Abstract
The introduction of the EU Water Framework Directive (WFD) is providing member state water resource managers with significant challenges in relation to meeting the deadline for ‘Good Ecological Status’ by 2015. Overall, instream physical habitat modelling approaches have advantages and disadvantages as management tools for member states in relation to the requirements of the WFD, but due to their different model structures they are distinct in their data needs, transferability, user-friendliness and presentable outputs. Water resource managers need information on what approaches will best suit their situations. This paper analyses the potential of different methods available for water managers to assess hydrological and geomorphological impacts on the habitats of stream biota, as requested by the WFD. The review considers both conventional and new advanced research-based instream physical habitat models. In parametric and non-parametric regression models, model assumptions are often not satisfied and the models are difficult to transfer to other regions. Research-based methods such as the artificial neural networks and individual-based modelling have promising potential as water management tools, but require large amounts of data and the model structure is complex. It is concluded that the use of habitat suitability indices (HSIs) and fuzzy rules in hydraulic–habitat modelling are the most ready model types to satisfy WFD demands. These models are well documented, transferable, user-friendly and have flexible data needs. They can easily be implemented in new regions using expert information or different types of local data. Furthermore, they are easily presentable to stakeholders and have the potential to be applied over large spatial scales. Integral care must be taken in the use of appropriate HSIs as these are the most sensitive part of the modelling and inaccurate results will be gained if not correctly formulated. If representative HSIs are not available, fuzzy rule-based modelling is recommended, but care must also be taken in the designing of the rule sets. For larger-scale modelling or when only few field data are available, generalized habitat models hold promise for quantifying habitat suitability based on average stream characteristics.
1 Introduction
Until the introduction of the EU Water Framework Directive (WFD), EU member states have had varying levels of water management legislation and assessment approaches for assessing the health of the biota–physical relationship of their streams (Dunbar et al. Citation2001, Tharme Citation2003, Harby et al. Citation2004). The WFD requires an integrated approach to manage three of the key components of stream ecosystems: water quantity, water quality and physical habitat that support the biology of the system (Commission Citation2000), with the assumption that there are links between stream biota and the three components of stream habitat (Poff et al. Citation1997, Humphries et al. Citation1999, Maddock Citation1999, Sullivan et al. Citation2004, Citation2006). The WFD implicitly assumes an underlying link between ecological status and abiotic quality elements; thus, a key aspect is to identify and assess the links between the physical and biological components of streams if a stream is found to be non-compliant biologically (Commission Citation2000). For example, some major variables to affect fish and invertebrate species carrying capacities in streams include physical habitat variables such as flow and velocity (Beecher et al. 1995, Citation2002, Poff et al. Citation1997, Humphries et al. Citation1999, Holm et al. Citation2001, Jowett et al. Citation2005, Suren and Jowett Citation2006, Schweizer et al. Citation2007), water depth (Egglishaw and Shackley Citation1982, Kennedy and Strange Citation1982, Beecher et al. 1995, Bardonnet et al. Citation2006, Schweizer et al. Citation2007), substrate composition (Heggenes Citation1988, Cobb et al. Citation1992, Knapp and Preisler Citation1999, Kondolf Citation2000, Vadas and Orth Citation2001) and cover (Heggenes and Traaen Citation1988, Vehanen et al. Citation2000, Vadas and Orth Citation2001). It is the knowledge of the relationships between such physical habitat variables and biota that is needed for many member states for both research purposes and implementation of the WFD (Commission Citation2000, Logan and Furse Citation2002).
Instream physical habitat modelling approaches could help water managers in the decision-making process in relation to biota–physical habitat links. They are modelling tools for predicting the suitability of a habitat to support a target species by quantifying the relationship between stream biota and physical habitat (Hardy Citation1998). Instream physical habitat modelling can be used to (i) identify the individual biota–physical habitat relationships, (ii) assess the quality of the physical habitat variables through its impact on the biota and (iii) predict likely biological responses if hydromorphological changes to a system are to occur, and make decisions on this information. They are currently used in both stream management and research worldwide (Tharme 1996, Citation2003, Jowett Citation1997, Dunbar et al. Citation1998, Parasiewicz and Dunbar Citation2001, Leclerc et al. Citation2003). However, instream physical habitat modelling types within the broader habitat modelling approaches differ in their applicability to biota–physical habitat assessments for EU water managers (Rosenfeld Citation2003, Harby et al. Citation2004, Jorgensen Citation2008). For models to be considered by water managers to be used now, they will need to specifically:
-
supply a clear understanding of the links between the biological and physical habitat elements using available data (e.g. monitoring data);
-
be user-friendly (i.e. managers use them without intensive training support);
-
be transparent (i.e. easily presented to different stakeholders); and
-
be widely applicable (i.e. can be applied on a large spatial scale and for different biota elements) (Pollard and Huxham Citation1998, Logan and Furse Citation2002).
It is not the intention of this paper to provide a detailed description of all the instream physical habitat modelling types (appropriate references are provided), but to provide an overview and analysis of the suitability and readiness of each habitat model type in relation to the above-mentioned criteria.
2 Instream physical habitat modelling types
2.1 Habitat suitability models
Habitat suitability index (HSI) modelling uses physical habitat variables to assess the physical quality of an area to support a target indicator species (Bovee Citation1986, Raleigh et al. Citation1986, Hardy Citation1998). HSIs are also referred to as ‘habitat suitability curves’. They are the biological basis of HSI modelling and represent the functional relationship between the selected physical habitat variable and the response of the selected species/lifestage (Bovee and Cochnauer Citation1977, Bovee et al. Citation1998). The suitability index varies between 0 (unsuitable) and 1 (most suitable) and provides a probability measure on how suitable a habitat is for a target species (Bovee and Cochnauer Citation1977, Bovee et al. Citation1998). They are commonly used for assessing site-specific suitability, but are also effective at a wider spatial range under a set of generalized criteria and can be used to predict outcomes if the input variables are manipulated (Raleigh et al. Citation1986).
HSIs can be derived from univariate suitability functions that consider individual habitat variables independently from one another () (Bovee Citation1986) or from multivariate functions (Ahmadi-Nedushan et al. Citation2006) that consider the interaction of several habitat variables to determine the species response for the cumulative effects of several habitat variables () (Vismara et al. Citation2001).
Figure 1 Examples showing (a) univariate HSI curve for the habitat variable ‘depth’ and (b) a multivariate HSI showing the species response to the cumulative effects of both habitat variables depth and velocity
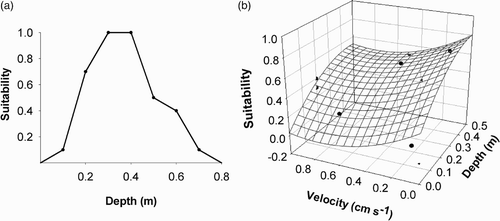
Univariate HSIs are the simplest and most commonly used and are derived in three ways depending on the available information (Bovee Citation1986, Bovee et al. Citation1998, Parasiewicz and Dunbar Citation2001, Ahmadi-Nedushan et al. Citation2006): Category I – expert opinion indices are derived from both professional judgement (expert opinion) and life history literature; Category II – habitat use indices are based on the frequency of occurrence from in situ studies of habit use of the target species and Category III – preference indices are Category II adjusted for in situ habitat availability measured at the same time as habitat use sampling (Bovee Citation1986). Categories II and III are the most ecological defensible due to their use of local in situ environmental data, but Category I is considered useful in data-poor situations and is also useful to generate ‘regional’ curves, applicable to all rivers of a certain hydrological and morphological ‘type’ within the same region. Category I HSIs can also be upgraded as more information becomes available. Category III HSIs can be derived using a number of ‘index of selection’ techniques; Ivlev index (Ivlev Citation1961), Jacobs selectivity index (Jacobs Citation1974) and forage ratio (Edmondson and Winberg Citation1971), the simplest and most common being the forage ratio. The equation for the forage ratio is as follows:
HSIs can be defined to take into account diel, seasonal and size class/age variation in habitat use or preference, all of which must be considered if suitable HSIs are to be used (e.g. a set of HSIs developed for a certain size class of fish may not be applicable to another, or a set of HSIs developed in summer will most likely not be applicable in winter). HSIs of individual habitat variables such as velocity, depth and substrate can be combined to give composite suitability indices (CSIs) expressing an overall assessment of the physical habitat quality of a defined river reach (Bovee Citation1986, Vadas and Orth Citation2001, Beecher et al. Citation2002). CSIs can be calculated in different ways with the ‘product equation’ being the most common. The equation assumes that habitat variables are independent of each other and of equal importance. The equation is
If one of the HSIs is zero, then the CSI will yield zero no matter the values of the other HSIs. This equation can be modified to include weighting factors that consider the relative importance of each variable to the target biota (Ahmadi-Nedushan et al. Citation2006).
Alternative methods for calculating CSI include the ‘arithmetic mean’, where high-quality habitat variables can compensate for poor quality habitat variables, as it takes an average of the HSI values. The ‘geometric mean’ is similar to the arithmetic mean, but instead of taking a direct average of the sum of the HSI values, the values are multiplied together and the nth root of the product taken, so some compensation is present, but similar to the product equation yields zero if one of the variables gets zero habitat suitability. The method ‘lowest HSI’ assumes that the lowest quality habitat variable is the most limiting, so determines the upper limit of the CSI, and high-quality HSIs cannot compensate for poorer ones (Korman et al. Citation1994).
