
ABSTRACT
A map of plant formations of the Sierra del Aramo (Asturian Central Massif) is presented at a scale of 1:25,000. The interest of this map lies in the idiosyncrasies of the area studied: it straddles two contrasting biogeographic districts, has carbonated and siliceous substrates, and displays various types of hillside deposits. It also has a height difference of more than 1300 m in 3 linear km, varied topography and orientations, and a history of human colonization prior to the Bronze Age. All this in just 220 km2, in a rural area in the process of abandonment but only 20 km from one of the regional metropolitan areas. In short, the Sierra del Aramo is a territory of high natural and cultural heritage, in which the vegetation map is essential as a fundamental scientific tool for territorial management.
1. Introduction
Vegetation maps have two main values: their own idiosyncrasy as a cartographic documentary source and their configuring a basic tool in both territorial management and scientific and technical work (CitationBarrera, 2017). Vegetation is the most perceptible element of the physical environment of much of the world's landscapes including the Sierra del Aramo, except in those areas where it is absent and the rock emerges (CitationBeato Bergua, 2018). In addition, it has an impact on water flows and modeling processes, and in this way plant formations play a major role in the composition of the landscape (CitationBertrand, 1966; CitationIbarra, 1993a, Citation1993b; CitationMartínez de Pisón, 1983). Through an understanding of structure and dynamics of vegetation, data are obtained not only on the ecological behavior, present situation, and physiognomy of communities, but also on the relationship between the vegetation in question and the anthropic factor. Thus, the vegetation offers us the chance to understand the evolution and the current state of a landscape system (CitationBertrand, 1964; CitationBertrand & Bertrand, 1986), especially in highly anthropized areas where there are elevated natural values that can be or become integrated into the Habitats Directive 92/43/EEC and the Natura 2000 Network (CitationBeato Bergua et al., 2017; CitationDalle Fratte et al., 2019; CitationEvans & Arvela, 2011; CitationGianguzzi et al., 2016; CitationViciani et al., 2016; CitationViciani et al., 2017, Citation2018). In this sense, vegetation mapping is indispensable in all eco-geographical analysis for nature conservation (CitationČernecký et al., 2020; CitationFreudenberger et al., 2013; CitationFührer, 2000; CitationIbarra & Yetano, 1998; CitationMaxted et al., 1997; CitationPanareda, 2000).
It is worth noting that the mapping of vegetation owes a great deal to the French biogeographer Marcel-Henri Gaussen (CitationKüchler, 1967). Among his contributions is the rational use of the color scheme; Gaussen gave an ecological-climatic significance to this by mixing the basic colors according to the combination of the environmental variables that determine the plant formations represented (CitationGaussen, 1936, Citation1947, Citation1958). Despite the age of his cartographic proposal, it is still very useful and in use, and is questioned only by the chromatic nuances that translate the dynamics of vegetation in relation to a given climatic situation, such as a finalist conception (CitationAlexandre & Génin, 2011; CitationRodríguez & Castañón, 2016).
The mapping of vegetation in Spain dates back to the map ‘Bodenund Vegetatonsverh-ltnisse der Iberischen Halbinsel’ by Willkomm of 1852 and the ‘Bosquejos dasográficos’ of the Comisión del Mapa Forestal de España created in 1868 (CitationBarrera, 2017). Since then, numerous maps have been published at the state level, both of real vegetation and of vegetation series (CitationRivas-Martínez, 1987), the forest maps of the national inventories being of special note (see, for example, CitationMAGRAMA, 2012 and CitationMAPAMA, 2018). Similarly, the production of regional maps at 1:50,000, 1:25,000, and even 1:10,000 scale, has been extensive. In the case of Asturias, the work of the CitationINDUROT (1994); its contribution has not yet been improved upon 25 years later, despite the need for precise and updated vegetation mapping because of the momentous changes that are taking place in the Asturian Massif (CitationBeato Bergua et al., 2017; CitationBeato Bergua et al., 2019b). The aim of this article, therefore, is to present the mapping of the vegetation of the Sierra del Aramo, an Asturian mountain range. Without coming under any figure of protection, the Aramo offers extensive natural and cultural richness, but is currently subject to important socioeconomic and landscape changes that need to be investigated.
2. Study area
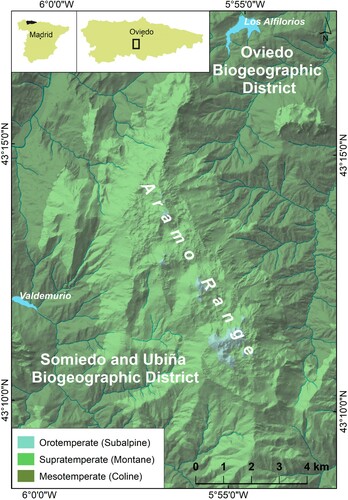
The Sierra del Aramo is situated at the intersection of the 43°16′N parallel and the 5°53′W meridian, that is, in the Asturian Central Massif of the Cantabrian Mountains (NW Spain), just 20 km southwest of Oviedo, the Asturian capital (). It is a mountain of medium altitude (the highest summit is the Gamoniterio, 1791 m a.s.l.), and is formed by a limestone ridge above the Trubia and Caudal river basins, which have been carved out shales, sandstones, limestones and coal strata. The verticalized sheets of Namurian limestones are being thrust over the Asturian Central Coal Basin, which explains the topographical and ecological differences. The Sierra del Aramo is crowned by a karst platform over 1300 m a.s.l. whereas the Aramo hillsides are steep and present rectilinear slopes between rocky escarpments and debris deposits (CitationBeato Bergua et al., 2019c).
