
ABSTRACT
We report 130 vertebrate fossils preserved as bony elements and the co-occurring assemblage of fish teeth and spines from the lower strata of the Pisco Formation exposed along the western side of the lower Ica Valley (East Pisco Basin, Peru). Geological mapping at 1:10,000 scale reveals that all these fossils originate from the Langhian–Serravallian P0 allomember. In the study area, P0 is up to ∼40 m thick and features a sandy lower portion, reflecting shoreface deposition, that fines upwards into a package of offshore silts. Marine vertebrates only occur in the lower sandy layers and include whales, dolphins, reptiles, birds, and bony and cartilaginous fishes. The reconstructed paleoenvironment is consistent with a warm-water, marginal-marine setting with a strong connection to the open ocean. This work helps to elucidate the rich yet still poorly understood middle Miocene portions of the Pisco Formation, and highlights the need to conserve this outstanding Fossil-Lagerstätte.
1. Introduction
The middle Miocene to Pliocene Pisco Formation of southern Peru is a shallow-marine sedimentary unit consisting of diatomaceous mudstones, siliciclastic sandstones, and siltstones, nodular dolomite layers, tephra beds, and minor conglomeratic and phosphatic intervals (Brand et al., Citation2011; de Muizon & DeVries, Citation1985; Di Celma, Malinverno, Cantalamessa, et al., Citation2016; Di Celma, Malinverno, Gariboldi, et al., Citation2016; Dunbar et al., Citation1990). The Pisco Formation is known to paleontologists worldwide for the richness, abundance and quality of its vertebrate fossils. The latter include sharks and rays, bony fishes, marine turtles and crocodiles, seabirds, cetaceans (featuring both baleen-bearing and echolocating toothed whales), pinnipeds (seals) and sloths, which collectively provide one of the best record of Neogene marine vertebrates anywhere in the world (e.g. Bianucci, Di Celma, Collareta, et al., Citation2016; Bianucci, Di Celma, Landini, et al., Citation2016; Brand et al., Citation2004; Di Celma et al., Citation2017; de Muizon, Citation1988; de Muizon & DeVries, Citation1985; Esperante et al., Citation2008, Citation2015; Lambert et al., Citation2010). Examples of exceptional preservation range from phosphatized whale baleen plates to stomach contents, regurgitations and even skeletons of cartilaginous fishes like sharks, characterizing the Pisco Formation as a true Konservat-Lagerstätte (Bianucci, Di Celma, Urbina, et al., Citation2016; Bosio et al., Citationin press; Collareta et al., Citation2015, Collareta, Landini, et al., Citation2017; Gariboldi et al., Citation2015; Gioncada et al., Citation2016; Gioncada, Gariboldi, et al., Citation2018; Gioncada, Petrini, et al., Citation2018; Lambert et al., Citation2015; Marx, Collareta, et al., Citation2017).
In the surroundings of the Ocucaje village (Ica desert), integrated studies aimed at elucidating the spatial and stratigraphic distribution of the fossil vertebrates from the Pisco Formation have been recently performed at the localities of Cerro Colorado, Cerro Los Quesos, Cerro Queso Grande and Cerro Ballena (Bianucci, Di Celma, Collareta, et al., Citation2016; Bianucci, Di Celma, Landini, et al., Citation2016; Esperante et al., Citation2015), resulting in the construction of a rather comprehensive overview of the late Miocene vertebrate assemblages from the Pisco Lagerstätte (Di Celma et al., Citation2017). Conversely, the middle Miocene basal strata of the Pisco Formation and their fossil vertebrate content have been only cursorily investigated (Bosio, Malinverno, Collareta, et al., Citation2020; Di Celma et al., Citation2017), although the lower Pisco strata are thought to be home to important fossils such as the holotypes of the basal delphinidan Incacetus broggi (Colbert, Citation1944; de Muizon, Citation1988) and the early-branching cetotheriid whale Tiucetus rosae (Marx, Lambert, et al., Citation2017).
The aim of this work is to report, through a detailed 1:10,000 scale geological map (Main Map), all the middle Miocene fossil vertebrates encountered during paleontological surveys at the localities of Cerro Submarino, Dos Cerritos, Cerro Los Tinajones and Cerro Las Tres Piramides, along the western side of the lower Ica Valley, where the lowermost Pisco strata are abundantly exposed (Di Celma, Malinverno, Bosio, et al., Citation2018). The paleoenvironmental significance of such a record is also discussed considering the distribution of the vertebrate remains in light of the local stratigraphy and facies associations.
2. Geological setting and regional stratigraphy
Since the Mesozoic, the tectonics of Peru have been controlled by the convergence of the oceanic Nazca/Farallon Plate and the continental South American Plate, inducing both the deformation of the leading edge of the latter and the development of a composite transform-convergent margin characterized by normal to strike-slip faults that formed elongated extensional/pull-apart basins along the Peruvian forearc (e.g. Dunbar et al., Citation1990; Kulm et al., Citation1982; León et al., Citation2008). According to Thornburg and Kulm (Citation1981), two trench-parallel structural highs formed on the continental shelf and upper slope in Late Cretaceous–early Paleogene times, namely, the Outer Shelf High and the Upper Slope Ridge. These two ridges segmented the Peruvian offshore into several sedimentary basins that are separated into an outer set of slope basins and an inner set of shelf basins; the latter include the southern Peruvian East Pisco Basin (hereinafter: EPB) and Caballas Basin. The EPB is a 180-km-long, northwest-to-southeast trending sedimentary basin whose sediments are now exposed along a narrow coastal plain between the towns of Pisco and Nazca, being located just landward of where the Peru-Chile trench is impinged on by the aseismic Nazca Ridge (a), a region of buoyant, topographically high oceanic crust (Hampel et al., Citation2004; Hsu, Citation1992; Macharé & Ortlieb, Citation1992; Pilger, Citation1981). The basin fill includes, from oldest to youngest, the Eocene Caballas, Paracas and Otuma formations, the lower Miocene Chilcatay Formation, and the middle–upper Miocene Pisco Formation (Coletti et al., Citation2018, Citation2019; DeVries, Citation1998, Citation2007; DeVries & Jud, Citation2018; DeVries et al., Citation2017; Di Celma et al., Citation2017; Di Celma, Malinverno, Bosio, et al., Citation2018; Di Celma, Malinverno, Collareta, et al., Citation2018; Dunbar et al., Citation1990). These units are heterogeneous in composition and bounded by regionally extensive unconformities, commonly demarcated by lags of igneous pebble- to boulder-sized clasts, which account for periods of subaerial exposure and represent major breaks of the sedimentary record of the EPB (DeVries, Citation1998). Consequently, Di Celma, Malinverno, Bosio, et al. (Citation2018) pointed out that these sedimentary units should be regarded as unconformity-bounded alloformations (NACSN, Citation2005) or, in the genetic sense, as depositional sequences (Catuneanu et al., Citation2009) rather than as formations.
