
Abstract
Tree ring cellulose oxygen isotopes (δ18O) were measured on 21 trees of Qilian juniper from the Animaqing Mountains, Tibetan Plateau, to investigate intra- and inter-tree variability, potential juvenile and elevation effects and climatic implications. There are no significant differences in mean and standard deviation of tree ring δ18O values at different heights in individual trees. Tree ring δ18O values from different directions show a high degree of coherence. The mean and standard deviation for vertical and circumferential δ18O time series are very similar, and δ18O data from different heights and directions are highly correlated (r > 0.88). The δ18O values of young trees are lower than those of old trees in the first 10 years of tree growth. Tree ring δ18O data from five different altitudes are highly correlated (r > 0.88) and share similar climatic signals. As such, an altitude effect on tree ring δ18O is not observed. Our results indicate that samples from one site, regardless of sampling height, direction or altitude, can be used to reconstruct a long-term δ18O record. Tree ring δ18O data from the Animaqing Mountains show a significant negative correlation (r = −0.67; p < 0.001) with May–July regional precipitation and appear to be a promising proxy for precipitation reconstruction.
1. Introduction
Climate change has profound societal and economic effects (Stocker et al., Citation2013). High-resolution paleoclimate records that can be calibrated and verified with instrumental climate data have been used to reconstruct past climate change with defined uncertainties. These have great potential to provide long-term records of past climate (McCarroll, Citation2015). In addition, such records are helpful in evaluating and improving climate models (McCarroll, Citation2015).
Given their high-resolution nature and ability to be accurately dated, tree rings have been widely used to reconstruct past climate change by calibrating and verifying the tree ring records with measured meteorological records (Cook et al., Citation2010; Shao et al., Citation2010). Presently, most of the longest tree ring-based reconstructions from China are based on the tree ring width of juniper (Gou et al., Citation2010; Shao et al., Citation2010; Yang et al., Citation2014; Zhang et al., Citation2015; Chen et al., Citation2016). Yang et al. (Citation2014) reconstructed annual precipitation variations over the past 3500 years based on a combined tree ring width study of sub-fossil, archaeological and living juniper samples from the north-eastern Tibetan Plateau. Chen et al. (Citation2016) reconstructed April–June maximum temperature variations based on a 2665-year-long tree ring width study of Qilian juniper from the upper treeline of the Animaqing Mountains on the eastern Tibetan Plateau. Compared with this single tree ring width index, other tree-related proxies have the potential to contain more paleoclimate information. For example, tree ring δ18O and δ13C data have previously been used as proxies for hydroclimatic parameters in Asia at both local and regional scales (Treydte et al., Citation2006; Grießinger et al., Citation2011; Sano et al., Citation2012a; An et al., Citation2014; Liu et al., Citation2014; Wernicke et al., Citation2015). Therefore, it is desirable to use multi-proxy approaches to dendroclimatology, which may yield a wide variety of paleo-environmental information from the same site (Gagen et al., Citation2006).
Stable oxygen isotopes (δ18O) in tree rings are controlled mainly by source water δ18O and relative humidity (Roden et al., Citation2000). Tree ring δ18O values have the potential to record climate signals and provide different climatic information as compared with tree ring width (Xu et al., Citation2011a, Citation2013a, Citation2015; Sano et al., Citation2012b). Furthermore, tree ring δ18O values have the potential to preserve low-frequency climate signals (Gagen et al., Citation2011). During the past decade, many tree ring cellulose oxygen isotope studies on the Tibetan Plateau have explored past climatic change (Treydte et al., Citation2006; Grießinger et al., Citation2011, Citation2017; Shi et al., Citation2011; Xu et al., Citation2011b; Sano et al., Citation2012a; Liu et al., Citation2013, Citation2014; An et al., Citation2014; Qin et al., Citation2014; Wernicke et al., Citation2015). Some of these data were used to reconstruct hydroclimate (Treydte et al., Citation2006; Grießinger et al., Citation2011, Citation2017; Sano et al., Citation2012a; An et al., Citation2014; Wernicke et al., Citation2015). Most of these stable oxygen isotope studies were based on spruce or fir species, which have relatively wide tree rings from which it is easy to extract cellulose.
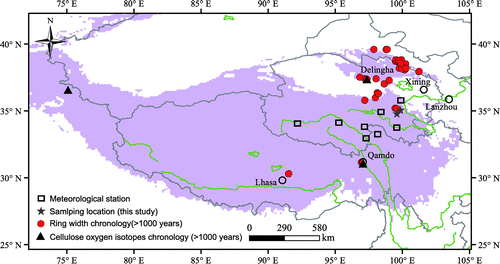
However, junipers grow very slowly, which makes it difficult to establish a δ18O time series with annual resolution (Wang et al., Citation2013), which is the reason that number of long-term ring width chronology (Fig. , red spots) is large, while the number of long-term oxygen isotope chronology is limited (Fig. , black triangles). Therefore, a better way to establish annually resolved time series from juniper is to combine multiple records from different trees/cores that have relatively wide rings in some periods. However, the cores with relatively wide tree rings are typically from different altitudes and sampling directions, and also have different ages. Climatic response of tree ring width for juniper at different altitudes might be variable. It has been reported that tree growth near the upper treeline is controlled by temperature, whereas at low altitudes it is governed by moisture (Takahashi and Yasue, Citation2003; Savva et al., Citation2006). In addition, age-related effects on tree ring δ18O values have been found for juniper in Pakistan and pine in Spain (Treydte et al., Citation2006; Esper et al., Citation2010), although such effects have not been observed for pine in south-east China and larch in Bhutan (Sano et al., Citation2013; Xu et al., Citation2016).
