Abstract
We present results of an international collaboration to survey American lobster Homarus americanus Milne-Edwards, 1837 nurseries in Atlantic Canada and the northeast United States from 2007 to 2009 under a standardized protocol involving two sampling methods, diver-based suction sampling and passive collectors. We surveyed young-of-year and older juveniles at 191 sampling sites over 39 sampling areas considerably expanding the known depth range and geographic limits of benthic recruitment. Young-of-year densities were strongly correlated in space with the abundance of older juveniles, signifying consistently strong settlement in the Gulf of Maine, lower Bay of Fundy, southwestern Nova Scotia and southern Gulf of St. Lawrence, and relatively weak settlement in southern New England, eastern coastal Nova Scotia and Newfoundland, a pattern consistent with commercial lobster harvests. Passive collectors elucidated bathymetric patterns of young-of-year recruitment in oceanographically contrasting regions. Although we observed young-of-year lobsters as deep as 80 m, they were most abundant above the thermocline in summer-stratified regions, such as the western Gulf of Maine and southern New England, and depth-wise differences were less extreme in thermally mixed waters of the eastern Gulf of Maine–Fundy region, a finding consistent with previous observations that postlarvae concentrate above the thermocline. Between the two samplers, we detected no sampling bias for young-of-year lobsters, although collectors may slightly under-represent older juveniles entering from the surrounding sea bed. Finally, we found that interactions between juvenile lobsters and suspected predators or competitors in collectors, such as crabs and fishes, are weak and unlikely to bias collector results.
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
Introduction
Early life-stage sampling is a key tool in population assessment and forecasting in marine ecological and fisheries science. Developing sampling methods to quantify newly settled cohorts in benthic population studies can be a powerful way of understanding processes operating before and after larval settlement that drive subsequent recruitment and population dynamics (Roughgarden et al. Citation1988; Pineda Citation2000; Balch & Scheibling Citation2000; Cowen et al. Citation2000; Navarrete et al. Citation2005). Studies of lobsters and crabs that track the fate of early benthic cohorts have provided critical insights into larval transport leading to benthic recruitment and the strength of inter- and intraspecific interactions influencing the fate of cohorts after settlement (Eggleston & Armstrong Citation1995; Pile et al. Citation1996; Incze et al. Citation1997; Moksnes & Wennhage Citation2001; Gardner et al. Citation2001; Briones-Fourzan et al. Citation2008). The western rock lobster, Panulirus cygnus George 1962, is a noteworthy example of a species for which annual monitoring of postlarval settlement has been an integral part of fishery management for more than four decades (Caputi et al. Citation1995). Progress has come much more slowly for other commercial species, however. In the case of the American lobster, Homarus americanus Milne-Edwards, 1837, one of the most important commercial species in the northwest Atlantic, early life-stage sampling started in the late 1980s and has yielded valuable information on early recruitment processes. To date, however, it has been difficult to assemble a picture of early benthic recruitment on the scale of the entire species’ range. New sampling tools and a wider international collaboration, however, are making larger-scale surveys possible.
The American Lobster Settlement Index is a monitoring programme of lobster nurseries in the northeast US and Atlantic Canada carried out by a collaboration of government marine resource agencies, academic institutions and the fishing industry (Wahle et al. Citation2010). Conducted once a year in the late summer or early autumn, sampling quantifies the abundance of young-of-year lobsters that have recruited during the summer postlarval settlement season, and all sizes of older juveniles residing in nurseries. Historically, monitoring has been conducted by diver-based suction sampling of shallow nurseries, which consist of naturally occurring cobble beds where young lobsters find plentiful shelter (Wahle & Steneck Citation1991; Incze & Wahle Citation1991). Monitoring by suction sampling began in 1989 in mid-coast Maine and has expanded to other study areas over time. However, it has been limited to the shallow waters of the coastal New England states in the US, and two locations in Atlantic Canada. Nursery populations in coastal New England have been reasonably well described, and these data have been used recently to validate spatial and temporal patterns of young-of-year recruitment (Xue et al. Citation2008; Wahle et al. Citation2009a; Incze et al. Citation2010). Although perhaps not as efficient as suction sampling at detecting the smallest lobsters, visual quadrat and transect surveys by divers are nonetheless effective means of quantifying older juveniles and inferring nursery locations (Bernstein & Campbell Citation1983; Hudon Citation1987; Steneck & Wilson Citation2001; Gendron & Sainte-Marie Citation2006). Still poorly understood, however, is the extent to which lobster postlarvae settle and recruit to other parts of the species’ geographic range, particularly to deep-water habitats beyond the reach of divers. As a result, population dynamic models that simulate regional production patterns would be forced to make untested assumptions about the numbers of lobsters recruiting to the benthos in deep water. Because the areas involved are so extensive, the difference in production estimates between those that assume no deep-water settlement and those that assume even a very low-density settlement could be dramatic.
In a cooperative research project with the US lobster fishing industry in 2005, vessel-deployed passive postlarval collectors, consisting of wire mesh trays filled with cobble, proved to be a viable alternative to suction sampling as means of quantifying lobster settlement (Wahle et al. Citation2009b). Collectors, therefore, provided the opportunity to evaluate settlement in locations that were either unsafe or impractical for divers. Renewed research support, in turn, provided the catalyst for an expanded collaboration between fishermen and scientists in the US and Canada beginning in 2007. The joint effort has greatly expanded the geographic and depth coverage by passive collectors and suction sampling.
Here we describe an international collaborative study spanning a large portion of the geographic range of the American lobster in Atlantic Canada and the northeast United States from 2007 to 2009. Our aim here is to describe patterns of young-of-year and older juvenile lobster abundance along the coast, and by depth, using the two quantitative tools currently in use within the collaborative programme: diver-based suction sampling and passive postlarval collectors. We also use this opportunity to build on our previous preliminary studies (Wahle et al. Citation2009b) comparing the performance and size-selectivity of the two methods, and evaluating species interactions in collectors that may bias abundance estimates.