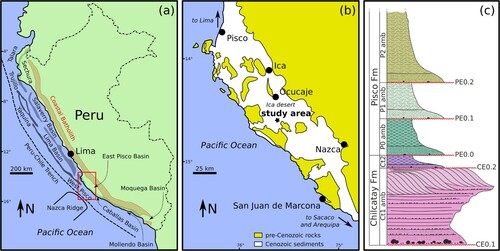
Figure 1. (a) Map of the major Cenozoic sedimentary basins along the coast of Peru. Major structural highs are the Coastal Batholith, the Outer Shelf High and the Upper Slope Ridge. Redrawn and modified after Travis et al. (Citation1976) and Thornburg and Kulm (Citation1981). (b) Schematic geological map of the East Pisco Basin, showing the areas of Cenozoic outcrop, the distribution of pre-Cenozoic rocks, and the location of the study area. Redrawn and modified after DeVries and Schrader (Citation1997). (c) Schematic stratigraphic column of the Chilcatay and Pisco formations and their internal subdivisions in allomembers. Modified after Di Celma, Malinverno, Collareta, et al. (Citation2018).
3. Surveying methods
The investigated area is located some 55–60 km SSE of the town of Ica and covers an area of approximately 33 km2 near the hills of Cerro Submarino, Dos Cerritos, Cerro Los Tinajones, and Cerro Las Tres Piramides (b). The physical stratigraphy and architecture of the studied outcrops were documented through conventional geological field methods, including high-resolution sedimentological logging of a 40-m-thick section, facies analysis, and geological mapping at 1:10,000 scale. The stratigraphic analysis was carried out by combining allostratigraphic and lithostratigraphic criteria, with unconformity-bounded allomembers as the fundamental units and lithostratigraphic units (facies associations) included for complementing the allostratigraphic framework.
As for previous surveys at other fossil-rich localities of the Ica desert (Bianucci, Di Celma, Collareta, et al., Citation2016; Bianucci, Di Celma, Landini, et al., Citation2016; Di Celma, Malinverno, Collareta, et al., Citation2018; Di Celma et al., Citation2019), we made taphonomic observations and preliminarily identified each fossil in the field, and only collected a few significant specimens for deposition at the Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MUSM) in Lima. For each vertebrate specimen encountered during the survey, the geographic position was recorded by means of a handheld GPS and the stratigraphic position along the measured section was established with an accuracy ranging between ±0.4 and ±3 m. The horizontal and vertical distribution of all the recorded fossils is reported in the Main Map. Concerning the isolated fish remains, only teeth of the mega-toothed shark Carcharocles megalodon for which precise GPS coordinates are known were regularly reported on the map and along the stratigraphic column.
4. Stratigraphic architecture and facies distribution
Two pre-Quaternary marine sedimentary units are exposed in the study area, namely, the Chilcatay (lower Miocene) and Pisco (middle–upper Miocene) formations (c).
In agreement with Di Celma, Malinverno, Collareta, et al. (Citation2018; Di Celma et al., Citation2019), the Chilcatay strata have been resolved into two allomembers (Ct1 and Ct2), which are separated by an intraformational unconformity (CE0.2). The lower Ct1 allomember consists of a basal boulder-bearing sandstone and conglomerate facies association (Ct1c), a package of medium- to fine-grained sandstones and siltstones punctuated by beds of coarse-grained sandstones and conglomerates (Ct1a), and an overlying interval of coarse-grained, southwestward dipping, mixed siliciclastic–carbonate clinobeds (Ct1b) (Di Celma, Malinverno, Collareta, et al., Citation2018; Di Celma et al., Citation2019). The Ct1a and Ct1b facies associations are separated by an abrupt lithological break (the KsA surface). The upper Ct2 allomember consists of massive and intensely bioturbated sandstones (Ct2a) changing upwards into massive siltstones (Ct2b). On the southwestern side of the study area, the undeformed Chilcatay deposits described above are truncated to varying degrees by a prominent surface infilled by a package of chaotic sediments, likely reflecting a large submarine slump scar bounded by well-defined lateral shear margins and associated mass transport deposits (MTDs), respectively. At outcrop, the oldest sediments truncated by the basal plane are those of Ct1a. Toward the distal edge of the basal plane, the base of the MTD steps upward, truncating successively younger Ct2a and Ct2b strata to the NE (Di Celma et al., Citation2019).
As observed by Di Celma, Malinverno, Cantalamessa, et al. (Citation2016; Di Celma, Malinverno, Gariboldi, et al., Citation2016; Di Celma, Malinverno, Bosio, et al., Citation2018), the Pisco Formation is locally comprised of three allomembers, designated P0, P1, and P2 from oldest to youngest, which progressively onlap a composite basal unconformity from southwest to northeast. Each Pisco allomember is comprised of a coarse-grained lower portion, reflecting distal lower shoreface to inner shelf deposition, that grades upwards into finer-grained offshore sediments. In particular, the P0 allomember, which is bounded at the base by the PE0.0 unconformity and at the top by the PE0.1 unconformity, comprises a 27-m-thick package of massive, medium-grained sandstones overlain by a 13-m-thick package of massive siltstones ().
Figure 2. Annotated panoramic views of the stratal architecture in the study area and some of the identified key stratal surfaces. (a) Southward view of the most westward hill at Cerro Las Tres Piramides (14°35′29″S–75°38′45″W) showing the upper portion of the Chilcatay Formation and the overlying lower portion of the Pisco Formation. (b) Southeastward view of the central hill at Cerro Las Tres Piramides (14°35′25″S–75°38′18″W). (c) Southward view of the most eastward hill at Cerro Las Tres Piramides (14°35′55″S–75°38′00″W). (d) Southwestward view from the northern nose of the elongated hill named Cerro Submarino (14°34′38″S–75°39′51″W). Encircled car for scale.

