Abstract
Based on material from the fourth cruise of the RV Akademik Mstislav Keldysh and the MAR-ECO cruise of the RV G.O. Sars, 31 species of Ophiuroidea were identified from the section of the Mid-Atlantic Ridge between the Reykjanes Ridge and the Azores. The taxonomy of the group of genera with reduced arm comb and tentacle pores restricted only to the proximal arm segments is reviewed. It is suggested that the genera Homalophiura H.L. Clark, 1915, Ophiurolepis Matsumoto, 1915, Theodoria Fell, 1961 and Homophiura Paterson, 1985 should be regarded as synonyms of the genus Ophioplinthus Lyman, 1878. From examination of the type specimens, it is demonstrated that the six-armed species Ophiacantha anomala G.O. Sars, Citation1871 also includes five-armed specimens previously identified as the separate species Ophiacantha cuspidata Lyman, 1878. The latter taxon is considered here as the junior synonym of O. anomala. Previously mentioned ‘deep-water populations of the common shallow water species Ophiacantha bidentata (Retzius, 1805)’ represent a separate species, Ophiacantha fraterna Verrill, 1885, which is well distinguished by the shape of the spicules of the disk, narrow outer oral papilla and narrower tentacle scale as well as a hermaphroditic nature. Three new species, i.e. Ophioplinthus pseudotessellata sp. nov., Ophiocamax patersoni sp. nov. and Ophiophyllum nesisi sp. nov., are described.
Introduction
The North Atlantic deep-water Ophiuroidea have a long history of study. The first deep-water ophiuroid from the North Atlantic was found during sounding of Baffin Bay at a depth of 1460 m in the early 19th century. This species was Gorgonocephalus eucnemis (Müller et Troschel, 1842) brought up wound around the plummet (Menzies et al. Citation1973). Regular collecting of ophiuroids took place during the famous deep-water expeditions, such as Porcupine and Lightning (Wyville Thomson Citation1873), Norwegian expeditions (Sars 1871), Challenger (Lyman 1878, Citation1882), Travailleur and Talisman (Koehler Citation1906, Citation1907a), Albatross (Verrill 1885a,b, 1894), Caudan (Koehler Citation1895, 1896a), Princesse Alice (Koehler Citation1896b, Citation1901a, Citation1909), L'Hirondelle (Koehler 1898), Michael Sars (Grieg Citation1921), Tjalfe, Thor, Dana, Ingolf (Mortensen Citation1913, Citation1933) and others. More recently, ophiuroids of the North Atlantic have been studied by Cherbonnier & Sibuet (Citation1972); cruises of the RV Jean Charcot), Gage et al. (Citation1983); mainly RRS Challenger), Paterson (Citation1985); campaign BIOGAS), and Bartsch (Citation1987, Citation1991) (RV Meteor). Paterson (Citation1985) summarized scattered data and reviewed all North Atlantic Ophiuroidea. Most recently, a new amphiurid species was described from deep waters around Iceland (Stöhr Citation2003). Stöhr & Segonzac (Citation2005) described several new species from hydrothermal vents and methane cold seeps of the North Atlantic, including also records from non-vent habitats. Tyler et al. (Citation2005) presented results of a study of Ophiuroidea around the Faeroe Islands, including both shallow- and deep-water species. Preliminary results of the study of the ophiuroid fauna from waters around Iceland were presented by Stöhr (oral communication).
Despite numerous studies, taxonomic problems still remain, even with common species. Species collected from the Reykjanes Ridge on two separate cruises were therefore examined in an effort to redefine the taxonomy of this and several other problematic ophiuroid families. Additionally, the genus Ophioplinthus was re-evaluated in order to elucidate its convoluted taxonomic history. Range extensions and species accounts are also given for the ophiuroid material collected from the Reykjanes Ridge taken by the RV Akademik Mstislav Keldysh in 1982 and the G.O. Sars cruise as part of the MAR-ECO 2004 project. For example, species of the genus Ophiocamax in the North Atlantic were recorded only from the Caribbean region and off the Sahara prior to this study. During the MAR-ECO programme a new species of this genus was discovered from the Charlie-Gibbs Fracture Zone and the Mid-Atlantic Ridge north of the Azores. Up to now, only one species of the genus Ophiophyllum was described from the Mid-Atlantic, O. atlanticum Stöhr & Segonzac, Citation2005. The present study discovered a new, second Atlantic species of Ophiophyllum from the Reykjanes Ridge.
Material and methods
The present work is based on numerous samples from the Reykjanes Ridge taken by the RV Akademik Mstislav Keldysh in 1982 and recent sampling during the G.O. Sars cruise under the framework of the MAR-ECO project in 2004 (www.mar-eco.no). Epibenthic fauna on the G.O. Sars MAR-ECO cruise was collected using a modified version of a semi-commercial otter trawl (the Campelen 1800 shrimp trawl) lined with a 5 mm mesh at the cod end (Bergstad & Gebruk Citation2008). The trawl had a mouth opening of 17 m × 12 m × 4.5 m. Ophiuroids in the MAR-ECO material occurred at 18 stations (). Specimens were preserved in 80% ethanol. The collection of the MAR-ECO ophiuroids was examined during a two-week visit to the Museum of Zoology, University of Bergen.
Table I. MAR-ECO trawl stations on which ophiuroids were sampled.
On the fourth cruise of Akademik Mstislav Keldysh to the Reykjanes Ridge in 1982 samples were collected using a 2.5 m Sigsbee trawl. Ophiuroids were found at 21 stations (). Specimens were preserved in 80% ethanol.
Table II. List of the stations of the fourth and 46th cruise of RV Akademik Mstislav Keldysh.
Additionally, ophiuroids from one trawl station in the north-west Atlantic taken in 2001 on the 46th cruise of Akademik Mstislav Keldysh were also examined ().
Abbreviations
IORAS, P.P. Shirshov Institute of Oceanology, Moscow, Russia; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, MA, USA; SMNH, Swedish Museum of Natural History, Stockholm, Sweden; USNM, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA; YPM, Yale Peabody Museum, Yale University, New Haven, CT, USA; ZMBN, Museum of Zoology at the University of Bergen, Norway; ZMMU, Zoological Museum, Moscow State University, Russia; ZSM, Zoologische Staatssammlung München, Germany.
