
ABSTRACT
Dynoides canadensis sp. nov. is described from the south-western coast of British Columbia, Canada. This species differs from its closely related species, D. elegans (Boone, 1923) ), by the presence of a smooth pleotelson without prominent rounded tubercle on the basal part of the pleotelsonic sinus, its sinus wall lacking crenulation; in having the penial processes length 7.7 × basal width and acute distal apex instead of 2.3 × basal width and rounded and blunt distal apex in D. elegans. This new species also has an appendix masculina with the distal apex extending somewhat beyond its base, instead of not reaching the base as is the case for D. elegans. A map of the distribution for the north-eastern Pacific species of Dynoides Barnard, 1914 (D. elegans (Boone, 1921); D. crenulatus and D. saldanai Carvacho & Haasmann, 1984; and D. dentisinus Shen 1929) in the north-eastern Pacific is provided. The new species is abundant in the western coastal zone of British Columbia. A revised generic diagnosis is provided for the genus Dynoides.
Publication LSID: lsid:http://zoobank.org:pub:ADF85BE9-C307-43EA-B270-1B2DB2F61883
Introduction
The genus Dynoides Barnard, Citation1914 has 18 accepted species (Boyko et al. Citation2008 onwards), which mostly occur in the Pacific Ocean from the north-eastern to the western Pacific intertidal and shallow water habitats. However, three species are known from the Indian Ocean: Dynoides serratisinus Barnard, Citation1914 from South Africa; Dynoides indicus Müller, Citation1991 from Sri Lanka; and Dynoides amblysinus Pillai, Citation1954 from India. Only Dynoides castroi Loyola and Silva, Citation1960 is reported from the South Atlantic Ocean (Brazil).
In the north-eastern Pacific, four species are known: D. crenulatus Carvacho and Haasman Citation1984, D. saldanai Carvacho and Haasmann, Citation1984 both from the Pacific Coast of Mexico; D. elegans (Boone, Citation1923) from the California coasts; and D. dentisinus Shen, Citation1929 from San Francisco Bay, which was originally described from the coast of North China. Additionally, Dynoides sp. also occurs in Mexico, Baja California Sur, Bahia de Concepcion (Wetzer et al. Citation2018). During the present study specimens of Dynoides collected along the west coast of Canada, off British Columbia, either incorrectly identified as Dynamenella perforata or only identified at the genus level (i.e. Dynoides), were reexamined. The specimens are morphologically distinct from any of the already described species of Dynoides. A new species of Dynoides is described herein.
Materials and methods
All specimens examined for this study are from British Columbia (), and were deposited in the Crustacea Collection of the Canadian Museum of Nature (Natural Heritage Campus, Gatineau, Quebec, Canada). Drawings were made with the aid of a camera lucida on Olympus BX51 compound microscope and illustrations were electronically inked with Corel Draw (version X6). Colour images of the specimens were taken using a Zeiss AxioCam ERc5s camera mounted on a Zeiss stereo microscope (Stemi 508).
Terminology of the morphological characters broadly follows Wetzer & Mowery (Citation2017). Abbreviations: CMNC – Canadian Museum of Nature, Canada; RS – robust seta/e; SPS – sensory palmate setae; PMS – plumose marginal setae.
Systematics
Family Sphaeromatidae Latreille, 1825
Genus Dynoides Barnard, Citation1914
The full synonymy for the genus was provided by Li (Citation2000).
Type species: Dynoides serratisinus Barnard, Citation1914 from Natal and Mozambique (by monotypy).