Radiometric and biostratigraphic dates from the underlying Burdigalian Chilcatay strata and the overlying Tortonian P1 strata (Bosio, Malinverno, Villa, et al., Citation2020; Gariboldi et al., Citation2017) bracket the age of the P0 allomember between ca. 18.0 and 9.5 Ma (Di Celma, Malinverno, Bosio, et al., Citation2018). Recently, Strontium Isotope Stratigraphy analyses on mollusk and barnacle shells as well as on shark teeth allowed to refine this age estimate, revealing that the P0 sequence belongs to the Langhian–Serravallian interval, between 14.80 and 12.45 Ma (Bosio, Malinverno, Collareta, et al., Citation2020; but see also DeVries & Jud, Citation2018 for a biostratigraphic approach to the age of the lower Pisco strata). Consequently, PE0.0 seemingly reflects a substantial time gap of about 3 Myr.
5. Vertebrate paleontology of the P0 allomember in the study area
One hundred and thirty vertebrate specimens preserved as bony elements were recorded from the P0 strata exposed at the study site (a). Cetaceans dominate this assemblage, being represented by 125 specimens (corresponding to 96.2% of the total number). Of all the cetacean specimens identifiable at the suborder level, totaling 109 partial skeletons, 98 (75.4%) belong to Mysticeti (baleen-bearing whales) and 11 (8.5%) to Odontoceti (echolocating toothed whales). Among mysticetes, 7 specimens show strong similarities with the Northern Hemisphere early-middle Miocene balaenopteroid genus Pelocetus (Kellogg, Citation1965; Kimura et al., Citation2007), whereas 3 compare favorably with the early branching cetotheriid Tiucetus rosae, whose holotype comes from deposits exposed along the eastern bank of the Ica River and possibly correlated with the P0 allomember (Marx, Lambert, et al., Citation2017) (a,b). Among the odontocetes, a single skeleton is identified as a non-kogiid physeteroid (sperm whale) and 5 as delphinidans (extinct relatives of the present-day dolphins and porpoises) (c); the latter include at least two distinct kentriodontid-like taxa, one of which displays some similarities with Incacetus broggi. Remains of non-cetacean tetrapods include 3 (2.3%) specimens identified as reptiles (including one turtle and one crocodylian) and 2 (1.5%) specimens identified as birds (including a skull attributed to the extinct bony-toothed seabird family Pelagornithidae; for a summary of the fossil occurrences of pelagornithids in the upper Miocene Pisco strata, see Chávez et al., Citation2007).
Figure 3. (a) Quantitative composition of the fossil tetrapod assemblage from the P0 strata of the Pisco Formation, based on systematic surface prospecting of 130 specimens preserved as bony elements. (b) Quantitative composition of the fossil fish assemblage from the P0 strata of the Pisco Formation, based on systematic surface prospecting of some 600 specimens preserved as dental or dermal elements.
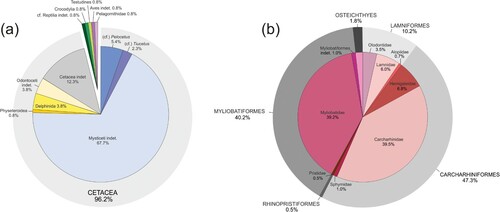
Figure 4. Field photographs depicting fossil tetrapods from the middle Miocene P0 allomember of the Pisco Formation exposed along the western side of the lower Ica Valley (East Pisco Basin, Peru). (a) MLP130, cf. Pelocetus sp., a cranium with articulated mandibles, preserved ventral side-up. (b) MLP36, cf. Tiucetus sp., a cranium with articulated mandibles, preserved dorsal side-up, plus some disarticulated vertebrae. (c) MLP36, Delphinida indet., a partially articulated, semi-eroded skeleton (vertebrae and ribs), with close-ups of three rib fragments with shark bite marks.

The spatial distribution of fossils is not homogeneous in the investigated area: 66 marine vertebrate specimens preserved as bony elements (50.8%, including the turtle and the crocodylian) have been encountered in the vicinities of Cerro Submarino, i.e. in the plain that extends northwest of Cerro Submarino and at the northern edge of Cerro Buque; 35 specimens (26.9%, including the birds and one specimen tentatively identified as a reptile) were found at Dos Cerritos (including the vicinities of Cerro Yesera de Amara); 17 (13.1%) at Cerro Los Tinajones; and 12 (9.2%) at Cerro Las Tres Piramides. The P0 tetrapod assemblage differs from those hosted in the Chilcatay strata exposed at Ullujaya and Zamaca (Bianucci et al., Citation2018; Di Celma, Malinverno, Collareta, et al., Citation2018; Di Celma et al., Citation2019) in its far greater proportion of baleen whales, perhaps reflecting a global trend in mysticete abundance and habitat occupancy (Marx et al., Citation2019), as well as by the absence of members of the odontocete clade Platanidelphidi (Bianucci et al., Citation2020). This assemblage also contrasts with that of P1 (Bianucci, Di Celma, Collareta, et al., Citation2016) by the lower abundance of odontocetes, and especially by the apparent absence of ziphiids (beaked whales), whereas it mainly differs from the P2 assemblage (Bianucci, Di Celma, Landini, et al., Citation2016) by the absence of pinnipeds (seals).
We recorded taphonomic observations for 97 of the cetacean specimens. Thirty-five of them (corresponding to 36.1% of the sample) are represented by articulated skull and postcranial remains, 12 (12.4%) by articulated postcrania, 45 (46.4%) by skull remains with no associated postcrania, and 5 (5.2%) by isolated postcranial or ear bones. Twelve specimens (12.4%) display a complete or partial carbonate nodule (either internal or external) and 11 (11.3%) were found in association with bivalves, including members of Veneridae (Dosinia sp.), Arcidae (Arca sp.), Lucinidae (Miltha sp.) and Pectinidae, and/or gastropods, including members of Architectonicidae, Conidae and Muricidae (Chorus sp.). Three cetacean specimens (3.1%) exhibit elongated incisions that are morphologically consistent with shark bite marks (c), and shark teeth were furthermore found in close association with another specimen.