2.2 Fuzzy rule-based modelling
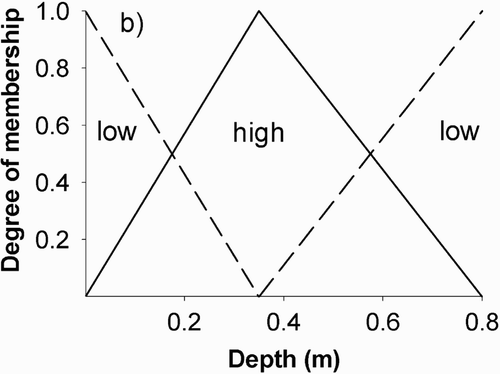
Fuzzy rule-based modelling is being proposed as an alternative to the HSI modelling types for stream biota–habitat analyses (Silvert Citation2000, Ahmadi-Nedushan et al. Citation2008). Fuzzy rule-based modelling does not use conventional input data sets known as ‘crisp sets’ (e.g. 0.1–0.2 m for depth), as in conventional HSI modelling (), but uses IF–THEN rules that are more flexible with biota suitability being able to partially belong to different sets () (Adriaenssens et al. Citation2004, 2008). Fuzzy rule-based suitability data are presented in the form of verbal type commands similar to the way a human brain thinks (e.g. low, medium, high), and the CSI attained is the result of how the sets of fuzzy rules are defined (Ahmadi-Nedushan et al. Citation2006). All possible combinations have to be considered and the output from the model is also fuzzy until a transformation process called ‘defuzzification’ (Ahmadi-Nedushan et al. Citation2008) is used and the ‘crisp output’ data presented as CSIs (Adriaenssens et al. Citation2004, Harby et al. Citation2004, Silvert Citation1997).
Figure 2 An example of a fuzzy set describing the relative degree of habitat suitability membership (i.e. low or high) for the habitat variable depth
Fuzzy rule-based approaches have been used for fish (Jorde et al. Citation2001, Kerle et al. Citation2002, Mouton et al. Citation2008) and benthic invertebrate studies (Adriaenssens et al. Citation2006, Van Broekhoven et al. Citation2006, Citation2007). They are also being hybridized with other types of approaches such as artificial intelligence-based models to increase their coverage beyond HSI modelling types (Adriaenssens et al. Citation2004, Ahmadi-Nedushan et al. Citation2006).
2.3 Linear models
Linear regression can be used to investigate biota–physical habitat relationships (Gordon et al. Citation2004, Harby et al. Citation2004). Linear regression approaches follow four main assumptions: (i) the observations are independent, (ii) the residuals are assumed to be identically and individually distributed, (iii) model residuals are assumed to follow a normal distribution and (iv) the regression function is a linear function of its parameters.
2.3.1. Simple linear regression
Simple linear regression (SLR) assesses habitat variables independently using a univariate function (Quinn and Keough Citation2006). The equation is
2.3.2. Multiple linear regression
Multiple linear regression (MLR) approaches consider the joint effect of multiple explanatory variables and their interactions. The equation is
2.3.3 Generalized linear models
Generalized linear models (GLMs) allow the dependent variable to have any distribution from the exponential family (e.g. binomial, Poisson, etc.); however, a transformation is used to achieve a linear dependency of the explanatory variables. This makes GLMs more flexible for analysing biota–physical habitat relationships based on input data that are not normally distributed (Agresti Citation1996). The equation is
2.3.4. Logistic regression
Logistic regression (LR) is appropriate for binary dependent variables such as the presence/absence (Jongman et al. Citation1995). Even though this methodology requires more data, an advantage is that binary data are common in instream studies (Filipe et al. Citation2002). LR is used similar to MLR (Guay et al. Citation2000); however, LR is based on fitting data to the logistic curve:
LR has been widely applied in the field of habitat modelling for fish (Geist et al. Citation2000, Guay et al. Citation2000, Garland et al. Citation2002) and benthic invertebrates (Udevitz et al. Citation1987, Peeters and Gardeniers 1998, Castella et al. Citation2001), and also macrophytes to a lesser extent (Narumalani et al. Citation1997, Wagner and Falter Citation2002, Van den Berg et al. Citation2003). It was successfully used to predict habitat selection by Apache trout in six east-central Arizona streams (Cantrall et al. Citation2005), also being instigated successfully to study fish–habitat relationships in a Californian river (Harvey et al. Citation2002), and used it to identify the most important environmental variables affecting freshwater fish in a semi-arid stream in Portugal (Filipe et al. Citation2002).
With all parametric-based analyses types, there are inherent problems when dealing with biota–physical habitat relationships as many of the assumptions of the modelling are not realized (i.e. data are not normally distributed data or there is no linear dependency), and transformations are also inherent with problems (i.e. more difficult to interpret results) (Townend Citation2002, Quinn and Keough Citation2006).
2.4 Non-parametric regression models
Non-parametric models are available and are starting to be used as an alternative to parametric-based types. If the assumptions of parametric regression approaches do not hold, then non-parametric techniques may be more appropriate (Quinn and Keough Citation2006, Jowett and Davey Citation2007).
Although these techniques have been in use for over 30 years, it is only in the last 10 years that they have been used in instream physical habitat modelling approaches. One technique that is gaining popularity is the general additive model (Jowett and Davey Citation2007) and to a lesser extent the multivariate adaptive regression splines (MARS) (Leathwick et al. Citation2005, Citation2006, Elith and Leathwick Citation2007).
The generalized additive models (GAMs) are a non-parametric extension of GLMs (Guisan et al. Citation2002). They use smooth functions of the explanatory variables, allowing the form of a relationship between the independent variable and the dependent variable to take on any shape as defined by the data (Jowett and Davey Citation2007). GAMs provide more flexibility in biota–physical habitat analyses because of their ability to deal with curvilinear relationships and non-monotonic relationships between independent and dependent variables. The only assumptions of GAMs are that functions are additive and the components are smooth (Guisan and Zimmermann Citation2000).
The MARS models are similar to GAMs as they make no assumption about the underlying functional relationship between the dependent and the independent variables, and they allow the independent variables to have any type of distribution (Friedman Citation1991, Hastie et al. Citation2001, Leathwick et al. Citation2005). MARS partitions the input space into regions, each with its own regression equation, which makes it suitable to solve problems involving more than two variables. MARS has been used to accurately describe the relationship between fish distribution and explanatory variables (Leathwick et al. Citation2006).
2.5 Artificial neural networks
Artificial neural networks (ANNs) evolved during the 1990s as a ‘black box’ nonlinear modelling type which is used to model complex nonlinear relationships between dependent and independent data, similar to the way a human brain works (Lek et al. Citation1996a, Citation1996b, Lek and Guegan Citation1999, May et al. Citation2008). Because biota–habitat relationships are often complex and non-linear, leading to problems with parametric regression-based approaches (Lek and Guegan Citation1999), ANNs have been proposed to overcome some of these problems (Lek et al. Citation1996b, Brosse et al. Citation1999a, Citation1999b, 2001). ANNs are able to learn and then identify relationships between input and output data, even if the data are imprecise and noisy. Owing to their ability to model (i) non-linear, (ii) complex multivariate data, (iii) noisy data and (iv) learning and adaptation ability, they are increasingly being used in aquatic habitat modelling (Dedecker et al. Citation2005, Dakou et al. Citation2006, Park et al. Citation2006). There are no specific raw physical habitat data requirements for ANNs, and similar to HSI and fuzzy rule-based modelling, any quantitative variable can be used as an input or output variable.
Two types of ANNs are being applied in habitat modelling approaches (Harby et al. Citation2004), that is, backpropagation networks (Lek and Guegan Citation1999) and Kohonen self-organizing maps (Lasne et al. Citation2007), with backpropagation networks being most widely used for fish (Lek et al. Citation1996a, Mastrorillo et al. Citation1997, Brosse et al. Citation1999b) and benthic invertebrates (Dedecker et al. Citation2001, Citation2005, Lencioni et al. Citation2006). Kohonen self-organizing maps have been used for benthic invertebrates (Lencioni et al. Citation2006).
2.6 Individual-based or agent-based modelling approaches
Individual-based models (IBMs) have been developed as an advanced alternative to conventional habitat suitability modelling (Bonabeau Citation2002, Grimm and Railsback Citation2005). They work at the individual level in the population and model biological responses (e.g. growth, fecundity, mortality, movement) of that individual to physical habitat variables (Stillman Citation2008). It is assumed that the target biota selects habitat to maximize their potential fitness (Railsback and Harvey Citation2002), and individuals (also called agents) will execute certain behaviour in relation to their interactions with their surroundings. The models are built by using IF–THEN rules to elucidate behavioural responses to changes in the habitat variables (Railsback and Harvey Citation2002). IBMs have been used in biota–physical habitat studies mostly for fish (Van Winkle et al. Citation1998, Railsback and Harvey Citation2002, Grand et al. Citation2006, Harvey and Railsback Citation2007).
