


ABSTRACT
Beliefs about the general character of the world – called primals – have received much attention in recent years. Primals (e.g. believing that the world is safe) are associated with psychological and social outcomes, such as personality traits, character strengths and life satisfaction. We performed the first study of genetic and environmental influences on primals, and explored phenotypic associations between primals and the big five personality traits, in a large (~9000) population-wide Swedish twin sample. Correlations between primals and personality traits were low to moderate. Broad-sense heritability estimates of primals ranged between 21% and 49%, suggesting that genetic factors contribute significantly to people’s beliefs about the general character of the world. In contrast, we found little support for shared environmental effects (e.g. rearing environment). Conclusively, we demonstrate that primals and personality are related, yet distinct, constructs and that individual differences in primals depend on both genetic and non-shared environmental factors.
Introduction
How internal beliefs can impact perceptions and, in turn, influence mental (and physical) well-being, is a core question in positive psychology and cognitive behavioral therapy (Beck, Citation2020; Karwoski et al., Citation2006; Seligman & Csikszentmihalyi, Citation2000). Yet, while past research has focused greatly on beliefs about the self, research regarding beliefs about the general character of the world is relatively limited (Chen et al., Citation2016), and has primarily focused on whether individuals believe the world is benevolent, meaningful (Janoff-Bulman, Citation1989) and safe or just (Clifton et al., Citation2019). In 2019, Clifton and colleagues set out to, for the first time, create a comprehensive questionnaire that would cover a broader range of world beliefs, apart from those mentioned above: so-called primal world beliefs – or primals, for short. The resulting Primals Inventory (PI-99) consists of 99 items. Psychometric analyses of US samples suggest that primals can be adequately modelled by a partly hierarchical structure with a large number (22) of tertiary primals at the lowest level. Most of these tertiary primals load onto three overarching secondary primals, namely, Safe, Enticing and Alive; with a single primary Good primal at the apex of the hierarchy (see Clifton et al., Citation2019). For more reasonable assessment times, Clifton and Yaden (Citation2021) shortened the original PI-99 into validated 18-Item and 6-Item versions (PI-18 and PI-6, respectively) to measure higher-order primals.
Besides creating questionnaires that allow for standardized empirical assessments of primal world beliefs, Clifton et al. (Citation2019) investigated the relationship of primals with various other psychological and social outcomes. Of notice, they found that more positive world beliefs were associated with higher levels of well-being and better mental health, i.e. less depression, anxiety and stress (Clifton et al., Citation2019). Furthermore, most primals were associated with the big five personality traits (i.e. Openness to experience, Conscientiousness, Extraversion, Agreeableness and Neuroticism, McCrae & John, Citation1992; for the relationship with primals see Clifton et al., Citation2019; Stahlmann et al., Citation2020), and with certain character strengths (e.g. hope, and gratitude; McGrath & Rich, Citation2023; Stahlmann & Ruch, Citation2023). This aligns with past research, finding that, for example, Extraversion and Agreeableness are positively, and Neuroticism is negatively, associated with the belief that the world is just (see meta-analysis by Nudelman, Citation2013), while benevolent world beliefs are positively related to Agreeableness (Roccas et al., Citation2002). Additionally, believing that the world is dangerous is positively associated with Neuroticism and negatively with Openness (Dallago et al., Citation2012; Langston & Sykes, Citation1997; Van Hiel et al., Citation2007). Similarly, higher well-being, less mental health problems and more life satisfaction have been associated with most primal world beliefs (Clifton & Kim, Citation2020; Clifton & Yaden, Citation2021; Clifton et al., Citation2019; Stahlmann et al., Citation2020), including the belief that the world is benevolent (Poulin & Cohen Silver, Citation2008), safe (Syropoulos, Citation2020) and just (Dzuka & Dalbert, Citation2006). Analyzing stress-related symptoms in response to the COVID-19 pandemic, additionally, revealed that believing in the goodness of the world (i.e. Good primal, assessed via PI-6; Clifton & Yaden, Citation2021) was associated with post-traumatic growth (Vazquez et al., Citation2021). However, primals did not significantly change with regard to stress/averse events during COVID-19, but remained relatively stable (Ludwig et al., Citation2023), which aligns with past research reporting high test–retest reliability of primals (Clifton et al., Citation2019). Contrary to popular beliefs, recent data indicate that primals may be relatively independent of socio-economic variables (Kerry et al., Citation2023), e.g. with people living in higher crime rate neighborhoods not reporting the world to be less safe. On the basis of these results, Clifton and colleagues have proposed that primals are a construct with similar stability to personality (see, e.g. https://myprimals.com/).