Fig. 1. Map of the study area. Red spots and black triangles indicate ring width and cellulose oxygen isotope data (>1000 yr), respectively. Stars and rectangles indicate sampling locations and meteorological stations, respectively.
In this study, oxygen isotopes of tree ring samples from different altitudes, and variable sampling heights, directions and ages were analysed to investigate: (1) intra- and inter-tree oxygen isotope variations; (2) potential juvenile effects on tree ring oxygen isotopes; (3) tree ring oxygen isotope changes along an altitudinal gradient; and (4) the climatic implications of Qilian juniper oxygen isotope data from the Animaqing Mountains on the eastern Tibetan Plateau.
2. Materials and methods
2.1. Sampling site
Tree ring samples were taken from Qilian juniper (Juniperus przewalskii Kom.) growing in natural forests in the Animaqing Mountains in Qinghai Province (Fig. ). The two sampling locations (YK and ND) are open canopy sites (YK = 34.76°N, 99.69°E and 3800 m asl; ND = 35.00°N, 100.07°E and 3400 m asl). Core samples of each tree were collected using a 5-mm-diameter increment borer. The cores were air dried and polished to make the tree ring borders clearly visible. The ring widths of the samples were then measured at a resolution of 0.01 mm using a binocular microscope with a linear stage interfaced with a computer (Velmex™; ACU-RITE). Cross-dating was performed in the laboratory by matching variations in ring width from all cores to determine the absolute annual age of each ring. Quality control was completed using the program COFECHA (Holmes, Citation1983).
2.2. Samples for oxygen isotope measurements
Four cores from one tree with different sampling directions (north, south, east and west) and two cores from one tree with different sampling heights (15 and 70 cm) at the ND site were used to check the circumferential and vertical variability of δ18O in a single tree. Twenty trees from different altitudes (4250, 4150, 4000, 3900 and 3800 m) were selected to test the effect of altitude on cellulose δ18O. Two relatively old trees (>200 yr) and four relatively young trees (<90 yr) with pith were used to evaluate potential juvenile effects on tree ring cellulose δ18O. All of these experiments were designed to develop a sampling protocol for reconstructing a long-term tree ring δ18O record in Qilian juniper.
The modified plate method (Xu et al., Citation2011a, Citation2013b), following the traditional cellulose extraction procedure of the Jayme–Wise method (Green, Citation1963), was used to extract α-cellulose. Cellulose samples weighing 80–260 μg were then wrapped in silver foil. Tree ring cellulose δ18O values were measured using an isotope ratio mass spectrometer (Delta V Advantage; Thermo Scientific) interfaced with a pyrolysis-type, high-temperature conversion elemental analyzer (TC/EA; Thermo Scientific) at the Research Institute for Humanity and Nature, Japan. Cellulose δ18O values were calculated by comparison with analysis of Merck cellulose (laboratory working standard), which was analysed after every eight tree ring samples. Oxygen isotope results are presented in δ notation as the per mil (‰) deviation from Vienna Standard Mean Ocean Water (VSMOW): δ18O = [(Rsample/Rstandard) − 1] × 1000, where Rsample and Rstandard are the 18O/16O ratios of the sample and standard, respectively. The analytical uncertainties on repeated measurements of the Merck cellulose were approximately ±0.2‰ (n = 231).
2.3. Climatic and statistical analyses
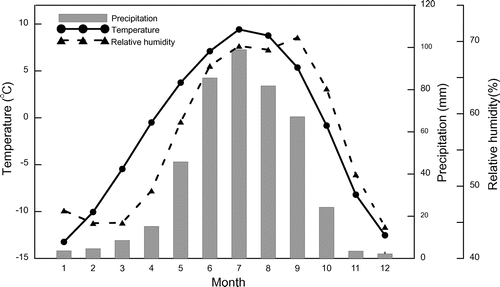
The sampling site is located in the transition zone of the Tibetan Plateau between the regions in the south dominated by the monsoon and those in the north dominated by westerly winds (Yao et al., Citation2013). Climatic parameters (monthly total precipitation, monthly mean temperature and monthly relative humidity) from eight meteorological stations (Xinghai, Maduo, Dari, Qingshuihe, Yushu, Qumalai, Shiqu and Tuotuohe, Table S1) during the period of 1960–2014 obtained from the China Meteorological Data Sharing Service System (http://data.cma.cn/) in the transition region (Tian et al., Citation2008; Yao et al., Citation2013) were averaged to represent the regional climate. The mean annual temperature of this region ranges from −2.5 to 0.2 °C, and the mean annual total precipitation ranges from 365 to 575 mm, with most precipitation falling from May to September (Fig. ). To investigate the relationship between tree ring δ18O and climatic factors, Pearson correlation coefficients were calculated between tree ring δ18O and these three climatic variables from January to October. Mean inter-trees correlation (Rbar) and Expressed Population Signal (EPS) were calculated to evaluate consistency and signal strength of δ18O time series from different trees (Wigley et al., Citation1984).
Fig. 2. Regional monthly mean temperature (black circles), precipitation (grey bars) and relative humidity (triangles) from 1960 to 2014.