Figure 1. Location of the study area.
In general, the analyzed area experiences Atlantic conditions with annual rainfall between 1100 and 1500 mm (around 2000 mm on the summits) and mild temperatures (between 6 and 13°C). According to CitationBeato Bergua (2018) and the Köppen classification there are four types of climates, Cfsb2 (properly temperate and rainy climate), Cfsb3 (transition to fresh climate), Cfsc (properly fresh climate), and Dfsc (typical cold mountain climate), as we ascend from the mild lower elevations to the climatic rigors of the summit area. Based on the bioclimatic classification of CitationRivas-Martínez et al. (2011), we can frame the Sierra del Aramo in the Temperate Oceanic Bioclimate, which in some low areas is presented with Submediterranean Variant. In addition, several bioclimatic belts can be distinguished, with specific plant coverings, depending on the changes in altitude of the thermoclimatic and ombroclimatic values. In terms of temperatures, the Sierra del Aramo displays three thermotypes which are determined from the thermicity index (It) and the positive annual temperature (Tp) (CitationRivas-Martínez, 2005) ().
Table 1. Thermotypes existing in the Sierra del Aramo. *Itc: compensated thermicity index. Based on CitationDíaz González (2015).
As regards humidity, the Sierra del Aramo has three differentiated ombrotypes (), which are obtained from the annual ombrothermic index (Io) (CitationDíaz González, 2015; CitationDíaz González & Fernández Prieto, 1994; CitationRivas-Martínez, 2005). A possible fourth, ultrahyperhumid ombrotype has been left out because of the lack of local data.
Table 2. Ombrotypes in the Sierra del Aramo. Based on CitationDíaz González (2015).
According to the units proposed by CitationRivas-Martínez et al. (2014), biogeographically the Sierra del Aramo is in the European Atlantic Province of the Atlantic-Central European Subregion. In particular, it is part of the two sub-provinces in which the Principality of Asturias is divided: although the northern and eastern slopes of the Aramo belong to the Oviedo District of the Cantabrian Atlantic Subprovince (Galician-Asturian Sector), the southern and the western slopes, together with the culminating platform, correspond to the Somiedo and Ubiña District of the Oro-Cantabrian Subprovince (Picoeuropean-Ubiniese Sector). Thus, in this area there are 24 potential series of vegetation (CitationDíaz González, 2009), which gives us a first idea of the plant richness that the Aramo can house. Specifically, potential plant formations are Polysticho setiferi-Fraxinetum excelsioris (Tüxen & Oberdorfer 1958) Rivas-Martínez ex C. Navarro 1982, Linario triornithophorae-Quercetum pyrenaicae Rivas-Martínez et al. 1984, Blechno spicant-Quercetum roboris Tüxen & Oberdorfer 1958, Carici sylvaticae-Fagetum sylvaticae (Rivas-Martinez 1965) C. Navarro 1982, Blechno spicanti-Fagetum sylvaticae (Tüxen & Oberdorfer 1958) Rivas-Martínez 1963, Hyperico androsaemi-Alnetum glutinosae (Br.-Bl. 1967) Rivas-Martínez in Loidi 1983, Festuco giganteae-Fraxinetum excelsioris F. Prieto & A. Bueno ass. nova, Salicetum angustifolio-albae T.E. Díaz & F. Prieto ass. nova, Daphno cantabricae-Arctostaphylo uvae-ursi Rivas-Martínez, Izco & Costa 1971, Cephalanthero longifoliae-Quercetum rotundifoliae Rivas-Martínez, T.E. Díaz, F. Prieto, Loidi & Penas 1984, Helleboro occidentalis-Tilietum platyphyllii (Prieto & Vázquez 1987) F. Prieto & Vázquez 2009, Luzulo henriquesii-Betuletum celtibericae Rivas-Martínez 1964 nom. mut. (art. 45) (addenda), Saniculo europaeae-Ilicetum aquifolii García-Baq. & Rivas-Martínez in Rivas-Martínez & col. 2011. However, the transformation of the plant cover from at least the Bronze Age (CitationBeato Bergua et al., 2019a) and the variety of local geo-ecological environments, explain the actual diversity of plant formations, many of which are susceptible to being valued and conserved as habitats of Community interest within the European Union (CitationBeato Bergua et al., 2017).
3. Materials and methods
For the characterization of the different plant formations present in the Sierra del Aramo we have used bibliographic sources relating to the framework of study itself (CitationBeato Bergua, 2018; CitationCortina, 2017; CitationMayor, 1996; CitationNavarro, 1974) and to the Asturian region (CitationDíaz González, 2014, Citation2015; CitationDíaz González & Fernández Prieto, 1988, Citation1994; CitationDíaz González & Vázquez, 2004). The information obtained during the field work carried out between 2010 and 2020 has also been essential, since it has allowed us to map the plant formations and make a biogeographic map (CitationBertrand, 1966; CitationFidalgo, 1997; CitationPanareda, 2000). Specifically, the methodology has been based on the identification and delimitation of plant formations by means of numerous transects, in which GPS data on location and delimitation were taken. In these ten years, more than a hundred inventories have been carried out trying to know all the plant formations and geoecological environments. The floristic composition and physonomic structure of the formations were determined according to phytosociological criteria of abundance-dominance of the species (CitationArozena & Molina, 2000; CitationBraun-Blanquet, 1979; CitationFidalgo, 1997). In addition, we have used the cartographic information and the reports of the Thematic Vegetation Map of the Principality of Asturias (CitationINDUROT, 1994), the Forest Map of Spain of the Fourth National Forest Inventory (CitationMAGRAMA, 2012), the latest Forest Map (CitationMAPAMA, 2018), and the maps of CitationNavarro (1974), CitationBeato Bergua et al. (2017) and CitationBeato Bergua (2018). Finally, the spatial boundaries of the different elements that make up the plant mosaic have been corrected and specified via the photointerpretation of the PNOA orthophoto (the Aerial Orthophoto National Program taken from 2017 have been used in ECW format through digital screen), using the computer software Arcgis 10.1, and according to criteria for color, texture, location and dynamics.