While primals seem to be related to many traits of interest to the field of psychology and have rapidly gained attention from both the research society and the public over the past few years, to-date, little is known about the genetic contributions to individual variation in primals and their associated traits, despite observed associations commonly being interpreted as causal (ignoring potential familial confounding). However, most psychological traits are substantially heritable, and associations between traits are known to be heavily confounded by familial factors (e.g. Maes et al., Citation2023; Mosing et al., Citation2014; Polderman et al., Citation2015; Schur et al., Citation2013). For instance, a meta-analysis of more than 2,500 twin studies by Polderman et al. (Citation2015) suggests that complex human traits are on average around 50% heritable (e.g. psychiatric 46.3% and social values 29.0%). In particular, personality traits show a mean heritability of around 40% (Vukasović & Bratko, Citation2015). A recent study by Vellani et al. (Citation2022), furthermore, found that individual differences in belief formation (e.g. updates of prior beliefs in light of actual probabilities for specific events) are 13% heritable. Against this background, we would expect primals to also be partly influenced by genetic factors. Here, we utilize a large, extensively phenotyped sample of approximately 9000 Swedish twins to, for the first time, explore the genetic and environmental nature of primal world beliefs. Specifically, we aimed to: (1) create a Swedish version of the Primals Inventory, including the full PI-18 and 11 additionally selected tertiary primal world beliefs, based on the PI-99, in a large Swedish population sample, (2) examine the relationship between primals and the big five personality traits by estimating their phenotypic correlations, (3) investigate the genetic and environmental influences on primal world beliefs.
Materials & methods
Participants
A Swedish version of the PI-18 and 11 selected tertiary primals from the PI-99 (62-items in total – for details on the scale translation see Measures and Materials section and Supplementary material) were administered in the latest wave of data collection – the ‘Humans-Making Music Study 2’ (HUMMUS 2) – of the Study of Twin Adults: Genes and Environment (STAGE) cohort from the Swedish Twin Registry (Lichtenstein et al., Citation2002, Citation2006; Magnusson et al., Citation2013; Zagai et al., Citation2019). The overall purpose of HUMMUS-2 was to investigate expertise and creativity, as well as relationships between cultural engagement, health and well-being. The total participating sample consisted of 9,538 Swedish twins. The final sample (excluding individuals with unspecified zygosity, N = 49; 0.5% exclusion rate) contained 4,651 twin singletons and 2,419 complete twin pairs (684 MZ females, 413 MZ males, 405 DZ females, 287 DZ males and 630 DZ opposite-sexed twin pairs). Singletons were included in the analyses as they contribute information to the estimation of means, variances, and covariances. For a more detailed description of the cohort, see Lichtenstein et al. (Citation2002, Citation2006). In total, the ages of the participants ranged from 36 to 63 years (Mage = 50.29, SDage = 7.75), with a slight bias of more biologically sexed females participating in the present study (43.39% males).
Before the survey was conducted, participants provided informed consent. The study was approved by the Swedish Ethical Review Authority (Dnr 2021–02014, 2022–00109–02, 2020–02575), and conformed to the Declaration of Helsinki.
Measures and materials
Primal world beliefs
The original PI-18 (Clifton & Yaden, Citation2021) was forth- and back-translated between Swedish and English in consultation with the original authors of the PI-99, following recently published translation guidelines (Clifton et al., Citation2023). For a more comprehensive explanation of the procedure, see Supplementary Material 1. In a similar approach and to keep assessment times in the twin cohort reasonable, 11 tertiary primals of interest were included (i.e. 44 additional items; leading to 62 items being assessed in our Swedish Primals Inventory, in total).
For each item, participants were asked to indicate how strongly they agreed or disagreed with the selected primal world belief item on a 6-point Likert scale ranging from 6 (‘strongly agree’) to 1 (‘strongly disagree’), with higher scores representing stronger agreement. To derive the PI-18 (i.e. Good, Safe, Enticing and Alive) as well as the 11 tertiary primals, average scores across the respective items were calculated, for each participant. Cronbach's alpha values for the PI-18 and the 11 tested tertiary primals are shown in Supplementary Table S1 and Table S2, respectively.
Personality
The Swedish translation of the 30-item Big Five Personality Trait inventories (BFI-2, Soto & John, Citation2017; see also https://situationslab.squarespace.com/big-five-inventory-2) was used to assess Openness, Conscientiousness, Extraversion, Agreeableness and Neuroticism (see Supplementary Table S3). Items were scored on a 5-point Likert scale, ranging from 0 (‘disagree strongly’) to 4 (‘agree strongly’). Following the original guidelines, average scores were calculated for each participant. Equal to the primal world belief assessment, higher scores indicated stronger agreement.
Statistical analysis
The present study and analyses were preregistered (see https://aspredicted.org/pd323.pdf). Data were analyzed using R (version 4.3.1; R Core Team, Citation2023). For the confirmatory factor analysis, we used the lavaan R package (version 0.6–15; Rosseel, Citation2012) and for the exploratory factor analysis the psych R package with an oblimin rotation (version 2.3.3; Revelle, Citation2023). To assess heritabilities, the OpenMx package (version 2.20.6; Boker et al., Citation2023) with the NPSOL optimizer was used. Plots were created using the corrplot (version 0.92; Wei & Simko, Citation2021) and ggplot2 package (version 3.4.2; Wickham, Citation2016). The data were reorganized using tidyverse (version 2.0.0; Wickham et al., Citation2019).
Data structure and exploration
We confirmed the original PI-18 structure in our Swedish sample running a confirmatory factor analysis, following the structure proposed by Clifton and Yaden (Citation2021). To investigate if we find a similar structure despite measuring fewer primals, we explored if a similar higher-order structure (i.e. secondary primals) emerged from our selected tertiary primals utilizing an exploratory factor analysis (a similar approach can be observed in Stahlmann et al., Citation2020; though we use only a selection of primals opposed to all 22 tertiary primals). Results of this factor analysis and heritabilities based on the resulting factors can be found in the supplements (Material 2, Tables and .excel sheet).