Species account
Family Asteronychidae
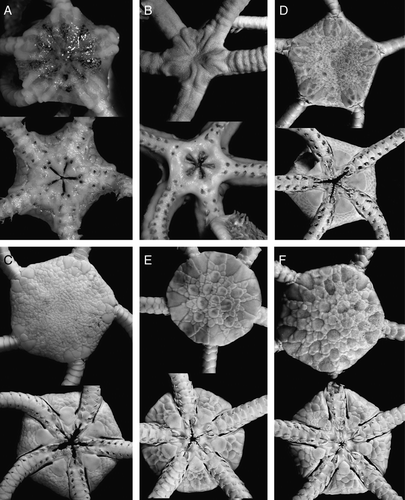
Astrodia tenuispina (Verrill, Citation1884)
(A)
Astronyx tenuispina: Verrill 1884: 219.
Astrodia tenuispina: Verrill 1899: 74; Paterson Citation1985: 15–6, .
Material
RV G.O. Sars, MAR-ECO cruise, St. 50/373, two specimens.
Distribution
North Atlantic: in the western part – the east coast of the USA, in the east Atlantic – Rockall Trough, Bay of Biscay, off Portugal, at 1560–3659 m (Paterson Citation1985). Present study: Mid-Atlantic Ridge, north of the Azores, at 2593–2607 m.
Family Asteroschematidae
Asteroschema inornatum Koehler, Citation1906
(B)
Asteroschema inornatum: Koehler Citation1906: 30–1, Plate 3, Figures 46, 47; Paterson Citation1985: 16, .
Material
RV G.O. Sars, MAR-ECO cruise, St. 56/378, one specimen; RV Akademik Mstislav Keldysh, fourth cruise, St. 364, two specimens.
Distribution
North Atlantic: Rockall Trough, Bay of Biscay, off north-west Spain, off Azores, 1478–2170 m (Paterson Citation1985). Present study: the Reykjanes Ridge and the Mid-Atlantic Ridge south of the Charlie-Gibbs Fracture Zone, at 1300–1950 m.
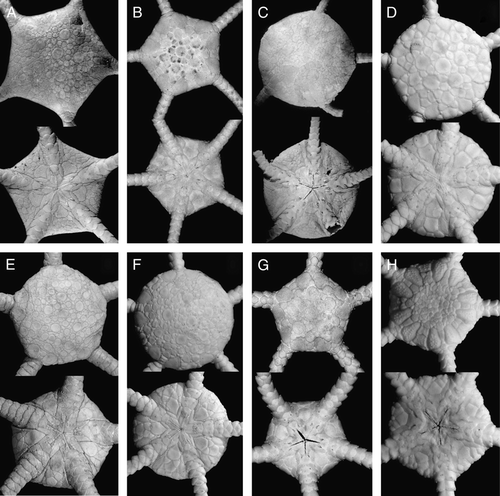
Figure 1. (A) Astrodia tenuispina (Verrill, Citation1884), MAR-ECO, St. 50/373, disk diameter 10 mm; (B) Asteroschema inornatum Koehler, Citation1906, ZMMU D-732, RV Akademik Mstislav Keldysh, St. 364, disk diameter 14 mm; (C) Ophiura irrorata (Lyman, 1878), MAR-ECO, St. 72/386, disk diameter 23 mm; (D) Ophiura ljungmani (Lyman, 1878), MAR-ECO, St. 40/367, disk diameter 10 mm; (E) Ophiura nitida Mortensen, Citation1933, MAR-ECO, St. 72/386, disk diameter 7.8 mm; (F) Ophiura violainae (Cherbonnier & Sibuet, Citation1972), MAR-ECO, St. 70/385, disk diameter 11 mm. Dorsal and ventral views are shown. Photographs: Tanya Korshunova (A, B); Konstantin Tabachnik (C–F).
Family Ophiuridae
Ophiura irrorata (Lyman, 1878)
(C)
Ophioglypha irrorata: Lyman 1878: 73, Plate 4, Figures 106–108.
Ophiura irrorata: H.L. Clark Citation1911: 62.
Ophiura (Ophioglypha) irrorata irrorata: Paterson Citation1985: 123–4, Figures 46, 47.
Material
RV G.O. Sars, MAR-ECO cruise, St. 46/372, one specimen; St. 50/373, eight specimens; St. 52/374, eight specimens; St. 72/386, 13 specimens.
Distribution
All oceans, except Arctic, at 403–7340 m. North Atlantic records: Cape Cod, south-western Iceland, Bay of Biscay – off North Africa. It is a predominantly abyssal species, inhabiting mostly depths below 2000 m (Paterson Citation1985). Present study: the Charlie-Gibbs Fracture Zone, the Mid-Atlantic Ridge north of the Azores, at 2522–3050 m.
Remarks
Paterson (Citation1985) considered three subspecies of O. irrorata within the same regions of the North Atlantic (e.g. O. irrorata irrorata inhabited the Bay of Biscay along with another subspecies O. irrorata concreta). Whether these subspecies represent separate species or rather a single polymorphic species is currently unclear. However, several subspecies cannot inhabit the same geographical region (e.g. Bay of Biscay), and we therefore consider O. irrorata without the subspecies division.
Ophiura ljungmani (Lyman, 1878)
(D)
Ophioglypha ljungmani: Lyman 1878: 71, Plate 3, Figure 77.
Ophiura ljungmani: Farran Citation1913: 31.
Ophiura (Ophiura) ljungmani: Paterson Citation1985: 118–20, Figure 44.
Material
RV G.O. Sars, MAR-ECO cruise, St. 40/367, two specimens; St. 53/375, one specimen; St. 54/377, five specimens; St. 66/383, two specimens; St. 72/386, two specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 268, 70 specimens; St. 316, seven specimens; St. 319, two specimens; St. 390, three specimens; St. 445, four specimens.
Distribution
Eastern Brazil, off Florida, at 101–2750 m. North Atlantic records: Labrador basin, south-eastern Iceland, North Africa (Paterson Citation1985). Present study: from the Reykjanes Ridge, including the Charlie-Gibbs Fracture Zone to the Azores, at 981–3527 m.
Ophiura nitida Mortensen, Citation1933
(E)
Ophiura nitida: Mortensen Citation1933: 84–6, Text figure 47, Plate 3: 18, 19.
Ophiura (Ophiura) nitida: Paterson Citation1985: 121–2, Figure 45.