The construction of the map of the Sierra del Aramo has been made over the digital data of the Spanish National Geographic Institute (IGN) (http://www.ign.es/). We have used the National Topographic Base at 1:25,000, vector format SHAPEFILE (.shp). The indexing of georeferenced data was made with ESRI ArcMap10.1 with a Universal Transverse Mercator (UTM) projection (Zone 30) and the European Terrestrial Reference System 1989 Datum (ETRS89); and the graphic project was performed with ADOBE Illustrator CS6. The selection of colors has been made under the inspiration of the Gaussen method: blue indicates moisture; yellow, drought; red, heat; violet and pink, intense luminosity, etc. (CitationRodríguez & Castañón, 2016). All plant formations are presented in a smooth color except those originating from a plantation; in these cases, a black horizontal stripe pattern has been superimposed. The location map of the study area is made with the Digital Terrain Model with 5-meter grid spacing, raster format ASCII (.asc) ESRI array, of the Spanish National Geographic Institute.
4. Cartographic representation
To obtain an adequate visualization of the cartographic units from the geographical point of view, 28 types of vegetation cover have been selected (Main Map). These classes are defined in the following sections (22) of this paper. shows the connections between the map and the phytosociological units.
Table 3. Relationship between each paper section, phytosociological units and map classes.
4.1. Fagus sylvatica Woodland (Beechwoods)
These develop on carbonated and siliceous substrates, where formations can be found corresponding to the series Carici sylvaticae-Fagetum sylvaticae S. and Blechno spicanti-Fagetum sylvaticae S., respectively. The tree cover of the beechwoods, which are demanding of rich soils, can become monospecific, although it is more common for beech to concur with other trees such as Quercus petraea, Fraxinus excelsior, Acer pseudoplatanus and Taxus baccata, accompanied by shrubs, sometimes tree-like, such as Corylus avellana, Crataegus monogyna,, Ilex aquifolium and Sorbus aria. Polistichum setiferum, Dryopteris filix-mas, Anemone nemorosa, Helleborous viridis, Daphne laureola, Carex sylvatica, Galium odoratum, Melica uniflora and Mercurialis perennis are abundant ().
Figure 2. (A) Eutrophic Pedunculate Oakwoods and Beechwoods in Morcín. (B) Braña Vallongo on the Aramo's summit platform: (a) Mixed forest with a dominance of yews; (b) Heath-gorse formation; (c) Hazelnut-dominated shrub formation; (d) Basophilous mesophytic grassland.

Likewise, acidophilic beechwoods are often mixed forests (Fagus sylvatica, Quercus petraea, Betula celtiberica, Taxus baccata, Corylus avellana, Ilex aquifolium and Sorbus aria), although the arboreal and shrub strata are poorer in species than in eutrophic forests. In addition, the sub-shrub stratum is composed of bushes of Erica arborea bushes and ferns (Dryopteris affinis, Dryopteris dilatata) that also appear in the herbaceous (Blechnum spicant) together with Luzula sylvatica subsp. henriquesii, Deschampsia flexuosa, Saxifraga spathularis, Vaccinium myrtillus and Euphorbia hyberna.
4.2. Mixed forests with a dominance of yews
Two interesting forest patches have been mapped on the Aramo's summit platform with an abundance of Taxus baccata. The southernmost is related to the series of eutrophic beech trees (Carici sylvaticae-Fagetum sylvaticae) because of the profusion of beech and holly. However, the other (in the central area of the summit platform) does not present any specimens of this syntaxa and stands out for its biodiversity (Quercus petraea, Fraxinus excelsior, Acer pseudoplatanus, Tilia platyphyllos and Tilia cordata) and the domain of Taxus baccata in some sectors. It has not been possible to assign this second forest patch to any of the phytosociological associations proposed to date (CitationBeato Bergua et al., 2019a).
4.3. Quercus petraea Woodland (Sessile Oakwoods)
The oakwoods of Quercus petraea develops in the middle mountain in the Somiedo and Ubiña biogeographic districts. Quercus petraea coexists with lime-trees on the best nourished soils (Helleboro occidentalis-Tilietum platyphyllii) and with birch and beech in the poorest (Luzulo henriquesii-Quercetum petraeae). The lime-tree forests (Tilia platyphyllos and Tilia cordata) are actually mixed forests with sessile oak, maples, mountain elms, and even beech, with very rich and diverse lower strata. On the other hand, the tree cover of the acidophilic forest is basically composed of Quercus petraea and Betula celtiberica, with some shrubs of Aucuparia Sorbus, Ilex aquifolium, Corylus avellana and Erica arborea. This type of orocantabric nemoral formation also forms mixed communities with Quercus pyrenaica in the sunnier areas.