Phenotypic correlations
In addition, the phenotypic relationships between assessed primals and personality were assessed using simple Pearson correlations. Relatedness was accounted for by assuring that for complete twin pairs only one randomly picked twin was included in the analyses.
Classical twin modelling
Classical twin models can be utilized to partition phenotypic variance into genetic (heritability) and environmental influences on a given trait (e.g. Posthuma et al., Citation2003; Rijsdijk & Sham, Citation2002). For this, first, basic twin model assumptions are tested by comparing a fully saturated model to more parsimonious submodels, using a stringent significance level of .01, while allowing for certain constrains (e.g. to test if means and variances within twins of a pair, between monozygotic and dizygotic twin pairs, and across the sexes can be equated; for a more detailed description see Posthuma et al., Citation2003). Univariate twin models estimate what proportion of the phenotypic variance is explained by additive genetic (A), non-additive/dominant genetic (D), common/shared environmental (C) and non-shared environmental (E) effects, using structure equation models in which these variance components correspond to latent variables (see ). The C component represents the environmental influences that are shared between twins of a pair and makes them more similar to one another (e.g. their family, and the neighborhood they grew up in). The non-shared environment (E) captures unique environmental influences which make twins of the pair different, as well as measurement error and idiosyncratic effects. Monozygotic (MZ) twins share, essentially, all genes. Dizygotic (DZ) twins on average share 50% of their segregating genes. Accordingly, for additive genetic effects DZ twins will show an average correlation of 0.50, whereas for dominance effects the expected correlation is 0.25 (for further explanation see Posthuma et al., Citation2003; and for a visualization see ). The difference in genetic similarity between MZ and DZ twins is the basis for heritability estimations. Specifically, if MZ twins are more similar on average than DZ twins, this indicates genetic influences on the investigated phenotype.
Figure 1. Heritability monozygotic (MZ) versus dizygotic (DZ) twins.
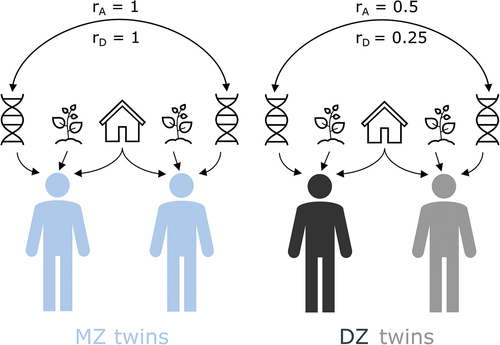
Notably, as classical twin designs do not include sufficient information to estimate C and D separately, either an ADE or ACE model is fitted. Specifically, if the DZ twin correlation is smaller than or equal to half the MZ twin correlation, dominant genetic effects are indicated and an ADE model is chosen; otherwise, an ACE model gets estimated. After fitting the full ACE/ADE model, it is assessed whether this full model or more parsimonious AE, CE, or E models fit the data best. For this, the D or C component gets equated to zero. To clarify, no DE models are fitted, as it is implausible to find non-additive genetic effects without additive genetic effects for complex human traits (Hill et al., Citation2008).
Transparency and openness
As we treated genetically informative data, which is highly sensitive data, data and materials are only available through application (see application procedure: https://ki.se/en/research/swedish-twin-registry-for-researchers). All analysis codes, however, are open-access and publicly available at https://osf.io/bd543/
Results
Data structure and exploration
Cronbach’s alphas for the PI-18 and tertiary primals ranged from .54 to .80 (details as well as means, and standard divisions are shown in Supplementary Table S1 and Table S2, respectively). The confirmatory factor analysis of the PI-18 structure revealed a CFI = 86.3, TLI = 81.6, SRMR = .06 and RMSEA (CI 90%) = .076 (.075; .078). Results of the hierarchical factor structure exploration based on the 11 tertiary primals can be found in the supplements (Supplementary Material 2).
Phenotypic correlations between primals and personality traits
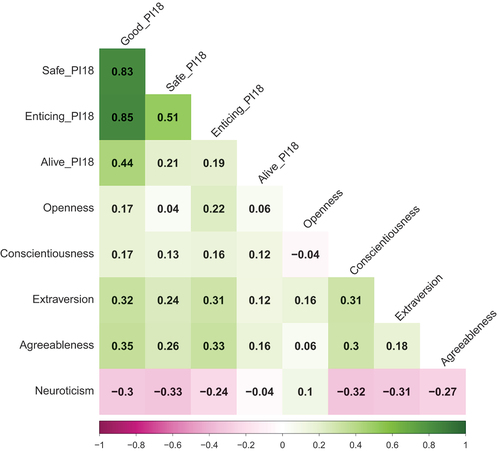
Phenotypic correlations between the big five personality traits were assessed for the PI-18 (see ). Here, Openness correlated most positively with Enticing (r = .22). Conscientiousness, Extraversion and Agreeableness all correlated most positively with the overarching Good primal (r = .17, r = .32, r = .35, respectively), while Neuroticism correlated most negatively with Safe (r =-.33).
Figure 2. Phenotypic correlation: PI-18 primals and personality traits.