Material
RV G.O. Sars, MAR-ECO cruise, St. 68/384, two specimens; St. 72/386, three specimens.
Distribution
Ophiura nitida is a rare ophiurid that has been recorded only from the Reykjanes Ridge, from a depth of 2065 m (Mortensen Citation1933). The present study revealed this species in the Charlie-Gibbs Fracture Zone at 2374–2567 m.
Remarks
This is the first record of this species since the description by Mortensen (Citation1933).
Ophiura violainae (Cherbonnier & Sibuet, Citation1972)
(F)
Homalophiura violainae: Cherbonnier & Sibuet Citation1972: 1378.
Ophiura (Ophiura) violainae: Paterson Citation1985: 122, Figure 45.
Material
RV G.O. Sars, MAR-ECO cruise, St. 70/385, one specimen; RV Akademik Mstislav Keldysh, fourth cruise, St. 465, two specimens.
Distribution
Bay of Biscay, off Portugal, at 2210–2399 m (Cherbonnier & Sibuet Citation1972; Paterson Citation1985). Present study: the Reykjanes Ridge, the Charlie-Gibbs Fracture Zone at 1620–1670 m.
Remarks
This is the first record of the species on the Reykjanes Ridge.
Ophiura saurura (Verrill, Citation1894) comb. nov.
(A–C)Ophioglypha saurura:Verrill 1894: 288.
Ophioglypha aspera: Koehler 1898: 40, Plate 6, Figures 19–21.
Amphiophiura saurura: H.L. Clark Citation1915: 315; Paterson Citation1985: 134–5, Figure 50.
Material
RV G.O. Sars, MAR-ECO cruise, St. 40/367, 19 specimens; St. 42/368, one specimen; St. 52/374, five specimens; St. 53/375, seven specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 465, four specimens; St. 489, two specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 445, one specimen.
Distribution
North Atlantic: off Nantucket and Newfoundland, south of Iceland to Bay of Biscay, at 844–2245 m (Paterson Citation1985). Present study: the Reykjanes Ridge, the Mid-Atlantic Ridge north of the Azores, at 981–2979 m.
Remarks
Specimens from St. 445 (B) are similar to the type specimen (YPM 9419) (A) in having numerous elevated disk plates, connected radial shields, straight distal edge of the dorsal arm plates, rounded proximal outline of the jaws and whitish colour. At the same time, most of the other specimens from various stations (e.g. C) somewhat differ from the type specimen and the specimen from St. 445 in the presence of a superficially smooth disk with non-elevated plates, radial shields conspicuously separated, triangular distal edge of the dorsal arm plates, triangular proximal outline of the jaws and maculated colour. Despite these differences, evidently intermediate specimens between these forms were discovered, therefore we consider both forms within a single species. Traditionally, O. saurura is considered within the genus Amphiophiura (Clark Citation1915; Mortensen Citation1933; Paterson Citation1985), but this placement was never discussed. Almost all species of the genus Amphiophiura (including the type species A. bullata (Wyville Thomson, 1877) have a peculiarly stocky general appearance with strong disk and arm plates and differ considerably from O. saurura. The latter is similar to the species of the genus Ophiura in general shape of the disk and arms, especially to O. irrorata. Differences between O. saurura and O. irrorata are smaller than with species from the genus Amphiophiura and clearly are non-generic. Therefore, we consider O. saurura as species of the genus Ophiura.
Figure 2. (A–D) Ophiura saurura (Verrill, Citation1894): (A) syntype, YPM 9419, RV Albatross, cruise of 1885, St. 2528, 41°47′N, 65°37′30′′W, depth 1232 m, disk diameter 14 mm; (B) IORAS 14.818, RV Akademik Mstislav Keldysh, St. 445, disk diameter 8.2 mm; (C) MAR-ECO, 40/367, disk diameter 17 mm; (D) Amphiophiura convexa (Lyman, 1878), IORAS 14.468, RV Akademik Mstislav Keldysh, St. 500, disk diameter 6.5 mm. (E) Ophiotjalfa vivipara Mortensen, Citation1913, ZMMU D-646, RV Akademik Mstislav Keldysh, St. 464, disk diameter 4 mm. Juvenile within genital slit indicated by arrow. (F) Ophiotypa simplex Koehler, Citation1897, IORAS 14.193, RV Akademik Mstislav Keldysh, St. 500, disk diameter 4 mm. Dorsal and ventral views are shown. Photographs: Alexander Martynov (A); Tanya Korshunova (B, D–F); Konstantin Tabachnik (C).
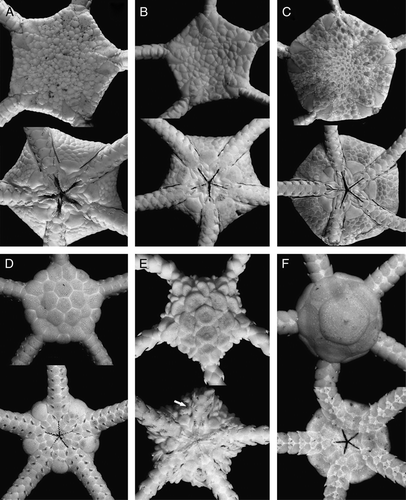
Amphiophiura convexa (Lyman, 1878)
(D)
Ophioglypha convexa: Lyman 1878: 84, Plate 3, Figures 83, 84.
Amphiophiura convexa: Matsumoto Citation1915: 77.
Amphiophiura bullata convexa: Paterson Citation1985: 132–3, Figure 51.
Material
RV G.O. Sars, MAR-ECO cruise, St. 64/381, two specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 500, two specimens; 46th cruise, St. 4226, three specimens.
Distribution
Atlantic, Pacific and Indian Oceans. North Atlantic records: Labrador Basin, Rockall Trough, Bay of Biscay, off the Azores, at 1997–6480 m (Litvinova Citation1971; Paterson Citation1985). Present study: Mid-Atlantic Ridge, south of Charlie-Gibbs Fracture Zone and the Biscay Bay region, at 3452–4297 m.
Ophiotjalfa vivipara Mortensen, Citation1913
(E)
Ophiotjalfa vivipara: Mortensen Citation1913: 40, Figures 4–6; Paterson Citation1985: 139, Figure 53.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 446, one specimen; St. 464, one specimen; St. 478, three specimens.