Two emerging IBM sub-types are biological process-based models and bioenergetic models (Harby et al. Citation2004). Biological process-based models are an extension of conventional instream physical habitat modelling assessing the habitat suitability for a species by using factors such as food availability, movement ability, foraging behaviour and metabolic processes (Hayes et al. Citation2000). A simple biological process-based model could be fish growth under different physical habitat conditions, whereas a more complex model might include other variables such as feeding, reproduction or digestion rates (Hayes and Jowett Citation1994).
Biological process-based models have been mostly developed for drift-feeding fish (Hughes and Dill Citation1990, Hughes Citation1992, Citation1998, Hayes et al. Citation2000, Hughes et al. 2003, Booker et al. Citation2004). Bioenergetic models are a special type of biological process-based model that defines the mass balance relationship between the amount of food consumed by a fish and its growth, and optimal fish location is based on energy budgets (Brant and Hartmann Citation1993, Hansen et al. Citation1993, Hill and Grossman Citation1993, Ney Citation1993). Bioenergetic models require inputs of bioenergetic parameters, water temperature data, growth data, energy density data and diet data (Brant and Hartmann Citation1993). Bioenergetics modelling has been used in fish habitat modelling to evaluate habitat quality (Booker et al. Citation2004), predator–prey dynamics (Stewart and Ibarra Citation1991), trophic interactions (Pothoven et al. Citation2001) and feeding patterns of fish on macroinvertebrates (Rand et al. Citation1995).
3 Hydraulic–habitat modelling
3.1 Conventional hydraulic–habitat models
Instream physical habitat models are often combined with a hydraulic model to form a ‘hydraulic–habitat’ model (Bovee and Milhous Citation1978, Stalnaker et al. Citation1995). The hydraulic–habitat model is very relevant as a water management tool under WFD conditions because it represents flow as the driver of the ecology within the river system (Poff et al. Citation1997, Bunn and Arthington Citation2002, Naiman et al. Citation2002), and this in turn affects the habitat quality available to the biota. Hydraulic–habitat models are mostly used when anthropogenic changes in flow regime (e.g. hydropower, water abstraction) are suspected to affect biota (Gibbins and Acornley Citation2000, Thorn and Conallin Citation2006, Olsen et al. Citation2009). Typically, hydraulic–habitat models are using HSI or fuzzy rule-based model types to represent the physical habitat requirements of species. Instead of giving a single HSI or CSI, the output of hydraulic–habitat models is a functional representation of the habitat availability quality for indicator biota under different flows (Bovee et al. Citation1998). For any surveyed river reach, the modelled habitat availability is expressed as a weighted usable habitat area (WUA) in units of square metre habitat per metre river length. Typically, as discharge increases from zero, WUA rises until an optimum flow, and then it decreases as discharge becomes too high to sustain optimal habitat conditions (Stalnaker et al. Citation1995). This functional representation can be converted into a hydraulic habitat suitability (HHS) index ranging between 0 and 1, similar to a HSI or CSI (Bovee et al. Citation1998).
Combined hydraulic–habitat models are probably the most widely applied instream physical habitat modelling type in the world in general and Europe in particular (Tharme Citation2003), and will be especially important for EU water managers where flow regime is being modified. There are already many well-established hydraulic–habitat models that have been tested on many stream types in Europe (Harby et al. Citation2004). The most famous and widely used approach is the instream flow incremental methodology and in particular one of its major components, the physical habitat simulation system (Bovee et al. Citation1998, Hardy Citation1998). This basic modelling system has been widely adapted and modified at the international level such as RHYHABSIM (Jowett Citation1992, Citation2006), CASIMIR (Schneider and Peter Citation1999, Jorde et al. Citation2001, Mouton et al. Citation2008), EVHA (Dunbar et al. Citation1998) and RHABSIM (Payne and Associates Citation2000). These models have been employed worldwide, and although they have been criticized for things such as transferability of HSIs and lack of ecological predictive capability (Orth and Maughen Citation1982, Mathur et al. Citation1985), they are also considered to be ‘the most scientifically and legally defensible methodology available’ for instream physical habitat assessments in many countries (Gore and Nestler Citation1988, Dunbar et al. Citation1998, Tharme Citation2003).
Two of the main advantages of hydraulic–habitat modelling are that multiple hydromorphological drivers (e.g. water use, morphological degradation) can be handled simultaneously, and physical habitat changes can be assessed at larger spatial scales (Cavendish and Duncan Citation1986).
3.2. Generalized habitat models
Generalized habitat models (also called statistical habitat models) are a modelling type being proposed as an alternative to conventional hydraulic–habitat modelling approaches (Lamouroux and Jowett Citation2005). Considerable field effort and experience is needed to run the conventional hydraulic–habitat models, whereas generalized habitat models obtain reach scale habitat values (HVs) based on a limited number of field measurements (Lamouroux et al. Citation1998), HVs range between 0 and 1 and are defined as the WUA divided by the wetted area of the reach. The generalized habitat models use reach hydraulic geometrical relationships (i.e. mean depth–discharge and mean width–discharge relationships) and simple average reach descriptions (i.e. mean particle size, Reynolds number) to estimate HV indices from discharge simulations (Lamouroux and Capra Citation2002, Stewardson and McMahon Citation2002, Stewardson Citation2005). It was shown that a limited reach scale data set collected at two different discharge levels was sufficient for a generalized habitat model to predict reach scale WUAs which were comparable with the ones predicted using the more data-demanding conventional hydraulic–habitat model types (Lamouroux and Capra Citation2002, Lamouroux and Jowett Citation2005). This makes the generalized habitat models particularly valuable for large-scale assessments or when only few reach data are available.
4 General discussion
Water managers are constantly in the process of adapting their existing national and regional assessment methods, modifying methods used in other countries or developing new methods to help them to meet the challenges of water resource management (Rosenfeld Citation2003), and for EU member states, it is essential for the implementation of the WFD (Wallin et al. Citation2003).
When deciding on modelling approaches for biota–physical habitat relationships, EU water managers must compromise between the accuracy of the approach and the policy, budget, timescale, expertise required, and ecological and hydromorphological data requirements of the approach (Jorgensen and Bendoricchio Citation2001). A trade-off between ‘optimizing accuracy’ versus ‘optimizing generality’ must be made in relation to management-based models (Guisan and Zimmermann Citation2000). Owing to a lack of environmental data, limited resources and the need to act immediately, certain modelling approaches are excluded or unsuitable as management-based models (Rosenfeld Citation2003).
Owing to the relatively short timeframe of the WFD, modelling types that require large amounts of data still to be collected will be substituted for model types that can use the already available data (e.g. monitoring data), even if this means decreasing the accuracy of the approach, and complex models substituted for simpler types (Rosenfeld Citation2003). provides an overview of the various instream habitat modelling approaches in relation to the four criteria set out in Section 1. A scoring system has been used to assess the overall utility of the models for water managers (), and a more detailed discussion is given for each approach below.
Table 1 Summary of different instream physical habitat modelling types in relation to their applicability as modelling tools for EU water managers
4.1 HSI models
HSI-based approaches have been in use for over 30 years and their popularity continues to grow probably due to their recognition worldwide as legitimate approaches when dealing with biota–hydromorphological analyses (Jowett Citation1997, Leclerc et al. Citation2003, Gordon et al. Citation2004, Harby et al. Citation2004). HSI approaches also deal with many types of data (e.g. continuous, discrete, binary, categorical) simultaneously and have been developed and tested for different biota groups (albeit mostly fish and benthic invertebrates). They can incorporate coarse monitoring data and expert opinion, and there are readily available user-friendly computer programs, many of which are used in combination with a hydraulic model (Bovee et al. Citation1998). They are also applicable over a wide spatial scale, but it is essential that care must be taken to make sure that the individual HSIs used in the modelling accurately represent the biota–physical habitat relationships of the area they are to be used in. Incorrect use of HSIs is a significant source of error in this modelling, mostly due to HSIs being transferred from one area to another area without adequate testing to determine transferability (Thomas and Bovee Citation1993, Moir et al. Citation2005).
Ideally, when using HSIs, they should be developed on a site-by-site basis, but this is impractical. Instead, Category III HSIs should be used as these habitat preference curves have been found to be more transferable than HSIs which are based on observed habitat use (Beecher et al. Citation1993, Dunbar et al. Citation2001). However, adequate evaluation of transferability should be conducted (Thomas and Bovee Citation1993, Groshens and Orth Citation1994), even if only in a literature review sense. If confidence is taken in the HSIs, then the conventional HSI methods are well suited for water managers, and the HSI approach is a clear candidate as a WFD management-based modelling approach.
4.2 Fuzzy rule-based modelling
Fuzzy rule-based approaches have advantages similar to HSI approaches, but can also better handle imprecise and qualitative data which are common with ecological data. The better use of imprecise and uncertain data including expert opinion makes the fuzzy rule-based approaches more appealing than HSI modelling, especially in data-poor situations (Adriaenssens et al. Citation2004, Mouton et al. Citation2008). Furthermore, they consider multivariate interactions of input variables, new parameters can easily be added and the system is relatively easy to implement (Ahmadi-Nedushan et al. Citation2008), all of which are attributes attractive to water managers.