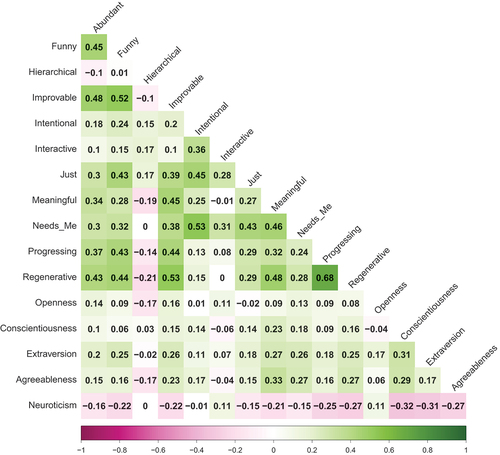
Tertiary primals showed low to moderate correlations with personality (see ). Notably, Openness correlated most positively with Improvable (r = .16). Conscientiousness, Extraversion and Agreeableness correlated most positively with Meaningful (r = .23, r = .27, r = .33, respectively). In contrast, Neuroticism was mostly negatively associated with tertiary primals, most strongly so with Regenerative (r = −.27).
Figure 3. Phenotypic correlation: tertiary primals and personality traits.
Variance explained by genetic influences
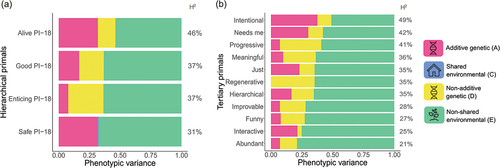
Assumption testing showed that all assumptions were met (see Supplementary.excel sheet). As there were no sex differences for any primal, simple ACE/ADE models were fit. In addition, for some primals age had a small but significant effect, thus, age was treated as a covariate for those primals (for more details see Supplementary.excel sheet). For all primals, apart from the PI-18 Safe primal (where an ACE model fits the data best), ADE models showed the best fit. However, D influences could, in most cases, be removed from the model without a significant deterioration of model fit (see Supplementary Table S4 and S5), resulting in AE models fitting most primals data best. Heritabilities of the PI-18 and tertiary primals were significant and similar. Twin correlations are shown in Supplementary Table S4 and Table S5, while the broad-sense heritability estimates, ranging from 21% to 49%, are visualized in (heritability estimates can be observed more detailed in Supplementary Figure S4). The remaining variance was explained by the non-shared environment (including measurement error and idiosyncratic effects) and, additionally, for Progressive and Regenerative due to non-additive dominance genetic effects. For further details on model fits and heritability estimates see Supplementary Table S4 and Table S5 and the Supplementary .excel sheet.
Figure 4. Full ACE/ADE models: PI-18 primals on the left (panel a) and tertiary primals on the right (panel b) with broad sense heritability (H2) shown for every primal.
Discussion
In recent years, there has been an increased scientific interest in beliefs people have about the general character of the world and how they relate to other variables, such as personality, health and well-being, yet the genetic influences on individual differences in primal world beliefs remain unknown. That is, one important question is to what extent such primal world beliefs are formed by an individual’s environment or by their genetic predispositions. Here, we address this for the first time by investigating the heritability of primal world beliefs in a Swedish population sample. We, firstly, created a Swedish version of the Primals Inventory and validated the Swedish PI-18. Second, primals showed, as expected, significant though small correlations with the big five personality traits. Finally, genetic factors had a significant contribution to variance in primal world beliefs. Broad sense heritabilities ranged between 21% (Abundant) and 49% (Intentional), with PI-18 primals generally showing similar heritabilities to tertiary primals.
The Swedish primals inventory
We created the first Swedish Primals Inventory and aimed at validating the PI-18 (for a full version of the questionnaire, see Supplementary Material 3). Though the original PI-18 structure in our Swedish sample showed a poor to moderate fit (e.g. for model fit criteria, see Hu & Bentler, Citation1999), Clifton and Yaden (Citation2021) argue this may be acceptable given items being intentionally restrained not only to maximize fit but also to provide a diversity of items indicating sub-dimensionality, required to retain systemic variance not attributable to factor scores, which inevitably lowers CFA fit indicators. Furthermore, exploring the hierarchical primal structures based on our selected 11 tertiary primals revealed a slightly different, yet surprisingly comparable (given that we only include 11 out of 22 tertiary primals, omitting several tertiary primals central to the higher-order primals), factor structure to the one originally described by Clifton et al. (Citation2019). Note that we are not the first to create and explore the Primals Inventory in a non-English-speaking sample. Stahlmann and colleagues, in 2020, equivalently analyzed the overarching structure of hierarchical primals in a German-speaking sample and found six overarching secondary primals. In other words, it may be that the hierarchical structure of primals varies between studies, languages and cultures, and whether primals show a universal hierarchical factor structure across cultures remains to be seen. There may be important cross-cultural differences in primal structures. Certainly, methodological issues may also cause divergent findings. For instance, there may be problems with the translation, e.g. some words being diligently translated may still be differently connoted or nuanced in different cultures. Additionally, reporting low internal consistency (i.e. low Cronbach’s alpha) in our sample and basing our analysis on selected tertiary primals (i.e. not the full 22 tertiary primals) may have further influenced the present findings. This highlights the importance of being careful with extrapolating research results beyond sample and culture.