4.4. Quercus pyrenaica Woodlands (Oro-Cantabrian Pyrenean Oakwoods)
Quercus pyrenaica is mixed with Castanea sativa on the coline belt in mixed formations with poor undergrowth with Polistichum setiferum, Pteridium aquilinum, Rubus sp.pl, Rosa sp.pl. and Lonicera periclymenum. However, as we ascend the formations are more monospecific (Linario triornithophorae-Quercetum pyrenaicae) or mix with sessile oak in the wettest areas and with holm-Oaks in the drier ones. The species reaches altitudes exceeding 1200 m a.s.l. on the sunny slopes with stocky, twisted trunks, and mixed in with other tree species such as Frangula alnus, Pyrus cordata, Ilex aquifolium, Pteridium aquilinum, Erica australis subsp. aragonensis, Erica arborea and Cytisus sp.pl.
4.5. Quercus robur Woodland (Pedunculate Oakwoods)
Quercus robur extends throughout the coline belt of the Oviedese District, usually in deciduous mixed formations. Worth noting is the presence of eutrophic series with maple and ash (Polysticho setiferi-Fraxinetum excelsioris), and oligotrophic series with birch (Blechno spicant-Quercetum roboris). The acidophilic series is dominated by Quercus robur, Betula celtiberica, Castanea sativa and Laurus nobilis in the lower areas, and is enriched with Fagus sylvatica and Quercus pyrenaica at higher altitudes. The eutrophic, pedunculate oakwood is more diverse in species (Quercus robur, Fraxinus excelsior, Acer pseudoplatanus, Ulmus glabra, Tilia platyphyllos, Fagus sylvatica) and presents an undergrowth that is composed by Laurus nobilis, Euonymus europaeus, Cornus sanguinea, Corylus avellana, Ilex aquifolium, Crataegus monogyna and Prunus spinosa.
4.6. Betula celtiberica Woodland (Birchwoods)
Although the young forest of Betula pubescens subsp. celtiberica or Betula alba enjoys a wide distribution, birchwoods (Luzulo henriquesii-Betuletum celtibericae) are scarcer and are limited to certain borders of the transition from scrub to forest. These are formations that are not excessively tall and participated by Fagus sylvatica, Quercus petraea, Sorbus aucuparia, Sorbus aria, Erica arborea, etc.
4.7. Riparian Woodlands (Alder, Ash and Willow Woods)
These formations develop at the bottom of the valleys and gullies where herbaceous strata with fontinal communities of Cardamine raphanifolia and Chrysosplenium oppositifolium are common together with Stellaria alsine, Cardamine flexuosa, Chaerophyllum hirsutum, Apium nodiflorum and Equisetum palustre. The riparian Ash woodlands are made up of Fraxinus excelsior, Acer pseudoplatanus, Ulmus glabra and Salix atrocinerea, and generate authentic galleries. The riverbank forests present various types of willow (Salix sp.pl.), usually bushy and arborescent in size in the areas closest to the river course and accompanied by Alnus glutinosa and Populus nigra in the lower levels. Salix fragilis, Salix eleagnos subsp. angustifolia and Salix caprea abound in the lower strata. Undoubtedly, it is a plant community of great biodiversity. Likewise, Alnus glutinosa forms small alder woods corresponding to the eastern Cantabrian riparian woodland series Hyperico androsaemi-Alnetum glutinosae S., one of the most biodiverse plant formations in the region (Fraxinus excelsior, Betula celtiberica, Acer pseudoplatanus, Ulmus glabra, Salix alba, Corylus avellana, Laurus nobilis, Sambucus nigra, Frangula alnus, Cornus sanguínea, Hypericum androsaemum, Saxifraga hirsuta, Circaea lutetiana, Silene dioica, Festuca gigantea, Hedera helix, Dryopteris affinis, etc.).
4.8. Holm oak wood lands
The holm oak woodlands are relict forests in the Cantabrian mountains (this general assertion has been questioned by CitationLozano et al., 2002), where to a large extent they are composed of Mediterranean-type vegetation, despite the presence of Atlantic taxa. Quercus ilex can be accompanied by Rhamnus alaternus, Arbutus unedo, Ruscus aculeatus, Asplenium adiantum-nigrum, Osyris alba, Amelanchier ovalis, Rubia peregrina, Jasminium fruticans and Smilax aspera. These communities are situated on carbonate rocks in sunny locations of the coline belt in the Oviedese District. In contrast, the Oro-Cantabrian holm oak woodlands of Quercus rotundifolia (Cephalanthero longifoliae-Quercetum rotundifoliae) grow on calcareous, sunny slopes in the Somiedo and the Ubiña biogeographic districts ().
Figure 3. (A) Alder Woods in Quirós valley. (B) Pena Tene and Bermiego countryside (Quirós): (a) Casmophitic vegetation on the Sierra de Las Carangas limestone outcrops; (b) Oro-Cantabrian holm oak woodlands of Quercus rotundifolia; (c) Pulvinate Scrublands; (d) Mixed forests and meadows; (e) Oligotrophic Pyrenean Oakwoods; (f) Fern fields.

4.9. Ilex aquifolium woodlands
The holly woods are located mainly in the montane belt; they are low density and do not reach tree height. Ilex aquifolium colonizes abandoned meadows alongside Crataegus monogyna and Corylus avellana, and, depending on the wealth or poverty of bases in the soil (Saniculo europaeae-Ilicetum aquifolii), in the presence of Sorbus aucuparia, Betula alba, Fraxinus excelsior or Taxus baccata. Either way, the herbaceous stratum is very poor and is made up of nemoral species such as Melica uniflora, Mercurialis perennis and Helleborus viridis subsp. occidentalis.