However, fuzzy rule-based models are still in their exploration stage and it may be difficult to convince water resource managers of the benefits of such subjective methods when they are looking for defendable data-driven-type approaches (Adriaenssens et al. Citation2004). Another problem is the determination and tuning of the membership functions and fuzzy rules. This is not a straightforward procedure and relies entirely on the opinion of experts (Chen and Mynatt Citation2003). Expert opinion is obviously a major concern, both in the choice of input variables and in the formulation of the rules, but for some biota groups (e.g. macrophytes, diatoms) expert opinion will be all water managers have, no matter what method is used. To overcome these problems (Cornelissen et al. Citation2001), (i) stressed criteria are needed to determine necessary qualifications of experts, (ii) proper elicitation of expert knowledge is necessary to construct membership functions and (iii) methods are needed to test the validity of a membership function.
Another solution that is being investigated to improve the flexibility of the system is the hybrid fuzzy approach, whereby fuzzy models are combined with techniques such as principal component analysis, genetic algorithms or ANNs (Meesters et al. 1998, Loia et al. Citation2000, Mackinson Citation2000), but at the same time it increases the complexity of the modelling process, something water managers want to avoid (Adriaenssens et al. Citation2004).
4.3. Parametric regression techniques
Linear regression models are robust if their assumptions are justified. Unfortunately, in stream ecology, these assumptions are rare, and even transformations of the data to help meet these assumptions are inherent with certain problems (Quinn and Keough Citation2006). However, linear regression techniques are well established, popular, available in most commercial computer packages and can deal with most types of data (Townend Citation2002).
SLR has been criticized because biota–hydromorphological relationships in streams are known to be a multivariate process (Jowett Citation1997, Geist et al. Citation2000, Guay et al. Citation2000, Vadas and Orth Citation2001, Vismara et al. Citation2001, Neumann and Wildman Citation2002, Leclerc et al. Citation2003) and treating variables as independent does not always give an adequate representation of the true situation (Nykanen et al. Citation2001). However, SLR is often used as a ‘first indication’ of which variables are important and then more appropriate methods are applied (Quinn and Keough Citation2006). MLR deals with more than one independent variable but a potential problem with MLR models is multicollinearity (Affifi and Clark Citation1996). Many variables are known to be highly correlated with each other (depth, velocity, substrate, etc.), and the final set of variables in the model may be a result of strong associations with other variables and can lead to redundant variables being included or important variables being excluded from the modelling (Ahmadi-Nedushan et al. Citation2006). Another drawback is that most parametric models are built up using local habitat data so they are site-specific, thereby leading to problems in transferring models between sites or comparing sites from different models, or extrapolating between sites, which are common requirements for water managers. Consequently, these types of techniques are not widely applicable and best suited in the explanatory phase for determining the importance of habitat variables to use in a later modelling phase.
4.4 Non-parametric techniques
GAMs have only been used within the last 5–10 years in instream physical habitat modelling. Jowett and Davey Citation(2007) compared GAMs against traditional habitat suitability criteria for both fish and invertebrates. They found that the GAM performed slightly better, but it was more complicated and the user had to be cautious not to over-fit the data in which case the model may cause minor fluctuations in the data to be exaggerated. The main issue with GAMs is that they need a lot of data to make their use worthwhile. If these data are available, then there may be situations where they give better results in identifying sharp thresholds in habitat selection (Jowett and Davey Citation2007). MARS works with data sets similar to GAMs but according to Leathwick et al. Citation(2006) they hold some advantages. They found that MARS results were easier to interpret ecologically. Furthermore, it is computationally much faster and more easily incorporated into other analyses using, for example, GIS modelling (Munoz and Fellicísmo Citation2004) than approaches such as GAMs and ANNs. However, both techniques require large data sets that may not be readily available.
As analysis types such as GAMs and MARS are used increasingly in research to supplement parametric-based models, they will become better documented and probably more acceptable as management-based analysis tools. But for the present, water managers should probably focus on more established parametric model types and only use more advanced techniques such as GAMs and MARS if transformations of the input data are not possible to fulfil the requirements of parametric models. However, this does not mean that they should be excluded if water managers have knowledge of them, as they hold advantages over those of parametric regression models.
4.5 Artificial neural networks
For water resource managers, ANNs have advantages in being able to process complex, nonlinear and noisy data, which is often the case with ecological data. Any type of raw data (i.e. discrete, continuous and dichotomous) can be used in ANNs (Lek and Guegan Citation1999).
Unfortunately, there are also some major limitations for water resource managers when it comes to using ANNs. First, ANNs often require large sets of training data. This is a problem as background ecological data are often limited, leading to too few data being used to train the model, and considerable time and resources can be spent gaining this information (Lek and Guegan Citation1999). Second, there are very few ANN ‘package’ models available. Because each study usually has varying amounts of input data, different amounts of input neurons, hidden layers, etc., are needed. This means that ANNs are usually designed specifically for each study being programmed using network toolboxes or by building new computer codes. For this purpose, the user needs to be experienced in computer programming and model building, and the model is therefore not easily transferable (Harby et al. Citation2004). Furthermore, there is also no standard method for choosing the optimal ANN characteristics, such as the number of hidden neurons and transfer functions. This leads to difficulties in following the pathways and defining the most important variables (Olden and Jackson Citation2002, Harby et al. Citation2004).
ANNs have also been criticized for their ‘black box’ approach, in the sense that ANNs do not provide a clear insight or understanding of the pathways from the input variables to the predicted output variables. This makes it very difficult to present the model to stakeholders. Therefore, ANNs are probably still better suited to the applied research field, but they may also be useful in site-specific situations where enough data are available. However, at this stage, they are not well suited for many member states as water management models, where the need to apply output at large spatial scales is required.
4.6 Individual-based approaches
IBMs provide advantages in which they require little in situ historical background data (e.g. field data) as they work off biological behavioural response–physical habitat relationships (Railsback et al. Citation2003, Harvey and Railsback Citation2007). Once these are established, they should be universal (Grimm and Railsback Citation2005). IBMs are also advantageous in that they are able to model complex nonlinear relationships where space is crucial (bottlenecks), and the positions of the individuals are not fixed (Railsback and Harvey Citation2002). The individuals exhibit complex behaviour which includes learning and adaption behaviour (Bonabeau Citation2002). This makes IBMs ideal for biota such as fish and to a lesser degree macroinvertebrates (Grimm and Railsback Citation2005). The major limitation is the determination of the critical biological responses to physical habitat (especially within bottlenecks) which are poorly known for most biota. For this purpose, expert opinion must be relied on to make the rules, thereby increasing subjectivity. Another problem is that IBM computer packages are not widely available and therefore not easily accessible.
From an EU water manager's view, IBMs are still not developed enough to be seriously considered as a WFD management model, as there are insufficient data to be applied at a large scale, and the hydraulic modelling is complex (usually 3D). Currently, the modelling is limited to drift feeding fish such as salmonids. Therefore, at this point in time, it is better suited for academic or research modelling approaches. However, it has the potential to be coupled with conventional physical habitat suitability modelling approaches to better understand and predict biota–physical habitat relationships or to study specific problems identified using coarser habitat model types (Grimm and Railsback Citation2005).
4.7 Hydraulic–habitat modelling
In order for the instream physical habitat models to be applicable at the larger scales often considered in water management, a combined hydraulic–habitat model should be used. This requires that the habitat model which is built using microscale data is applicable at river or reach scale. For this purpose, the fuzzy logic and HSI-based habitat models are most useful because of their flexible data use and their functional representation which means that the transferability can be accomplished using expert opinion.
Because of model assumptions that cannot be satisfied and a close empirical data dependency of parametric and non-parametric regression methods, it is very uncertain that these models can be transferred to other regions. Owing to their flexible data use, ANNs also have a high potential for larger-scale applications; however, this model type requires a great deal of data for training. If the model has been trained in a region with different data availabilities, experience in computer programming and model building is needed to adopt the model to a new region. IBMs also hold promising perspectives as they are based on biological behaviour responses and therefore less dependent on in situ data. However, detailed information on the spatial variation in physical habitat variables is required, and the hydraulic model needs a 3D structure which complicates its operation.
Among the two most suitable instream physical habitat model candidates, the HSI and fuzzy logic-based methods, the use of representative HSIs is very important for successful HSI modelling. However, the fuzzy rule-based model can be based solely on expert opinions, but this method can be very subjective and perhaps less convincing in the eyes of managers and other stakeholders. Consequently, if HSIs are validated, the HSI model is probably preferable, but if data availability is sparse, the fuzzy rule-based method should be used. Fuzzy-based rules can also be based on available field data. If field observations are imprecise, fuzzy rule-based curves are the preferred option.
4.8 Generalized habitat models
Generalized habitat models essentially perform the same function as hydraulic–habitat models, but hold advantages over them as they do not inherit their complexity and their high field and biological data requirements (Lamouroux et al. Citation1998, Lamouroux and Capra Citation2002). However, due to their relative simplicity, information is sacrificed. This loss of information must be weighed up against the intensive fieldwork requirements of conventional hydraulic–habitat models, especially if HSIs have to be devised for the conventional modelling approach. Therefore, this model type is very attractive to water managers as they are widely applicable with a limited amount of field work (Lamouroux and Jowett Citation2005, Lamouroux and Cattaneo Citation2006).