Generic diagnosis
Body dorsal surfaces smooth, covered with small tubercles and setosed in some species. Head longer than broad, with small median rostral process. Pereonites 2–7 without coxal plate sutures. Pleon with or without median prominent dorsal process, posterior margin with two separate sutures on either side. Pleotelson wider than long, posterior margin with a slit, sinus, notch, or foramen. Epistome anteriorly and posteriorly rounded. Maxilliped palp articles 2–4 medial margins slightly extended. Maxillula mesial endite with four long comb setae. Maxilla with four 4 curved pectinate RS on middle and lateral endites. Left mandible incisor and lacinia mobilis with 3–4 cusps, spine row present; palp articles 2 bearing two biserrate setae on distolateral margin in most of known species. Pereopods 1–7 ambulatory, inferior margins of merus to propodus bearing dense fine setae fringe. Penial processes long, entirely fused along the basal third to half of their length. Appendix masculina elongate, about twice the length of the endopod; distal third to more than distal half-length doubled back on proximal part. Pleopod 3 exopod without transverse suture. Pleopods 4 and 5 endopods bearing thickened transverse ridges, exopods thickened transverse ridges absent. Uropods with both rami broad and lamellar, subequal in length.
Dynoides canadensis sp. nov.
()
Figure 1. Map of the geographic distribution of the genus Dynoides Barnard, Citation1914, in the north-eastern Pacific. A, species distribution on the north-eastern Pacific coast; B, locations of Dynoides canadensis sp. nov. distribution; C, location of D. dentisinus distribution; D, location of D. elegans (red circle) and Dynoides sp. (blue square); E, D. crenulatus (red circle) and D. saldanai (green square).
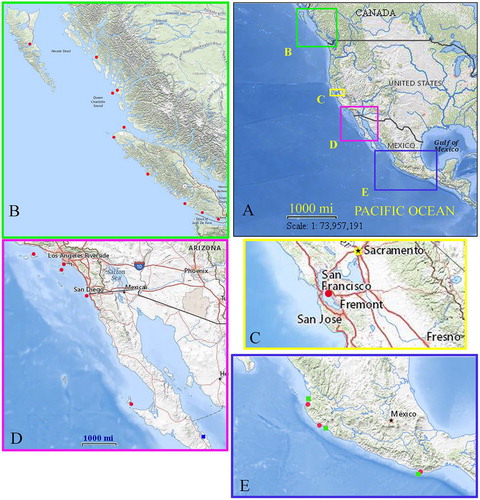
Figure 2. Dynoides canadensis sp. nov., holotype (CMNC 1985-0667.1); A, dorsal view; B, lateral view; C, epistome; D, antennula; E, antenna; F, maxillula; G, maxilla; H, left mandible; I, palp of mandible; J, maxilliped; K, ovigerous female (CMNC 1985-0667.2).
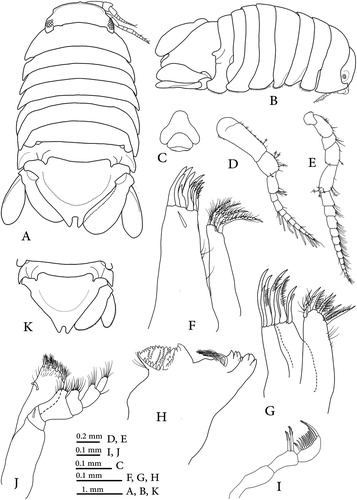
Figure 3. Dynoides canadensis sp. nov., holotype (CMNC 1985-0667.1); A–G, pereopods 1–7.
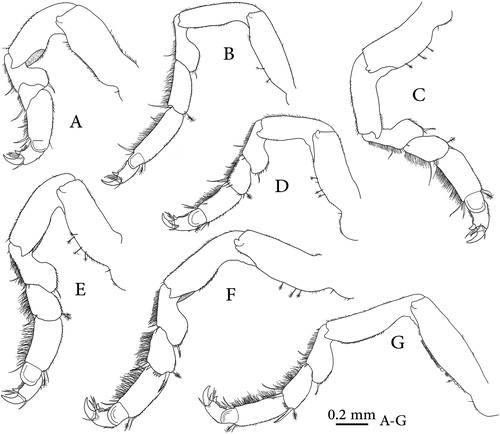
Figure 4. Dynoides canadensis sp. nov., holotype (CMNC 1985-0667.1); A, penes; B–F, pleopods 1–5; G, uropod.