4.10. Chestnut Woodlands (Castanea sativa-dominated forests)
The extension of Castanea sativa is very much worthy of note since it occupies much of the coline belt of the Sierra del Aramo and is favored by the use of its fruit and wood (). The density of the treetops generates a dark, humid environment similar to that of beechwoods, in which certain species of ferns (Polistichum setiferum, Dryopteris affinis Dryopteris dilatata), brambles, climbing plants, herbaceous plants and mosses barely develop. The chestnut woodlands may be dotted with Tilia platyphyllos, Quercus robur, Corylus avellana, Betula alba, Fraxinus excelsior, Acer pseudoplatanus, etc., and can even constitute mixed, nemoral formations that enjoy great naturalness and biological richness despite originally being plantations.
Figure 4. (A) A sector of the eastern slope of the Aramo Range: (a) Fagus sylvatica Woodland of La Matona, with an abundance of Taxus baccata; (b) Montane grasslands; (c) Fern fields and Heath-gorse formations; (d) Meadows; (e) Chestnut Woodlands; (f) Riparian Woodlands. (B) Castanea sativa-dominated forests and plantations in Riosa.

4.11. Conifer plantations
Conifer plantations are very scarce. The plantations of Pinus radiata in some plots in Riosa and Quirós are of note. They are practically monospecific formations with a very poor floristic composition of species typical of anthropized spaces. On the southern slope of the Aramo there are also some experimental plots with Pseudotsuga menziesii, Chamaecyparis lawsoniana and Larix sp.pl.
4.12. Other tree plantations
Leaving aside the chestnut woodlands, forest repopulation is not extensive in the Sierra del Aramo. However, within the study area there are small mosaics of eucalyptus (Eucaliptus globulus) as well as poplar and hardwood leafy plantations.
4.13. Hazelnut-dominated shrub formations
Corylus avellana develops inside all types of forests and colonizes screes and other inactive deposits. In the sunniest areas within the coline belt it grows alongside Laurus nobilis, Crataegus monogyna or Fraxinus excelsior and with Rhamnus alaternus, Ligustrum vulgare, Arbutus unedo and Phillyrea latifolia in xeric biotopes. In soils rich in bases the hazel grow up alongside Crataegus monogyna, Rosa sp.pl. and Prunus spinosa. However, the large extent of the plant formations with Rhamnus alpina, Berberis vulgaris subsp. cantabrica and Ribes alpinum on the Aramo summit platform (spiny shrublands from the association Pruno spinosae-Berberidetum cantabricae) is of note. These are very interesting communities, as are those in which the hazel coexists with Juniperus alpina, Daphne laureola subsp. cantabrica, Genista hispanica subsp. occidentalis and Taxus baccata (Oro-Cantabrian subalpine creeping juniper woods with bearberry and spurge-laurel, Daphno cantabricae-Arctostaphyletum uvae-ursi).
4.14. Heath-gorse formations
The intensity of anthropic use and its subsequent cessation has favored the extension of heath-gorse formations on oligotrophic soils. In particular, in their facies with Ulex europaeus heath-gorse formations dominate on the coline belt of the northern area (Ulici europaei-Ericetum vagantis), and in that of Ulex galii, for all mountain grass areas of the intermediate stations (Daboecio cantabricae-Ulicetum gallii). Among these high-density, low-height formations, that can sometimes exceed two meters in height, Ulex sp.pl., Erica vagans, Daboecia cantabrica, Erica cinerea, and Erica ciliaris, Calluna vulgaris, etc. are especially relevant.
4.15. Pulvinate Scrublands
These are made up of low, padded bushes in open formations dominated by Genista hispanica subsp. occidentalis. We can distinguish between basophilous broom formations on the coline and montane belts with or without Ulex europaeus (with Helianthemum nummularium, Lithodora diffusa, Teucrium pyrenaicum, Brachypodium pinnatum subsp. rupestre, Erica vagans, etc.), and a third type on the summit platform characterized by the Cantabrian endemism Genista legionensis, with Globularia nudicalis, Thymelaea ruizii and Euphorbia flavicoma subsp. occidentalis (Lithodoro diffusae-Genistetum legionensis). The Sierra del Aramo is the westernmost location of Genista legionensis ().
Figure 5. (A) Limestone grasslands on cacuminal skeletal soils with Genista hispanica subsp. occidentalis communities. (B) Genista legionensis patch.

4.16. Fern fields
Pteridium aquilinum easily colonizes meadows and pastures from the coline to the montane belt on both acidic and basic soils. It appears in extensive almost monospecific formations, or in mixed formations with Rubus sp.pl., Cytisus scoparius, Cytisus cantabricus, Erica arborea, Erica vagans, Daboecia cantabrica, Ulex europaeus, Ulex galii, Genista hispanica subsp. occidentalis, etc.
4.17. Broom scrublands
These scrub communities are limited to few sectors of oligotrophic soils in the Quirós valley at altitudes between 500 and 800 m a.s.l., and are characterized by Genista florida subsp. polygaliphylla, Cytisus cantabricus, Erica arborea and Pteridium aquilinum. Cytisus cantabricus also appears in the highest areas of the Aramo, and even dominates some high-mountain limestone outcrops.
4.18. Fruit tree plantations and agricultural crops
In this section we have included some small plots dedicated to agriculture in the valley bottoms, as well as other plantations of shrubs and fruit trees in the hills (apples, red fruits, etc.). The scarcity of flat areas, the primary orientation of production towards livestock, and the depopulation of the Asturian rural environment, explain the limited area given over to plantations and crops.