3. Results and discussion
3.1. Juvenile effects on cellulose δ18O
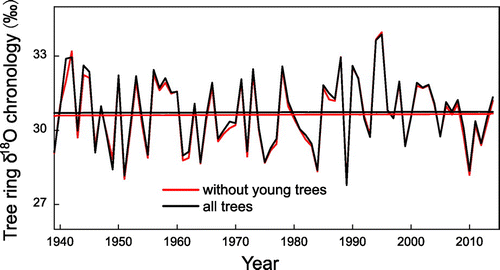
Tree ring δ18O time series from four young trees with pith are compared with δ18O values from old trees in Fig. . The tree ring δ18O values of YK173 (Fig. a) are ~3.95‰ and 1.24‰ lower than the δ18O values from the old trees when the young trees are two and three years old, respectively. The mean and standard deviation of δ18O values of YK173 and the old trees are very similar after the first four years of growth. δ18O values of YK184 (Fig. b) are ~2.68‰ lower than the δ18O values from old trees during the first nine years of growth and the δ18O values of YK180 (Fig. c) are ~2.03‰ lower than the δ18O values from the old trees during the first 10 years of growth. δ18O values of YK171 exhibit similar variations to the old trees (Fig. d). Figure reveals that the δ18O values of young trees are sometimes lower than those of old trees, but this phenomenon is limited to the first 10 years of growth. Despite this, δ18O variations from old and young trees are highly correlated when cambial ages are <10 years. δ18O values of young Qilian juniper with cambial ages of >10 years are very similar to those of old Qilian juniper. To test the feasibility of building up chronology using young trees (>10 years part), we build up two tree ring δ18O chronologies that include young trees (>10 years part) and exclude young trees. The mean values and long-term trends of these two tree ring δ18O chronologies have not significant difference (Fig. ), and these two tree ring δ18O chronologies are highly correlated, which indicates young trees (>10 years part) could be used for paleoclimate reconstruction as old trees.
Fig. 3. Tree ring δ18O values from young trees (red lines) and old trees (black lines). The dashed line indicates the pith year.
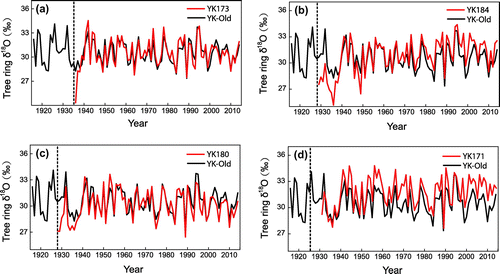
Fig. 4. Tree ring δ18O chronology based on all trees (black) and non-young trees (red).
Cellulose δ18O values of young oak trees in France increase in their first 30 years of growth (Labuhn et al., Citation2014), which is similar to our results but over a longer time span. In contrast, δ18O values of young juniper from central Asia have higher values than old juniper, and the age-related decreasing trends persist for several hundred years (Treydte et al., Citation2006). δ18O values of young larch trees from the Kamchatka Peninsula, Russia (Nakatsuka et al., Citation2008), and pine from Spain (Esper et al., Citation2010) also show long-term decreasing δ18O trends. However, age-related effects for pine from northern Fennoscandia are not observed after a juvenile phase of about 50 years (Young et al., Citation2011). There is also no apparent juvenile effect on δ18O values of larch in Bhutan (Sano et al., Citation2013), and no significant differences between mean values, standard deviations and climate responses of δ18O between young and old pine trees in south-east China (Xu et al., Citation2016). Juvenile effects on δ18O depend on species and location.
In this study, the lower δ18O values of young trees may be related to xylogenesis. Young trees with an earlier onset of xylogenesis may uptake some meltwater characterized by depleted δ18O (Azetsu-Scott and Tan, Citation1997), which results in lower xylem water δ18O and cellulose δ18O. As the trees grow older, the delayed onset of cambium division may minimize the influences from meltwater (Li et al., Citation2013). However, the detailed cause of the juvenile effect on tree ring δ18O values remains unclear, and further work is needed to identify the mechanisms involved. Nevertheless, if the first 10 years of tree growth are not included, then paleoclimate reconstruction using young trees is robust.
3.2. Intra-tree δ18O variability
Tree ring δ18O values at different sampling heights (15 and 70 cm) are shown in Fig. a. The mean and standard deviation of δ18O values at 15 cm (70 cm) are 32.64‰ (32.63‰) and 1.69‰ (1.59‰), respectively. Tree ring δ18O values at different heights show a high degree of synchronicity (r = 0.93; n = 43), and an analysis of variance (ANOVA) shows no significant differences in mean or standard deviation between tree ring δ18O values at different heights. Although previous studies have revealed vertical δ13C isotopic variations (Leavitt, Citation2010), our results show no vertical δ18O changes in Qilian juniper. In addition, recent study on two species (Abies georgei and Hippophae tibetana) in south-east Tibet Plateau revealed that the mean and standard deviation of tree ring oxygen isotope at different sampling heights do not have significant differences Xu et al. (Citation2017).
Fig. 5. Tree ring δ18O values from different sampling heights (a) and directions (b).
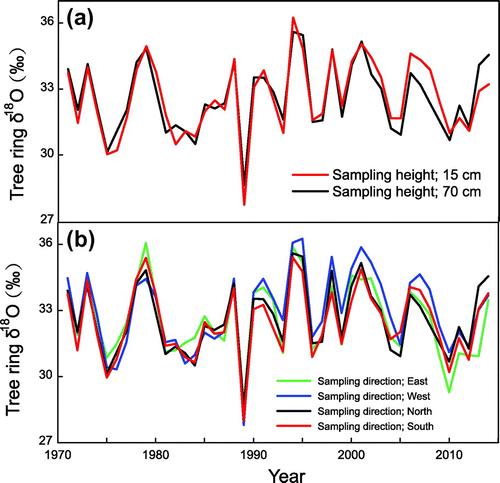
Tree ring δ18O variations in different directions (north, south, west and east) are shown in Fig. b. The mean δ18O values from the four directions are 32.63, 32.49, 32.98 and 32.60‰, respectively (Table ), and ANOVA results show no significant differences in mean or standard deviation between tree ring δ18O values in different directions. The yearly standard deviations obtained from the four directions vary between 0.12 and 1.33‰ (mean = 0.48‰), and these values are equivalent to the ranges reported for Abies pindrow radii (0.5–2‰) and Quercus petraea (0.5–1.5‰) (Ramesh et al., Citation1985; Robertson et al., Citation1995). The δ18O time series from the four directions are also positively correlated (Table ), and the mean inter-series correlation for the four directions is 0.92. These results show that the mean and standard deviation of vertical and circumferential δ18O time series are very similar, and tree ring δ18O values from different heights and directions are highly correlated, which indicates that we can reconstruct long-term δ18O records by combining samples from different heights and directions.