Methods
Sampling methods and geographic coverage
Sampling for early benthic juveniles was conducted by two methods: diver-based suction, and passive collector-based sampling (). Suction sampling has been used to monitor lobster settlement since the late 1980s, and the monitoring protocol has been described in detail elsewhere (Wahle & Incze Citation1997). Briefly, the method involves a pair of divers working together to vacuum the contents of 0.5-m2 quadrats placed in cobble–boulder habitat. The quadrat is semi-enclosed with a mesh apron to impede escape of lobsters and crabs. The polyvinyl chloride pipe frame is sealed to be air-tight and buoyant and to give the quadrat's shape; and an apron hanging from the frame is weighted along the bottom with chain that conforms to the irregularities of the seabed. Divers removed rocks within the quadrat during the sampling process to expose hidden infauna. Samples were retained in a mesh bag secured to the top of the sampler; bags were changed between quadrats. Typically 12 0.5-m2 quadrats were sampled per site, although more were used in some locations to enhance detection of lobsters occurring at low density. Half-square-meter quadrats have been used in all study areas, except Beaver Harbour, New Brunswick, where 0.25-m2 quadrats have been sampled over the years. Selection of the smaller quadrat size there is based on balancing the needs for sample replication and diving factors given the extreme tidal range found within the Bay of Fundy. Sample bags are returned to the laboratory where they are sorted fresh or were frozen to be sorted later. Lobster carapace length, sex and number of claws are recorded. Associated fauna, such as crabs and fish, are also counted, measured and identified to species.
Figure 1. Two sampling methods for young-of-year and older juvenile lobsters and associated fauna: (a) suction sampling; (b) passive, vessel-deployed, postlarval collectors.

Passive, vessel-deployed postlarval collectors were recently developed as an alternative to suction sampling (Wahle et al. Citation2009b). Collectors were primarily designed to evaluate lobster settlement in locations that are either unsafe or impractical for diver sampling, although they have become the preferred sampling device, even in shallow water, in some regions. Details of their design and method of deployment are provided in Wahle et al. (Citation2009b). Collectors are made of 10-gauge vinyl-coated wire with a 38-mm (1.5 inch) mesh. They measure 61.0 cm×91.5 cm×15.0 cm deep, providing 0.56 m2 in floor area. A cover made of the same wire mesh retains the rocks and the free passage of settling postlarvae. Two oak runners are fastened to the bottom for additional rigidity and to aid sliding and stacking on the vessel's deck. The floor and walls are lined inside with a rugged 2-mm plastic mesh (PetMesh™) to retain lobsters, crabs and other organisms during retrieval. Collectors are filled with clean, rounded cobbles ranging in diameter from 10 to 15 cm acquired from a local excavator equipped to sort cobble for the desired size range. Collectors filled with rocks weigh approximately 80 kg. Each collector is fitted with a bridle to permit lifting in a horizontal position, which is important to retaining collections during retrieval. In previous pilot studies, we demonstrated that losses of organisms were negligible when the collectors were retrieved in this way (Wahle et al. Citation2009b). Once on deck, the wire mesh covers are removed from the collectors. The rocks are rinsed with seawater from a deck hose to rinse away sediment, and then are carefully removed to inspect for lobsters and other organisms. Occasionally, collectors become swamped with sand over the course of the season because of storms; these collectors are excluded from subsequent analysis. All lobsters, crabs and fishes are measured. Lobsters are further inspected to determine sex, and number of claws. Size definitions for young-of-year lobsters vary by study area because of regional differences in growth rate (; Wahle & Fogarty Citation2006). In the present study collectors were deployed progressively from south to north corresponding to the regional availability of lobster postlarvae. Thus deployments in Rhode Island were from late May to early September, and from early July to late October from the Gulf of Maine to Newfoundland.
Table I. Results of nested ANOVAs for depth, year and site effects on densities of young-of-year and older juvenile lobsters in collectors in four regions, illustrated in . The F-ratio for the fixed-effect factor depth was assessed using for denominator the mean-square of the random variable site [depth]. The year and year×depth factors were assessed using year×site [depth] as error term, whereas site [depth] and year×site [depth] were assessed on the model residuals. Significant p-values (p<0.05) indicated in bold. Young-of-year could not be analysed for Rhode Island because of insufficient number of settlers.
From 2007 to 2009 surveys by suction sampling and passive collector were conducted over a broad geographic area encompassing most of the range of the American lobster, from coastal southern New England to Newfoundland, as well as offshore locations on Platts Bank in the central Gulf of Maine and the western flank of Georges Bank on the southern New England shelf (). Replicate quadrats or collectors were nested within sites, which in turn were nested within study areas. By this design, quadrats within sites were separated by metres, sites within study areas were separated by approximately one-half to a few kilometres and study areas were separated by several tens of kilometres or more. In a number of cases, collector deployments and suction sampling were conducted at the same site, permitting side-by-side comparison of the samples. Over the three years, 191 sites were sampled in 39 study areas, although not all sites or study areas were sampled each year.
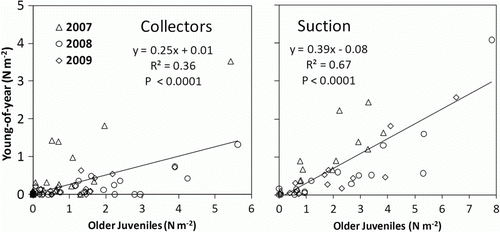
To determine whether there tended to be locations of consistently high or low settlement, we evaluated the strength of the relationship between young-of-year recruitment and older juvenile densities at the same sites in the same year. We conducted a correlation analysis of site means both for the suction sampling surveys and the collector data separately for each of the three years.
Within this broad geographic sampling program, the design requirements for certain analyses required selection of subsets of data, such as locations where both collector deployments and suction sampling were conducted at the same site. Specific study areas for which particular analyses apply are noted below.