4.19. Grasslands
Different types of herbaceous formations can be distinguished starting at around 1000–1100 m a.s.l. – mountain pastures (mesophilous and hygrophilous), calcareous grasslands, perennial Nardus grasslands, and psicroxerophitic basophilus grasslands in the culminating platform. More specifically, we have: a) discontinued limestone grasslands on cacuminal skeletal soils with summer dryness dominated by Festuca burnatii and Erodium glandulosum; b) basophilous mesophytic grasslands on deep soils above 1200 m a.s.l. (Bromus erectus, Helianthemum nummularium, Brachypodium pinnatum subsp. rupestris, etc.); c) oligotrophic grassland of Jasione laevis and Galium saxatile on washed lime-related substrates above 1200 m a.s.l.; d) perennial nitrophilous grasslands in the high mountain altered by cattle (Chenopodium bonus-henricus, Senecio duriaei, Urtica dioica, Malva sylvestris, etc.).
4.20. Meadows
The herbaceous formations managed for mowing and cattle grazing were one of the largest formations on the basal slopes and valleys of the Aramo until the cessation of agro-livestock activity (). The meadows surfaces of this type present numerous species of grasses, herbaceous species, and legumes. However, they are now dominated by certain forage species such as ‘vallico’ (Lolium perenne, Lolium multiflorum), and monospecificity is prevalent. According to CitationNavarro (1974) and CitationMayor (1996) the following types of meadows can be found: a) Mesophilous meadows of Linum bienne and Cynosurus cristatus in the coline belt; b) Cantabrian mesophilous hay meadows dominated by Malva moschata and Arrhenatherum elatius subsp. bulbosum on wet eutrophic soils in the transition from the coline to the montane belt; c) pastures on the dry eutrophic and skeletal soils of the coline belt (Saxifraga tridactylites, Minuartia hybrida, Euphorbia exigua, etc.); d) grazing meadows, wet and nitrophic altered coline belt areas (Lolium perenne, Plantago major, Polygonum aviculare, Poa annua, Potentilla reptans, etc.).
Figure 6. (A) Cytisus cantabricus communities and casmophitic vegetation on the Aramo summit platform (Llagu Robles). (B) Meadows in Dosango (Quirós).

4.21. Casmophitic vegetation
There are ample areas of calcareous rocks in the Sierra del Aramo, especially in the highest areas where limestones appear practically devoid of vegetation, except in some sectors that are colonized by dispersed spiny shrublands and subalpine creeping juniper woods. However, even on the barest rocks, plants develop into fissure communities of great interest. In fact, the rupicolous vegetation has a fairly wide floristic diversity in keeping with the geoecological variability of its locations and the wealth in species, with a large number of endemisms. These plant formations correspond to the alliance Saxifragion trifurcato-canaliculatae, specifically, to the association Crepido-Erinetum (Erinus alpinus, Crepis albida subsp. asturica, Saxifraga canaliculata, Saxifraga trifurcata, Asplenium trichomanes, etc.).
4.22. Scree vegetation
At the foot of the limestone walls there are numerous screes due to pleistocene periglacial phenomena and gravity. Many of them, now inactive, are being colonized by shrub formations. However, in the pioneering stage the vegetation that protagonizes this phenomenon in the Cantabrian mountain corresponds to the species of the Linarion filicaulis alliance. Arabis alpina, Iberis pruitii, Rumex scutatus, Vincetoxicum hirundinaria, Geranium columbinum, Digitalis parviflora and Galeopsis angustifolia have been documented in the Sierra del Aramo.
5. Conclusions
This article describes the plant formations and their mapping for the Sierra del Aramo, whose wealth of biodiversity is outstanding. The map has a legend of 34 classes, of which 28 correspond to plant formations developed over three biogeographical belts and, within these, 4 are tree and crop plantations. These include habitats of Community interest for the European Union and one fundamental issue: the passage between two subprovinces and contrasting biogeographic districts (the Oviedese District of the Cantabrian Atlantic Subprovince and the Somiedo and Ubiña District of the Oro-Cantabrian Subprovince). In addition, the Sierra del Aramo presents other singularities such as the plant formations described earlier in which Taxus baccata participates, the importance of the extent of the pastures despite the process of expansion of the scrub (with a cover that is also outstanding in its different facies), etc. All this constitutes a natural and cultural heritage which needs to be mapped, given that vegetation maps must contribute both to territorial management and to the future monitoring of the evolution of the different plant covers in relation to socio-economic changes.
Software
The digital cartographic base and the digitalization of the vegetation mapping used in the preparation of this map were performed with ESRI ArcMap10.1. The final design was carried out with the graphic program ADOBE Illustrator CS6.
ARAMO_vegetation_MAP2021_3.pdf
Download PDF (2.7 MB)Acknowledgements
We want to express our gratitude for their collaboration to carry out this work to Raquel Cunill Artigas, José Manuel Berdasco Rodríguez, Héctor Rato Martín, Adriano Álvarez Méndez and Robin Walker.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Related Research Data
References
- Alexandre, F., & Génin, A. (2011). Géographie de la végétation terrestre. Modèles hérités, perspectives, concepts et méthodes. Armand Colin. http://www.rsehn.es/index.php?d=publicaciones&num=66&w=387&ft=1
- Arozena, M. E., & Molina, P. (2000). Estructura de la vegetación. In G. Meaza (Dir.), Metodología y práctica de la Biogeografía (pp. 77–147). Serbal.
- Barrera, I. (2017). La vegetación en los mapas. Memorias de la Real Sociedad Española de Historia Natural, 2ª ép., 14, 317–342.