Distribution
Previously known only from two records – in the Davis Strait and off south-west Iceland, at 1438 m (Mortensen Citation1913, Citation1933). In the present study this species was recorded on the Reykjanes Ridge at 1650–1800 m.
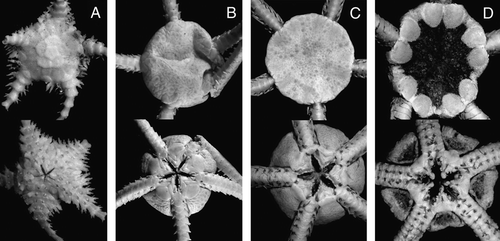
Figure 3. (A) Ophiophycis mirabilis Koehler, 1901, IORAS 14.718, RV Akademik Mstislav Keldysh, St. 526, disk diameter 2.8 mm; (B) Ophiopleura inermis (Lyman, 1878), MAR-ECO, St. 53/375, disk diameter 8 mm; (C) Ophiocten hastatum Lyman, 1878, ZMMU D-730, RV Akademik Mstislav Keldysh, St. 390, disk diameter 14 mm; (D) Ophiernus vallincola Lyman, 1878, ZMMU D-731, RV Akademik Mstislav Keldysh, St. 415, disk diameter 13 mm. Dorsal and ventral views are shown. Photographs: Alexander Martynov (A); Konstantin Tabachnik (B); Tanya Korshunova (C, D).
Ophiotypa simplex Koehler, 1897
(F)
Ophiotypa simplex Koehler 1897: 281–3, Plate 5, Figures 1–3; Paterson Citation1985: 144–5, Figure 55.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 464, two specimens; St. 500, two specimens.
Distribution
Atlantic, Pacific and Indian Oceans. North Atlantic records: off the Azores and Cape Verde at 3595–4366 m (Paterson Citation1985). Present study: the Reykjanes Ridge and the Bay of Biscay at 1670–4297 m.
Remarks
The present findings are the most northern record for the species.
Ophiophycis mirabilis Koehler, 1901
(A)
Ophiophycis mirabilis Koehler Citation1901a: 222, Figures 1, 2; Paterson Citation1985: 141, Figure 54.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 445, one specimen; St. 478, one specimen; St. 489, five specimens; St. 526, one specimen.
Distribution
Prior to this study the species was recorded only from the Bay of Biscay at 1175–1662 m (Paterson Citation1985). Present study: the Reykjanes Ridge and off Portugal, 920–1860 m.
Ophiopleura inermis (Lyman, 1878)
(B)
Ophioglypha inermis: Lyman 1878: 95, Plate 5, Figures 123–125.
Ophiopleura inermis: Gage et al. Citation1983: 295; Paterson Citation1985: 128, Figure 48.
Material
RV G.O. Sars, MAR-ECO cruise, St. 53/375, one specimen.
Distribution
North Atlantic: off Martha's Vineyard, New England north to western Greenland, east of Faeroe Islands, south of the Azores, at 150–1740 m (Paterson Citation1985). Present study: the Mid-Atlantic Ridge south of the Charlie-Gibbs Fracture Zone, at 981–1003 m.
Ophiocten hastatum Lyman, 1878
(C)
Ophiocten hastatum: Lyman 1878: 103, Plate 5, Figures 133, 134; Paterson et al. Citation1982: 117–9, ; Paterson Citation1985: 129, Figure 49.
Material
RV G.O. Sars, MAR-ECO cruise, St. 42/368, one specimen; St. 44/369, one specimen; St. 72/368, two specimens; St. 268, eight specimens; RV Akademik Mstislav Keldysh, fourth cruise; St. 316, 11 specimens; St. 317, one specimen; St. 390, 68 specimens; St. 445, one specimen; St. 465, five specimens.
Distribution
Atlantic, Pacific and Southern Oceans, at depths of 1600–4700 m. North Atlantic records: off Rockall Trough, south of the Azores, at 1970–4700 m (Paterson et al. Citation1982). Present study: the Reykjanes Ridge and the Mid-Atlantic Ridge south of the Charlie-Gibbs Fracture Zone, the Mid-Atlantic Ridge, north of the Azores, at 1620–2951 m.
Ophiernus vallincola Lyman, 1878
(D)
Ophiernus vallincola Lyman 1878: 122, Plate 6, Figures 170–172.
Ophiernus abyssalis Koehler 1896b: 242.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 415, one specimen; St. 445, 12 specimens; St. 464, one specimen; St. 465, two specimens.
Distribution
East Atlantic, from south of Ireland to off South Africa, Indian Ocean, western Pacific, at 840–4065 m (Madsen Citation1977). Present study: the Reykjanes Ridge, at 1535–1860 m.
Ophioplinthus tessellata (Verrill, Citation1894) comb. nov.
(Figures 4A, C, 5A–C)
Ophioglypha tessellata: Verrill 1894: 290.
Homalophiura tessellata H.L. Clark Citation1915: 327.
Homophiura tessellata: Paterson Citation1985: 137–8, Figure 52.
Material
RV G.O. Sars, MAR-ECO cruise, St. 50/373, 11 specimens; St. 53/375, 47 specimens; St. 54/377, three specimens; St. 72/386, one specimen.
Distribution
North Atlantic: off New England to Labrador Basin, southern Iceland to Azores, at 433–4706 m (Paterson Citation1985). Present study: the Charlie-Gibbs Fracture Zone, the Mid-Atlantic Ridge south of the Charlie-Gibbs Fracture Zone and north of the Azores at 981–3527 m. It is interesting to note that despite numerous samples on the Reykjanes Ridge during the fourth cruise of Akademik Mstislav Keldysh this species was not found.