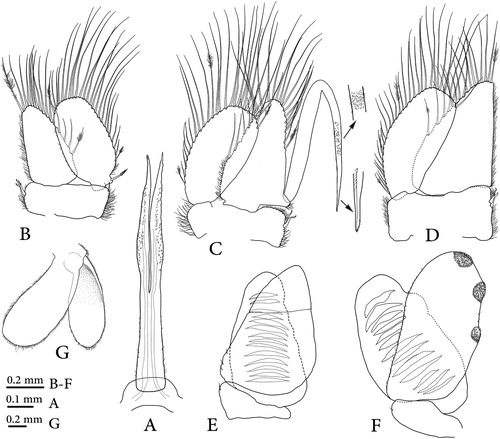
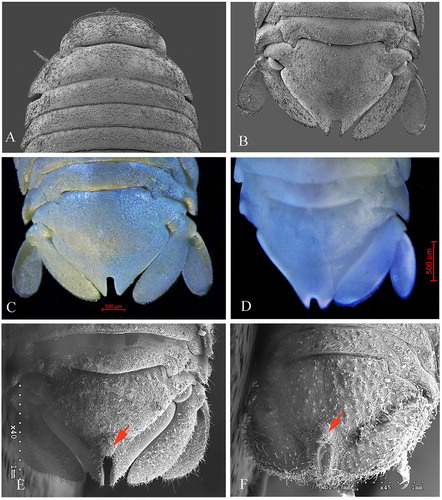
Figure 5. Dynoides canadensis sp. nov.; A, male, anterior dorsal; B, male, posterior dorsal; C, male, paratype, pleotelson; D, Ovigerous female, paratype, pleotelson; E, Dynoides elegans, male, pleotelson (after Wetzer & Mowery, Citation2017); F, Dynoides elegans, female, pleotelson (after Wetzer & Mowery, Citation2017). Red arrows show a prominent rounded tubercle on the base of pleotelsonic sinus.
Material examined
Holotype. ♂ (4.2 mm), Canada, British Columbia, Barclay Land District, Cape Beale, coll. Bousfield, Edward L., 19 July 1970 (CMNC 1985-0667.1).
Paratypes. 4 ♂♂(up to 4.2 mm), 18♀♀ (up to 4.3 mm), 1 ovigerous♀ (4.5 mm), same data as holotype, (CMNC 1985-0667.2). 2♂♂ (up to 5 mm), 2 ovigerous ♀♀ (3.5 mm); 2 ♀♀ (3.5 mm), British Columbia, Sooke Land District, Sooke Harbour, Whiffin Spit, coll. Bousfield, Edward L., 17 August 1955 (CMNC 1990-0064). 3 ♀♀ (up to 5 mm), British Columbia, Rupert Land Distr., Vancouver Island, Cape Scott, coll. Bousfield, Edward L., 18 July 1959 (CMNC 1990-0066). 20 ♂&♀, British Columbia, Renfrew Land Distr., Vancouver Island, Port Renfrew, coll. Bousfield, Edward L., 1 August 1970 (CMNC 1990-0068). 1♀ (3 mm), British Columbia, Queen Charlotte Islands Land Distr., Graham Island, 27 July 1957 (CMNC 1990-0067). 5 ♂♂ (up to 4.9), 6 ♀♀ (up to 4.2), British Columbia, Barclay Land District, British Columbia, Barclay Land Distr., Trevor Channel, Tzartus Island, coll. Bousfield, Edward L., 21 July 1970 (CMNC 1985-0664). 4 ♂♂ (up to 4.5 mm), 5 ♂♂ (up to 4.2 mm), British Columbia, Metchosin Land Distr., Sooke Basin, Becher Bay, coll. Bousfield, Edward L., 31 July 1970 (CMNC 1990-0057). 4♂♂ (up to 4.5 mm), 2 ♀♀ (3 mm), British Columbia, Nootka Land Distr., Nootka Island, coll. Bousfield, Edward L., 20 August 1959 (CMNC 1990-0059). 1 ♂ (4.2 mm), 1 ♂ (3.5 mm), British Columbia, Rupert Land Distr., Hope Island, coll. Bousfield, Edward L., 22 August 1959 (CMNC 1990-0060). 1 ♂ (5.1 mm), 5 ♂♂ (up to 4 mm), 1 juvenile, British Columbia, Range 2 Coast Land Distr., Goose Island, coll. Bousfield, Edward L., 6 August 1964 (CMNC 1990-0061). 6 ♀♀ (up to 3 mm), British Columbia, Range 2 Coast Land Distr., Hunter Island, coll. Bousfield, Edward L., 8 August 1964 (CMNC 1990-0063). 8 ♂♂ (up to 5 mm), 20 ♀♀ (up to 4.1 mm), 10 juveniles, British Columbia, Range 3 Coast Land Distr., Princess Royal Island, coll. Bousfield, Edward L., 20 July 1964 (CMNC 1990-0065).