- Beato Bergua, S. (2018). El patrimonio natural de la Sierra del Aramo (Montaña Central Asturiana) y la evolución de su paisaje. (Unpublished PhD Thesis). University of Oviedo, 596 pp.
- Beato Bergua, S., Marino Alfonso, J. L., & Poblete Piedrabuena, M. Á. (2017). El paisaje vegetal y los hábitats forestales de interés comunitario en la Montaña Central Asturiana. Cuadernos Geográficos, 56(1), 26–52. https://revistaseug.ugr.es/index.php/cuadgeo/article/view/4834
- Beato Bergua, S., Poblete Piedrabuena, M. Á., & Cunill Artigas, R. (2019a). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81(2772), 1–30. https://doi.org/https://doi.org/10.21138/bage.2772
- Beato Bergua, S., Poblete Piedrabuena, M. Á., & Marino Alfonso, J. L. (2019b). La dinámica del paisaje en la Sierra del Aramo (Macizo Central Asturiano): procesos naturales y antrópicos. Pirineos, 174, e041. https://doi.org/https://doi.org/10.3989/pirineos.2019.174001
- Beato Bergua, S., Poblete Piedrabuena, M. Á., Rodríguez Pérez, C., & Marino, J. L. (2019c). Geomorphology of the Sierra del Aramo (Asturian Central Massif, Cantabrian Mountains, NW Spain). Journal of Maps, 15(2), 590–600. https://doi.org/https://doi.org/10.1080/17445647.2019.1646675
- Bertrand, C., & Bertrand, G. (1986). La végétation dans le géosystème. Phytogéographie des montagnes cantabriques centrales (Espagne). Revue Géographique des Pyrénées et du Sud-ouest, 57(3), 291–312. https://doi.org/https://doi.org/10.3406/rgpso.1986.4949
- Bertrand, G. (1964). Esquisse biogéographique de la Liébana (Massif Cantabrique, Espagne). La dynamique actuelle des paysages. Revue Géographique des Pyrénées et du Sud-Ouest, 35(3), 225–262. https://doi.org/https://doi.org/10.3406/rgpso.1964.4776
- Bertrand, G. (1966). Pour une étude géographique de la végétation. Revue Géographique des Pyrénées et du Sud-Ouest, T. XXXVII, Fasc. 2, 129–144. https://doi.org/https://doi.org/10.3406/rgpso.1966.4544
- Braun-Blanquet, J. (1979). Fitosociología. Bases para el estudio de las comunidades vegetales. Blume.
- Černecký, J., Gajdoš, P., Špulerová, J., Halada, L., Mederly, P., Ulrych, L., Ďuricová, V., Švajda, J., Černecká, L., Andráš, P., &, & Rybanič, R. (2020). Ecosystems in Slovakia. Journal of Maps, 16(2), 28–35. https://doi.org/https://doi.org/10.1080/17445647.2019.1689858
- Cortina, J. (2017). Flora medicinal y hábitats de la Sierra del Aramo (Asturias). (Unpublished Degree Thesis). University of Salamanca.
- Dalle Fratte, M., Brusa, G., & Cerabolini, B. E. L. (2019). A low-cost and repeatable procedure for modelling the regional distribution of Natura 2000 terrestrial hábitats. Journal of Maps, 15(2), 79–88. https://doi.org/https://doi.org/10.1080/17445647.2018.1546625
- Díaz González, T. E. (2009). Caracterización de los Distritos Biogeográficos del Principado de Asturias (Norte de España). In F. LLamas, y C. Acedo (Eds.), Botánica Pirenaico-Cantábrica en el siglo XXI (pp. 423–455). Área Publ. Univ. León.
- Díaz González, T. E. (2014). Mapas de vegetación de las series, geoseries y geopermaseries de España. 1:250.000: Asturias. Global Geobotany, 3, 1–34. https://editaefa.com/mostrarArticulo.php?num=3&articulo=163
- Díaz González, T. E. (2015). Guía para la identificación de los bosques, matorrales y series de vegetación (Vegetación Potencial) de Asturias mediante bioindicadores fitocenológicos. Boletín de Ciencias de la Naturaleza, 53, 5–94. http://www.ridea.org/catalogo/catalogo/item/boletin-de-ciencias-de-la-naturalezan-53
- Díaz González, T. E., & Fernández Prieto, J. A. (1988). Caracterización de las unidades fitogeográficas de Asturias. Monografías del Instituto Pirenaico de Ecología, 4, 517–528.
- Díaz González, T. E., & Fernández Prieto, J. A. (1994). El paisaje vegetal de Asturias. Itinera Geobotánica, 8, 5–242.
- Díaz González, T. E., & Vázquez, A. (2004). Guía de los bosques de Asturias. Ediciones Trea.
- Evans, D., & Arvela, M. (2011). Assessment and reporting under Article 17 of the Habitats Directive. Explanatory Notes & Guidelines for the period 2007-2012. European Commission, Brussels.
- Fidalgo, C. (1997). El trabajo de campo en Biogeografía. Didáctica geográfica, 2, 33–44.
- Freudenberger, L., Hobson, P., Schluck, M., Kreft, S., Vohland, K., Sommer, H., Reichle, S., Nowicki, C., Barthlott, W., & Ibisch, P. L. (2013). Nature conservation: Priority-setting needs a global change. Biodiversity and Conservation, 22(5), 1255–1281. https://doi.org/https://doi.org/10.1007/s10531-012-0428-6
- Führer, E. (2000). Forest functions, ecosystem stability and management. Forest Ecology and Management, 132(1), 29–38. https://doi.org/https://doi.org/10.1016/S0378-1127(00)00377-7
- Gaussen, H. (1936). Le choix des couleurs dans les cartes botaniques. Bulletin de la Société Botanique de France, 83(6-7), 474–480. https://doi.org/https://doi.org/10.1080/00378941.1936.10837343
- Gaussen, H. (1947). L'emploi des couleurs en cartographie. Comptes Rendus de L'Académie des Sciences, Paris, 224, 450–452.