5 Conclusion
As a management-based modelling approach, there are substantial differences between the instream physical habitat modelling types available to water managers. HSI and fuzzy logic-based methods appear to be most ready for operational application by water managers. These models are well documented, user-friendly, easily presentable to stakeholders and transferable to be applied over large spatial scales. Furthermore, they can deal with different types of data and they are applicable to a range of biota. However, for HSI modelling, it is essential that care is taken in the use of appropriate HSIs as these are the most sensitive part of the modelling. Inaccurate results will be gained if HSIs are not correctly formulated.
Other methods are also suitable, especially parametric and non-parametric approaches but restrictive model assumptions and their applicability over a limited spatial scale remains a substantial disadvantage. Fuzzy-based models hold promise in either replacing HSIs or supplementing them and may be a better alternative than HSIs for biota where little is known of their biota–physical habitat relationships (e.g. macrophytes and diatoms). The remaining model types will certainly increase in use as they become more developed, refined and independently tested in the research field, but at present they remain research or academic-based. However, modelling types such as ANNs and IBMs that take away the problems of in situ restrictions that many other model types have hold promise and should be researched further as they are universal and therefore widespread applicable, a major attribute for all water managers. In relation to hydraulic–habitat modelling, generalized habitat modelling approaches hold promise for reducing the fieldwork and experience needed in conventional hydraulic–habitat approaches.
Acknowledgements
The authors of this paper would like to thank Ian Jowett, Thom Hardy and Mike Dunbar for their comments and help with the paper. This study was partly funded by the project ‘Climate change impact on ecological conditions in streams’, contract no. 274-06-0474.
Related Research Data
References
- Adriaenssens , V. 2004 . Fuzzy rule-based models for decision support in ecosystem management . Science of the Total Environment , 319 ( 1–3 ) : 1 – 12 .
- Adriaenssens , V. , De Baets , B. , Goethals , P. L.M. and De Pauw , N. 2004 . Fuzzy rule-based models for decision support in ecosystem management . Science of the Total Environment , 319 ( 1–3 ) : 1 – 12 .
- Adriaenssens , V. , Goethals , P. L.M. and De Pauw , N. 2006 . Fuzzy knowledge-based models for prediction of Asellus and Gammarus in watercourses in Flanders (Belgium) . Ecological Modelling , 195 ( 1–2 ) : 3 – 10 .
- Affifi , A. A. and Clark , V. 1996 . Computer-aided multivariate analysis , New York : Chapman and Hall .
- Agresti , A. 1996 . An introduction to categorical data analysis , New York : John Wiley and Sons .
- Ahmadi-Nedushan , B. 2006 . A review of statistical methods for the evaluation of aquatic habitat suitability for instream flow assessment . River Research and Applications , 22 ( 5 ) : 503 – 523 .
- Ahmadi-Nedushan , B. 2008 . Instream flow determination using a multiple input fuzzy-based rule system: a case study . River Research and Applications , 24 ( 3 ) : 279 – 292 .
- Annoni , P. , Saccardo , I. , Gentili , G. and Guzzi , L. 1997 . A mulivariate model to relate hydological, chemical and biologocial parameters to salmonid biomass in Italian rivers . Fisheries Management and Ecology , 4 ( 6 ) : 439 – 452 .
- Bardonnet , A. , Poncin , P. and Roussel , J. M. 2006 . Brown trout fry move inshore at night: a choice of water depth or velocity? . Ecology of Freshwater Fish , 15 ( 3 ) : 309 – 314 .
- Beecher , H. A. , Caldwell , B. A. and Demond , S. B. 2002 . Evaluation of depth and velocity preferences of juvenile coho salmon in Washington streams . North American Journal of Fisheries Management , 22 ( 3 ) : 785 – 795 .
- Beecher , H. A. , Johnson , T. H. and Carleton , J. P. 1993 . Predicting microdistributions of steelhead (Oncorhynchus-mykiss) parr from depth and velocity preference criteria – test of an assumption of the instream flow incremental methodology . Canadian Journal of Fisheries and Aquatic Sciences , 50 ( 11 ) : 2380 – 2387 .
- Bonabeau , E. 2002 . Agent-based modeling: methods and techniques for simulating human systems . Proceedings of the National Academy of Sciences of the United States of America , 99 ( Suppl. 3 ) : 7280 – 7287 .
- Booker , D. J. , Dunbar , M. J. and Ibbotson , A. 2004 . Predicting juvenile salmonid drift-feeding habitat quality using a three-dimensional hydraulic-bioenergetic model . Ecological Modelling , 177 : 157 – 177 .
- Bovee , K. D. 1998 . Stream habitat analysis using the instream flow incremental methodology , Fort Collins : US Geological Survey . Biological Resources Discipline Information and Technology Report USGS/BRD-1998-0004
- Bovee , K. D. 1986 . Development and evaluation of habitat criteria for use in instream flow incremental methodology , Fort Collins : US Fish and Wildlife Service . Instream Flow Information Paper 21, FWS/OBS/86/07
- Bovee , K. D. and Cochnauer , T. 1977 . Development and evaluation of weighted criteria, probability of use curves for instream flow assessment: fisheries , Fort Collins : US Fish and Wildlife Service . FWS/OBS-77/63
- Bovee , K. D. and Milhous , R. T. 1978 . Hydraulic simulation in instream flow studies: theory and techniques , Fort Collins : US Fish and Wildlife Service . Instream Flow Information Paper 5, FWS/OBS/78/33
- Brant , S. B. and Hartmann , K. J. 1993 . Innovative approaches with bioenergetics models: future applications to fish ecology and management . Transactions of American Fisheries Society , 122 : 731 – 735 .
- Brey , T. , Jarreteichmann , A. and Borlich , O. 1996 . Artificial neural network versus multiple linear regression: Predicting P/B ratios from empirical data . Marine Ecology-Progress Series , 140 ( 1–3 ) : 251 – 256 .
- Brosse , S. , Guegan , J. F. , Tourenq , J. N. and Lek , S. 1999a . The use of artificial neural networks to assess fish abundance and spatial occupancy in the littoral zone of a mesotrophic lake . Ecological Modelling , 120 ( 2–3 ) : 299 – 311 .
- Brosse , S. , Lek , S. and Dauba , F. 1999b . Predicting fish distribution in a mesotrophic lake by hydroacoustic survey and artificial neural networks . Limnology and Oceanography , 44 ( 5 ) : 1293 – 1303 .
- Bult , T. P. , Riley , S. C. , Haedrich , R. L. , Gibson , R. J. and Heggenes , J. 1999 . Density-dependent habitat selection by juvenile Atlantic salmon (Salmo salar) in experimental riverine habitats . Canadian Journal of Fisheries and Aquatic Sciences , 56 ( 7 ) : 1298 – 1306 .
- Bunn , S. E. and Arthington , A. H. 2002 . Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity . Environmental Management , 30 ( 4 ) : 492 – 507 .
- Cantrall , C. J. , Robinson , A. T. and Avenetti , L. D. 2005 . Habitat selection by Apache trout in six east-central Arizona streams . Transactions of American Fisheries Society , 134 : 1382 – 1388 .
- Castella , E. , Adalsteinsson , H. , Brittain , J. E. , Gislason , G. M. , Lehmann , A. , Lencioni , V. , Lods-Crozet , B. , Maiolini , B. , Milner , A. M. , Olafsson , J. S. , Saltveit , S. J. and Snook , D. L. 2001 . Macrobenthic invertebrate richness and composition along a latitudinal gradient of European glacier-fed streams . Freshwater Biology , 46 ( 12 ) : 1811 – 1831 .
- Cavendish , M. G. and Duncan , M. I. 1986 . Use of the instream flow incremental methodology: a tool for negotiation . Environmental Impact Assessment Review , 6 : 347 – 363 .
- Chen , Q. and Mynatt , A. E. 2003 . Integration of data mining techniques and heuristic knowledge in fuzzy logic modelling of eutrophication in Taihu Lake . Ecological Modelling , 162 : 55 – 67 .
- Cobb , D. G. , Galloway , T. D. and Flannagan , J. F. 1992 . Effect of discharge and substrate stability on density and species composition of stream insects . Canadian Journal of Fisheries and Aquatic Sciences , 49 ( 9 ) : 1788 – 1795 .
- Commission , E. 2000 . Directive 2000/60/EC. Establishing a framework for community action in the field of water policy , Luxembourg : European Commission . PE-CONS 3639/1/100 Rev 1
- Cornelissen , A. M.G. 2001 . Assessment of the contribution of sustainability indicators to sustainable development: a novel approach using fuzzy set theory . Agricultural and Ecosystem Environment , 8 : 173 – 185 .
- Dakou , E. , Goethals , P. L.M. , D'heygere , T. , Dedecker , A. P. , Gabriels , W. , De Pauw , N. and Lazaridou-Dimitriadou , M. 2006 . Development of artificial neural network models predicting macroinvertebrate taxa in the river Axios (Northern Greece) . Annales De Limnologie-International Journal of Limnology , 42 ( 4 ) : 241 – 250 .