Description (based on holotype male)
Body smooth, 1.84 times as long as greatest width, widest at pereonite 5 (A). Head with small rostral point. Pereonites 2–7 coxal plates fringed with small marginal setae; coxae of pereonite 6 reaching as far back as to overlap lateral parts of pereonite 7. Pleonite 1 with 2 anterolateral visible sutures (B). Pleon without median dorsal process. Pleotelson about 1.6 times as long as width, with a clear dorsal dome, pleotelsonic sinus with straight-sided margins, and slightly raised.
Epistome (C) with rounded anterior and posterior margins; lateral margins concave.
Antennula (D) not extending to posterior of pereonite 1; peduncle article 1, 2.6 times as long as article 2, inferior distal margin with 1 palm setae; peduncle article 2 inferior distal margin with 3 palm and some simple setae; peduncle articles 3 longer than article 2, inferior distal margin with 2 palm and 3 simple setae; flagellum with 9 articles, articles 2–8 each bearing an aesthetasc.
Antenna (E) extending to posterior of pereonite 1; peduncle articles 1 and 3 and peduncle articles 2 and 4 sub-equal in length; peduncle article 5 longest, with some palm setae on distal margin; flagellum with 14 articles.
Maxillula (F) lateral endite with 10 simple and serrated RS; mesial endite with 4 long comb and 1 small simple setae, 1 submarginal simple seta present.
Maxilla (G) lateral and middle endites each with 4 curved pectinate RS; mesial endite with 4 long rarely plumose and 3 long robust comb and 2 slender simple setae.
Left mandible (H) incisor with 4 cusps; lacinia mobilis with 3 cusps; spine row of 6 curved, serrate spines; molar process with serrate teeth; palp articles 2 with 2 biserrate setae, palp article 3 with 13 biserrate setae (I).
Maxilliped (J) endite lateral margin sinuate, distomesial margin with single coupling hook, distal margin with 4 long circumplumose, 2 densely circumplumose and 2 simple RS; palp articles 2–4 with weak distally lobe; palp article 4 with single long seta on superiodistal angle.
Pereopod 1 (A) basis about 2.5 times as long as greatest width, superior margin fringed with short, acute scale-setae; ischium, merus and propodus superior margin fringed with acute scale-setae; merus superodistal angle with single long RS; carpus triangular, inferodistal angle with single long RS; propodus 2.4 times as long as wide, inferodistal angle with 2 long RS, superodistal corner with single sensory palmate seta; dactylus inferior margin with cuticular scales, secondary unguis simple, with 2 simple setae at base, distal margin with 2 simple sub-marginal setae.
Pereopod 2 (B) basis about 3.2 times as long as greatest width, with 2 small SPS; ischium, merus, carpus and propodus superior margin fringed with acute scale-setae; ischium about 3.6 times as long as width; merus subequal in length to carpus, superodistal and inferodistal angles with 1 long simple seta; carpus inferodistal angle with a long SPS, superodistal angle with a long simple RS; propodus about 1.5 times as long as carpus, inferodistal angle with a long SPS; dactylus inferior margin with cuticular scales, secondary unguis simple, with 3 simple setae at base, distal margin with 2 simple sub-marginal setae.