Bathymetric patterns
Bathymetric patterns of young-of-year and older juveniles were evaluated during 2007 and 2008 at a subset of four study areas. A coherent study was designed to compare three study areas of contrasting oceanography off coastal New England, USA: eastern Maine (study area 21, see ; ), near the mouth of the Bay of Fundy; mid-coast Maine (26), centred in the Gulf of Maine; and Rhode Island (38), adjacent to southern New England's shelf waters. The three-area comparison includes a description of the thermal structure of the water column over the summer settlement season. An ancillary depth-wise comparison was made in Lobster Bay (27), southwestern Nova Scotia, Canada, over a smaller range of depths somewhat expanding the geographic coverage of this analysis, but with no matching thermal analysis. Sites used in these analyses are listed in .
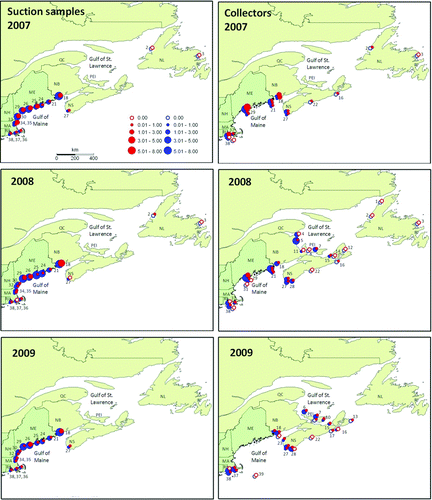
Figure 2. Spatial patterns of lobster density determined by suction sampling and collector. Average densities (N m−2) of young-of-year (red) and older juvenile lobsters (blue) found in suction samples (left), and collectors (right) deployed in 2007, 2008 and 2009. Symbols represent multi-site averages for each study area. Numbers denote study areas listed in .
In each of the three New England study areas (eastern Maine, mid-coast Maine, and Rhode Island) 100 collectors were divided among three depth strata (shallow=5–18 m; mid=25–45; deep=55–90 m), and then further divided among two sites within each depth stratum, with each site receiving between 14 and 18 collectors; sites in deeper strata receiving the higher number of collectors to offset potential losses. The geographic position of each collector was recorded at deployment. Eastern Maine is strongly influenced by tidal mixing of the Bay of Fundy, and is thermally homogeneous from surface to bottom during most of the year, with summer temperatures normally not exceeding 12°C. In contrast, mid-coast Maine becomes thermally stratified to a shallow depth during the warm season with surface temperatures reaching 16–18°C, but with temperatures at a depth of 50 m not exceeding 7°C. Coastal Rhode Island and the southern New England shelf represent a warmer regime with summer surface temperatures typically exceeding 20°C, and a deeper thermally stratified layer than forms in the Gulf of Maine. Temperature profiles were taken with a Sea-Bird (Bellevue, WA, USA) conductivity–temperature–depth (CTD) profiler at the time of deployment and retrieval. At least one collector in each depth stratum for each study area was equipped with a Hobo (Onset Computer, Inc., Bourne, MA, USA) temperature logger secured to the collector at the sea bed and the corresponding surface buoy. Loggers recorded temperature hourly.
In Lobster Bay, Nova Scotia, during the summers of 2007 and 2008, 193 collectors were set at four sites in the shallow stratum as defined above, and 80 collectors were set at two sites in the mid-depth stratum. No temperature data were recorded.
In a separate statistical analysis for each of the three regions in New England we analysed variation in density of settlers or juveniles using a mixed-model nested ANOVA, with year and depth stratum as fixed-effect factors, site [depth] (read site nested within depth) as random-effect factor and the year×depth and year×site [depth] interactions as fixed-effect factors. We derived the error terms for these different factors from first principles (Underwood Citation1997). Most importantly, the F-ratio for depth was assessed using the mean-square of the random variable site [depth] as the denominator the level of replication assumed in assessing this term was thus the different sites within a given depth and region, and not the different collectors. We elucidated significant depth and depth×year effects using Tukey-type multiple comparisons.
The fourth region, southwestern Nova Scotia, had to be analysed with a slightly modified model because it comprised an insufficient number of sites to support the full model (insufficient degrees of freedom). For this region, we removed the year×site [depth] factor from the model and tested the year and the year×site factors on the model residuals. However, and most importantly, the depth term was still tested on site [depth].
We ran all analyses on log(x+1)-transformed density values, as this was necessary in some cases to ensure the data satisfied the assumption of equal variances for main effects, as determined by a Levene's Test (Levene Citation1960). Statistical analyses were conducted with JMP computer program version 6 software.
Comparison of samplers
To assess the two sampling methods for evidence of sampling bias, we conducted two analyses to compare collector and suction-sampling data generated at the same sites. The Beaver Harbour suction-sampling data were excluded from this analysis because of the smaller quadrat area sampled there, only half the size of collectors. First, we used a paired t-test to compare pairs of site means obtained by the two methods with a hypothesized difference of zero if no sampling bias existed. Tests were conducted separately for young-of-year and older juveniles. Over the three years of sampling, we accumulated 25 pairs of independent site means rendering a statistical power exceeding 0.8 to resolve an effect size equal to 1 standard deviation of the group mean at α=0.05. Next, to more finely evaluate potential differences size selectivity, we compared size distributions of lobsters generated by the two methods with a Kolmogorov–Smirnov (KS) two-sample test, which tests the null hypothesis that two independent samples have been drawn from the same population (Conover Citation1971). The test statistic, D, is the maximal difference between cumulative frequency distributions of two samples. Size categories were 1 mm in width. We used the function lfclus in the R software package fishmethods; this statistically compares length frequencies from simple random cluster sampling, and uses randomization procedures to derive a null probability distribution (Nelson Citation2011).