Primals and personality
Finding that a large proportion of variance in primal world beliefs is unrelated to the big five personality traits, reflected by low to moderate correlations, supports the idea that primals, although related to personality, are also different (Clifton et al., Citation2019). Although correlations of slightly larger magnitudes between primals and personality have been reported previously (Clifton et al., Citation2019; Stahlmann & Ruch, Citation2023; Stahlmann et al., Citation2020), our sample convinces by its size and generalized wide population. An interesting question for future research will be to examine this specific personality–primals relationship, how it is shaped by gene-environment interplay, and how it relates to other outcomes such as health and well-being. For this purpose, further work on genetically informative samples, using multi-variate twin modelling and other genetic analyses will be essential (Posthuma, Citation2009; Turkheimer & Harden, Citation2014).
Heritability of primal world beliefs
Strikingly, we find that beliefs about the general character of the world (i.e. primals) are substantially genetically influenced. This suggests that the formation and implementation of world beliefs is a complex process that depends, on the one hand, on genetic predispositions (familial effects) and, on the other hand, on environmental influences, e.g. individual experiences that may influence a person’s impression of the world. Further analyzing this gene-environment interplay will be a major task for future primals research. As such, finding that primals are partly heritable is in line with general conclusions from behavior genetic research (Polderman et al., Citation2015). Interestingly, while some primals (e.g. Intentional: 49%) are found to be as heritable as personality (~40%; Polderman et al., Citation2015; Vukasović & Bratko, Citation2015), other primals seem to be moderately less genetically influenced, with Abundant at 21% being the lowest broad-sense heritability estimate of all, in the present study. While these lower heritabilities align with estimates for other belief formation heritabilities (e.g. belief updating at 13%; Vellani et al., Citation2022), low heritabilities could also be due to higher measurement errors, reflected in inflated E components (e.g. Rijsdijk & Sham, Citation2002). ADE, and for most primals – apart from Progressive and Regenerative – more parsimonious AE models fitted the data best, with one noticeable exception. That is, for the PI-18 Safe primal the full model that was estimated was an ACE model. While fitting an ACE model for the PI-18 Safe primal could indicate that shared environmental effects may play a role, a simpler AE model explained the data better. Overall, these findings once more align with personality heritability research where often ADE or AE models explain the data best (e.g. Bouchard & McGue, Citation2003; Eaves et al., Citation1998; Keller et al., Citation2005; but see; Weinschenk et al., Citation2022), with shared- or rearing-environment having negligible influences. Indeed, finding that primals seem relatively unaffected by different socio-economic factors (Kerry et al., Citation2023) strengthens the assumption that family environment does not play a systematic role for transmitting primals between parents and their children, though not finding small, yet significant shared environmental effects may be also due to a lack of power (for further discussion see limitation section).
It is important to note that finding significant genetic influences on individual differences in primal world beliefs does not mean that primals cannot change. Rather, it suggests that individuals have a predisposition or tendency which, together with the environmental factors they are exposed to, will form their individual beliefs. Ultimately, and mirroring previous statements (Clifton & Kim, Citation2020; Clifton et al., Citation2019; Stahlmann & Ruch, Citation2023), this gives hope that given the right intervention primals can be changed, yet, what type of intervention works (best) needs to be studied and whether changed primals, indeed, lead to the expected effects, e.g. higher well-being, additionally, remains to be seen.
Limitations
Following the aforementioned restrain of low Cronbach’s alphas for most primals, suggesting relatively low general reliability, several further limitations should be noted. First, as briefly touched upon above, measurement errors and low(er) Cronbach’s alphas might have inflated the E components in our results so that they reflected measurement errors rather than true non-shared environmental effects. Second, our exploratory analysis of the hierarchical structure in our sample is based on only 11 of the 22 tertiary primals, limiting the generalizability of these results. Remarkably, however, even in our selected primals sample a comparable structure to Clifton et al. (Citation2019) original structure was found. As the main focus of the present paper was to assess the heritabilities of primal world beliefs and how they are studied most commonly at present, we based our analyses on the originally proposed tertiary primal structure (see Clifton et al., Citation2019) to create comparable findings. Whether tertiary primal structures hold at item levels across cultures could be, additionally, investigated by future research (see approach Stahlmann et al., Citation2020); however, this is beyond the scope and focus of this paper. Next, we find little to no support for non-additive genetic and shared environmental effects. To note, finding significant C or D components is rare in classical twin modelling, and large power or extended twin models are often needed (Keller et al., Citation2010; Posthuma & Boomsma, Citation2000). Thus, future extended twin models are necessary to validate that shared environmental effects (e.g. parenting styles) have no effect on most primals. However, our sample being middle-aged and the C component being known to decrease with increased age, particularly from childhood to adulthood (Bergen et al., Citation2007), supports our present findings of negligible shared environmental effects. Following from our sample being middle-aged, it should further be noted that our sample is a Swedish, birth cohort restricted sample. Thus, and most importantly, findings may be less generalizable to other birth cohorts, cultures and/or ethnicities, and future research should try to replicate our findings in other populations. Despite these limitations, we would like to highlight that this is, to our knowledge, the first twin study on primal world beliefs, with the sample size being large (even for a twin study).
Conclusion
This study shows that beliefs about the general character of the world are moderately heritable, with broad-sense heritabilities ranging between 21% and 49%. These findings highlight that primal world beliefs are not only formed by ones individual environment but are also genetically influenced. Additionally, we created the first version of the Swedish Primals Inventory, paving the way for future primals research in the Swedish population. Furthermore, positive beliefs about the world were found to be associated with, but substantially distinct from, personality. Altogether, we shed more light onto the genetic nature of primal world beliefs and laid the foundation for future work to analyze the etiology of primals, their associations with numerous other related traits, and the nature of such associations.