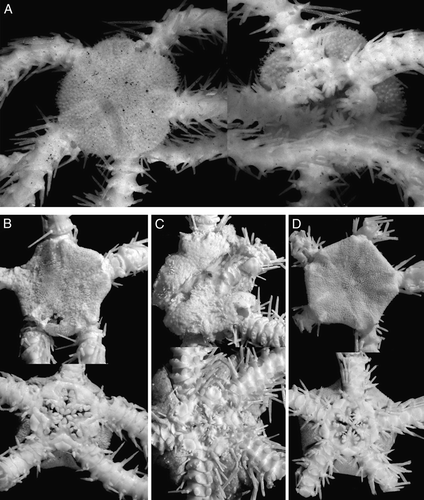
Figure 4. (A) Ophioplinthus tessellata (Verrill, Citation1894), MAR-ECO, St. 53/375, disk diameter 17 mm; (B) Ophioplinthus pseudotessellata sp. nov., holotype ZMBN 77852, MAR-ECO, St. 40/367, disk diameter 17.5 mm; (C) Ophioplinthus tessellata (Verrill, Citation1894), MAR-ECO, St. 53/375, disk diameter 10 mm; (D) Ophioplinthus pseudotessellata sp. nov., paratype ZMBN 77853, MAR-ECO, St. 40/367, disk diameter 7 mm. Dorsal and ventral views are shown. Photographs: Alexander Martynov (A–C); Konstantin Tabachnik (D).
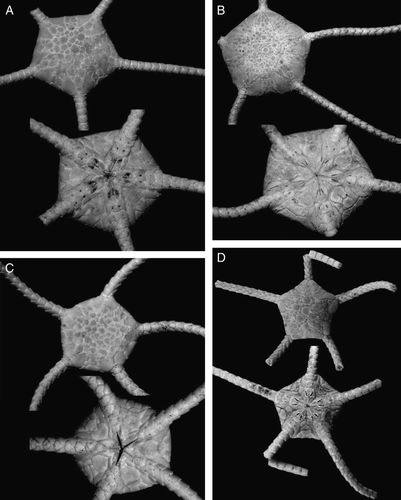
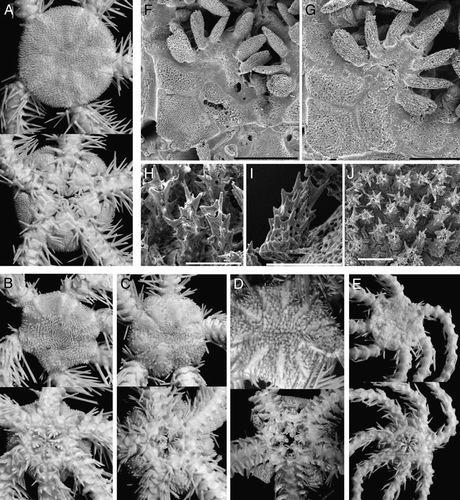
Figure 5. (A–C) Ophioplinthus tessellata (Verrill, Citation1894): (A) lectotype YPM 39095, RV Albatross, cruise of 1886, St. 2710, off Nantucket I., 40°06′00′′N, 68°01′30′′W, depth 1800 m, disk diameter 19 mm, dorsal and ventral views; (B) paralectotype USNM 9070, RV Albatross, St. 2083, Massachusetts, Georges Bank, 40°26′40′′N, 67°05′15′′W, depth 1754, disk diameter 20 mm, dorsal and ventral views; (C) MAR-ECO, St. 50/373, disk diameter 19 mm, dorsal view. (D–G) Ophioplinthus pseudotessellata sp. nov.: (D) paratype ZMMU D-749, MAR-ECO, St. 40/367, disk diameter 17.5 mm, dorsal view; (E) scanning electron micrograph of lateral arm plate, basal segments, showing arm spine articulation ridges; (F) scanning electron micrograph of lateral arm plate, basal segments; (G) scanning electron micrograph of dorsal arm plate, basal segments. Scale bars: 300 µm (E–G). Photographs: Eric Lazo-Wasem (A); Cynthia Ahearn (B); Alexander Martynov (C–G).

Remarks
Ophiurids with strongly reduced arm comb and tentacle pores restricted only to the few proximal segments have a long and tangled taxonomic history. Lyman (1878, 1882) erected the genus Ophioplinthus for two species with the above-mentioned characters. However, Lyman considered other species with similar characters (e.g. Ophioglypha inornata Lyman, 1878, O. abyssorum Lyman, 1883) to be within his large genus Ophioglypha (now a synonym of the genus Ophiura). Several Ophioplinthus-like species were described by Studer (1876) and Koehler (Citation1901b, Citation1908 Citation1911) also within the genus Ophioglypha. In 1915, Matsumoto and H.L. Clark created two genera, Ophiurolepis and Homalophiura, almost simultaneously. The type species of these genera are different, but both have reduced arm combs and tentacle pores restricted to proximal arm segments. Clark (Citation1915) listed a number of species previously assigned to the genus Ophiura (or Ophioglypha) under the generic name Homalophiura. Hertz (Citation1927a Citationb) for the first time suggested that all three genera, Ophioplinthus, Ophiurolepis and Homalophiura, be united under the same name, using the oldest name Ophioplinthus. Despite this Hertz (Citation1927a Citationb), as well as Koehler (Citation1922, Citation1923), for most Antarctic species of this group, used the generic name Ophiurolepis. Later this decision was widely accepted (Mortensen Citation1936; Fell Citation1961; Madsen Citation1967; McKnight Citation1967; Bernasconi & D'Agostino Citation1975; Smirnov Citation2006). Bartsch (Citation1982) gave a short historical overview of the taxonomy of the Homalophiura–Ophiurolepis group and placed H. inornata (type species of the genus Homalophiura) within the genus Ophiurolepis. In addition, Madsen (Citation1967) listed without discussion Homalophiura inornata (Lyman, 1878) under the generic name Ophiurolepis and thus acted as the first reviser. Fell (Citation1961) described one more genus with tentacle pores restricted only to proximal segments of the arms, Theodoria. This genus is distinguished from Ophiurolepis mainly by the degree of development of the tentacle scales and excavated jaws.