Species associations in collectors
Because a diversity of species other than lobsters also recruit to collectors, we evaluated whether the frequency of collectors with young-of-year and older juvenile lobsters varied independently of those with crabs and fishes large enough to eat or compete with lobsters. For study areas with sufficiently large sample sizes, we used Chi-square contingency tables to evaluate the strength of the association between the frequency of lobsters in the two size categories and crabs>30 mm carapace width or carnivorous fishes>50 mm total length, sizes at which individuals are suspected to be large enough to pose a threat to early benthic lobsters. We reasoned that a negative interaction would be suggested if lobsters were found co-occurring with potential predators less often than would be expected by chance. A comparable assessment was not conducted with suction-sampling data because suction samplers do not capture fish as effectively as collectors (Wahle et al. Citation2009b).
Results
Geographic patterns
Mean densities of young-of-year and older juveniles are mapped in by study area (). The combination of suction sampling and collector data compiled here give the largest synoptic view of lobster young-of-year recruitment and older juveniles available to date. Densities of older juveniles, an aggregate of several age classes mostly between 1 and 3 years old, were generally higher than those of young-of-year lobsters. Densities of young-of-year lobsters were strongly correlated with older juveniles in the same study area, both in collectors and in suction samples, suggesting a consistent spatial pattern of recruitment from year to year (). Although juvenile densities were very sparse at the northern and offshore extremes of the species’ range, it is noteworthy that a single young-of-year lobster was found in Bonne Bay, Newfoundland, in collectors in 2007, and in suction samples in 2008, marking the northern extreme of young-of-year recruitment on record for the species. It is also noteworthy that our sampling forays with collectors to offshore areas such as Platts Bank in the Gulf of Maine in 2008 and the eastern flank of Georges Bank in 2009 produced no young-of-year or older juvenile lobsters ().
Figure 3. Correlation between young-of-year and older juvenile lobster densities at the same site from collectors and suction sampling. Each point represents a site average.
Bathymetric patterns
In general the number of young-of-year and older juveniles was depth-dependent, with the highest densities found in collectors deployed in the shallowest strata (; ). However, the magnitude and significance of the depth factor varied among regions, in a manner that generally mirrored the pattern of summer thermal stratification (; ). In thermally stratified mid-coast Maine, the depth effect was very pronounced and statistically significant for both young-of-year and older juveniles; settlers were significantly and markedly more abundant in shallow than in mid or deep strata, whereas juvenile abundance followed this pattern in 2007 and differed significantly among all three depth strata in 2008. Similarly, in thermally stratified Rhode Island, juveniles were markedly and significantly more abundant at the two shallower depth strata than the deeper one; young-of-year recruitment was relatively high in the shallow stratum in 2007, but it was otherwise extremely sparse, precluding statistical analysis of this group. In contrast, in the well-mixed regime of eastern Maine, near the Bay of Fundy, the effect of depth on lobster abundance was weaker and only bordered statistical significance for both young-of-year and juveniles, and southwestern Nova Scotia (also well mixed) it was weakest and clearly non-significant, at least as a main effect. Nonetheless, depth did have some effect on lobster settlement in Nova Scotia. Despite the narrower depth range sampled there, the significant depth×year interaction for young-of-year lobsters, inferred that larval settlement was significantly greater in shallow than mid depths in 2007 when settlement was relatively high, but did not differ between depths in 2008, when settlement was much lower (). Finally, it is noteworthy that these collector deployments produced the deepest record of young-of-year densities to date at 80 m, although we found it surprising that young-of-year and older juvenile lobsters were not detected in offshore deployments, such as Georges and Platt's Banks, even at intermediate depths ().
Figure 4. Bathymetric distributions of young-of-year and older juvenile lobsters sampled by collectors during 2007 and 2008 in eastern and mid-coast Maine, Nova Scotia and Rhode Island. Shallow = 5–18 m; mid = 25–45 m; deep = 55–90 m.
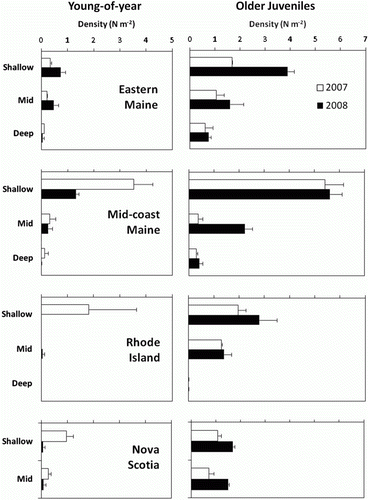
Figure 5. Temperature regimes in the three oceanographically contrasting study areas of New England where collectors were deployed in three depth strata and temperature records were taken (). (a) Temperature profiles taken 2007 and 2008 when collectors were retrieved at the end of the settlement season in each study area. (b) Temperature time series from temperature loggers fastened to collectors at different depths on the sea bed and at the surface in each study area during the 2007 sampling season.
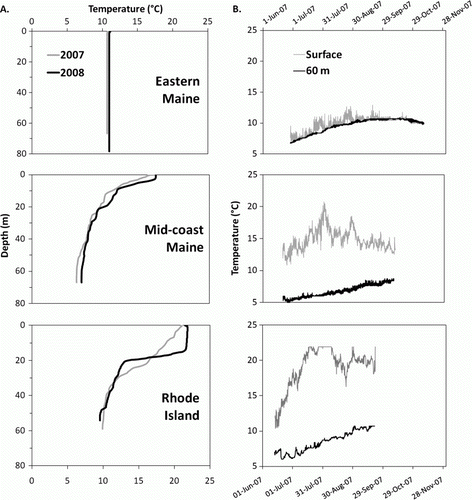
Table II. Results of Kolmogorov–Smirnov test to compare cumulative size frequency distribution of lobsters from collector and suction sampling by study area and for all study areas combined (see ). Separate analyses conducted for lobsters of all sizes and those≤25 mm carapace length for which under-sampling by collectors was suspected. Significant p-values (p<0.05) indicated in bold.