Shark tooth-rich horizons as those seen in the Chilcatay Formation (Bianucci et al., Citation2018; Landini et al., Citation2019) and the P1 allomember of the Pisco Formation (Landini, Altamirano-Sierra, et al., Citation2017; Landini, Collareta, et al., Citation2017) have not been recognized in P0. Nevertheless, teeth and dermal elements of various types of fishes are rather commonly found in P0 and comprise a relatively diverse assemblage (b). Sharks and rays contribute the vast majority of the total fish assemblage, and are represented by Lamniformes (accounting for 10.2% of the fish specimens and including Alopias vulpinus, Carcharocles megalodon, Cosmopolitodus hastalis, and Isurus oxyrinchus), Carcharhiniformes (47.3%, including Carcharhinus brachyurus, Carcharhinus cf. leucas, Carcharhinus sp., Galeocerdo aduncus, Hemipristis serra, Physogaleus contortus, and Sphyrna cf. zygaena), Myliobatiformes (40.2%, including Aetobatus cf. narinari, Myliobatidae indet., and Myliobatiformes indet., the latter consisting of caudal spines) and Rhinopristiformes (0.5%, consisting of rostral spines of Anoxypristis cf. cuspidata). Sparid-like molariform teeth of bony fishes (1.8%) are also found in P0. This fish assemblage () markedly differs from those from the Chilcatay Formation (Bianucci et al., Citation2018; Landini et al., Citation2019) and the P1 and P2 allomembers of the Pisco Formation (Landini, Altamirano-Sierra, et al., Citation2017; Landini, Collareta, et al., Citation2017) by the very rare presence of teeth of the shortfin mako I. oxyrinchus, as well as by the great abundance of teeth referred to the extinct snaggletooth shark H. serra, which is in turn absent from the upper Miocene strata of the Pisco Formation. Myliobatiform stingrays are also characteristically abundant in P0, accounting for more than two fifths of the elasmobranch specimens.
Figure 5. Dental and dermal remains of bony and cartilaginous fishes from the middle Miocene P0 allomember of the Pisco Formation exposed along the western side of the lower Ica Valley (East Pisco Basin, Peru). (a) Osteichthyan (sparid?) molariform tooth. (b) Carcharocles megalodon tooth. (c) Alopias vulpinus tooth. (d) Cosmopolitodus hastalis tooth. (e) Myliobatoidei indet. caudal spine. (f) Hemipristis serra tooth. (g) Carcharhinus cf. leucas tooth. (h) Galeocerdo aduncus tooth. (i) Physogaleus contortus tooth. (j) Carcharhinus brachyurus tooth. (k) Sphyrna cf. zygaena tooth. (l) Anoxypristis cf. cuspidata rostral spine. Specimen in panel A is shown in apical view; specimens in panels E and L are shown in dorsal view; all other specimens are shown in lingual view.

From a stratigraphic point of view, all the vertebrate fossils from P0 were recovered from the lower sandy portion of this sequence, and most of them come from the basal meters of this stratal division: indeed, 111 out of 130 tetrapod specimens were found in the lowermost 10 m of the P0 sequence.
6. Paleoecological and paleoenvironmental inferences
The elasmobranch assemblage from P0 exhibits an obviously thermophilic taxonomic imprint that differs from the cooler conditions that are hypothesized for the upper Miocene portion of the Pisco Formation (Amiot et al., Citation2008; DeVries & Frassinetti, Citation2003; Dunbar et al., Citation1990). The co-occurrence of taxa such as Crocodylia indet., Aetobatus cf. narinari, Anoxypristis cf. cuspidata, Carcharhinus cf. leucas and Hemipristis serra, the latter being known from a rather high number of teeth, allows to reconstruct tropical to subtropical thermal affinities for the middle Miocene Ica desert paleo-area (Bosio, Malinverno, Collareta, et al., Citation2020). This interpretation is supported by the presence of remains of warm-water invertebrates, such as architectonicid, conid and cypraeid gastropods, the gastropod species Ficus distans, and the sole coral colony ever collected so far from the EPB, which was referred to the family Rhizangiidae (Bosio, Malinverno, Collareta, et al., Citation2020; DeVries et al., Citation2006; DeVries & Frassinetti, Citation2003; Di Celma et al., Citation2017). The shift to seemingly cooler conditions corresponding to the PE0.1 surface might reflect the exit from the Middle Miocene Climatic Optimum as well as a late Miocene strengthening of the Humboldt Current coupled with an intensification of the coastal upwelling regime (Bosio, Malinverno, Collareta, et al., Citation2020).
The shark and ray assemblage furthermore suggests a shallow-marine environment close to the coast. Coupled with the likely presence of the bull shark (C. cf. leucas), the occurrence of the knifetooth sawfish A. cf. cuspidata might indicate proximity to a river mouth. On the other hand, the occurrence of teeth of the common thresher shark (Alopias vulpinus) and the abundance of large-sized (estimated total body length around 11.8–13.7 m; Bianucci et al., Citation2019) Pelocetus-like mysticetes imply a well-developed connection with the open sea (IUCN, Citation2020).
The abundance of eagle ray (i.e. myliobatine) teeth, as well as the relatively frequent observation of molar-like teeth of osteichthyans, matches well the observation of a rather rich hard-shell benthic biota that would have provided an important food source for the P0 marginal-marine vertebrates (an observation that also implies mostly oxic conditions at the seafloor). The bite marks observed on some cetacean specimens indicate predation or scavenging by sharks upon baleen whales and delphinidans, as in younger deposits of the Sacaco area that have been usually referred to the Pisco Formation (e.g. Collareta, Lambert, et al., Citation2017).
The striking abundance of baleen whale specimens in the P0 shoreface sands, as well as their preservation patterns (e.g. the prevalence of isolated skulls), might partly be explained by some process of accumulation of floating mysticete carcasses in this marginal-marine paleoenvironment (see also Schäfer, Citation1972).
7. Concluding remarks
The abundant fossils of the EPB provide a uniquely detailed window into the history of marine life, and as such, they form a crucial part of Peru’s natural heritage. Protecting this heritage requires a better understanding of the location, characteristics, and relevance of its component sites and strata. Here, we present the first detailed account of the richness and structure of the fossil vertebrate assemblage of the still poorly known P0 strata. As such, this study represents a first detailed census of the richness, abundance and diversity of the fossil vertebrates from the middle Miocene portion of the Pisco Formation, which fills a crucial gap between the better known records from the early Miocene Chilcatay deposits and the upper Miocene P1 and P2 Pisco strata. Thematic maps like ours significantly contribute to a growing body of geological and paleontological knowledge, whose eventual synthesis will hopefully lend pivotal support to the protection of the extraordinary geoheritage of the Ica Desert.
