 h and 24 h but repressed at 30 min in SlXTH13 and SlXTH19.
Figure 2. Expression induction and repression of SlXTHs of tomato in rainfall - (R), wind - (W), and touch - (T) treated plants at different time point (0 hour, 15 min, 30 min and 24 hours), assessed by real-time PCR (Average of Log2FC values from three independent samples are presented by bars. Standard deviation is represented by error bars).
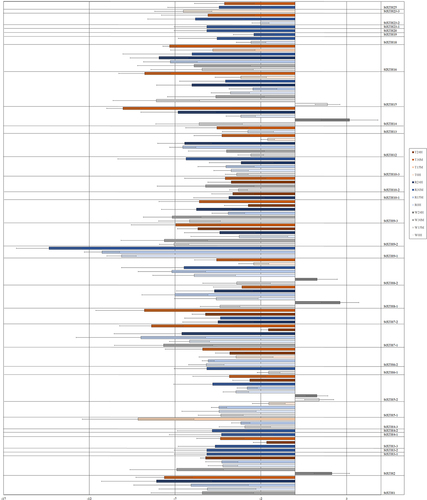
Potato. The 32 StXTHs responded to rainfall treatment by significantly changing the expression level of each (; ). The expression level of StXTH6–3, StXTH10–2 and StXTH23–3 showed no significant changes. The expression level of other StXTHs showed a significant decrease.
Figure 3. Expression induction and repression of StXTHs of potato in rainfall- (R), wind- (W), and touch- (T) treated plants at different time point (0 hour, 15 min, 30 min and 24 hours), assessed by real-time PCR (Average of Log2FC values from three independent samples are presented by bars. Standard deviation is represented by error bars).
3.2. The effect of wind treatment on the gene expression of XTHs
Tomato. Seven out of 25 SlXTHs (SlXTH1, SlXTH2, SlXTH3, SlXTH4, SlXTH9, SlXTH20, SlXTH24) were not sensitive to wind treatment (). Two SlXTHs (SLXTH10 and SLXTH23) responded to the airflow only at the first time point (0 h) by increased gene expression, while the expression level of four others (SlXTH6, SlXTH13, SlXTH16 and SlXTH19) changed only at the time point of 24 h. The expression of both SlXTH13 and SlXTH19 was induced, while that of SlXTH6 and SlXTH16 was repressed. SlXTH8, SlXTH12, SlXTH18 and SlXTH22 were induced at 30 min, as well as SlXTH18 at 15 min, while SlXTH7, SlXTH14, SlXTH15 and SlXTH21 were repressed at 30 min, as well as SlXTH14 at 0 h and 24 h. The expression level of SlXTH5, SlXTH11, SlXTH17 and SlXTH25 was either increased or decreased depending on the time point.
Potato. The 18 out of 35 StXTHs (StXTH3–1, StXTH3–2, StXTH3–3, StXTH4–1, StXTH4–2, StXTH6–2, StXTH6–3, StXTH7–2, StXTH9–1, StXTH10–1, StXTH13, StXTH18, StXTH19, StXTH20, StXTH23–1, StXTH23–2, StXTH23–3 and StXTH25) were not sensitive to wind treatment (). The expression level of StXTH5–2 was induced at time point of 0 h. The expression of StXTH8–1, StXTH8–2 and StXTH14 was decreased at 15 min but was increased at 24 h. The gene expression level of StXTH15–2 was increased at time point of 0 h and was decreased at 15 min.
3.3. The effect of touch treatment on the gene expression of XTHs
Tomato. Nine SlXTHs (SlXTH2, SlXTH4, SlXTH9, SlXTH15, SlXTH19, SlXTH20, SlXTH21, SlXTH24 and SlXTH25) were not sensitive to touch treatment (). The expression level of SlXTH17 altered significantly at each time point after touch treatment; it was repressed immediately after touch treatment but was induced later in each remaining time point. SlXTH5, SlXTH11, SlXTH14 and SlXTH23 responded immediately after touching (at 0 h); SlXTH23 was induced, while the three others were repressed. Three other XTHs (SlXTH8, SlXTH13, and SlXTH18) altered their gene expression only at the last time point (at 24 h after touching), SlXTH8 and SlXTH13 were induced, while SlXTH18 was repressed. The expression level of SlXTH1, SlXTH10 and SlXTH22 increased, while that of SlXTH3, SlXTH6, SlXTH12 and SlXTH16 decreased at one or two time points after the touch treatment. SlXTH7 was repressed at 0 h but then induced at 15 min.