- Gaussen, H. (1958). Integration of data by means of vegetation maps. Ninth Pacific Congress, 20, 67–74.
- Gianguzzi, L., Papini, F., & Cusimano, D. (2016). Phytosociological survey vegetation map of Sicily (Mediterranean region). Journal of Maps, 12(5), 845–851. https://doi.org/https://doi.org/10.1080/17445647.2015.1094969
- Ibarra, P. (1993a). Una propuesta metodológica para el estudio del paisaje integrado. Geographicalia, 30, 229–242.
- Ibarra, P. (1993b). Naturaleza y hombre en el sur del campo de Gibraltar: un análisis paisajístico integrado. Junta de Andalucía, Consejería de Cultura y Medio Ambiente, Agencia de Medio Ambiente.
- Ibarra, P., & Yetano, M. (1998). El estudio de la vegetación en geografía. Geographicalia, 26, 165–174.
- INDUROT. (1994). Cartografía temática ambiental. Mapa de vegetación. Escala 1:25.000. Consejería de Medio Ambiente, Ordenación del Territorio e Infraestructuras del Gobierno del Principado de Asturias.
- Küchler, A. W. (1967). Vegetation mapping. The Ronald Press Company.
- Lozano, P. J., Meaza, G., & Cadiñanos, J. A. (2002). Paleobiogeografía de la Reserva de la Biosfera de Urdaibai (Vizcaya). Boletín de la Asociación de Geógrafos Españoles, 34, 193–211. https://bage.age-geografia.es/ojs/index.php/bage/article/view/436/0
- MAGRAMA. (2012). Principado de Asturias. Cuarto Inventario Forestal Nacional. Dirección General de Desarrollo Rural y Política Forestal.
- MAPAMA (Dirección General de Desarrollo Rural). (2018). Mapa Forestal de España de máxima actualidad 1:25.000 (MFE25). Ministerio para la Transición Ecológica. https://www.miteco.gob.es/es/cartografia-y-sig/ide/descargas/biodiversidad/mfe.aspx
- Martínez de Pisón, E. (1983). Cultura y Ciencia del Paisaje. Agricultura y sociedad, 27, 9–32.
- Maxted, N., Ford-Lloyd, B. V., & Hawkes, J. G. (1997). Plant genetic conservation. Springer. https://doi.org/https://doi.org/10.1007/978-94-009-1437-7
- Mayor, M. (1996). Indicadores ecológicos y grupos socioecológicos en el Principado de Asturias (Sierra del Aramo). Universidad de Oviedo.
- Navarro, F. (1974). Estudio de la flora y vegetación de la sierra del Aramo y sus estribaciones (Asturias). (Unpublished PhD Thesis). University of Oviedo.
- Panareda, J.M. (2000). Cartografía y representación fitogeográfica. In G. Meaza (Dir.), Metodología y práctica de la Biogeografía (pp. 273–316). Ediciones del Serbal.
- Rivas-Martínez, S. (1987). Memoria del mapa de series de vegetación de España 1: 400.000. ICONA. Ministerio de Agricultura, Pesca y Alimentación.
- Rivas-Martínez, S. (2005). Avances en Geobotánica. Discurso de Apertura del Curso Académico de la Real Academia Nacional de Farmacia del año 2005. Madrid: Real Academia Nacional de Farmacia del año 2005.
- Rivas-Martínez, S., Penas, A., Díaz González, T. E., Costa, J. C., Pinto Gomes, C., Herrero, L., & del Río, S. (2014). Biogeography of Spain and Portugal. Update typology. Int J Geobot Res, 4(1), 1–64. http://editaefa.com/mostrarArticulo.php?articulo=166
- Rivas-Martínez, S., Rivas Sáenz, S., & Penas, A. (2011). Worldwide bioclimatic classification system. Global Geobotany, 1(1), 1–634. https://doi.org/https://doi.org/10.5616/gg110001
- Rodríguez, C., & Castañón, J. C. (2016). Modos de representación cartográfica de las unidades de paisaje: revisión y propuestas. Ería, 99-100-100 bis, 15–40. https://doi.org/https://doi.org/10.17811/er.99.2016.15-40
- Viciani, D., Dell'Olmo, L., Ferretti, G., Lazzaro, L., Lastrucci, L., & Foggi, B. (2016). Detailed Natura 2000 and CORINE biotopes habitat maps of the island of Elba (Tuscan Archipelago, Italy). Journal of Maps, 12(3), 492–502. https://doi.org/https://doi.org/10.1080/17445647.2015.1044040
- Viciani, D., Dell’Olmo, L., Foggi, B., Ferretti, G., Lastrucci, L., & Gennai, M. (2018). Natura 2000 habitat of Mt. Argentario promontory (southern Tuscany, Italy). Journal of Maps, 14(2), 447–454. https://doi.org/https://doi.org/10.1080/17445647.2018.1489903
- Viciani, D., Dell’Olmo, L., Vicenti, C., & Lastrucci, L. (2017). Natura 2000 protected habitats, Massaciuccoli Lake (northern Tuscany, Italy). Journal of Maps, 13(2), 219–226. https://doi.org/https://doi.org/10.1080/17445647.2017.1290557