Figure 6. (A) Ophioplinthus medusa Lyman, 1878, syntype, MCZ 378, HMS Challenger, St. 156, Antarctic, off Kaiser Wilhelm II Land, depth 3594 m, disk diameter 12.5 mm; (B) Ophioplinthus abyssorum (Lyman, 1883), holotype, MCZ 532, Blake, St. 140, off Virgin Gorda, depth 1997 m, disk diameter 11 mm; (C) Ophioplinthus grisea Lyman, 1878, syntype, MCZ 377, HMS Challenger, St. 156, Antarctic, off Kaiser Wilhelm II Land, depth 3594 m, disk diameter 18 mm; (D) Ophioplinthus confragosa (Lyman, 1878), flattened specimen, ZMMU D-733, RV Akademik Kurchatov, cruise 11, St. 928, 52°15′S, 56°51′W, off Falkland Is., depth 1105 m, disk diameter 13 mm; (E) Ophioplinthus carinata (Studer, Citation1876), IORAS 14.672, RV Ob’, St. 122, Kerguelen I., disk diameter 18 mm; (F) Ophioplinthus confragosa (Lyman, 1878), swollen specimen, ZMMU D-733, RV Akademik Kurchatov, cruise 11, 52°15′S, 56°51′W, off Falkland Is., depth 1105 m, disk diameter 15 mm. (G) Ophioplinthus inornata (Lyman, 1878), syntype, MCZ 611, HMS Challenger, St. 106, tropical Atlantic, depth 3367 m, disk diameter 8 mm; (H) Ophiomusium lymani Wyville Thompson, 1873, ZMMU D-744, RV Akademik Mstislav Keldysh, St. 385, disk diameter 19 mm. Dorsal and ventral views are shown. Photographs: Alexander Martynov (A, C); Jessica Cundiff (B, G); Tanya Korshunova (D–F, H).
Paterson (Citation1985) briefly reviewed the genus Homalophiura and divided it into four informal groups. Paterson argued that species with reduced arm comb and tentacle pores restricted only to the proximal segments belonged to several different genera. According to Paterson, the genus Ophiurolepis included species with short genital slits (including the type species of the genus Homalophiura, H. inornata), whereas for species with long genital slits (Homalophiura tessellata, H. abyssorum) Paterson created a new genus Homophiura. Other species from the former genus Homalophiura were referred to the genus Ophiura s.l. The relation between Homalophiura, Ophiurolepis, Homophiura and Ophioplinthus was not investigated. According to the present study, the length of the genital slits is not a very reliable character. First of all, the type species of the genus Ophiurolepis, O. carinata (Studer, Citation1876) (E), and most of the other species of the genus Ophiurolepis have long genital slits. Secondly, the type species of the genus Homalophiura, H. inornata (Lyman, 1878) (G) (according to illustration given by Lyman Citation1882: Plate 3, ), and the type species of the genus Homophiura, H. tessellata (Figures 4A, C, 5A–C) both have long genital slits. In general, it is hard to evaluate an exact length of the genital slits due to closing of the distal part, and this may cause misinterpretation. For instance, Lyman (1882) figured the type species of the genus Ophioplinthus, O. medusa, as having short genital slits. However, re-examination of the type material of O. medusa (A) revealed the presence of long genital slits at least in one interradius, whereas in other interradii the genital slits are closed distally. Thus, there are no real differences in length of the genital slits (according to Paterson Citation1985 one of the main diagnostic characters) between type species of the genera Ophioplinthus, Homalophiura, Ophiurolepis and Homophiura. Homophiura Paterson, Citation1985, therefore, is a clear synonym of the genus Homalophiura H.L. Clark, Citation1915. The latter, in turn does not have reliable differences from Ophioplinthus Lyman, 1878. Bartsch (Citation1991) did not support the genus Homophiura and maintained H. abyssorum (Lyman, 1883) within the genus Homalophiura.
Many authors mention Ophiurolepis as closely related to Homalophiura and Ophioplinthus (Hertz Citation1927a Citationb; Mortensen Citation1933, Citation1936; Paterson Citation1985). Apart from the tentacle pores being restricted to the proximal segments, Ophiurolepis can be distinguished by a round swollen disk, relatively few (compared to Ophioplinthus) distinct large disk plates including quite distinct primary plates in a rosette pattern, relatively large radial shields, slender arms, carinate dorsal arm plates and a fragmented oral shield. At first glance, all these characters taken together distinguish Ophiurolepis from Ophioplinthus. However, if all species referred to Ophiurolepis are considered, the above-mentioned characters turn into a tendency rather than a clear separation. For instance, some species do not have carinate dorsal arm plates and fragmented oral shield (e.g. O. martensi (Studer, 1885)). We examined a number of specimens of Homalophiura confragosa (Lyman, 1878), the species with unclear generic placement, collected off the Falkland Islands. Some specimens of this species had round and conspicuously swollen disks (F) and were similar to species of the genus Ophiurolepis, whereas other specimens, with round or pentagonal and flattened disks (D) did not differ from species of the genera Ophioplinthus and Homophiura. On the other hand, some small specimens of the type species of the genus Homophiura, H. tessellata, have quite well-developed distinct disk plates and weakly carinated dorsal arm plates. The type species of the genus Homalophiura, H. inornata (G), has quite distinct disk plates and thick dorsal arm plates with a weak ridge, similar to the species of the genus Ophiurolepis with well-developed dorsal ridges, as well as relatively small oral papillae characteristic for the type species of the genera Ophioplinthus and Homophiura. An additional character mentioned by Paterson as separating Homophiura from Ophiurolepis is the fragmented oral and adoral shields in the latter genus. Despite this, in the present material a new species Ophioplinthus pseudotessellata sp. nov. (Figures 4B, D, 5D–G), closely related to the type species of the genus Homophiura, H. tessellata, has partially fragmented oral and adoral shields. Thus Homalophiura inornata can be equally referred to the genera Homophiura or Ophiurolepis.
Moreover, the genus Theodoria Fell, Citation1961 is closely related to the genus Ophiurolepis in most characters including distinct dorsal plates of the disk. A difference of the type species of the genus Theodoria, T. relegata (Koehler, Citation1922), from the type species of the genus Ophiurolepis, O. carinata (and from the genus Ophioplinthus), is well-defined proximal tentacle pores and tentacle scales. However, this character is not stable. Even among species referred to the genus Theodoria, for instance T. wallini (Mortensen, 1925), tentacle scales are much less developed than in the type species, and are more similar to those in some species of the genus Ophiurolepis. At the same time, in one species, initially included by Lyman in his genus Ophioplinthus, O. grisea Lyman, 1878 (C), tentacle scales are well developed. The general shape of the disk and absence of distinct plates in O. grisea separates it from Theodoria and Ophiurolepis. Thus, O. grisea by morphology of the tentacle scales is similar to Theodoria, whereas the shape of the disk corresponds to the type species of the genus Ophioplinthus. Similarly, Ophiurolepis scissa (Koehler, Citation1908) has an intermediate position between Theodoria and Ophiurolepis based on the degree of development of tentacle scales. Homalophiura nana (Lütken & Mortensen, Citation1899) and H. madseni Belyaev & Litvinova, Citation1972 have quite well-developed tentacle scales. Homalophiura abyssorum (Lyman, 1883) (B) in the structure and shape of disk is similar to H. madseni, but has strongly reduced tentacle scales. Another character, mentioned as a distinguishing feature of Theodoria, is the excavated jaws. Despite the fact that many species of the Ophioplinthus–Ophiurolepis group have rather flattened jaws, we found distinctly excavated jaws in small specimens of Ophioplinthus pseudotessellata sp. nov. (D), whereas one species of Theodoria, T. wallini, has somewhat flattened jaws. One of the species originally included by Lyman in the genus Ophioplinthus, O. grisea, also has slightly excavated jaws. Finally, Homalophiura madseni has clearly excavated jaws.