Pereopod 3 (C) basis about 2.25 times as long as greatest width, with 3 small SPS; merus longer than carpus, inferodistal angles with 1 long serrated RS; carpus inferodistal angle with 1 SPS, superodistal angle with a long simple RS; propodus about 2 times as long as carpus, inferodistal angle with a long SPS; dactylus inferior margin with cuticular scales, secondary unguis simple.
Pereopods 4 and 5 (D and E) are similar to pereopod 3 as illustrated.
Pereopod 6 (F) basis about 3 times as long as greatest width, superior margin with 3 SPS; ischium, merus, carpus and propodus superior margin fringed with acute scale-setae; merus superodistal margin with 1 biserrate RS, inferior margin with 2 RS; carpus superodistal margin with 1 biserrated RS and 1SPS, inferior margin with 1 biserrate RS, distomedial margin with 4 robustacute scale-setae; propodus 1.6 times as long as carpus, superodistal corner with 1 slender and 1 SPS.
Pereopod 7 (G) basis about 4 times as long as greatest width, superior margin fringed with cuticular scale; ischium, merus, carpus and propodus are similar to pereopod 6 as illustrated.
Penial processes (A) about 7.7 times as long basal width entirely fused along basal 0.42 of length, distal margin bearing setules on most of surface except on apical parts, proximal lateral margin with some scale setae.
Pleopod 1 (B) exopod and endopod with 25 and 17 PMS respectively, endopod shorter than exopod; exopod proximal lateral corner with single biserrated seta. Sympodite mesial margin with 2 coupling hooks.
Pleopod 2 (C) exopod and endopod sub-equal in length, with 27 and 25 PMS respectively; appendix masculina proximally slightly swollen, distally narrowing, extending beyond endopod apex, more than distal half length doubled back on the proximal half and extending somewhat beyond its base; distal half bearing setules on most of surface medially, apex with some setules laterally; sympodite mesial margin with 2 coupling hooks.
Pleopod 3 (D) exopod and endopod subequal in length, with 34 and 16 PMS respectively; sympodite mesial margin with 2 coupling hooks.
Pleopod 4 (E) both rami subequal in length, exopod wider than endopod, with transverse suture, transverse folds absent.
Pleopod 5 (F) exopod with 3 scale patches, lateral margin with slender simple marginal setae.
Uropodal rami (G) extending just beyond pleotelsonic apex, fringed with dense small marginal setae.
Female: Similar to male except in sexual characters; body smooth, 1.9 times as long as greatest width; (K, D). Uropod not extending to posterior margin of pleotelson.
Etymology
The specific name is an adjective referring to the country of the type locality.
Remarks
Dynoides canadensis sp. nov. is most similar to D. elegans from California and D. saldanai and D. crenulatus from Mexico, Oaxaca. Based on the redescription of D. elegans given by Wetzer et al. (Citation2018), this new species differs from D. elegans, in having the penial processes length 7.7 × basal width and acute distal apex (instead of 2.3 × basal width and rounded and blunt distal apex), the doubled back part on the proximal half of the appendix masculina extending somewhat beyond its base (instead of not reaching the base). The pleotelson of D. elegans is covered with small tubercles, has a pleotelsonic sinus with a prominent rounded tubercle on the base of sinus and the sinus walls are finely crenulated (E, F). Conversely, the new species has a pleotelson without small tubercles on dorsal surface, lacking a prominent rounded tubercle on the basal part of the pleotelsonic sinus and its sinus wall has no crenulation. Dydoides crenulatus is readily distinguished from this new species by its highly setose and more slender body. The other species from Mexico, D. saldanai, can be easily recognized by the heart-shaped slit on the pleotelson apex.
The analyses of 16S rDNA by Wetzer et al. (Citation2018) clearly support the existence of two distinct Dynoides species along the coastal zone of California area; Dynoides elegans from Southern California, USA, and Dynoides sp. from Baja California Sur, Mexico. Based on distribution report of Dynoides species in the Mexican Pacific by Espinosa-Pérez and Hendrickx (Citation2001), Dynoides sp. could be D. crenulatus or its sympatric species, D. saldanai.