Open scholarship


This article has earned the Center for Open Science badges for Open Materials and Preregistered. The materials are openly accessible at https://osf.io/bd543/
Heritability_Primals_Supplements_Details_Twinmodelling.xlsx
Download MS Excel (176.8 KB)Heritability_Primals_Supplements_JoPP.docx
Download MS Word (1.6 MB)Heritability_Primals_Supplements_JoPP.pdf
Download PDF (901.6 KB)Acknowledgments
We would like to acknowledge Erik Pettersson for helping with the translations of the original PI-99, and Klaus Frieler for his consultation regarding the factor analysis statistics. Furthermore, we would like to express our gratitude to all Swedish twins that participated and to the Swedish Twin Registry for providing the data.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
As highly sensitive genetic data were analyzed, data can only be applied for, see https://ki.se/en/research/swedish-twin-registry-for-researchers for more information.
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/17439760.2024.2387340
Additional information
Funding
References
- Beck, J. S. (2020). Cognitive behavior therapy: Basics and beyond. Guilford Publications.
- Bergen, S. E., Gardner, C. O., & Kendler, K. S. (2007). Age-related changes in heritability of behavioral phenotypes over adolescence and Young Adulthood: A meta-analysis. Twin Research and Human Genetics, 10(3), 423–433. https://doi.org/10.1375/twin.10.3.423
- Boker, S. M., Neale, M. C., Maes, H. H., Spiegel, M., Brick, T. R., Estabrook, R., Bates, T. C., Gore, R. J., Hunter, M. D., Pritikin, J. N., Zahery, M., & Kirkpatrick, R. M. (2023). OpenMx: Extended structural equation modelling. https://CRAN.R-project.org/package=OpenMx
- Bouchard, T. J., Jr., & McGue, M. (2003). Genetic and environmental influences on human psychological differences. Journal of Neurobiology, 54(1), 4–45. https://doi.org/10.1002/neu.10160
- Chen, S. X., Lam, B. C. P., Wu, W. C. H., Ng, J. C. K., Buchtel, E. E., Guan, Y., & Deng, H. (2016). Do people’s world views matter? The why and how. Journal of Personality & Social Psychology, 110(5), 743–765. https://doi.org/10.1037/pspp0000061
- Clifton, A. B., Stahlmann, A. G., Hofmann, J., Chirico, A., Cadwallader, R., & Clifton, J. D. (2023). Improving scale equivalence by increasing access to scale-specific information. Perspectives on Psychological Science, 18(4), 843–853. https://doi.org/10.1177/17456916221119396
- Clifton, J. D. W., Baker, J. D., Park, C. L., Yaden, D. B., Clifton, A. B. W., Terni, P., Miller, J. L., Zeng, G., Giorgi, S., Schwartz, H. A., & Seligman, M. E. P. (2019). Primal world beliefs. Psychological Assessment, 31(1), 82–99. https://doi.org/10.1037/pas0000639
- Clifton, J. D. W., & Kim, E. S. (2020). Healthy in a crummy world: Implications of primal world beliefs for health psychology. Medical Hypotheses, 135, 109463. https://doi.org/10.1016/j.mehy.2019.109463
- Clifton, J. D., & Yaden, D. B. (2021). Brief measures of the four highest-order primal world beliefs. Psychological Assessment, 33(12), 1267. https://doi.org/10.1037/pas0001055
- Dallago, F., Mirisola, A., & Roccato, M. (2012). Predicting right-wing authoritarianism via personality and dangerous world beliefs: Direct, indirect, and interactive effects. The Journal of Social Psychology, 152(1), 112–127. https://doi.org/10.1080/00224545.2011.565384
- Dzuka, J., & Dalbert, C. (2006). The belief in a just world and subjective well-being in old age. Aging & Mental Health, 10(5), 439–444. https://doi.org/10.1080/13607860600637778
- Eaves, L. J., Heath, A. C., Hewitt, J. K., Martin, N. G., & Neale, M. C. (1998). Sex differences and non-additivity in the effects of genes on personality. Twin Research, 1(3), 131–137. https://doi.org/10.1375/twin.1.3.131
- Hill, W. G., Goddard, M. E., Visscher, P. M., & Mackay, T. F. C. (2008). Data and theory point to mainly additive genetic variance for complex traits. PLOS Genetics, 4(2), e1000008. https://doi.org/10.1371/journal.pgen.1000008
- Hu, L. T., & Bentler, P. M. (1999). Cutoff criteria for fit indexes in covariance structure analysis: Conventional criteria versus new alternatives. Structural Equation Modeling: A Multidisciplinary Journal, 6(1), 1–55. https://doi.org/10.1080/10705519909540118
- Janoff-Bulman, R. (1989). Assumptive worlds and the stress of traumatic events: Applications of the schema construct. Social Cognition, 7(2), 113–136. https://doi.org/10.1521/soco.1989.7.2.113
- Karwoski, L., Garratt, G. M., & Ilardi, S. S. (2006). On the integration of cognitive-behavioral therapy for depression and positive psychology. Journal of Cognitive Psychotherapy, 20(2), 159–170. https://doi.org/10.1891/jcop.20.2.159