Software
The geological map was compiled by scanning hand drafts as black and white TIF files, and then digitizing the line art using the Corel Draw X3 graphics package. We used the GIS Data processing application Global Mapper 12 to generate contour lines for the 1:10,000 scale topographic base map. To do so, we relied on digital elevation model (DEM) based on the Shuttle Radar Topography Mission 26 (SRTM), as released by the United States Geological Survey (SRTM3 USGS version 2.1). The background aerial imagery is from World Imagery (ESRI, https://services.arcgisonline.com/ArcGIS/rest/services/World_Imagery/MapServer).
JoM_P0_Main_Map_revised.pdf
Download PDF (16.3 MB)Acknowledgments
Our gratitude to Anna Gioncada, Gabriella Bagnoli, Giancarlo Molli, Giovanni Sarti, Christian de Muizon, Rodolfo Salas-Gismondi, Rafael M. Varas-Malca, Thomas J. DeVries, Walter Aguirre, Piero Giuffra, and Francesco Talini for their support, either in the field or in the lab (sometimes both), as well as for much fruitful discussions on the geology and paleontology of the P0 strata. Not least, the journal reviewers Ricardo N. Melchor, Chris Orton, Ronald Steel, and the Associate Editor Arthur Merschat are warmly acknowledged for their thoughtful and constructive criticism that strongly contributed to sharpen the focus of the present study.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Amiot, R., Göhlich, U. B., Lécuyer, C., de Muizon, C., Cappetta, H., Fourel, F., Héran, M. A., & Martineau, F. (2008). Oxygen isotope compositions of phosphate from Middle Miocene–Early Pliocene marine vertebrates of Peru. Palaeogeography, Palaeoclimatology, Palaeoecology, 264(1–2), 85–92. https://doi.org/https://doi.org/10.1016/j.palaeo.2008.04.001
- Bianucci, G., Collareta, A., Bosio, G., Landini, W., Gariboldi, K., Gioncada, A., Lambert, O., Malinverno, E., de Muizon, C., Varas-Malca, R., Villa, I. M., Coletti, G., Urbina, M., & Di Celma, C. (2018). Taphonomy and palaeoecology of the lower Miocene marine vertebrate assemblage of Ullujaya (Chilcatay Formation, East Pisco Basin, southern Peru). Palaeogeography, Palaeoclimatology, Palaeoecology, 511, 256–279. https://doi.org/https://doi.org/10.1016/j.palaeo.2018.08.013
- Bianucci, G., Di Celma, C., Collareta, A., Landini, W., Post, K., Tinelli, C., de Muizon, C., Bosio, G., Gariboldi, K., Gioncada, A., Malinverno, E., Cantalamessa, G., Altamirano-Sierra, A., Salas-Gismondi, R., Urbina, M., & Lambert, O. (2016). Fossil marine vertebrates of Cerro Los Quesos: Distribution of cetaceans, seals, crocodiles, seabirds, sharks, and bony fish in a late Miocene locality of the Pisco Basin, Peru. Journal of Maps, 12(5), 1037–1046. https://doi.org/https://doi.org/10.1080/17445647.2015.1115785
- Bianucci, G., Di Celma, C., Landini, W., Post, K., Tinelli, C., de Muizon, C., Gariboldi, K., Malinverno, E., Cantalamessa, G., Gioncada, A., Collareta, A., Salas-Gismondi, R., Varas-Malca, R. M., Urbina, M., & Lambert, O. (2016). Distribution of fossil marine vertebrates in Cerro Colorado, the type locality of the giant raptorial sperm whale Livyatan melvillei (Miocene, Pisco Formation, Peru). Journal of Maps, 12(3), 543–557. https://doi.org/https://doi.org/10.1080/17445647.2015.1048315
- Bianucci, G., Di Celma, C., Urbina, M., & Lambert, O. (2016). New beaked whales from the late Miocene of Peru and evidence for convergent evolution in stem and crown Ziphiidae (Cetacea, Odontoceti). PeerJ, 4, e2479. https://doi.org/https://doi.org/10.7717/peerj.2479
- Bianucci, G., Marx, F. G., Collareta, A., Di Stefano, A., Landini, W., Morigi, C., & Varola, A. (2019). Rise of the titans: Baleen whales became giants earlier than thought. Biology Letters, 15(5), article #20190175. https://doi.org/https://doi.org/10.1098/rsbl.2019.0175
- Bianucci, G., de Muizon, C., Urbina, M., & Lambert, O. (2020). Extensive diversity and disparity of the early Miocene Platanistoids (Cetacea, Odontoceti) in the Southeastern Pacific (Chilcatay Formation, Peru). Life, 10, article #27. https://doi.org/https://doi.org/10.3390/life10030027
- Bosio, G., Gioncada, A., Gariboldi, K., Bonaccorsi, E., Collareta, A., Pasero, M., Di Celma, C., Malinverno, E., Urbina, M., & Bianucci, G. (in press). Mineralogical and geochemical characterization of fossil bones from a Miocene marine Konservat-Lagerstätte. Journal of South American Earth Sciences, in press. doi:https://doi.org/10.1016/j.jsames.2020.102924
- Bosio, G., Malinverno, E., Collareta, A., Di Celma, C., Gioncada, A., Parente, M., Berra, F., Marx, F. G., Vertino, A., Urbina, M., & Bianucci, G. (2020). Strontium isotope stratigraphy and the thermophilic fossil fauna from the middle Miocene of the East Pisco Basin (Peru). Journal of South American Earth Sciences, 97, article #102399. https://doi.org/https://doi.org/10.1016/j.jsames.2019.102399
- Bosio, G., Malinverno, E., Villa, I. M., Di Celma, C., Gariboldi, K., Gioncada, A., Barberini, V., Urbina, M., & Bianucci, G. (2020). Tephrochronology and chronostratigraphy of the Miocene Chilcatay and Pisco formations (East Pisco Basin, Peru). Newsletters on Stratigraphy, 53(2), 213–247. https://doi.org/https://doi.org/10.1127/nos/2019/0525
- Brand, L. R., Esperante, R., Chadwick, A. V., Porras, O. P., & Alomía, M. (2004). Fossil whale preservation implies high diatom accumulation rate in the Miocene–Pliocene Pisco Formation of Peru. Geology, 32(2), 165–168. https://doi.org/https://doi.org/10.1130/G20079.1
- Brand, L. R., Urbina, M., Chadwick, A. V., DeVries, T. J., & Esperante, R. (2011). A high-resolution stratigraphic framework for the remarkable fossil cetacean assemblage of the Miocene/Pliocene Pisco Formation, Peru. Journal of South American Earth Sciences, 31(4), 414–425. https://doi.org/https://doi.org/10.1016/j.jsames.2011.02.015
- Catuneanu, O., Abreu, V., Bhattacharya, J. P., Blum, M. D., Dalrymple, R. W., Eriksson, P. G., Fielding, C. R., Fisher, W. L., Galloway, W. E., Gibling, M. R., Giles, K. A., Holbrook, J. M., Jordan, R., Kendall, C. G. S. C., Macurda, B., Martinsen, O. J., Miall, A. D., Neal, J. E., Nummedal, D., … Winker, C. (2009). Towards the standardization of sequence stratigraphy. Earth-Science Reviews, 92(1-2), 1–33. https://doi.org/https://doi.org/10.1016/j.earscirev.2008.10.003
- Chávez, M., Stucchi, M., & Urbina, M. (2007). El registro de Pelagornithidae (Aves: Pelecaniformes) y la avifauna neógena del Pacífico sudeste. Bulletin de l’Institut français d’Etudes andines, 36(2), 175–197. https://doi.org/https://doi.org/10.4000/bifea.3780
- Colbert, E. H. (1944). A new fossil whale from the Miocene of Peru. Bulletin of the American Musem of Natural History, 83, 195–216.