Table 1. Statistical analysis of data for each tree in this study.
Table 2. Correlation coefficients among tree ring δ18O time series from different directions.
3.3. Inter-tree δ18O variability
All tree ring δ18O time series in YK site are illustrated in Fig. a–e and Table . The lowest value for samples from the YK site is 28.82‰ (YK105) at 3900 m, and the highest value is 32.04‰ (YK173) from 3800 m (Fig. ). The largest difference amongst all the δ18O time series from the YK site is 3.22‰, which falls within the range of 1–4‰ for inter-tree δ18O variability (Leavitt, Citation2010). Given that the microclimatic conditions associated with these trees are similar, genetic variability and the crown/root architecture of individuals may contribute to some of the observed inter-tree variability (Leavitt, Citation2010).
Fig. 6. Tree ring δ18O time series from different altitudes. Coloured lines in Figure a–e indicate oxygen isotope time series from different trees in each altitude, and coloured lines in Figure f indicate tree ring oxygen isotope time series from different altitudes.
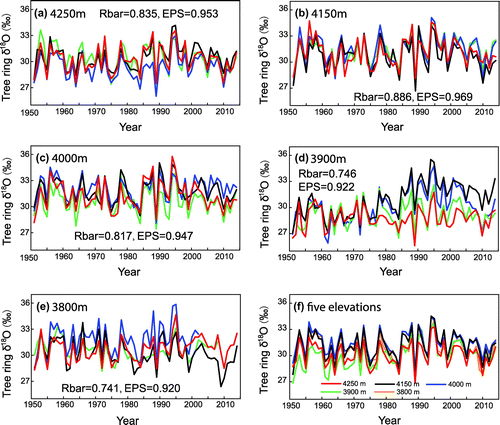
The first-order autocorrelation for all trees except YK109 and YK114 is lower than 0.2 (Table ). Unlike tree ring width, tree ring oxygen isotopes in the current year are not affected significantly by tree ring oxygen isotopes in the previous year (Hill et al., Citation1995). Tree ring δ18O records from individual trees in Laos, Vietnam, Bhutan, Thailand and south-eastern and northern China have lower first-order autocorrelations (<0.2) in monsoonal Asia (Sano et al., Citation2012b, Citation2013; Xu et al., Citation2013a, Citation2013b, Citation2015, Citation2016). On the Tibetan Plateau, the first-order autocorrelation of Qilian juniper δ18O records in the Qilian Mountains, as constructed by individual trees and pooling, is 0.29 (Qin et al., Citation2014) and 0.69 (Xu et al., Citation2011b), respectively.
The first-order autocorrelations of tree ring δ18O values for YK109 and YK114 are 0.53 and 0.39, respectively, which are much higher than other δ18O time series at this site. δ18O data for YK109 (Fig. d; black line) and YK114 (Fig. d; blue line) show significant increasing trends during the period of 1985–2014, but increasing trends were not found in other trees at the same altitude or at other altitudes at the YK site. The increasing trend may contribute to the observed high first-order autocorrelations for these two trees. The reasons for the increasing trends in YK109 and YK114 are not clear. Climatic conditions cannot explain the trends, because climate is expected to be the same for all trees at the same altitude, and δ18O values for YK105 (Fig. d; green line) and YK115 (Fig. d; red line) do not show significant trends. Tree age also cannot explain the increasing trends, because YK114 and YK109 are >200 years old. One possible explanation may be related to genetic variability or physiological factors. The δ18O values of YK114 and YK109 are higher (2–3‰) than the δ18O values of YK105 and YK115 during the period of 1985–2005. Based on a tree ring oxygen isotope fractionation model (Roden et al., Citation2000), enriched cellulose δ18O may be due to enhanced transpiration, enriched precipitation δ18O or a reduced exchange ratio with xylem water during cellulose formation. However, the underlying mechanisms that result in different trends for trees from the same site are unclear. Such different trends for the same species at the same site and time have also been observed for Japanese cedar from Yakushima Island, Japan (Dr. Sano, personal communication). Multi-stable isotope analysis, including carbon and hydrogen isotope data, might shed more light on the causes of these different trends.
In addition, δ18O values for YK109 and YK114 show significant positive correlations with those of the remaining 18 trees, although such trends were not observed in the other trees. It should be noted that such trends which are unrelated to climate are need to be identified and removed for the purposes of paleoclimate reconstructions. As such, measuring δ18O values from individual trees appears to be necessary for robust reconstructions.