Summarizing, it should be noted that the degree of development of the tentacle scales varies considerably among different species from the genera Ophioplinthus, Homalophiura and Ophiurolepis. This means that Theodoria cannot remain as a separate genus. Even if we try to retain the genus Theodoria as independent, including only species with more or less well-defined tentacle scales, there is no evidence of monophyly.
A characteristic feature of the genus Ophiura is a rapid decrease in size of the tentacle pores and number of tentacle scales in the distal direction. Paterson (Citation1985): 135), discussing the taxonomy of the genus Homalophiura, noted that ‘this character is more widespread…’. In fact, the most characteristic feature of the Ophioplinthus–Ophiurolepis group is a reduction of distal tentacle pores. This tendency is most pronounced in the genus Ophioplinthus and it is present in the genus Ophiura. There is a possibility that species similar in morphology to the group Ophioplinthus–Ophiurolepis may have evolved several times from different species of the genus Ophiura by means of further reduction of the arm comb and tentacle pores on distal segments. Thus, it is necessary to have additional characters for confirmation or for disproof of the monophyly of the Ophioplinthus–Ophiurolepis group. One such character could be the position of the second tentacle pore. Species of the genus Ophiura with reduced arm comb and reduced tentacle scales, like O. maculata, have the second tentacle pore rather opened inside the mouth between the jaws. At the same time, in species of the genera Ophioplinthus, Ophiurolepis, Theodoria and Homophiura, as well as in the type species of the genus Homalophiura, the second tentacle pore does not open inside the mouth and is nearly completely enclosed by the jaw. This character corresponds to the shape of the jaws: in the genus Ophiura the jaws have nearly parallel edges distally and rounded edges proximally, whereas in Ophioplinthus, Ophiurolepis, Theodoria and Homophiura the jaws are triangular.
Thus, Ophioplinthus, Homalophiura, Ophiurolepis, Theodoria and Homophiura represent a single morphological unit supported by several characters. The type species of the genera Ophioplinthus and Homophiura differ only slightly from each other and these genera are considered here as clear synonyms. From most characters Ophiurolepis is also similar to Ophioplinthus. There are differences, mainly the more distinct and more regular shape of the plates in Ophiurolepis. However, this character is quite hard to use as diagnostic since it is difficult to determine the degree of the distinctness of disk plates. As already mentioned, Homalophiura inornata can equally belong to the genus Homophiura or the genus Ophiurolepis.
In addition, some species, for instance Ophiurolepis martensi (Studer, 1885), Homalophiura abyssorum (Lyman, 1883), Homalophiura confragosa (Lyman, 1878) and Homalophiura madseni Belyaev & Litvinova, Citation1972, are hardly unambiguously referred to Ophioplinthus or Ophiurolepis. An interesting ecological characteristic of Ophioplinthus sensu lato are commensal hydroids recorded on e.g. O. medusa, O. tessellata and O. relegata (Lyman Citation1882; Svoboda et al. Citation1995). These species earlier were included in the genera Ophioplinthus, Homophiura and Theodoria. Most recently, Hunter (Citation2007) has presented a phylogenetic analysis based on morphological characteristics of the genus Ophiurolepis. According to this study genus Ophiurolepis is not a monophyletic group comprising from three clades including species from other closely related genera. For instance, Ophiurolepis scissa is more closely related to Homalophiura confragosa and H. inornata than to other species of the genus Ophiurolepis. These data also support the taxonomic division suggested here.
In summary, we consider Homalophiura H.L. Clark, Citation1915, Ophiurolepis Matsumoto, Citation1915, Theodoria Fell, Citation1961 and Homophiura Paterson, Citation1985 as synonyms of the genus Ophioplinthus Lyman, 1878. Lyman (1878) used the name Ophioplinthus as a noun of the feminine gender. The following species we consider within the genus Ophioplinthus Lyman, 1878:
Ophioplinthus abyssorum (Lyman, 1883) comb. nov.
Ophioplinthus accomodata (Koehler, Citation1922) comb. nov.
Ophioplinthus anceps (Koehler, Citation1908) comb. nov.
Ophioplinthus banzarei (Madsen, Citation1967) comb. nov.
Ophioplinthus brevirima (Mortensen, Citation1936) comb. nov.
Ophioplinthus brucei (Koehler, Citation1908)
Ophioplinthus carinata (Studer, Citation1876) comb. nov.
Ophioplinthus clasta (H.L. Clark, Citation1911) comb. nov.
Ophioplinthus confragosa (Lyman, 1878) comb. nov.
Ophioplinthus divisa (Lütken & Mortensen, 1899) comb. nov.
? Ophioplinthus euryplax (H.L. Clark, 1939) comb. nov.
Ophioplinthus frigida (Koehler, 1901) comb. nov.
Ophioplinthus gelida (Koehler, 1901) comb. nov.
? Ophioplinthus glypta (H.L. Clark, 1939) comb. nov.
Ophioplinthus granulifera (Bernasconi & D'Agostino, 1973) comb. nov.
Ophioplinthus grisea Lyman, 1878
Ophioplinthus inflata (Koehler, Citation1897) comb. nov.
Ophioplinthus inornata (Lyman, 1878) comb. nov.
Ophioplinthus intorta (Lyman, Citation1878) comb. nov.
Ophioplinthus madseni (Belyaev & Litvinova, Citation1972) comb. nov.
Ophioplinthus medusa Lyman, Citation1878
Ophioplinthus martensi (Studer, 1885) comb. nov.