- Coletti, G., Bosio, G., Collareta, A., Buckeridge, J. S., Consani, S., & El Kateb, A. (2018). Palaeoenvironmental analysis of the Miocene barnacle facies: Case studies from Europe and South America. Geologica Carpathica, 69(6), 573–592. https://doi.org/https://doi.org/10.1515/geoca-2018-0034
- Coletti, G., Bosio, G., Collareta, A., Malinverno, E., Bracchi, V., Di Celma, C., Basso, D., Stainbank, S., Spezzaferri, S., Cannings, T., & Bianucci, G. (2019). Biostratigraphic, evolutionary, and paleoenvironmental significance of the southernmost lepidocyclinids of the Pacific coast of South America (East Pisco Basin, southern Peru). Journal of South American Earth Sciences, 96, article #102372. https://doi.org/https://doi.org/10.1016/j.jsames.2019.102372
- Collareta, A., Lambert, O., Landini, W., Di Celma, C., Malinverno, E., Varas-Malca, R., Urbina, M., & Bianucci, G. (2017). Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru. Palaeogeography, Palaeoclimatology, Palaeoecology, 469, 84–91. https://doi.org/https://doi.org/10.1016/j.palaeo.2017.01.001
- Collareta, A., Landini, W., Chalcatana, C., Valdivia, W., Altamirano-Sierra, A., Urbina-Schimtt, M., & Bianucci, G. (2017). A well preserved skeleton of the fossil shark Cosmopolitodus hastalis from the late Miocene of Peru, featuring fish remains as fossilized stomach contents. Rivista Italiana di Paleontologia e Stratigrafia, 123, 11–22. https://doi.org/https://doi.org/10.13130/2039-4942/8005
- Collareta, A., Landini, W., Lambert, O., Post, K., Tinelli, C., Di Celma, C., Panetta, D., Tripodi, M., Salvadori, P. A., Caramella, D., Marchi, D., Urbina, M., & Bianucci, G. (2015). Piscivory in a Miocene Cetotheriidae of Peru: First record of fossilized stomach content for an extinct baleen-bearing whale. The Science of Nature, 102(11-12), article #70. https://doi.org/https://doi.org/10.1007/s00114-015-1319-y
- de Muizon, C. (1988). Les Vertébrés de la Formation Pisco (Pérou). Troisième partie: Les Odontocètes (Cetacea, Mammalia) du Miocène. Travaux de l’Institut Français d’Études Andines, 42, 1–2-44.
- de Muizon, C. & DeVries, T. J. (1985). Geology and paleontology of late Cenozoic marine deposits in the Sacaco area (Peru). Geologische Rundschau, 74(3), 547–563. https://doi.org/https://doi.org/10.1007/BF01821211
- DeVries, T. J. (1998). Oligocene deposition and Cenozoic sequence boundaries in the Pisco Basin (Peru). Journal of South American Earth Sciences, 11(3), 217–231. https://doi.org/https://doi.org/10.1016/S0895-9811(98)00014-5
- DeVries, T. J. (2007). Molluscan evidence bearing on Cenozoic warm upwelling off southern Peru. Geological Society of America Abstracts with Programs, 39, 78.
- DeVries, T. J., & Frassinetti, D. (2003). Range extensions and biogeographic implications of Chilean Neogene mollusks found in Peru. Boletín del Museo Nacional de Historia Natural, Chile, 52, 119–135.
- DeVries, T. J., Groves, L. T., & Urbina, M. (2006). A new early miocene Muracypraea Woodring, 1957 (Gastropoda: Cypraeidae) from the Pisco Basin of southern Peru. The Nautilus, 120, 101–105.
- DeVries, T. J., & Jud, N. A. (2018). Lithofacies patterns and paleogeography of the Miocene Chilcatay and lower Pisco depositional sequences (East Pisco Basin, Peru). Boletín de la Sociedad Geológica del Perú, Volumen Jubilar, 8, 124–167.
- DeVries, T. J., & Schrader, H. (1997). Middle Miocene marine sediments in the Pisco basin(Peru). Boletín de la Sociedad Geológica del Perú, 87, 1–13.
- DeVries, T. J., Urbina, M., & Jud, N. A. (2017). The Eocene-Oligocene Otuma depositional sequence (East Pisco Basin, Peru): Paleogeographic and paleoceanographic implications of new data. Boletín de la Sociedad Geológica del Perú, 112, 14–38.