- Dedecker , A. P. , Goethals , P. L.M. , D'heygere , T. , Gevrey , M. , Lek , S. and De Pauw , N. 2005 . Application of artificial neural network models to analyse the relationships between Gammarus pulex L. (Crustacea, Amphipoda) and river characteristics . Environmental Monitoring and Assessment , 111 ( 1–3 ) : 223 – 241 .
- Dedecker , A. P. , Goethals , P. L.M. , Gabriels , W. and Pauwn , D. 2001 . River management applications of ecosystem models predicitng aquatic macroinvertebrate communities based on artificial neural networks (ANNs) . Mededelingen Faculteit Landbouwkundige en Toegepaste Biolgische Wetenschappen , 66 ( 4 ) : 207 – 211 .
- Dunbar , M. 2001 . Further validation of PHABSIM for the habitat requirements of salmonid fish , UK : Centre for Ecology and Hydrology . R&D Report W6-036
- Dunbar , M. J. 1998 . Overseas approaches to setting river flow objectives , Wallingford : Environment Agency and NERC . R&D Technical Report W6-161
- Edmondson , W. T. and Winberg , G. G. 1971 . A manual on methods for the assessment of secondary productivity in fresh waters , Oxford : Blackwell .
- Egglishaw , H. J. and Shackley , P. E. 1982 . Influence of water depth on dispersion of juvenile salmonids, Salmo-salar L and Salmo-trutta-L, in a Scottish stream . Journal of Fish Biology , 21 ( 2 ) : 141 – 155 .
- Elith , J. and Leathwick , J. 2007 . Predicting species distributions from museum and herbarium records using multiresponse models fitted with multivariate adaptive regression splines . Diversity and Distributions , 13 ( 3 ) : 265 – 275 .
- Filipe , A. F. , Cowx , I. G. and Collares-Pereira , M. J. 2002 . Spatial modelling of freshwater fish in semi-arid river systems: a tool for conservation . River Research and Applications , 18 ( 2 ) : 123 – 136 .
- Friedman , J. H. 1991 . Multivariate Adaptive Regression Splines (with discussion) . Annals of Statistics , 19 : 1 – 141 .
- Garland , R. D. , Tiffan , K. F. , Rondorf , D. W. and Clark , L. O. 2002 . Comparison of subyearling fall chinook salmon's use of riprap, revetments and unaltered habitats in Lake Wallula of the Columbia River . North American Journal of Fisheries Management , 22 ( 4 ) : 1283 – 1289 .
- Geist , D. R. , Jones , J. , Murray , C. J. and Dauble , D. D. 2000 . Suitability criteria analyzed at the spatial scale of redd clusters improved estimates of fall chinook salmon (Oncorhynchus tshawytscha) spawning habitat use in the Hanford Reach, Columbia River . Canadian Journal of Fisheries and Aquatic Sciences , 57 ( 8 ) : 1636 – 1646 .
- Gibbins , C. N. and Acornley , R. M. 2000 . Salmonid habitat modelling studies and their contribution to the development of an ecologically acceptable release policy for Kielder Reservoir, North-East England . Regulated Rivers-Research and Management , 16 ( 3 ) : 203 – 224 .
- Gordon , N. D. 2004 . Stream hydrology: an introduction for ecologists , 2 , Chichester : John Wiley and Sons .
- Gore , J. A. and Nestler , J. M. 1988 . Instream flow studies in perspective . Regulated Rivers – Research and Management , 2 : 93 – 101 .
- Grand , T. C. , Railsback , S. F. , Hayse , J. W. and Lagory , K. E. 2006 . A physical habitat model for predicting the effects of flow fluctuations in nursery habitats of the endangered Colorado pikeminnow (Ptychocheilus lucius) . River Research and Applications , 22 ( 10 ) : 1125 – 1142 .
- Gray , B. R. 2005 . Selecting a distributional assumption for modelling relative densities of benthic macroinvertebrates . Ecological Modelling , 185 ( 1 ) : 1 – 12 .
- Grimm , V. and Railsback , S. F. 2005 . Individual-based modelling and ecology , Princeton University .
- Groshens , T. P. and Orth , D. J. 1994 . Transferability of habitat suitability criteria for smallmouth bass . Micropterus dolomieu. Rivers , 4 ( 3 ) : 194 – 212 .
- Guay , J. C. 2000 . Development and validation of numerical habitat models for juveniles of Atlantic salmon (Salmo salar) . Canadian Journal of Fisheries and Aquatic Sciences , 57 ( 10 ) : 2065 – 2075 .
- Guisan , A. , Edwards , T. C. and Hastie , T. 2002 . Generalised linear and generalized additive models in studies of species distributions: setting the scene . Ecological Modelling , 157 : 89 – 100 .
- Guisan , A. and Zimmermann , N. E. 2000 . Predictive habitat distribution models in ecology . Ecological Modelling , 135 ( 2–3 ) : 147 – 186 .
- Hansen , M. J. , Boisclair , D. , Brandt , S. B. , Hewett , S. W. , Kitchell , J. F. , Lucas , M. C. and Ney , J. J. 1993 . Applications of bioenergetics models to fish ecology and management - where do we go from here . Transactions of the American Fisheries Society , 122 ( 5 ) : 1019 – 1030 .
- Harby , A. 2004 . Cost 626 report: state-of-the-art in data sampling, modelling analysis and applications of river habitat modelling , France : European Aquatic Modelling Network .
- Hardy , T. B. 1998 . The future of habitat modeling and instream flow assessment techniques . Regulated Rivers – Research and Management , 14 ( 5 ) : 405 – 420 .
- Harvey , B. C. and Railsback , S. F. 2007 . Estimating multi-factor cumulative watershed effects on fish populations with an individual-based model . Fisheries , 32 ( 6 ) : 292 – 298 .
- Harvey , B. C. , White , J. L. and Nakamoto , R. J. 2002 . Habitat relationships and larval drift of native and nonindigenous fishes in neighboring tributaries of a coastal California river . Transactions of American Fisheries Society , 131 : 159 – 170 .
- Hastie , T. , Tibshirani , R. J. and Friedman , J. H. 2001 . “ The elements of statistical learning: data mining, Inference and Prediction ” . Springer Series in Statistics New York : Springer-Verlag .
- Hayes , J. W. and Jowett , I. G. 1994 . Microhabitat models of large drift-feeding brown trout in three New Zealand rivers . North American Journal of Fisheries Management , 14 : 710 – 725 .
- Hayes , J. W. , Stark , J. D. and Shearer , K. A. 2000 . Development and test of a whole lifetime foraging and bioenergetics model for drift-feeding brown trout . Transactions of American Fisheries Society , 129 : 315 – 332 .
- Heggenes , J. 1988 . Substrate preferences of brown trout fry (Salmo-trutta) in artificial stream channels . Canadian Journal of Fisheries and Aquatic Sciences , 45 ( 10 ) : 1801 – 1806 .
- Heggenes , J. and Traaen , T. 1988 . Daylight responses to overhead cover in stream channels for fry of 4 salmonid species . Holarctic Ecology , 11 ( 3 ) : 194 – 201 .
- Hill , J. and Grossman , G. D. 1993 . An energetic model of microhabitat use for rainbow-trout and rosyside dace . Ecology , 74 ( 3 ) : 685 – 698 .
- Holm , C. F. , Armstrong , J. D. and Gilvear , D. J. 2001 . Investigating a major assumption of predictive instream habitat models: is water velocity preference of juvenile Atlantic salmon independent of discharge? . Journal of Fish Biology , 59 ( 6 ) : 1653 – 1666 .
- Hughes , N. F. 1998 . A model of habitat selection by drift-feeding stream salmonids at different scales . Ecology , 79 ( 1 ) : 281 – 294 .
- Hughes , N. F. 1992 . Selection of positions by drift-feeding salmonids in dominance hierarchies - model and test for arctic grayling (Thymallus-arcticus) in sub-arctic mountain streams, interior Alaska . Canadian Journal of Fisheries and Aquatic Sciences , 49 ( 10 ) : 1999 – 2008 .
- Hughes , N. F. and Dill , L. M. 1990 . Position choice by drift-feeding salmonids - model and test for arctic grayling (Thymallus-arcticus) in sub-arctic mountain streams, interior Alaska . Canadian Journal of Fisheries and Aquatic Sciences , 47 ( 10 ) : 2039 – 2048 .
- Humphries , P. , King , A. J. and Koehn , J. D. 1999 . Fish, flows and flood plains: links between freshwater fishes and their environment in the Murray–Darling River system, Australia . Environmental Biology of Fishes , 56 ( 1–2 ) : 129 – 151 .
- Inoue , M. and Nakano , S. 2001 . Fish abundance and habitat relationships in forest and grassland streams, northern Hokkaido, Japan . Ecological Research , 16 ( 2 ) : 233 – 247 .
- Ivlev , V. S. 1961 . Experimental ecology of the feeding fishes , New Haven : Yale University Press .
- Jacobs , J. 1974 . Quantitative measurement of food selection: a modification of the forage ratio and Ivlev's electivity index . Oecologia , 14 : 413 – 417 .