3.4. Altitude effects on cellulose δ18O
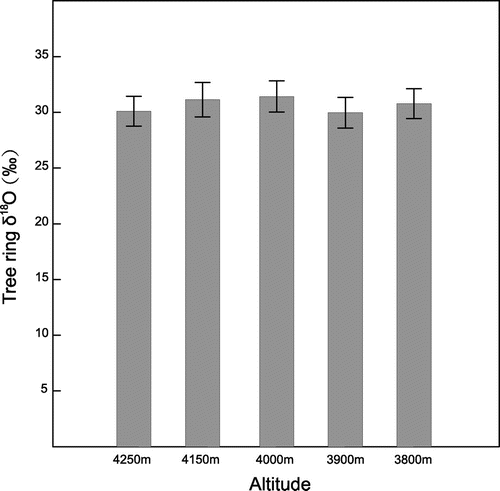
Averaged tree ring δ18O time series from five altitudes (3800, 3900, 4000, 4150, 4250 m) were shown in Fig. f and Table . ANOVA results indicate that there are no significant differences in standard deviation between tree ring δ18O values from five altitudes. The mean value of δ18O from five altitudes ranges from 29.98 to 31.42‰, with lowest value of 29.98‰ in 3900 m and highest value of 31.42‰ in 4000 m. There is no significant relationship between tree ring δ18O and altitude (Fig. ). Recent study showed that observed lapse rate is very small (−0.06‰/100 m) in the transition zone where are affected by shifting influences between the westerlies and Indian monsoon (Yao et al., Citation2013). The insignificant correlation between altitude and precipitation δ18O in study area may be the reason that there is no obvious altitude effect on tree ring δ18O. Previous study showed that there is 2.3‰ difference between earlywood δ18O of Smith fir at the high and low altitude in Sygera Mountains of the south-eastern Tibetan Plateau (Zeng et al., Citation2014), because observed lapse rate is strong (−0.13‰/100 m) in the monsoon domain (Yao et al., Citation2013).
Fig. 7. Average and standard deviation of tree ring δ18O data from different altitudes.
The Rbar and EPS of δ18O from each altitude is shown in Fig. . The Rbar of δ18O from each altitude is from 0.741 to 0.886, which is higher than Rbar of Qilian juniper δ18O (0.62) in Qilian mountain (Qin et al., Citation2014) and of Juniper (0.66) in central Asia (Treydte et al., Citation2006). EPS of tree ring δ18O from all altitudes ranges from 0.92 to 0.96, which is higher than 0.85 that is considered that the composite chronology represents the mean variance of the population and yields a signal relatively free of noise due to individual variation (Wigley et al., Citation1984), and is also higher than EPS of Qilian Juniper (0.88) in Qilian mountain (Qin et al., Citation2014). In addition, tree ring δ18O from different altitudes are highly correlated (r > 0.88, Table ). These results indicate that altitude effects for tree ring δ18O of Qilian Juniper in study area are not observed and samples from each altitude can represent the local signal. Therefore, tree ring δ18O from all altitudes were averaged to build up the regional δ18O chronology.
Table 3. Correlation coefficient among tree ring δ18O time series from different altitudes.
3.5. Climatic implications of tree ring δ18O data
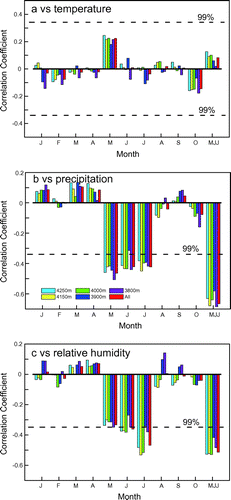
Previous studies have shown that tree ring δ18O values mainly reflect climate in the current growth season (Xu et al., Citation2011a; Barbour and Song, Citation2014; Wernicke et al., Citation2015). A correlation analysis was carried out between tree ring cellulose δ18O data from different altitude (light blue, yellow, green, dark blue and purple bars in Fig. )/all altitudes (red bar in Fig. ) and regional climatic parameters in the transition zone for the period 1960–2014. Given that the tree ring δ18O data from each altitude show similar climatic responses (Fig. ), the climatic implications are considered to be regional in nature. There are no significant correlations between tree ring δ18O and temperature (Fig. a). The δ18O data are significantly correlated with May–July precipitation (r = −0.67; n = 55; p < 0.001), and the correlation coefficients between precipitation and δ18O are similar for the months of May, June and July (Fig. b). The regional δ18O values show a negative correlation with May–July humidity (r = −0.51; n = 55; p < 0.001), where the main signal originates from June and July (Fig. c). The significant negative correlation between tree ring δ18O and relative humidity is readily explained by the tree ring oxygen isotope fractionation model (Roden et al., Citation2000). Lower relative humidity results in enhanced evapotranspiration, which causes the leaf water δ18O to be enriched and leads to the evaporation of soil water. Therefore, enriched leaf water and xylem water δ18O cause the higher cellulose δ18O.
Fig. 8. Correlations between tree ring δ18O from different altitudes and regional temperature (a), precipitation (b) and relative humidity (c) obtained from eight instrumental stations during 1960–2014.
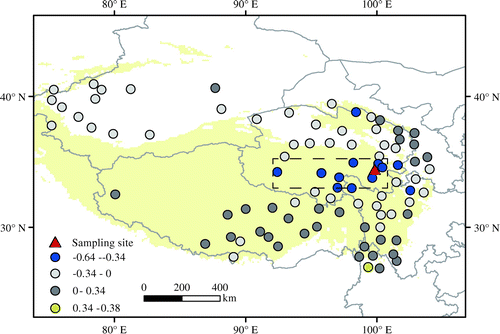
Positive correlations between relative humidity and precipitation (r = 0.44; n = 55; p < 0.01) indicate that precipitation can affect tree ring δ18O by influencing relative humidity. The correlation between precipitation and tree ring δ18O is higher than that between relative humidity and tree ring δ18O. Thus, there are other processes that precipitation influence tree ring δ18O. Spatial correlations between tree ring δ18O and precipitation (Fig. ) show that tree ring δ18O has a significant negative correlation with precipitation near the study area and west of the sampling location, which may reflect the rainout effect during water vapour transportation (Rozanski et al., Citation1992). Previous studies have also indicated that precipitation δ18O is linked to rainout processes in surrounding regions (Lawrence et al., Citation2004; Kurita et al., Citation2009). Greater rainfall in surrounding or upstream areas associated with rainout of heavy isotopes results in more depleted precipitation δ18O and tree ring δ18O. This negative correlation between tree ring δ18O and regional precipitation has also been observed in several sites elsewhere in Asia (Sano et al., Citation2013; Xu et al., Citation2013b, Citation2015, Citation2016).