- Keller, M. C., Coventry, W. L., Heath, A. C., & Martin, N. G. (2005). Widespread evidence for non-additive genetic variation in Cloninger’s and Eysenck’s personality dimensions using a twin plus sibling design. Behavior Genetics, 35(6), 707–721. https://doi.org/10.1007/s10519-005-6041-7
- Keller, M. C., Medland, S. E., & Duncan, L. E. (2010). Are extended twin family designs worth the trouble? A comparison of the bias, precision, and accuracy of parameters estimated in four twin family models. Behavior Genetics, 40(3), 377–393. https://doi.org/10.1007/s10519-009-9320-x
- Kerry, N., White, K., O’Brien, M. L., Perry, L. M., & Clifton, J. D. (2023). Despite popular intuition, positive world beliefs poorly reflect several objective indicators of privilege, including wealth, health, sex, and neighborhood safety. Journal of Personality, 92(4), 1129–1142. https://doi.org/10.1111/jopy.12877
- Langston, C. A., & Sykes, W. E. (1997). Beliefs and the big five: Cognitive bases of broad individual differences in personality. Journal of Research in Personality, 31(2), 141–165. https://doi.org/10.1006/jrpe.1997.2178
- Lichtenstein, P., De Faire, U., Floderus, B., Svartengren, M., Svedberg, P., & Pedersen, N. L. (2002). The Swedish twin registry: A unique resource for clinical, epidemiological and genetic studies. Journal of Internal Medicine, 252(3), 184–205. https://doi.org/10.1046/j.1365-2796.2002.01032.x
- Lichtenstein, P., Sullivan, P. F., Cnattingius, S., Gatz, M., Johansson, S., Carlström, E., Björk, C., Svartengren, M., Wolk, A., Klareskog, L., Faire, U. D., Schalling, M., Palmgren, J., & Pedersen, N. L. (2006). The Swedish twin registry in the third millennium: An update. Twin Research and Human Genetics, 9(6), 875–882. https://doi.org/10.1375/183242706779462444
- Ludwig, V. U., Crone, D. L., Clifton, J. D., Rebele, R. W., Schor, J. A., & Platt, M. L. (2023). Resilience of primal world beliefs to the initial shock of the COVID‐19 pandemic. Journal of Personality, 91(3), 838–855. https://doi.org/10.1111/jopy.12780
- Maes, H. H. M., Lapato, D. M., Schmitt, J. E., Luciana, M., Banich, M. T., Bjork, J. M., Hewitt, J. K., Madden, P. A., Heath, A. C., Barch, D. M., Thompson, W. K., Iacono, W. G., & Neale, M. C. (2023). Genetic and environmental variation in continuous phenotypes in the ABCD Study®. Behavior Genetics, 53(1), 1–24. https://doi.org/10.1007/s10519-022-10123-w
- Magnusson, P. K., Almqvist, C., Rahman, I., Ganna, A., Viktorin, A., Walum, H., Halldner, L., Lundström, S., Ullén, F., Långström, N., Larsson, H., Nyman, A., Gumpert, C. H., Råstam, M., Anckarsäter, H., Cnattingius, S., Johannesson, M., Ingelsson, E., … Pedersen, N. L. (2013). The Swedish twin registry: Establishment of a biobank and other recent developments. Twin Research and Human Genetics, 16(1), 317–329. https://doi.org/10.1017/thg.2012.104
- McCrae, R. R., & John, O. P. (1992). An introduction to the five-factor model and its applications. Journal of Personality, 60(2), 175–215. https://doi.org/10.1111/j.1467-6494.1992.tb00970.x
- McGrath, R. E., & Rich, G. (2023). World beliefs, character strengths, and hope for the future. PLOS ONE, 18(6), e0286531. https://doi.org/10.1371/journal.pone.0286531
- Mosing, M. A., Madison, G., Pedersen, N. L., Kuja-Halkola, R., & Ullén, F. (2014). Practice does not make perfect: No causal effect of music practice on music ability. Psychological Science, 25(9), 1795–1803. https://doi.org/10.1177/0956797614541990
- Nudelman, G. (2013). The belief in a just world and personality: A meta-analysis. Social Justice Research, 26(2), 105–119. https://doi.org/10.1007/s11211-013-0178-y
- Polderman, T. J., Benyamin, B., De Leeuw, C. A., Sullivan, P. F., Van Bochoven, A., Visscher, P. M., & Posthuma, D. (2015). Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nature Genetics, 47(7), 702–709. https://doi.org/10.1038/ng.3285
- Posthuma, D. (2009). Multivariate genetic analysis. In Y.-K. Kim (Ed.), Handbook of behavior genetics (pp. 47–59). Springer New York. https://doi.org/10.1007/978-0-387-76727-7_4
- Posthuma, D., Beem, A. L., De Geus, E. J., Van Baal, G. C. M., Von Hjelmborg, J. B., Iachine, I., & Boomsma, D. I. (2003). Theory and practice in quantitative genetics. Twin Research and Human Genetics, 6(5), 361–376. https://doi.org/10.1375/136905203770326367
- Posthuma, D., & Boomsma, D. I. (2000). A note on the statistical power in extended twin designs. Behavior Genetics, 30(2), 147–158. https://doi.org/10.1023/A:1001959306025
- Poulin, M., & Cohen Silver, R. (2008). World benevolence beliefs and well-being across the life span. Psychology and Aging, 23(1), 13–23. https://doi.org/10.1037/0882-7974.23.1.13