- Jacobsen , D. 2005 . Temporally variable macroinvertebrate-stone relationships in streams . Hydrobiologia , 544 ( 201–214 )
- Jongman , R. H.G. , Peres-Neto , P. A. and Olden , J. D. 1995 . Data analysis in community and landscape ecology , Cambridge University Press .
- Jorde , K. 2001 . “ Fuzzy based models for the evaluation of fish habitat quality and instream flow assessment ” . In Proceedings of the 3rd international symposium on environmental hydraulics Tempe, AZ
- Jorgensen , S. E. and Bendoricchio , G. 2001 . Fundamentals of ecological modelling , 3 , Oxford : Cambridge University Press .
- Jorgensen , S. E. 2008 . Overview of the model types available for development of ecological models . Ecological Modelling , 215 ( 1–3 ) : 3 – 9 .
- Jowett , I. 1992 . Models of the abundance of large brown trout in New Zealand rivers . North American Journal of Fisheries Management , 12 : 417 – 432 .
- Jowett , I. G. 1997 . Instream flow methods: A comparison of approaches . Regulated Rivers-Research & Management , 13 ( 2 ) : 115 – 127 .
- Jowett , I. G. 2006 . RHYHABSIM river hydraulics and habitat simulation , Software manual
- Jowett , I. G. and Davey , A. J.H. 2007 . A comparison of composite habitat suitability indices and generalized additive models of invertebrate abundance and fish presence–habitat availability . Transactions of the American Fisheries Society , 136 ( 2 ) : 428 – 444 .
- Jowett , I. G. , Richardson , J. and Bonnett , M. L. 2005 . Relationship between flow regime and fish abundances in a gravel-bed river, New Zealand . Journal of Fish Biology , 66 ( 5 ) : 1419 – 1436 .
- Kennedy , G. J.A. and Strange , C. D. 1982 . The distribution of salmonids in upland streams in relation to depth and gradient . Journal of Fish Biology , 20 ( 5 ) : 579 – 591 .
- Kerle , F. , Zöllner , M. , Schneider , B. , Kappus and Baptist , M. J. Monitoring of long-term fish habitat changes in restored secondary floodplains channels of the River Rhine . Fourth International Ecohydraulics Symposium . Cape Town, South Africa. 3–8 March
- Knapp , R. A. and Preisler , H. K. 1999 . Is it possible to predict habitat use by spawning salmonids? A test using California golden trout (Oncorhynchus mykiss aguabonita) . Canadian Journal of Fisheries and Aquatic Sciences , 56 ( 9 ) : 1576 – 1584 .
- Kondolf , G. M. 2000 . Assessing salmonid spawning gravel quality . Transactions of the American Fisheries Society , 129 ( 1 ) : 262 – 281 .
- Korman , J. , Perrin , C. J. and Lekstrum , T. 1994 . A guide for the selection of standard methods for quantifying sportfish habitat capability and suitability in streams and lakes of British Columbia , Vancouver, BC : BC Environment Fisheries Branch .
- Labonne , J. , Allouche , S. and Gaudin , P. 2003 . Use of a generalised linear model to test habitat preferences: the example of Zingel asper, an endemic endangered percid of the River Rhone . Freshwater Biology , 48 ( 4 ) : 687 – 697 .
- Lamouroux , N. and Capra , H. 2002 . Simple predictions of instream habitat model outputs for target fish populations . Freshwater Biology , 47 ( 8 ) : 1543 – 1556 .
- Lamouroux , N. , Capra , H. and Pouilly , M. 1998 . Predicting habitat suitability for lotic fish: linking statistical hydraulic models with multivariate habitat use models . Regulated Rivers-Research and Management , 14 ( 1 ) : 1 – 11 .
- Lamouroux , N. and Cattaneo , F. 2006 . Fish assemblages and stream hydraulics: consistent relations across spatial scales and regions . River Research and Applications , 22 ( 7 ) : 727 – 737 .
- Lamouroux , N. and Jowett , I. G. 2005 . Generalized instream habitat models . Canadian Journal of Fisheries and Aquatic Sciences , 62 ( 1 ) : 7 – 14 .
- Lasne , E. 2007 . Fish zonation and indicator species for the evaluation of the ecological status of rivers: example of the Loire Basin (France) . River Research and Applications , 23 ( 8 ) : 877 – 890 .
- Leathwick , J. R. , Elith , J. and Hastie , T. 2006 . Comparative performance of generalized additive models and multivariate adaptive regression splines for statistical modelling of species distributions . Ecological Modelling , 199 ( 2 ) : 188 – 196 .
- Leathwick , J. R. , Rowe , D. , Richardson , J. , Elith , J. and Hastie , T. 2005 . Using multivariate adaptive regression splines to predict the distributions of New Zealand's freshwater diadromous fish . Freshwater Biology , 50 ( 12 ) : 2034 – 2052 .
- Leclerc , M. A. , St-Hilaire , A. and Bechara , J. A. 2003 . State-of-the-art and perspectives on habitat modelling . Canadian Water Resources Journal , 28 ( 2 ) : 153 – 172 .
- Lek , S. , Belaud , A. , Baran , P. , Dimopoulos , I. and Delacoste , M. 1996a . Role of some environmental variables in trout abundance models using neural networks . Aquatic Living Resources , 9 ( 1 ) : 23 – 29 .
- Lek , S. , Delacoste , M. , Baran , P. , Dimopoulos , I. , Lauga , J. and Aulagnier , S. 1996b . Application of neural networks to modelling nonlinear relationships in ecology . Ecological Modelling , 90 ( 1 ) : 39 – 52 .
- Lek , S. and Guegan , J. F. 1999 . Artificial neural networks as a tool in ecological modelling, an introduction . Ecological Modelling , 120 ( 2–3 ) : 65 – 73 .
- Lencioni , V. 2006 . Macroinvertebrate assemblages in glacial systems: a comparison of linear multivariate methods with artificial neural networks . Ecological Modelling , 203 ( 1–2 ) : 119 – 131 .
- Logan , P. and Furse , M. 2002 . Preparing for the European Water Framework Directive – making the links between habitat and aquatic biota . Aquatic Conservation-Marine and Freshwater Ecosystems , 12 ( 4 ) : 425 – 437 .
- Loia , V. , Sessa , S. , Staiano , A. and Tagliaferri , R. 2000 . Merging fuzzy logic, neural networks, and genetic computation in the design of a decision-support system . International Journal of Intelligent Systems , 15 ( 7 ) : 575 – 594 .
- Mackinson , S. 2000 . An adaptive fuzzy expert system for predicting structure, dynamics and distribution of herring shoals . Ecological Modelling , 126 ( 2–3 ) : 155 – 178 .
- Maddock , I. 1999 . The importance of physical habitat assessment for evaluating river health . Freshwater Biology , 41 ( 2 ) : 373 – 391 .
- Mastrorillo , S. , Dauba , F. , Oberdorff , T. , Guégan , J. F. and Lek , S. 1997 . Prediciting local fish species in the Garonne River basin . C.R. Académie des Science , 321 : 423 – 428 .
- Mathur , D. 1985 . A critique of the instream flow incremental methodology . Canadian Journal of Fisheries and Aquatic Sciences , 42 ( 4 ) : 825 – 831 .
- May , R. J. , Maier , H. R. , Dandy , G. C. and Fernando , T. 2008 . Non-linear variable selection for artificial neural networks using partial mutual information . Environmental Modelling & Software , 23 ( 10–11 ) : 1312 – 1326 .
- Meesters , E. H. , Bak , R. P.M. , Westmacott , S. , Ridgley , M. and Dollar , S. 1997 . A fuzzy logic model to predict coral reef development under nutrient and sediment stress . Conservation Biology , 12 ( 5 ) : 957 – 965 .
- Moir , H. J. 2005 . PHABSIM modelling of Atlantic salmon spawning habitat in an upland stream: testing the influence of habitat suitability indices on model output . River Research and Applications , 21 ( 9 ) : 1021 – 1034 .
- Mouton , A. M. 2008 . Optimisation of a fuzzy physical habitat model for spawning European grayling (Thymallus thymallus L.) in the Aare river (Thun, Switzerland) . Ecological Modelling. 215(1–3), 122–132
- Munoz , J. and Fellicísmo , A. M. 2004 . Comparison of statistical methods used in predictive modelling . Journal of Vegetation Science , 15 : 285 – 292 .
- Naiman , J. R. , Bunn , S. E , Nilsson , C. , Petts , G. E. , Pinay , G. and Thompson , L. C. 2002 . Legitimising fluvial ecosystems as users of water: An overview . Environmental Management , 30 ( 4 ) : 455 – 467 .
- Narumalani , S. , Jensen , J. R. , Althausen , J. D. , Burkhalter , S. and Mackey , H. E. 1997 . Aquatic macrophyte modeling using GIS and logistic multiple regression . Photogrammetric Engineering and Remote Sensing , 63 ( 1 ) : 41 – 49 .
- Neumann , R. M. and Wildman , T. L. 2002 . Relationships between trout habitat use and woody debris in two southern New England streams . Ecology of Freshwater Fish , 11 ( 4 ) : 240 – 250 .