Comparing samplers
Among the 25 sites where collector and suction sampling were conducted at the same time, we found no significant difference between the two sampling methods with respect to the mean density of young-of-year lobster per square metre that would have settled directly from the plankton (collector: 0.76±0.99 SD, suction: 0.67±0.94 SD; t=2.06, p=0.32). Older juveniles, which immigrated to collectors from the surrounding sea bed, were significantly more abundant in collectors than in suction samples collected nearby (collector: 2.17±1.84 SD, suction: 1.66±1.74 SD; t=2.06, p=0.01). Although the overall density of older juveniles was greater in collectors than suction samples, there is evidence that collectors under-sample in a specific intermediate size range of older juveniles relative to suction sampling. For the pooled data from all study areas, the KS test detected a significant difference between the cumulative size frequencies of the two methods (); there tended to be relatively few lobsters in the<25 mm range than in suction samples, suggesting collectors under-sample lobsters in this range relative to suction sampling (). Whether we considered all sizes or only lobsters≤25 mm, the size distributions for the two sampling methods were statistically indistinguishable using the KS test; and only for eastern Maine did the test statistic approach significance (p=0.08). For the more heavily sampled mid-coast Maine and Lobster Bay, Nova Scotia study areas, visual inspection of the data suggested collectors under-sampled lobsters between 12 and 25 mm, but not those<12 mm that would have been more likely to have settled into collectors from the plankton than to have migrated in from the surrounding sea bed. Possible explanations for under-sampling of the 12–25 mm size are examined in the discussion.
Figure 6. Cumulative size distributions and fitted logistic curves for lobsters collected by suction sampling (triangles) and collectors (squares) in four study areas where both methods were used at the same sites. Statistical analysis in .
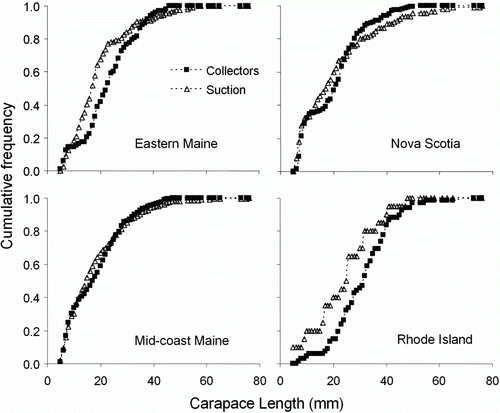
Table III. (a) Occurrence in collectors of fish and crab species considered potential predators or competitors in four study areas where they were evaluated. (b) Results of Chi-square contingency tests evaluating the strength of the association in collectors of young-of-year or older juvenile lobsters with potential crab and fish predators or competitors. Indicated in bold are significant p-values (p<0.05) along with the sign (+ or −) of the association. Analyses shown for study areas having large enough samples to give expected frequencies of no less than five collectors per cell of a 2×2 contingency table.
Species associations in collectors
We found a diversity of associated fauna in collectors, principally fishes and crabs (a). The complement of species varied among study areas with somewhat greater diversity in southern New England (Rhode Island) than the Gulf of Maine (Maine and Nova Scotia). We had sufficient numbers of collectors in eastern and mid-coast Maine, Rhode Island and Lobster Bay, Nova Scotia to conduct the Chi-square contingency tests to evaluate whether the occurrence of lobsters in collectors was independent of the occurrence of crabs and fish. We found no significant association in most cases (b). The only case of a significantly negative association occurred for older juvenile lobsters and crabs in Rhode Island. Statistically significant positive associations were found for young-of-year lobsters and fish in mid-coast Maine, and older juveniles and fish in eastern Maine, mid-coast Maine and Lobster Bay, Nova Scotia. The collector data in most cases, except Rhode Island, do not support the hypothesis that associated fish or crabs negatively affect the recruitment of lobsters to collectors. Questions remain about the mechanism behind the observed positive associations that will be addressed in the Discussion.
Discussion
By virtue of this international collaboration, our study has characterized patterns of American lobster young-of-year recruitment and older juvenile abundance on an unprecedented spatial scale using two standardized sampling approaches: diver-based suction sampling and passive, vessel-deployed collectors. It has refined our understanding of the performance of the samplers, affirmed previous hypotheses and stimulated new ones on the determinants of postlarval settlement patterns.
The large-scale pattern revealed study areas of low and high benthic recruitment. Results of the surveys suggest that postlarval settlement is most consistent and concentrated in the Gulf of Maine, Bay of Fundy, southwestern Nova Scotia and southern Gulf of St. Lawrence regions. We suspect it is no coincidence that these regions support some of the most productive lobster fisheries in the species range (Fisheries and Oceans Canada Citation2011; NOAA Fisheries Citation2011). The significant correlation of young-of-year and older juveniles at the same sites, together with our longer-term time series at these locations and previous larval transport modelling (Incze et al. Citation2010), suggests spatial differences in recruitment are relatively consistent year to year, resulting in recruitment hot and cold spots. Advective processes influencing larval and postlarval transport could be responsible for consistent along-shore heterogeneity in settlement to coastal nurseries (Wahle & Incze Citation1997; Xue et al. Citation2008; Chassé & Miller Citation2010; Incze et al. Citation2010). However, as has been shown for other benthic species (Burke Citation1986; Pearce & Scheibling Citation1990; Raimondi Citation1991), the possibility should not be ruled out that postlarvae may concentrate in nurseries by a behavioural attraction to high densities of benthic conspecifics (Boudreau et al. Citation1991; Burdett-Coutts Citation2010). Although sampling has been conducted for more than two decades at some of these locations (Incze et al. Citation1997; Wahle et al. Citation2009a), the present study represents the first sampling at others. The project therefore broadens the baseline that will be valuable in assessing recruitment success to lobster nurseries over the species’ range in the coming decades.