- R Core Team. (2023). _R: A language and environment for statistical computing_. R Foundation for Statistical Computing. https://R-project.org/
- Revelle, W. (2023). Psych: Procedures for psychological, psychometric, and personality research. https://CRAN.R-project.org/package=psych
- Rijsdijk, F. V., & Sham, P. C. (2002). Analytic approaches to twin data using structural equation models. Briefings in Bioinformatics, 3(2), 119–133. https://doi.org/10.1093/bib/3.2.119
- Roccas, S., Sagiv, L., Schwartz, S. H., & Knafo, A. (2002). The big five personality factors and personal values. Personality & Social Psychology Bulletin, 28(6), 789–801. https://doi.org/10.1177/0146167202289008
- Rosseel, Y. (2012). Lavaan: An R package for structural equation modeling. Journal of Statistical Software, 48(2), 1–36. https://doi.org/10.18637/jss.v048.i02
- Schur, E., Godfrey, K. M., Dansie, E., Buchwald, D., Pagoto, S., & Afari, N. (2013). Can familial factors account for the association of body mass index with poor mental health in men or women? General Hospital Psychiatry, 35(5), 502–507. https://doi.org/10.1016/j.genhosppsych.2013.04.004
- Seligman, M. E. P., & Csikszentmihalyi, M. (2000). Positive psychology: An introduction. The American Psychologist, 55(1), 5–14. https://doi.org/10.1037/0003-066X.55.1.5
- Soto, C. J., & John, O. P. (2017). The next big five inventory (BFI-2): Developing and assessing a hierarchical model with 15 facets to enhance bandwidth, fidelity, and predictive power. Journal of Personality & Social Psychology, 113(1), 117. https://doi.org/10.1037/pspp0000096
- Stahlmann, A. G., Hofmann, J., Ruch, W., Heintz, S., & Clifton, J. D. (2020). The higher-order structure of primal world beliefs in German-speaking countries: Adaptation and initial validation of the German primals inventory (PI-66-G). Personality & Individual Differences, 163, 110054. https://doi.org/10.1016/j.paid.2020.110054
- Stahlmann, A. G., & Ruch, W. (2023). Primal world beliefs correlate strongly but differentially with character strengths. The Journal of Positive Psychology, 18(4), 481–491. https://doi.org/10.1080/17439760.2022.2070532
- Syropoulos, S. (2020). Personal safety and positive life outcomes: Cross-national evidence from the world values survey. Peace and Conflict: Journal of Peace Psychology, 26(3), 281–292. https://doi.org/10.1037/pac0000444
- Turkheimer, E., & Harden, K. P. (2014). Behavior genetic research methods. Handbook of Research Methods in Social and Personality Psychology, 159–187. https://doi.org/10.1017/CBO9780511996481.012
- Van Hiel, A., Cornelis, I., & Roets, A. (2007). The intervening role of social worldviews in the relationship between the five-factor model of personality and social attitudes. European Journal of Personality, 21(2), 131–148. https://doi.org/10.1002/per.618
- Vazquez, C., Valiente, C., García, F. E., Contreras, A., Peinado, V., Trucharte, A., & Bentall, R. P. (2021). Post-traumatic growth and stress-related responses during the COVID-19 pandemic in a national representative sample: The role of positive core beliefs about the world and others. Journal of Happiness Studies, 22(7), 2915–2935. https://doi.org/10.1007/s10902-020-00352-3
- Vellani, V., Garrett, N., Gaule, A., Patil, K. R., & Sharot, T. (2022). Quantifying the heritability of belief formation. Scientific Reports, 12(1), 11833. https://doi.org/10.1038/s41598-022-15492-0
- Vukasović, T., & Bratko, D. (2015). Heritability of personality: A meta-analysis of behavior genetic studies. Psychological Bulletin, 141(4), 769–785. https://doi.org/10.1037/bul0000017
- Wei, T., & Simko, V. (2021). R package ‘corrplot’: Visualization of a correlation matrix. https://github.com/taiyun/corrplot
- Weinschenk, A., Rasmussen, S. H. R., Christensen, K., Dawes, C., & Klemmensen, R. (2022). The five factor model of personality and heritability: Evidence from Denmark. Personality & Individual Differences, 192, 111605. https://doi.org/10.1016/j.paid.2022.111605
- Wickham, H. (2016). ggplot2: Elegant graphics for data analysis. Springer-Verlag New York. https://ggplot2.tidyverse.org
- Wickham, H., Averick, M., Bryan, J., Chang, W., McGowan, L. D. A., François, R., Grolemund, G., Hayes, A., Henry, L., Hester, J., Kuhn, M., Pedersen, T. L., Miller, E., Bache, S. M., Müller, K., Ooms, J., Robinson, D., Seidel, D. P., … Yutani, H. (2019). Welcome to the tidyverse. Journal of Open Source Software, 4(43), 1686. https://doi.org/10.21105/joss.01686
- Zagai, U., Lichtenstein, P., Pedersen, N. L., & Magnusson, P. K. (2019). The Swedish twin registry: Content and management as a research infrastructure. Twin Research and Human Genetics, 22(6), 672–680. https://doi.org/10.1017/thg.2019.99