Fig. 9. Spatial correlations between tree ring δ18O and May–July precipitation from meteorological stations on the Tibetan Plateau from 1960 to 2014.
4. Conclusion and perspectives
In this study, we analysed cellulose δ18O in 21 trees of Qilian juniper from the Animaqing Mountains to investigate potential juvenile and altitude effects, intra- and inter-tree δ18O variability and climatic implications of cellulose δ18O. The δ18O values of young trees are sometimes lower than the δ18O values of old trees, but this effect is limited to the first 10 years of growth. δ18O values of young Qilian juniper with cambial ages >10 years are very similar to those of old Qilian juniper. Excluding the first 10 years of growth allows paleoclimate reconstructions from young trees. ANOVA results show no significant differences in the mean and standard deviation of tree ring δ18O values at different heights and directions, and tree ring δ18O time series from different heights and directions are highly correlated.
The mean inter-series correlation (Rbar) and EPS for tree ring δ18O data from each altitude are very high, and tree ring δ18O data from five altitudes are highly correlated and share a similar climatic response. Altitude effects on tree ring δ18O values are not observed in the study area. These findings indicate that reconstructing long-term δ18O records using samples from one site regardless of sampling height, direction and altitude is a robust approach. Given that the tree ring width of Qilian juniper is sometimes narrow, using data from different trees/cores with relatively wide rings is a viable way of building a composite δ18O record. Such an approach may also be applicable to juniper from the northern and southern Tibetan Plateau. Tree ring δ18O data from the Animaqing Mountains exhibit a significant negative correlation (r = −0.67; p < 0.001) with May–July regional precipitation. Future studies should be able to use tree ring δ18O data as a proxy of paleo-precipitation and tree ring width as a proxy of paleo-temperature in the Animaqing Mountains.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was jointly funded by the National Natural Science Foundation of China [grant number 41630529], [grant number 41672179], and [grant number 41690114]; the Chinese Academy of Sciences (CAS) Pioneer Hundred Talents Program; the National Key R&D Program of China [grant number 2016YFA0600502]; and a Grant-in-Aid for Scientific Research [grant number 2310262] from JSPS, Japan.
Supplemental data
Supplemental data for this article can be accessed here https://doi.org/10.1080/16000889.2017.1391663.
Table S1. Details of meteorological stations.
Download MS Word (52.7 KB)Acknowledgements
We deeply appreciate the helpful comments from two anonymous reviewers and the editor to improve the manuscript.
References
- An, W., Liu, X., Leavitt, S. W., Xu, G., Zeng, X. and co-authors. 2014. Relative humidity history on the Batang-Litang Plateau of western China since 1755 reconstructed from tree-ring δ18O and δD. Clim. Dyn. 42, 2639–2654.10.1007/s00382-013-1937-z
- Azetsu-Scott, K. and Tan, F. C. 1997. Oxygen isotope studies from Iceland to an East Greenland Fjord: behaviour of glacial meltwater plume. Marine Chem. 56, 239–251.10.1016/S0304-4203(96)00078-3
- Barbour, M. and Song, X. 2014. Do tree-ring stable isotope compositions faithfully record tree carbon/water dynamics? Tree Physiol. 34, 792–795.10.1093/treephys/tpu064
- Cook, E. R., Anchukaitis, K. J., Buckley, B. M., D’Arrigo, R. D., Jacoby, G. C. and co-authors. 2010. Asian monsoon failure and megadrought during the last millennium. Science. 328, 486–489.
- Chen, F., Zhang, Y., Shao, X., Li, M. Q. and Yin, Z. Y. 2016. A 2000-year temperature reconstruction in the Animaqin Mountains of the Tibet Plateau, China. Holocene 26(12), 1904–1913.10.1177/0959683616646187
- Esper, J., Frank, D., Battipaglia, G., Büntgen, U., Holert, C. and co-authors. 2010. Low-frequency noise in δ13C and δ18O tree ring data: a case study of Pinus uncinata in the Spanish Pyrenees. Global Biogeochem. Cycles 24, GB4018. DOI: 10.1029/2010GB003772.
- Gagen, M., McCarroll, D., Edouard, J. L. 2006. Combining ring width, density and stable carbon isotope proxies to enhance the climate signal in tree-rings: an example from the Southern French Alps. Clim. Change 78, 363–379(317).10.1007/s10584-006-9097-3
- Gagen, M., McCarroll, D., Loader, N. J. and Robertson, I. 2011. Stable isotopes in dendroclimatology: moving beyond ‘potential’. In Dendroclimatology. Developments in Paleoenvironmental Research (eds. M. Hughes, T. Swetnam and H. Diaz H) Vol. 11 . Springer, Dordrecht.
- Gou, X., Deng, Y., Chen, F., Yang, M., Fang, K. and co-authors. 2010. Tree ring based streamflow reconstruction for the Upper Yellow River over the past 1234 years. Chin. Sci. Bull. 55, 4179–4186.10.1007/s11434-010-4215-z
- Green, J. 1963. Wood cellulose. In: Method in Carbohydrate Chemistry (ed. R. L. Whistler). Academic, San Diego, CA, pp. 9–21.
- Grießinger, J., Bräuning, A., Helle, G., Thomas, A. and Schleser, G. 2011. Late Holocene Asian summer monsoon variability reflected by δ18O in tree-rings from Tibetan junipers. Geophys. Res. Lett. 38, L03701.