Potato. The 12 out of 35 StXTHs (StXTH3–1, StXTH3–2, StXTH4–1, StXTH4–2, StXTH6–1, StXTH6–3, StXTH9–1, StXTH10–3, StXTH18, StXTH19, StXTH20 and StXTH23–1) were not sensitive to touch treatment (). The expression level of other StXTHs showed significant decrease.
4. Discussion and Conclusions
Plants are lifelong exposed to environmental mechanical factor, like rainfall, wind or touch. Plants are able to perceive and respond to these stimuli, via influencing plant growth, development thereby affecting their survival, as well as their evolutionary success.Citation2,Citation9,Citation10 Responses of plants to these cues can be short term, like Ca2+-dependent signaling or molecular changesCitation14,Citation33,Citation34 and as a consequence of those, long term, as well, like changes in growth and development.Citation8 The three environmental cues, which transcriptional effects on the XTHs of potato and tomato were investigated in the present study, are very common during the life of plants. They can be both beneficial and harmful but definitely affect the metabolism, growth and development of plants. Even if rainfall is one of the sources of essential, life-giving water, it also carries potential dangers for the plants via possible transport and spread of pathogens, or parasites.Citation12,Citation35 It should be mentioned as a limitation of the study examining the effects of rain that its effect is a combined effect due to the nature of rain. Since the plants are not only mechanically affected by raindrops falling on them but the rainwater falling on the plants and reaching the soil from there may also affect the plants. Wind also affects the structure, morphology and growth of plants and triggers functional and developmental responses.Citation12,Citation36 Touch stimuli can be originated from a wide range of environmental components, such as from animals, neighboring plants, humans or abiotic surroundings, and can similarly elicit growth-developmental responses.Citation12,Citation13,Citation37
Considering the fundamental roles of xyloglucan endotransglucosylases/hydrolases in cell growth,Citation15–18 it should not been surprising if the gene expression of at least some of them were sensitive to environmental mechanical factors. Transcriptional responses were described for some XTH genes in Arabidopsis (in response to touch and sound),Citation11,Citation14 cucumber (in response to touch or injury)Citation19 and potato (in response to ultrasound).Citation20 However, to the best of our knowledge no other studies have been reported before that have addressed either the mechano-sensitivity of tomato XTH genes (SlXTHs) or mechano-inducible/suppressive properties of potato XTH genes (StXTHs), beyond our earlier study on the ultrasound-responsive expression of 11 potato StXTH genes.Citation20
The expression of all SlXTHs was proven to be mechano-sensitive by at least one of the mechanical forces (rainfall, wind, touch) we studied. All SlXTHs changed their expression level in response to rainfall treatment, while 72% and 64% of the SlXTHs responded to the wind and touch treatment, respectively. The expression level of SlXTH2, SlXTH4, SlXTH9, SlXTH20 and SlXTH24 could be altered only by the rainfall treatment, but they were insensitive to the wind and touch.
Eleven out of these 35 putative StXTHs were proven earlierCitation20 to be responsive to ultrasound (US). The 91%, 49% and 66% of the StXTHs responded to the rain, wind and touch treatment, respectively. The expression level of StXTH1, StXTH2, StXTH5–1, StXTH7–1 and StXTH9–2 decreased at all the treatments similar to the US treatment.Citation20 The StXTH12 and StXTH16 were induced in response to the US treatment, while were repressed at rainfall, wind and touch treatments ().
Table 2. Comparative table of mechano-sensitivity of different SlXTHs and StXTHs investigated. Red indicates when the expression level increased, blue indicates when it decreased at any time points. SlXTHs are based on Saladié et al.21, StXTHs are based on Hidvégi et al.Citation20.
The SlXTH3 and SlXTH4 similarly to StXTH3–1, StXTH3–2, StXTH3–3, StXTH4–1 and StXTH4–2 were not sensitive to wind treatment. The SlXTH9 and StXTH9–1 were not sensitive to rainfall and wind treatments. Neither StXTH19 nor SlXTH19 was sensitive to wind treatment. Similarly, neither SlXTH20 nor StXTH20 was sensitive to wind and touch treatments ().
Further investigations are required to discover how these mechano-sensitive XTHs can participate in the adaptive response to the environmental cues studied, in both species. Moreover, further searching for potential and presumably XTHs is necessary that may be induced mechanically, and further investigations are needed to discover how they can participate in the growth and developmental responses to environmental cues.
Author contributions
NH and JD conceived the experiments. NH, JD and AG designed the experiments and established the plant cultures. NH, AG and BT conducted the RT-qPCR analyses, JD conducted the statistical analysis. JD, NH, AG, and BT analyzed the results, and co-wrote all versions of the paper and take responsibility for the content of the paper.