Although not a direct test of patterns of postlarval settlement behaviour, our results are consistent with the hypothesis that postlarval settlement tends to concentrate above the thermocline in thermally stratified waters (Boudreau et al. Citation1992; Wilson Citation1999). The first concerted effort to evaluate depth-wise patterns of lobster settlement was conducted by divers using suction samplers and diver-deployed collectors to depths of 20 m (Wilson Citation1999). Wilson's findings and earlier laboratory behavioural studies with postlarvae (Boudreau et al. Citation1992) suggested that the depth of settlement might be determined by the depth of thermal stratification. In direct underwater field observations of individual postlarvae diving through thermoclines, Annis (Citation2005) found that postlarvae tended to terminate dives near the 12°C isotherm. Our deployments of collectors over a range of depths to 80 m in oceanographically contrasting settings along the coast indicate that in thermally stratified study areas, young-of-year and older juvenile lobsters concentrated in zones above the thermocline. Given the limited range of movement of young-of-year lobsters, it is unlikely they moved to shallower water after postlarval settlement. Nonetheless, a few young-of-year lobsters were found below the thermocline, indicating that the thermocline is not an absolute barrier to postlarvae.
Our discovery that although settlement tends to concentrate above the thermocline some settlers transcend the thermocline raises questions about the fate of these lobsters, as growth potential would be limited at temperatures perpetually well below 10°C. Alternatively, in well-mixed regions, such as the Bay of Fundy, temperatures are paradoxically warmer at depths>30 m during the summer than they are in those further south that stratify thermally. Therefore, deep waters in these well-mixed areas may provide large areas of suitable nursery habitat. Together with seabed maps providing the availability of suitable habitat, collector-based sampling could improve our ability to extrapolate recruitment over large areas of the sea floor. Finally, two points are worth highlighting regarding depth distributions: (1) even in the thermally uniform water column of the Bay of Fundy, young-of-year lobsters still concentrate in the shallowest strata, and (2) we did not detect settlement on the offshore banks, even though collectors were shallower than some of the nearshore sites and placed on cobble habitat. Together these facts suggest that lobster postlarvae settle preferentially in nearshore habitat, regardless of the thermocline. Sampling of the offshore banks was limited in this study, however, and further exploration is necessary.
To be a nursery, the sea bed must have both an adequate postlarval supply and suitable habitat. Suction sampling enables us to assess young-of-year recruitment to suitable habitat where we find it. As portable cobble habitat units, passive collectors represent an important new tool enabling us to assess the potential for young-of-year recruitment even where the natural sea bed is unsuitable. Adequate sea bed mapping, therefore, would be essential for a full understanding of the potential of the sea bed for lobster production, and to identify locations where habitat enhancement would be likely to intercept a consistent supply of settling postlarvae.
Our findings are also consistent with previous work suggesting collectors and suction samplers compared favourably with respect to the density and size structure of lobsters collected (Wahle et al. Citation2009b). We found no difference in the density of young-of-year lobsters sampled by the two methods, although our results suggest that collectors may slightly under-sample larger juveniles entering collectors from the surrounding sea bed. A possible explanation for this size selectivity is that lobsters in their first or second year of life are cryptic and sedentary and would have less tendency to move into collectors from the surrounding habitat than somewhat older juveniles (Lawton & Lavalli Citation1995 Wahle & Incze Citation1997). Continued side-by-side sampling by the two methods in the coming years will provide additional statistical power to resolve these differences.
We found little evidence that suspected predators such as crabs or fish interact negatively with either young-of-year or older juvenile lobsters in collectors. Indeed, among the four study areas analysed, only in Rhode Island did we find a significant negative association between older juvenile lobsters and crabs. Crabs can be especially abundant in southern New England, and may represent a potential predator, prey and competitor for food, habitat or shelter (Richards & Cobb Citation1986; Barshaw & Lavalli Citation1988; Loher & Whitlatch Citation2002).
Despite the potential for negative interactions, positive associations within collector should perhaps be expected more often than not for the simple reason that most of the species associated with lobsters are also naturally associated with cobble habitat. Therefore, co-occurrence of species in collectors could simply reflect the heterogeneity of the habitat surrounding the collector. Conversely, infilling of a collector by sand, as occurred on occasion, could reduce available shelter space, and could make it equally unsuitable for lobster, fishes and crabs alike. Evaluating species associations within samples should continue to be an important part of understanding the sampling process.
Other sampling tools have been employed in population studies of the American lobster, including traps and trawls, diver-based and remote visual surveys. A wealth of data have been generated by various studies in many geographic areas. These data are not necessarily accessible or available in a form that allows ready comparison from study area to study area. In a few cases, relatively long-time series monitoring or surveys have been designed with the express purpose of following cohorts through time. For example, in the Magdelaine Islands, Quebec, Canada, multi-year, methodical transect surveys of lobster nurseries by experienced divers in clear water have provided a valuable time series of the fate of juvenile cohorts from the time of initial postlarval settlement (Hudon Citation1987; Gendron & Sainte-Marie 2007). Such painstaking visual censuses are not likely to be practical in monitoring of young-of-year lobsters, however, because of time constraints, differences in diver experience, variable visibility and the need for spatial replication. As long as we accept the reduced sampling efficiency of visual surveys for the smallest lobsters, however, visual quadrat and transect methods can nonetheless contribute meaningfully to our understanding of patterns and processes of shallow-water American lobster populations (Bernstein & Campbell Citation1983; Hudon Citation1987; Wahle Citation1993; Steneck & Wilson Citation2001; Gendron & Sainte-Marie 2007). In the case of the Steneck & Wilson (Citation2001) study in Maine, spatial patterns of young-of-year and older juveniles sampled by suction and visual methods, respectively, were convincingly linked to spatial patterns in the commercial catch of adults. Only recently have young-of-year time series generated by suction sampling been used in forecasting time trends in the abundance of lobster about to recruit to the fishery (Wahle et al. Citation2009a), as has long been practiced in the western Australia rock lobster fishery (Caputi et al. Citation1995). Therefore, efforts to standardize monitoring protocols across regions and integrate sampling methods across the life stages will allow broader-scale comparisons and forecasting potential.