- Grießinger, J., Bräuning, A., Helle, G., Hochreuther, P. and Schleser, G. 2017. Late Holocene relative humidity history on the southeastern Tibetan plateau inferred from a tree-ring δ18O record: recent decrease and conditions during the last 1500 years. Quat. Int. 430, 52–59.10.1016/j.quaint.2016.02.011
- Hill, S., Waterhouse, J., Field, E., Switsur, V. and Ap Rees, T. 1995. Rapid recycling of triose phosphates in oak stem tissue. Plant, Cell Environ. 18, 931–936.10.1111/pce.1995.18.issue-8
- Holmes, R. 1983. Computer-assisted quality control in tree-ring dating and measurement. Tree-ring Bull. 43, 69–78.
- Kurita, N., Ichiyanagi, K., Matsumoto, J., Yamanaka, M. D. and Ohata, T. 2009. The relationship between the isotopic content of precipitation and the precipitation amount in tropical regions. J. Geochem. Explor. 102, 113–122.10.1016/j.gexplo.2009.03.002
- Labuhn, I., Daux, V., Pierre, M., Stievenard, M., Girardclos, O. and co-authors. 2014. Tree age, site and climate controls on tree ring cellulose δ18O: a case study on oak trees from south-western France. Dendrochronologia 32, 78–89.10.1016/j.dendro.2013.11.001
- Lawrence, J. R., Gedzelman, S. D., Dexheimer, D., Cho, H. K., Carrie, G. D. and co-authors. 2004. Stable isotopic composition of water vapor in the tropics. J. Geophys. Res. 109, 539–547.
- Leavitt, S. W. 2010. Tree-ring C–H–O isotope variability and sampling. Sci. Total Environ. 408, 5244–5253.10.1016/j.scitotenv.2010.07.057
- Li, X., Liang, E., Gricar, J., Prislan, P., Rossi, S. and co-authors. 2013. Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol. 33, 48–56.10.1093/treephys/tps113
- Liu, X., Zeng, X., Leavitt, S. W., Wang, W., An, W. and co-authors. 2013. A 400-year tree-ring δ18O chronology for the southeastern Tibetan Plateau: implications for inferring variations of the regional hydroclimate. Global Planet Change 104, 23–33.10.1016/j.gloplacha.2013.02.005
- Liu, X., Xu, G., Grießinger, J., An, W., Wang, W. and co-authors. 2014. A shift in cloud cover over the southeastern Tibetan Plateau since 1600: evidence from regional tree-ring δ18O and its linkages to tropical oceans. Quat. Sci. Rev. 88, 55–68.10.1016/j.quascirev.2014.01.009
- Mccarroll, D. 2015. ‘Study the past, if you would divine the future’: a retrospective on measuring and understanding Quaternary climate change. J. Quat. Sci. 30, 154–187.10.1002/jqs.2775
- Nakatsuka, T., Ohnishi, K. and Hara, T. 2008. Reconstruction of summer temperature variation by hydrogen and oxygen isotopic ratios of larch tree-ring cellulose in Kamchatka Peninsula (in Japanese). Mon. Chikyu 30(4), 207–215.
- Qin, C., Bao, Y., Bräuning, A., Grießinger, J. and Wernicke, J. 2014. Drought signals in tree-ring stable oxygen isotope series of Qilian juniper from the arid northeastern Tibetan Plateau. Global Planet. Change 125, 48–59.
- Ramesh, R., Bhattacharya, S. K. and Gopalan, K. 1985. Dendroclimatological implications of isotope coherence in trees from Kashmir Valley, India. Nature 317, 802–804.10.1038/317802a0
- Robertson, I., Field, E. M., Heaton, T. H. E., Pilcher, J. R., Pollard, M. and co-authors. 1995. Isotope coherence in oak cellulose. In: Problems of Stable Isotopes in Tree-rings, Lake Sediments and Peat-bogs as Climatic Evidence for the Holocene (eds. B. Frenzel, B. Stauffer and M. M. Weib). G. Fischer, Stuttgart, Vienna and New York, pp. 141–155.
- Roden, J. S., Lin, G. and Ehleringer, J. R. 2000. A mechanistic model for interpretation of hydrogen and oxygen isotope ratios in tree-ring cellulose. Geochim. Cosmochim. Acta 64, 21–35.10.1016/S0016-7037(99)00195-7
- Rozanski, A. A., Araguás-Araguás, L. and Gonfiantini, R. 1992. Isotopic patterns in global precipitation. Washington DC Am. Geophys. Union Geophys. Monograph 78, 1–36.
- Sano, M., Ramesh, R., Sheshshayee, M. and Sukumar, R. 2012a. Increasing aridity over the past 223 years in the Nepal Himalaya inferred from a tree-ring δ18O chronology. Holocene 22(7), 809–817.10.1177/0959683611430338
- Sano, M., Xu, C. and Nakatsuka, T. 2012b. A 300-year Vietnam hydroclimate and ENSO variability record reconstructed from tree ring δ18O. J. Geophys. Res. 117, D12115.
- Sano, M., Tshering, P., Komori, J., Fujita, K., Xu, C. and co-authors. 2013. May–September precipitation in the Bhutan Himalaya since 1743 as reconstructed from tree ring cellulose δ18O. J. Geophys. Res.: Atmos. 118, 8399–8410.