Acknowledgments
Project no. TKP2021-EGA-20 has been implemented with the support provided by the Ministry of Culture and Innovation of Hungary from the National Research, Development and Innovation Fund, financed under the TKP2021-EGA funding scheme.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Calvo P, Gagliano M, Souza GM, Trewavas A. Plants are intelligent, here’s how. Ann Bot. 2020;125(1):11–9. doi:10.1093/aob/mcz155.
- Dobránszki J. Application of naturally occurring mechanical forces in in vitro plant tissue culture and biotechnology. Plant Signal Behav. 2021;16:1902656. doi:10.1080/15592324.2021.1902656.
- Gagliano M, Mancuso S, Robert D. Towards understanding plant bioacoustics. Trends Plant Sci. 2012;17(6):323–325. doi:10.1016/j.tplants.2012.03.002.
- Karban R. Plant behaviour and communication. Ecol Lett. 2008;11(7):727–739. doi:10.1111/j.1461-0248.2008.01183.x.
- Chehab EW, Eich E, Braam J. Thigmomorphogenesis: a complex plant response to mechano-stimulation. J Exp Bot. 2009;60(1):43–56. doi:10.1093/jxb/ern315.
- Coutand C, Mitchell SJ. Editorial: Mechanical signaling in plants: From perception to consequences for growth and morphogenesis (Thigmomorphogenesis) and ecological significance. Front Plant Sci. 2016;7:1441. doi:10.3389/fpls.2016.01441.
- Gagliano M, Grimonprez M, Depczynski M, Renton M. Tuned in: plant roots use sound to locate water. Oecologia. 2017;184(1):151–160. doi:10.1007/s00442-017-3862-z.
- Hamant O, Haswell ES. Life behind the wall: sensing mechanical cues in plants. BMC Biol. 2017;15(1):1–9. doi:10.1186/s12915-017-0403-5.
- Monshausen GB, Haswell ES. A force of nature: molecular mechanisms of mechanoperception in plants. J Exp Bot. 2013;64(15):4663–4680. doi:10.1093/jxb/ert204.
- Telewski FW. A unified hypothesis of mechanoperception in plants. Am J Bot. 2006;93(10):1466–1476. doi:10.3732/ajb.93.10.1466.
- Ghosh R, Mishra RC, Choi B, Kwon YS, Bae DW, Park S-C, Jeong M-J, Bae H. Exposure to sound vibrations lead to transcriptomic, proteomic and hormonal changes in Arabidopsis. Sci Rep. 2016;6(1):1–17. doi:10.1038/srep33370.
- Braam J, Davis RW. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis. Cell. 1990;60(3):357–364. doi:10.1016/0092-8674(90)90587-5.
- Johnson KA, Sistrunk ML, Polisensky DH, Braam J. Arabidopsis thaliana responses to mechanical stimulation do not require ETR1 or EIN2. Plant Physiol. 1998;116(2):643–649. doi:10.1104/pp.116.2.643.
- Lee D, Polisensky DH, Braam J. Genome-wide identification of touch- and darkness-regulated Arabidopsis genes: a focus on calmodulin-like and XTH genes. New Phytol. 2005;165(2):429–444. doi:10.1111/j.1469-8137.2004.01238.x.
- Dyson R, Band L, Jensen O. A model of crosslink kinetics in the expanding plant cell wall: yield stress and enzyme action. J Theor Biol. 2012;307:125–136. doi:10.1016/j.jtbi.2012.04.035.
- Miedes E, Suslov D, Vandenbussche F, Kenobi K, Ivakov A, Van Der Straeten D, Lorences EP, Mellerowicz EJ, Verbelen J-P, Vissenberg K. Xyloglucan endotransglucosylase/hydrolase (XTH) overexpression affects growth and cell wall mechanics in etiolated Arabidopsis hypocotyls. J Exp Bot. 2013;64(8):2481–2497. doi:10.1093/jxb/ert107.
- Simmons TJ, Mohler KE, Holland C, Goubet F, Franková L, Houston DR, Hudson AD, Meulewaeter F, Fry SC. Hetero‐trans‐β‐glucanase, an enzyme unique to Equisetum plants, functionalizes cellulose. Plant Journal. 2015;83(5):753–769. doi:10.1111/tpj.12935.
- van Sandt Vst, Suslov D, Verbelen J-P, Vissenberg K, Van Sandt VST. Xyloglucan endotransglucosylase activity loosens a plant cell wall. Ann Bot. 2007;100(7):1467–1473. doi:10.1093/aob/mcm248.