Comments on the geographic distribution
Based on the previous studies, four species of Dynoides are distributed in the north-eastern Pacific: D. crenulatus is sympatric with D. saldanai in most localities and distributed in the coastal zone of the south-west of Mexico from Oaxacan to near the Gulf of California (E); D. dentisinus occurs in San Francisco Bay, Western California (C); D. elegans reported from Cedros Island in Baja California Norte to Santa Cruz Island in Santa Barbara County, Southern California (D). The new species is distributed along the western coasts of British Columbia from Victoria area to Graham Island (B).
Acknowledgements
We are particularly grateful to Dr Robert Anderson for provision of the laboratory and to Mr. Philippe Ste-Marie for is assistance facilities during the first author visit to the Canadian Museum of Nature, Ottawa, in 2019. Dr Niel Bruce (Queensland Museum), Dr. Saskia Brix (Senckenberg Research Institute, Germany) and Dr Brenda Doti (Universidad de Buenos Aires, Argentina) are thanked for their constructive suggestions and comments that improved the manuscript. Financial support for this study was provided by Canadian Museum of Nature Visiting Scientist Awards 2019, and Shahrekord University, Iran.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Barnard KH. 1914. Contributions to the crustacean fauna of South Africa. 1. Additions to the marine Isopoda. Annals of the South African Museum. 10(7):197–230. doi:10.5962/bhl.part.9314.
- Boone PL. 1923. New marine tanaid and isopod Crustacea from California. Proceedings of the Biological Society of Washington. 36:147–156. https://www.biodiversitylibrary.org/part/43449.
- Boyko CB, Bruce NL, Hadfield KA, Merrin KL, Ota Y, Poore GCB, Taiti S, Schotte M, Wilson GDF, editors. 2008 onwards. World marine, freshwater and terrestrial isopod crustaceans database. Dynoides Barnard, 1914. Accessed through: World Register of Marine Species from: http://marinespecies.org/aphia.php?p=taxdetails&id=204371on2020-12-21.
- Carvacho A, Haasmann Y. 1984. Isopodos litorales de Oaxaca, Pacifico Mexico. Cahiers de Biologie Marine. 25:15–32.
- Espinosa-Pérez MC, Hendrickx ME. 2001. A new species of Exosphaeroma Stebbing (Crustacea: Isopoda: Sphaeromatidae) from the Pacific coast of Mexico. Proceedings of the Biological Society of Washington. 114:640–648. https://www.biodiversitylibrary.org/part/43978.
- Li L. 2000. A new species of Dynoides (Crustacea: Isopoda: Sphaeromatidae) from the Caped’ Aguilar Marine Reserve, Hong Kong. Records of the Australian Museum. 52:137–149. doi:10.3853/j.0067-1975.52.2000.1311
- de Loyola e Silva J. 1960. Sphaeromatidae do litoral Brasiliero (Isopoda-Crustacea). Boletim da Universidade do Parana, Zoologia. 4:1–182.
- Müller HG. 1991. Two new species of Cerceis and Dynoides from a sabellid reef at Sri Lanka (Crustacea: Isopoda: Sphaeromatidae). Zoologischer Anzeiger. 226(5/6):307–318.
- Pillai NK. 1954. A preliminary note on the Tanaidacea and Isopoda of Travancore. Bulletin of the Central Research Institute, University of Travancore (Natural Science). 3:1–23.
- Shen CJ. 1929. Description of a new isopod, Dynoides dentisinus, from the Coast of North China. Bulletin of the Fan Memorial Institute of Biology, Peiping. 1:65–75.
- Wetzer R, Bruce NL, Pérez-Losada M. 2018. Relationships of the Sphaeromatidae genera (Peracarida: Isopoda) inferred from 18S rDNA and 16S rDNA genes. Arthropod Systematics & Phylogeny. 76(1):1–30.
- Wetzer R, Mowery G. 2017. Redescription of Dynoides elegans (Boone, 1923) (Crustacea, Isopoda, Sphaeromatidae) from the north-eastern Pacific. ZooKeys. 646:1–16. doi:10.3897/zookeys.646.10626