- Di Celma, C., Malinverno, E., Bosio, G., Collareta, A., Gariboldi, K., Gioncada, A., Molli, G., Basso, D., Varas-Malca, R. M., Pierantoni, P. P., Villa, I. M., Lambert, O., Landini, W., Sarti, G., Cantalamessa, G., Urbina, M., & Bianucci, G. (2017). Sequence stratigraphy and paleontology of the upper Miocene Pisco Formation along the western side of the lower Ica valley (Ica Desert, Peru). Rivista Italiana Paleontologia e Stratigrafia, 123, 255–274. https://doi.org/https://doi.org/10.13130/2039-4942/8373
- Di Celma, C., Malinverno, E., Bosio, G., Gariboldi, K., Collareta, A., Gioncada, A., Landini, W., Pierantoni, P. P., & Bianucci, G. (2018). Intraformational unconformities as a record of late Miocene eustatic falls of sea level in the Pisco Formation (southern Peru). Journal of Maps, 14(2), 607–619. https://doi.org/https://doi.org/10.1080/17445647.2018.1517701
- Di Celma, C., Malinverno, E., Cantalamessa, G., Gioncada, A., Bosio, G., Villa, I. M., Gariboldi, K., Rustichelli, A., Pierantoni, P. P., Landini, W., Tinelli, C., Collareta, A., & Bianucci, G. (2016). Stratigraphic framework of the late Miocene Pisco Formation at Cerro Los Quesos (Ica Desert, Peru). Journal of Maps, 12(5), 1020–1028. https://doi.org/https://doi.org/10.1080/17445647.2015.1115783
- Di Celma, C., Malinverno, E., Collareta, A., Bosio, G., Gariboldi, K., Lambert, O., Landini, W., Gioncada, A., Villa, I. M., Coletti, G., de Muizon, C., Urbina, M., & Bianucci, G. (2018). Facies analysis, stratigraphy and marine vertebrate assemblage of the lower Miocene Chilcatay Formation at Ullujaya (Pisco basin, Peru). Journal of Maps, 14(2), 257–268. https://doi.org/https://doi.org/10.1080/17445647.2018.1456490
- Di Celma, C., Malinverno, E., Gariboldi, K., Gioncada, A., Rustichelli, A., Pierantoni, P. P., Landini, W., Bosio, G., Tinelli, C., & Bianucci, G. (2016). Stratigraphic framework of the late Miocene to Pliocene Pisco Formation at Cerro Colorado (Ica Desert, Peru). Journal of Maps, 12(3), 515–529. https://doi.org/https://doi.org/10.1080/17445647.2015.1047906
- Di Celma, C., Pierantoni, P. P., Malinverno, E., Collareta, A., Lambert, O., Landini, W., Bosio, G., Gariboldi, K., Gioncada, A., de Muizon, C., Molli, G., Marx, F. G., Varas-Malca, R. M., Urbina, M., & Bianucci, G. (2019). Allostratigraphy and paleontology of the lower Miocene Chilcatay Formation in the Zamaca area, East Pisco basin, southern Peru. Journal of Maps, 15(2), 393–405. https://doi.org/https://doi.org/10.1080/17445647.2019.1604439
- Dunbar, R. B., Marty, R. C., & Baker, P. A. (1990). Cenozoic marine sedimentation in the Sechura and Pisco basins, Peru. Palaeogeography, Palaeoclimatology, Palaeoecology, 77(3-4), 235–261. https://doi.org/https://doi.org/10.1016/0031-0182(90)90179-B
- Esperante, R., Brand, L., Nick, K. E., Poma, O., & Urbina, M. (2008). Exceptional occurrence of fossil baleen in shallow marine sediments of the Neogene Pisco Formation, Southern Peru. Palaeogeography, Palaeoclimatology, Palaeoecology, 257(3), 344–360. https://doi.org/https://doi.org/10.1016/j.palaeo.2007.11.001
- Esperante, R., Brand, L. R., Chadwick, A. V., & Poma, O. (2015). Taphonomy and paleoenvironmental conditions of deposition of fossil whales in the diatomaceous sediments of the Miocene/Pliocene Pisco Formation, southern Peru—a new fossil-lagerstätte. Palaeogeography, Palaeoclimatology, Palaeoecology, 417, 337–370. https://doi.org/https://doi.org/10.1016/j.palaeo.2014.09.029
- Gariboldi, K., Bosio, G., Malinverno, E., Gioncada, A., Di Celma, C., Villa, I. M., Urbina, M., & Bianucci, G. (2017). Biostratigraphy, geochronology and sedimentation rates of the upper Miocene Pisco Formation at two important marine vertebrate fossil-bearing sites of southern Peru. Newsletters on Stratigraphy, 50(4), 417–444. https://doi.org/https://doi.org/10.1127/nos/2017/0345
- Gariboldi, K., Gioncada, A., Bosio, G., Malinverno, E., Di Celma, C., Tinelli, C., Cantalamessa, G., Landini, W., Urbina, M., & Bianucci, G. (2015). The dolomite nodules enclosing fossil marine vertebrates in the East Pisco Basin, Peru: Field and petrographic insights into the Lagerstätte formation. Palaeogeography, Palaeoclimatology, Palaeoecology, 438, 81–95. https://doi.org/https://doi.org/10.1016/j.palaeo.2015.07.047
- Gioncada, A., Collareta, A., Gariboldi, K., Lambert, O., Di Celma, C., Bonaccorsi, E., Urbina, M., & Bianucci, G. (2016). Inside baleen: Exceptional microstructure preservation in a late Miocene whale skeleton from Peru. Geology, 44(10), 839–842. https://doi.org/https://doi.org/10.1130/G38216.1
- Gioncada, A., Gariboldi, K., Collareta, A., Di Celma, C., Bosio, G., Malinverno, E., Lambert, O., Pike, J., Urbina, M., & Bianucci, G. (2018). Looking for the key to preservation of fossil marine vertebrates in the Pisco Formation of Peru: New insights from a small dolphin skeleton. Andean Geology, 45(3), 379–398. https://doi.org/https://doi.org/10.5027/andgeoV45n3-3122
- Gioncada, A., Petrini, R., Bosio, G., Gariboldi, K., Collareta, A., Malinverno, E., Bonaccorsi, E., Di Celma, C., Pasero, M., Urbina, M., & Bianucci, G. (2018). Insights into the diagenetic environment of fossil marine vertebrates of the Pisco Formation (late Miocene, Peru) from mineralogical and Sr-isotope data. Journal of South American Earth Sciences, 81, 141–152. https://doi.org/https://doi.org/10.1016/j.jsames.2017.11.014
- Hampel, A., Kukowski, N., Bialas, J., Heubscher, C., & Heinbockel, R. (2004). Ridge subduction at an erosive margin: The collision zone of the Nazca Ridge in southern Peru. Journal of Geophysical Research: Solid Earth, 109(B2), article #B02101. https://doi.org/https://doi.org/10.1029/2003JB002593
- Hsu, J. T. (1992). Quaternary uplift of the Peruvian coast related to the subduction of the Nazca Ridge: 13.5 to 15.6 degrees South latitude. Quaternary International, 15-16, 87–97. https://doi.org/https://doi.org/10.1016/1040-6182(92)90038-4
- International Union for Conservation of Nature (IUCN). (2020). The IUCN Red List of Threatened Species. Version 2020-1. Retrieved March 19, 2020, from https://www.iucnredlist.org
- Kellogg, R. (1965). Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia, part 1: A new whalebone whale from the Miocene Calvert Formation. U.S. National Museum Bulletin, 247, 1–45.