- Malinowski R, Fry SC, Zuzga S, Wiśniewska A, Godlewski M, Noyszewski A, Barczak-Brzyżek A, Malepszy S, Filipecki M. Developmental expression of the cucumber Cs-XTH1 and Cs-XTH3 genes, encoding xyloglucan endotransglucosylase/hydrolases, can be influenced by mechanical stimuli. Acta Physiol Plant. 2018;40(7):1–11. doi:10.1007/s11738-018-2707-7.
- Hidvégi N, Gulyás A, Dobránszki J, da Silva Ja T. Mining sequences with similarity to XTH genes in the Solanum tuberosum L. transcriptome: introductory step for identifying homologous XTH genes. Plant Signaling & Behavior. 2020;15(10):1797294. doi:10.1080/15592324.2020.1797294.
- Braam J. In touch: plant responses to mechanical stimuli. New Phytol. 2005;165(2):373–389. doi:10.1111/j.1469-8137.2004.01263.x.
- Saladié M, Rose JK, Cosgrove DJ, Catalá C. Characterization of a new xyloglucan endotransglucosylase/hydrolase (XTH) from ripening tomato fruit and implications for the diverse modes of enzymic action. Plant Journal. 2006;47(2):282–295. doi:10.1111/j.1365-313X.2006.02784.x.
- Okonechnikov K, Golosova O, Fursov M, Team U. UniPro UGENE: a unified bioinformatics toolkit. Bioinformatics. 2012;28(8):1166–1167. doi:10.1093/bioinformatics/bts091.
- Tang X, Zhang N, Si H, Calderón-Urrea A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods. 2017;13(1):1–8. doi:10.1186/s13007-017-0238-7.
- Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7):1–12. doi:10.1186/gb-2002-3-7-research0034.
- Andersen CL, Jensen JL, Tf Ø. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004;64(15):5245–5250. doi:10.1158/0008-5472.CAN-04-0496.
- Pfaffl MW, Tichopad A, Prgomet C, Neuvians TP. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol Lett. 2004;26(6):509–515. doi:10.1023/B:BILE.0000019559.84305.47.
- Xie F, Xiao P, Chen D, Xu L, Zhang B. miRdeepfinder: a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol Biol. 2012;80(1):75–84. doi:10.1007/s11103-012-9885-2.
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262.
- Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, Xia R. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13(8):1194–1202. doi:10.1016/j.molp.2020.06.009.
- Dobránszki J, Hidvégi N, Gulyás A, Teixeira da Silva JA. mRNA transcription profile of potato (Solanum tuberosum L.) exposed to ultrasound during different stages of in vitro plantlet development. Plant Mol Biol. 2019;100(4–5):511–525. doi:10.1007/s11103-019-00876-0.
- Teixeira da Silva JA, Hidvégi N, Gulyás A, Tóth B, Dobránszki J. Transcriptomic response of in vitro potato (Solanum tuberosum L.) to piezoelectric ultrasound. Plant Mol Biol Report. 2020;38(3):404–418. doi:10.1007/s11105-020-01204-3.
- Monshausen GB, Bibikova TN, Weisenseel MH, Gilroy S. Ca2+ regulates reactive oxygen species production and pH during mechanosensing in Arabidopsis roots. Plant Cell. 2009;21(8):2341–2356. doi:10.1105/tpc.109.068395.
- Xu Y, Berkowitz O, Narsai R, De Clercq I, Hooi M, Bulone V, Van Breusegem F, Whelan J, Wang Y. Mitochondrial function modulates touch signalling in Arabidopsis thaliana. Plant Journal. 2019;97(4):623–645. doi:10.1111/tpj.14183.
- van Moerkercke A, Duncan O, Zander M, J Š, Broda M, Bossche RV, Lewsey MG, Lama S, Singh KB, Ljung K. A MYC2/MYC3/MYC4-dependent transcription factor network regulates water spray-responsive gene expression and jasmonate levels. Proceedings of the National Academy of Sciences; Vol. 116. 2019. p. 23345–23356. doi:10.1073/pnas.1911758116.
- Gardiner B, Berry P, Moulia B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016;245:94–118. doi:10.1016/j.plantsci.2016.01.006.
- Wang K, Yang Z, Qing D, Ren F, Liu S, Zheng Q, Liu J, Zhang W, Dai C, Wu M. Quantitative and functional posttranslational modification proteomics reveals that TREPH1 plays a role in plant touch-delayed bolting. Proceedings of the National Academy of Sciences; Vol. 115. 2018. p. E10265–E10274. doi:10.1073/pnas.1814006115.