- Ney , J. J. 1993 . Bioenergetics modeling today - growing pains on the cutting edge . Transactions of the American Fisheries Society , 122 ( 5 ) : 736 – 748 .
- Nykanen , M. , Huusko , A. and Maki-Petays , A. 2001 . Seasonal changes in the habitat use and movements of adult European grayling in a large subarctic river . Journal of Fish Biology , 58 ( 2 ) : 506 – 519 .
- Olden , J. D. and Jackson , D. A. 2002 . Illuminating the ‘black box’: understanding variable contributions in artificial neural networks . Transactions of American Fisheries Society , 154 : 135 – 150 .
- Olsen , M. 2009 . Groundwater abstraction impacts on physical habitat of brown trout (Salmo trutta) in a small Danish stream . Hydrology Research , 40 ( 4 ) : 394 – 405 .
- Orth , D. J. and Maughen , O. E. 1982 . Evaluation of the incremental methodology for recommending instream flows for fishes . Transactions of American Fisheries Society , 111 : 413 – 445 .
- Parasiewicz , P. and Dunbar , M. J. 2001 . Physical habitat modelling for fish - a developing approach . Large Rivers , 12 : 239 – 268 .
- Park , Y. S. , Grenouillet , G. , Esperance , B. and Lek , S. 2006 . Stream fish assemblages and basin land cover in a river network . Science of the Total Environment , 365 ( 1–3 ) : 140 – 153 .
- Payne and Associates . 2000 . RHABSIM . Fisheries Biology. Consulting and Software Publishing, Arcata, CA. Available from: http://trpafishbiologists.com/rindex.html
- Poff , N. L. 1997 . The natural flow regime . Bioscience , 47 ( 11 ) : 769 – 784 .
- Pollard , P. and Huxham , M. 1998 . The European Water Framework Directive: a new era in the management of aquatic ecosystem health? . Aquatic Conservation-Marine and Freshwater Ecosystems , 8 ( 6 ) : 773 – 792 .
- Pothoven , S. A. , Fahnenstiel , G. L. and Vanderploeg , H. A. 2001 . Population dynamics of Bythotrephes cederstroemii in south-east Lake Michigan, 1995–1998 . Freshwater Biology , 46 ( 11 ) : 1491 – 1501 .
- Quinn , G. P. and Keough , M. J. 2006 . Experimental design and data analysis for biologists , Cambridge University Press .
- Railsback , S. F. , Stauffer , H. B. and Harvey , B. C. 2003 . What can habitat preference models tell us? Tests using a virtual trout population . Ecological Applications , 13 ( 6 ) : 1580 – 1594 .
- Railsback , S. F. and Harvey , B. C. 2002 . Analysis of habitat-selection rules using an individual-based model . Ecology , 83 ( 7 ) : 1817 – 1830 .
- Raleigh , R. F. , Zuckerman , L. D. and Nelson , P. C. 1986 . Habitat suitability models and instream flow suitability curves: Brown Trout , Washington, DC : US Department of the Interior, Fish and Wildlife Service . Biological Report 82(10.124)
- Rand , P. S. 1995 . Effect of lake-wide planktivory by the pelagic prey fish community in lakes Michigan and Ontario . Canadian Journal of Fisheries and Aquatic Sciences , 52 ( 7 ) : 1546 – 1563 .
- Rosenfeld , J. 2003 . Assessing the habitat requirements of stream fishes: an overview and evaluation of different approaches . Transactions of American Fisheries Society , 57 ( 4 ) : 766 – 774 .
- Schneider , M. and Peter , A. Okostrom: field study and use of the simulation model CASIMIR for determination of fish habitat in River Brenno . Proceedings of the 3rd international symposium on ecohydraulics . Salt Lake City, Utah.
- Schweizer , S. 2007 . Predicting joint frequency distributions of depth and velocity for instream habitat assessment . River Research and Applications , 23 ( 3 ) : 287 – 302 .
- Silvert , W. 1997 . Ecological impact classification with fuzzy sets . Ecological Modelling , 96 ( 1–3 ) : 1 – 10 .
- Silvert , W. 2000 . Fuzzy indices of environmental conditions . Ecological Modelling , 130 ( 1–3 ) : 111 – 119 .
- Stalnaker , C. B. 1995 . The instream flow incremental methodology: a primer for IFIM , Technical Report Series Fort Collins : National Biological Service, Biological Report 29 .
- Stewardson , M. 2005 . Hydraulic geometry of stream reaches . Journal of Hydrology , 306 ( 1–4 ) : 97 – 111 .
- Stewardson , M. J. and McMahon , T. A. 2002 . A stochastic model of hydraulic variations within stream channels . Water Resources Research , 38 ( 1 ) : 14
- Stewart , D. J. and Ibarra , M. 1991 . Predation and production by salmonine fishes in lake-Michigan, 1978–88 . Canadian Journal of Fisheries and Aquatic Sciences , 48 ( 5 ) : 909 – 922 .
- Stillman , R. A. 2008 . MORPH – an individual-based model to predict the effect of environmental change on foraging animal populations . Ecological Modelling , 216 ( 3–4 ) : 265 – 276 .
- Sullivan , S. M.P. , Watzin , M. C. and Hession , W. C. 2006 . Influence of stream geomorphic condition on fish communities in Vermont, USA . Freshwater Biology , 51 ( 10 ) : 1811 – 1826 .
- Sullivan , S. M.P. , Watzin , M. C. and Hession , W. C. 2004 . Understanding stream geomorphic state in relation to ecological integrity: evidence using habitat assessments and macroinvertebrates . Environmental Management , 34 ( 5 ) : 669 – 683 .
- Suren , A. M. and Jowett , I. G. 2006 . Effects of floods versus low flows on invertebrates in a New Zealand gravel-bed river . Freshwater Biology , 51 ( 12 ) : 2207 – 2227 .
- Tharme , R. E. 2003 . A global perspective on environmental flow assessment: emerging trends in the development and application of environmental flow methodologies for rivers . River Research and Applications , 19 ( 5–6 ) : 397 – 441 .
- Thomas , J. A. and Bovee , K. D. 1993 . Application and testing of a procedure to evaluate transferability of habitat suitability criteria . Regulated Rivers – Research and Management , 8 ( 3 ) : 285 – 294 .
- Thorn , P. and Conallin , J. C. 2006 . RHYHABSIM as a stream management tool: case study in the River Kornerup Catchment, Denmark . The Journal of Transdisciplinary Environmental Studies , 5 ( 1–2 ) : 1 – 17 .
- Townend , J. 2002 . Practical statistics for environmental and biological scientists , Chichester, UK : John Wiley and Sons, Ltd .
- Udevitz , M. S. , Bloomfield , P. and Apperson , C. S. 1987 . Prediction of the occurrence of four species of mosquito larvae with logistic regression on water chemistry variables . Environmental Entomology , 16 : 281 – 285 .
- Vadas , R. L. and Orth , D. J. 2001 . Formulation of habitat suitability models for stream fish guilds: do the standard methods work? . Transactions of the American Fisheries Society , 130 ( 2 ) : 217 – 235 .
- Van Broekhoven , E. , Adriaenssens , V. and De Baets , B. 2007 . Interpretability-preserving genetic optimization of linguistic terms in fuzzy models for fuzzy ordered classification: An ecological case study . International Journal of Approximate Reasoning , 44 ( 1 ) : 65 – 90 .
- Van Broekhoven , E. , Adriaenssens , V. , De Baets , B. and Verdonschot , P. F.M. 2006 . Fuzzy rule-based macroinvertebrate habitat suitability models for running waters . Ecological Modelling , 198 ( 1–2 ) : 71 – 84 .
- Van Den Berg , M. S. , Joosse , W. and Coops , H. 2003 . A statistical model predicting the occurrence and dynamics of submerged macrophytes in shallow lakes in the Netherlands . Hydrobiologia , 506 ( 1–3 ) : 611 – 623 .
- Van Winkle , W. , Jager , H. I. , Railsback , S. F. , Holcomb , B. D. , Studley , T. K. and Baldrige , J. E. 1998 . Individual-based model of sympatric populations of brown and rainbow trout for instream flow assessment: model description and calibration . Ecological Modelling , 110 ( 2 ) : 175 – 207 .
- Vehanen , T. , Bjerke , P. L. , Heggenes , J. , Huusko , A. and Maki-Petays , A. 2000 . Effect of fluctuating flow and temperature on cover type selection and behaviour by juvenile brown trout in artificial flumes . Journal of Fish Biology , 56 ( 4 ) : 923 – 937 .
- Vismara , R. 2001 . Habitat suitability curves for brown trout (Salmo trutta fario L.) in the river Adda, Northern Italy: comparing univariate and multivariate approaches . Regulated Rivers – Research and Management , 17 ( 1 ) : 37 – 50 .
- Wagner , T. and Falter , C. M. 2002 . Response of an aquatic macrophyte community to fluctuating water levels in an oligotrophic lake . Lake Reserve Management , 18 ( 1 ) : 52 – 65 .
- Wallin , M. , Wiederholm , D. and Johnson , R. K. 2003 . Guidance on establishing reference conditions and ecological status class boundaries for inland surface waters . CIS Working Group 2.3 – REFCOND, 7th version