- Kimura, T., Hasegawa, Y., Ohzawa, H., Yamaoka, T., Furukawa, Y., Ueda, T., Kiyoshi, T., Sugihara, M., & Sakuda, M. (2007). A mysticete whale (Cetacea) skeleton from the middle Miocene Bihoku Group, Shobara, Hiroshima, Japan. Miscellaneous Reports of the Hiwa Museum for Natural History, 48, 1–10.
- Kulm, L. D., Resig, J. M., Thornburg, T. M., & Schrader, H. J. (1982). Cenozoic structure, stratigraphy and tectonics of the central Peru forearc. In J. K. Legget (Ed.), Trench and forearc geology: Sedimentation and tectonics on modern and ancient plate margins (pp. 151–169). Blackwells.
- Lambert, O., Bianucci, G., Post, K., De Muizon, C., Salas-Gismondi, R., Urbina, M., & Reumer, J. (2010). The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nature, 466(7302), 105–108. https://doi.org/https://doi.org/10.1038/nature09067
- Lambert, O., Collareta, A., Landini, W., Post, K., Ramassamy, B., Di Celma, C., Urbina, M., & Bianucci, G. (2015). No deep diving: Evidence of predation on epipelagic fish for a stem beaked whale from the Late Miocene of Peru. Proceedings of the Royal Society B: Biological Sciences, 282(1815), article #20151530. https://doi.org/https://doi.org/10.1098/rspb.2015.1530
- Landini, W., Altamirano-Sierra, A., Collareta, A., Di Celma, C., Urbina, M., & Bianucci, G. (2017). The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru). Journal of South American Earth Sciences, 73, 168–190. https://doi.org/https://doi.org/10.1016/j.jsames.2016.12.010
- Landini, W., Collareta, A., Di Celma, C., Malinverno, E., Urbina, M., & Bianucci, G. (2019). The early Miocene elasmobranch assemblage from Zamaca (Chilcatay Formation, Peru). Journal of South American Earth Sciences, 91, 352–371. https://doi.org/https://doi.org/10.1016/j.jsames.2018.08.004
- Landini, W., Collareta, A., Pesci, F., Di Celma, C., Urbina, M., & Bianucci, G. (2017). A secondary nursery area for the copper shark Carcharhinus brachyurus from the late Miocene of Peru. Journal of South American Earth Sciences, 78, 164–174. https://doi.org/https://doi.org/10.1016/j.jsames.2017.07.003
- León, W., Aleman, A., Torres, V., Rosell, W., & De la Cruz, O. (2008). Estratigrafía, sedimentología y evolución tectónica de la cuenca Pisco Oriental. Boletín INGEMMET, 27, 144 pp.
- Macharé, J., & Ortlieb, L. (1992). Plio-Quaternary vertical motions and the subduction of the Nazca Ridge, central coast of Peru. Tectonophysics, 205(1-3), 97–108. https://doi.org/https://doi.org/10.1016/0040-1951(92)90420-B
- Marx, F. G., Collareta, A., Gioncada, A., Post, K., Lambert, O., Bonaccorsi, E., Urbina, M., & Bianucci, G. (2017). How whales used to filter: Exceptionally preserved baleen in a Miocene cetotheriid. Journal of Anatomy, 231(2), 212–220. https://doi.org/https://doi.org/10.1111/joa.12622
- Marx, F. G., Fitzgerald, E. M., & Fordyce, R. E. (2019). Like phoenix from the ashes: How modern baleen whales arose from a fossil “dark age”. Acta Palaeontologica Polonica, 64, 231–238. https://doi.org/https://doi.org/10.4202/app.00575.2018
- Marx, F. G., Lambert, O., & de Muizon, C. (2017). A new Miocene baleen whale from Peru deciphers the dawn of cetotheriids. Royal Society Open Science, 4, article #170560. https://doi.org/https://doi.org/10.1098/rsos.160542
- NACSN (North American Commission on Stratigraphic Nomenclature). (2005). North American stratigraphic code. AAPG Bulletin, 89(11), 1547–1591. https://doi.org/https://doi.org/10.1306/07050504129
- Pilger, R. H. (1981). Plate reconstructions, aseismic ridges, and low-angle subduction beneath the Andes. Geological Society of America Bulletin, 92(7), 448–456. https://doi.org/https://doi.org/10.1130/0016-7606(1981)92<448:PRARAL>2.0.CO;2
- Schäfer, W. (1972). Ecology and palaeoecology of Marine environments. University of Chicago Press.
- Thornburg, T. M., & Kulm, L. D. (1981). Sedimentary basins of the Peru continental margin: Structure, stratigraphy, and Cenozoic tectonics from 6°S to 16°S latitude. In L. D. Kulm, J. Dymond, E. J. Dasch, & D. M. Hussong (Eds.), Nazca plate: Crustal formation and Andean convergence (pp. 393–422). Washington: Geological Society of America.
- Travis, R. B., Gonzales, G., & Pardo, A. (1976). Hydrocarbon potential of coastal basins of Peru. AAPG Memoir, 25, 331–338. https://doi.org/https://doi.org/10.1306/83D918CB-16C7-11D7-8645000102C1865D