- Savva, Y., Oleksyn, J., Reich, P. B., Tjoelker, M. G., Vaganov, E. A. and co-authors. 2006. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees 20, 735–746.10.1007/s00468-006-0088-9
- Shao, X., Xu, Y., Yin, Z. Y., Liang, E., Zhu, H. and co-authors. 2010. Climatic implications of a 3585-year tree-ring width chronology from the northeastern Qinghai-Tibetan Plateau. Quaternary Science Reviews 29, 2111–2122.10.1016/j.quascirev.2010.05.005
- Shi, C., Masson-Delmotte, V., Risi, C., Eglin, T., Stievenard, M. and co-authors. 2011. Sampling strategy and climatic implications of tree-ring stable isotopes on the southeast Tibetan Plateau. Earth and Planetary Science Letters 301(1–2), 307–316.10.1016/j.epsl.2010.11.014
- Stocker, T., Qin, D., Plattner, G., Tignor, M., Allen, S. and co-authors. 2013. IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK and New York, USA.
- Takahashi, K. and Yasue, H. A. 2003. Effects of climate on the radial growth of tree species in the upper and lower distribution limits of an altitudinal ecotone on Mount Norikura, central Japan. Ecol. Res. 18, 4–6.
- Tian, L. D., Ma, L. L., Yu, W. S., Liu, Z. F., Yin, C. L. and co-authors. 2008. Seasonal variations of stable isotope in precipitation and moisture transport at Yushu, eastern Tibetan Plateau. Sci. China Earth Sci. 51, 1121–1128.10.1007/s11430-008-0089-1
- Treydte, K., Schleser, G., Helle, G., Frank, D., Winiger, M. and co-authors. 2006. The twentieth century was the wettest period in northern Pakistan over the past millennium. Nature 440, 1179–1182.10.1038/nature04743
- Wang, W., Liu, X., Xu, G., Shao, X., Qin, D. and co-authors. 2013. Moisture variations over the past millennium characterized by Qaidam Basin tree-ring δ18O. Chin. Sci. Bull. 58, 3956–3961.10.1007/s11434-013-5913-0
- Wernicke, J., Grießinger, J., Hochreuther, P. and Bräuning, A. 2015. Variability of summer humidity during the past 800 years on the eastern Tibetan Plateau inferred from δ18O of tree-ring cellulose. Clim. Past 11, 327–337.10.5194/cp-11-327-2015
- Wigley, T., Briffa, K. and Jones, P. 1984. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 23, 201–213.10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2
- Xu, C., Sano, M. and Nakatsuka, T. 2011a. Tree ring cellulose δ18O of Fokienia hodginsii in northern Laos: a promising proxy to reconstruct ENSO? J. Geophysical Research 116, D24109.
- Xu, G., Chen, T., Liu, X., An, W., Wang, W. and co-authors. 2011b. Potential linkages between the moisture variability in the northeastern Qaidam Basin, China, since 1800 and the East Asian summer monsoon as reflected by tree ring δ18O. J. Geophys. Res. Atmos. 116, D09111.
- Xu, C., Sano, M. and Nakatsuka, T. 2013a. A 400-year record of hydroclimate variability and local ENSO history in northern Southeast Asia inferred from tree-ring δ18O. Palaeogeogr. Palaeoclimatol. Palaeoecol. 386, 588–598.10.1016/j.palaeo.2013.06.025
- Xu, C., Zheng, H., Nakatsuka, T. and Sano, M. 2013b. Oxygen isotope signatures preserved in tree ring cellulose as a proxy for April–September precipitation in Fujian, the subtropical region of southeast China. J. Geophys. Res. 118, 12805–12815.
- Xu, C., Pumijumnong, N., Nakatsuka, T., Sano, M. and Li, Z. 2015. A tree-ring cellulose δ18O-based July–October precipitation reconstruction since AD 1828, northwest Thailand. J. Hydrol. 529, 433–441.10.1016/j.jhydrol.2015.02.037
- Xu, C., Ge, J., Nakatsuka, T., Yi, L., Zheng, H. and co-authors. 2016. Potential utility of tree ring δ18O series for reconstructing precipitation records from the lower reaches of the Yangtze River, southeast China. J. Geophys. Res.: Atmos. 121, 3954–3968.
- Xu, C., Zhu, H., Nakatsuka, T., Sano, M., Li, Z. and co-authors. In press. Sampling strategy and climatic implication of tree-ring cellulose oxygen isotopes of Hippophae tibetana, and Abies georgei, on the southeastern Tibetan Plateau. Int. J. Biometeorol. DOI: 10.1007/s00484-017-1365-6.
- Yang, B., Qin, C., Wang, J., He, M., Melvin, T. M., and co-authors. 2014. A 3,500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. Proc. Nat. Acad. Sci. United States Am. 111, 2903–2908.10.1073/pnas.1319238111
- Yao, T., Masson-Delmotte, V., Gao, J., Yu, W., Yang, X., and co-authors. 2013. A review of climatic controls on δ18O in precipitation over the Tibetan Plateau: observations and simulations. Rev. Geophys. 51, 525–548.10.1002/rog.v51.4
- Young, G. H. F., Demmler, J. C., Gunnarson, B. E., Kirchhefer, A. J., Loader, N. J. and co-authors. 2011. Age trends in tree ring growth and isotopic archives: a case study of Pinus sylvestris L. from northwestern Norway. Global Biogeochem. Cycles 25, GB2020.
- Zeng, X., Liu, X., Wang, W., Xu, G., An, W. and co-authors. 2014. No altitude-dependent effects of climatic signals are recorded in Smith fir tree-ring δ18O on the southeastern Tibetan Plateau, despite a shift in tree growth. Boreas 43, 588–599.10.1111/bor.2014.43.issue-3
- Zhang, H., Shao, X. and Zhang, Y. 2015. Which climatic factors limit radial growth of Qilian juniper at the upper treeline on the northeastern Tibetan Plateau? J. Geog. Sci. 25, 1173–1182.10.1007/s11442-015-1226-3