In conclusion, we have compiled the results of widespread sampling of young-of-year and older juvenile American lobsters populations spanning a 1500-km segment of the species’ geographic range, from 41° to 49°N, by suction sampling and passive collector. Our results showed regional and bathymetric differences in the intensity of benthic recruitment by underscoring the importance of nearshore shallow cobble habitats as prime nurseries for lobsters. Observed low levels of settlement to depths as great as 80 m suggest the potential that vast areas of sea bed in deep water may contribute to overall recruitment, notwithstanding the lesser potential for growth at those depths, and the failure to detect evidence of settlement on offshore banks so far. Furthermore, our results build on previous comparisons of the performance of passive collectors and suction samplers. Although we report no sampling bias for young-of-year lobsters that settle from the plankton, we found collectors to slightly under-sample intermediate-size juveniles that are less mobile than larger juveniles. Aside from these slight differences in size selectivity, the two methods provide a consistent picture of lobster recruitment that together with an understanding of habitat availability will be useful in estimating regional differences in lobster production. Finally, although collectors sample a broad spectrum of fishes, crabs and other invertebrates along with lobsters, we found limited evidence that associated fish or crabs negatively affect the occurrence of young-of-year lobsters in collectors. Taken together, the two sampling methods represent key quantitative approaches within the tool kit needed to monitor changes in lobster nursery populations throughout the species’ geographic range.
Editorial responsibility: Roy Kropp
Acknowledgements
Long-term and broad geographic analyses are predicated on a large group of dedicated scientific support teams and industry participants. We acknowledge financial support from the participating states and provinces in the US and Canada; a US National Oceanic and Atmospheric Administration–Northeast Consortium grant to R. Wahle; Natural Sciences and Engineering Research Council of Canada grants to R. Rochette and P. Snelgrove; Fisheries and Oceans Canada; the Fishermen and Scientists Research Society; and Community-University Research for Recovery Alliance grant to B. Neis from the Social Sciences and Humanities Research Council of Canada. Special thanks go to the fishing industry collaborators who provided advice and vessel support, and to the many students, interns and technicians who assisted in the field and laboratory. Finally, we thank R. Kropp, P. Yund and two anonymous reviewers for helpful comments on the manuscript.
Notes
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
References
- Annis , ER. 2005 . Temperature effects on the vertical distribution of lobster postlarvae (Homarus americanus) . Limnology and Oceanography , 50 : 1972 – 82 .
- Balch , T and Scheibling , RE. 2000 . Temporal and spatial variability in settlement and recruitment of echinoderms in kelp beds and barrens in Nova Scotia . Marine Ecology Progress Series , 205 : 139 – 54 .
- Barshaw , DE and Lavalli , KL. 1988 . Predation upon postlarval lobsters Homarus americanus by cunners Tautogolabrus adspersus and mud crabs Neopanope sayi on three different substrates, eelgrass, mud and rocks . Marine Ecology Progress Series , 48 : 119 – 23 .
- Bernstein BB , Campbell A . 1983 . Contribution to the development of methodology for sampling and tagging small juvenile lobsters . Canadian Technical Report in Fisheries and Aquatic Sciences 1741 . 34 pages .
- Boudreau , B , Simard , Y and Bourget , E. 1991 . Behavioural responses of the planktonic stages of the American lobster Homarus americanus to thermal gradients, and ecological implications . Marine Ecology Progress Series , 76 : 13 – 23 .
- Boudreau , B , Simard , Y and Bourget , E. 1992 . Influence of a thermocline on vertical distribution and settlement of post-larvae of the American lobster Homarus americanus Milne-Edwards . Journal of Experimental Marine Biology and Ecology , 162 : 35 – 49 .
- Briones-Fourzan , P , Candela , J and Lozano-Alvarez , E. 2008 . Postlarval settlement of the spiny lobster Panulirus argus along the Caribbean coast of Mexico: Patterns, influence of physical factors, and possible sources of origin . Limnology and Oceanography , 53 : 970 – 85 .
- Burdett-Coutts , V. 2010 . Larval supply, settlement, and recruitment of American lobster, Homarus americanus . MSc thesis , Memorial University , St. Johns , Newfoundland , Canada . 171 pages .
- Burke , RD. 1986 . Pheromones and the gregarious settlement of marine invertebrate larvae . Bulletin of Marine Sciences , 39 : 323 – 31 .
- Caputi , N , Brown , RS and Phillips , BF. 1995 . Predicting catches of the western rock lobster (Panulirus cygnus) based on indices of puerulus and juvenile abundance . ICES Marine Science Symposium , 199 : 287 – 93 .
- Chassé , J and Miller , RJ. 2010 . Lobster larval transport in the southern Gulf of St. Lawrence . Fisheries Oceanography , 19 : 319 – 38 .
- Conover WJ. 1971 . Practical Nonparametric Statistics . New York , NY : Wiley . 462 pages .
- Cowen , RK , Lwiza , KMM , Sponaugle , S , Paris , CB and Olson , DB. 2000 . Connectivity of marine populations: Open or closed? . Science , 287 : 857 – 59 .
- Eggleston , DB and Armstrong , DA. 1995 . Pre- and post-settlement determinants of estuarine dungeness crab recruitment . Ecological Monographs , 65 : 193 – 216 .
- Fisheries and Oceans Canada . 2011 . Commercial fisheries landings . http://www.dfo-mpo.gc.ca/stats/commercial/sea-maritimes-eng.htm (accessed 7 December 2011) .
- Gardner , C , Frusher , SD , Kennedy , RB and Cawthorn , A. 2001 . Relationship between settlement of southern rock lobster pueruli, Jasus edwardsii, and recruitment to the fishery in Tasmania, Australia . Marine and Freshwater Research , 52 : 1271 – 75 .
- Gendron , L and Sainte-Marie , B. 2006 . Growth of juvenile lobster Homarus americanus off the Magdalen Islands (Quebec, Canada) and projection of instar and age at commercial size . Marine Ecology Progress Series , 326 : 221 – 33 .
- Hudon , C. 1987 . Ecology and growth of postlarval and juvenile lobster, Homarus americanus, off Iles de la Madeleine (Quebec) . Canadian Journal of Fisheries and Aquatic Sciences , 44 : 1855 – 69 .
- Incze , LS and Wahle , RA. 1991 . Recruitment from pelagic to early benthic phase in lobsters Homarus americanus . Marine Ecology Progress Series , 79 : 77 – 87 .
- Incze , LS , Wahle , RA and Cobb , JS. 1997 . Quantitative relationships between postlarval supply and benthic recruitment in the American lobster, Homarus americanus . Fisheries Bulletin , 48 : 729 – 43 .
- Incze , L , Xue , H , Wolff , N , Xu , D , Wilson , C Steneck , R . 2010 . Connectivity of lobster (Homarus americanus) populations in the coastal Gulf of Maine: Part II. Coupled biophysical dynamics . Fisheries Oceanography , 19 : 1 – 20 .
- Lawton , P and Lavalli , K. 1995 . “ Postlarval, juvenile, adolescent, and adult ecology ” . In Biology of the Lobster Homarus americanus , Edited by: Factor , JR . 47 – 88 . New York , NY : Academic Press .
- Levene H. 1960 . In Olkin I , Ingram I , Ghurye SG , Hoeffding W , Madow WG , Mann HB Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling . Stanford , CA ; Stanford University Press , 278 – 92 .
- Lohrer , AM and Whitlatch , RB. 2002 . Interactions among aliens: Apparent replacement of one exotic species by another . Ecology , 83 : 719 – 33 .
- Moksnes , PO and Wennhage , H. 2001 . Methods for estimating decapod larval supply and settlement: importance of larval behavior and development stage . Marine Ecology Progress Series , 209 : 257 – 73 .
- NOAA Fisheries . 2011 . Commercial fisheries landings . http://www.st.nmfs.noaa.gov/st1/commercial/index.html (accessed 7 December 2011) .
- Navarrete , SA , Wieters , EA , Broitman , BR and Castilla , JC. 2005 . Scales of benthic–pelagic coupling and the intensity of species interactions: From recruitment limitation to top-down control . Proceedings of the National Academy of Sciences USA , 102 : 18046 – 51 .
- Nelson GA . 2011 . fishmethods: Fisheries Methods and Models in R. R package version 1.2-0 . http://CRAN.R-project.org/package=fishmethods (accessed 22 October 2012). Computer program.
- Pearce , CM and Scheibling , RE. 1990 . Induction of settlement and metamorphosis in the sand dollar Echinaranchnius parma: Evidence for an adult-associated factor . Marine Biology , 107 : 363 – 69 .
- Pile , AJ , Lipcius , RN , Van Montfrans , J and Orth , RJ. 1996 . Density-dependent settler–recruit–juvenile relationships in blue crabs . Ecological Monographs , 66 : 277 – 300 .
- Pineda , J. 2000 . Linking larval settlement to larval transport: Assumptions, potentials, and pitfalls . Oceanography of the Eastern Pacific , I : 84 – 105 .
- Raimondi , P. 1991 . Settlement behavior of Chthamalus anisopoma larvae largely determines the adult distribution . Oecologia , 85 : 349 – 60 .
- Richards , A and Cobb , JS. 1986 . Competition for shelter between lobsters (Homarus americanus) and Jonah crabs (Cancer borealis): Effects of relative size . Canadian Journal of Fisheries and Aquatic Sciences , 43 : 2250 – 55 .
- Roughgarden , J , Gaines , S and Possingham , H. 1988 . Recruitment dynamics in complex life cycles . Science , 241 : 1460 – 66 .
- Steneck , RS and Wilson , CJ. 2001 . Large-scale and long-term spatial and temporal patterns in demography and landings of the American lobster, Homarus americanus, in Maine . Fisheries Bulletin , 52 : 1303 – 20 .
- Underwood AJ. 1997 . Experiments in Ecology: Their Logical Design and Interpretation using Analysis of Variance . Cambridge : Cambridge University Press . 524 pages .
- Wahle , RA. 1993 . Recruitment to American lobster populations along an estuarine gradient . Estuaries , 16 : 731 – 38 .
- Wahle , RA , Cobb , JS , Incze , LS , Lawton , P , Gibson , M Glenn , R . 2010 . The American lobster settlement index at 20 years: Looking back – looking ahead . Journal of the Marine Biological Association of India , 52 : 180 – 88 .
- Wahle RA , Fogarty M . 2006 . Growth . Chapter 1 in Phillips B Lobsters: Biology, Management, Aquaculture and Fisheries . Sunderland , MA : Sinauer , 1 – 44 .
- Wahle , RA and Incze , LS. 1997 . Pre- and post-settlement processes in recruitment of the American lobster . Journal of Experimental Marine Biology and Ecology , 217 : 179 – 207 .
- Wahle , RA and Steneck , RS. 1991 . Recruitment habitats and nursery grounds of the American lobster Homarus americanus – A demographic bottleneck . Marine Ecology Progress Series , 69 : 231 – 43 .
- Wahle , RA , Gibson , C and Fogarty , MJ. 2009a . Distinguishing disease impacts from larval supply effects in a lobster fishery collapse . Marine Ecology Progress Series , 376 : 185 – 92 .
- Wahle , RA , Wilson , C , Parkhurst , M and Bergeron , CE. 2009b . A vessel-deployed passive postlarval collector to assess settlement of the American lobster Homarus americanus . New Zealand Journal of Marine and Freshwater Research , 43 : 465 – 74 .
- Wilson CJ . 1999 . Bathymetric and spatial patterns of settlement in American lobster, H. americanus, in the Gulf of Maine: Insights into processes controlling abundance . Master of Science thesis , University of Maine . 37 pages .
- Xue , H , Incze , L , Xu , D , Wolff , N and Pettigrew , N. 2008 . Connectivity of lobster populations in the coastal Gulf of Maine, Part I: Circulation and larval transport potential . Ecological Modeling , 210 : 193 – 211 .