Ophioplinthus mordax (Koehler, Citation1922) comb. nov.
Ophioplinthus nexila (Kyte, 1987) comb. nov.
Ophioplinthus olstadi (Madsen, 1955) comb. nov.
Ophioplinthus partita (Koehler, Citation1908) comb. nov.
Ophioplinthus relegata (Koehler, Citation1922) comb. nov.
Ophioplinthus scissa (Koehler, Citation1908) comb. nov.
? Ophioplinthus scutata (Lyman, Citation1883) comb. nov.
Ophioplinthus tessellata (Verrill, 1994) comb. nov.
Ophioplinthus tuberosa (Mortensen, Citation1936) comb. nov.
Ophioplinthus tumescens (Koehler, Citation1922) comb. nov.
Ophioplinthus turgida (Mortensen, Citation1936) comb. nov.
Ophioplinthus wallini (Mortensen, 1925) comb. nov.
Ophioplinthus pseudotessellata sp. nov.
(Figures 4B, D, 5D–G)
Material
RV G.O. Sars, MAR-ECO cruise, St. 40/367, holotype (ZMBN 77852, dried), 27 paratypes (ZMBN 77853, in ethanol), one paratype (ZMMU D-749, dried).
Etymology
Species epithet pseudotessellata (from the Greek pseudos – false – and species epithet tessellata) refers to close similarity of this species to Ophioplinthus tessellata.
Description of holotype
The disk diameter of the holotype is 17.5 mm. It is pentagonal, moderately swollen and slightly concave in the interradii. The central area of the disk is distinctly depressed. The disk scales are numerous (about 150–160 in number), small, but various in size, irregular-rounded in shape. There is a conspicuous primary rosette of six plates, clearly separated from most disk plates by their larger sizes. Radial shields are irregularly pear-shaped in outline, proximally narrowed, distally considerably widened and lie close to each other, but do not touch. Maximum width of radial shields is 3 mm. Along both internal and external edges of the radial shields there are irregular rows of small plates. In some radii these plates are smaller, in others larger. Distally in-between the radial shields few irregularly rectangular plates are placed which together with plates along the internal edges form a more or less evident Y-shaped structure between a pair of radial shields. The surface of the radial shields is smooth. Each jaw bears two or three quite irregularly placed conical or blunt apical oral papillae and seven or eight pairs of rectangular lateral papillae. Distal papillae are more elongated, block-shaped, whereas more proximally placed papillae are rounded. The oral shield is large, irregularly oval or irregularly pear-shaped and fragmented different edges in different interradii: ventrally, dorsally or laterally. The distal edge of the oral shield is bordered by an irregular line of quite small scales. Adoral shields are triangular narrow bands, about three times as long as broad. Between adoral shields and oral shields sometimes an insertion plate appears as an additional adoral shield, whereas in some interradii between adoral shields there are several irregular small plates. Genital slits are long, extending from first to fourth ventral arm plates, but for most of their length tightly closed, therefore appearing as quite short or inconspicuous. The papillae of the arm comb are greatly reduced and very short, but distinguishable. There are several short papillae placed on the distal part of the genital slit, arranged in two more or less distinct rows, up to 10 papillae per row. In some interradii these papillae are almost indistinct. Dorsally few tubercle-shaped papillae are placed at the distal edge of the radial shields. Arms slender, broken, about 1.5 times disk diameter. The dorsal arm plates are small, a few basal segments bear spear-shaped plates, while starting from the third or fifth segments the plates become almost rhomboid and very narrow. The first ventral arm plate is trapezoid. Most ventral arm plates are irregularly rhomboidal, lengthened triangular and widely separated from each other. The two first plates are nearly contiguous. Distal ventral arm plates are irregularly rhombic or wide spear-shaped, and widely separated by lateral plates. Lateral arm plates are massive. Arm spines on most segments are absent and eroded, more or less retained only on few basal segments. Spines generally are four (in a single segment five), extremely short and conical. The ventral spine on more distal segments (starting approximately from 15th segment) is considerably longer than the others and also longer than the corresponding ventral spine on the proximal segments. Arm spine articulation ridges are separated from each other by conspicuous elevations and more dorsally placed ridges are separated from each other by a distinct gap, while ventral ridges are closer together. The second tentacle pore is slit-shaped and surrounded by two rows of four to seven tentacle scales. The lateral plate has smaller and more numerous square scales, whereas the rows on the ventral plate include few larger almost oval swollen scales. The same pattern of two different rows of tentacle scales is found on three following basal segments but to a considerably lesser degree. The third segment has three scales on the lateral plate and three to four on the ventral plate. Fourth and fifth segments usually have one or two on the lateral plate and two or three on the ventral plate. Starting from the sixth segment there are no tentacle pores and tentacle scales.
Paratype variations
Generally, all paratypes, including small specimens, are essentially similar to the holotype. Small paratypes of Ophioplinthus pseudotessellata sp. nov. have about 100 dorsal plates on the disk and even more narrow, almost ovoid dorsal arm plates. The tentacle scales of the small specimens are more distinct than in the holotype. Large paratypes have a more flattened disk and more distinct disk plates. Tentacle scales are more numerous – up to eight scales at the second pore, up to five, usually four scales at the third pore.
Distribution
Mid-Atlantic Ridge north of the Azores, at 2954–2968 m.
Remarks
Ophioplinthus pseudotessellata sp. nov. is clearly different from the sympatric congener Ophioplinthus tessellata in having more numerous and smaller plates on the disk (comparing specimens of similar size), small narrow rhomboid dorsal arm plates and partially fragmented oral and adoral shields. For comparative purposes type series from different stations of O. tessellata from USNM and YPM were studied. All six studied specimens ranging 10–21 mm in disk diameter invariably have wide dorsal arm plates, whereas both small and large specimens of Ophioplinthus pseudotessellata sp. nov. have very narrow dorsal arm plates. The shape of the dorsal arm plate is thus the most reliable character for distinguishing Ophioplinthus pseudotessellata sp. nov. from O. tessellata. Small specimens of Ophioplinthus pseudotessellata sp. nov. differ even more in this character from O. tessellata of comparable size in having very narrow, almost ovoid dorsal arm plates (compare C and D). Large specimens of Ophioplinthus pseudotessellata sp. nov. (17.5 and 18.5 mm disk diameter) have 150–160 dorsal disk plates, whereas large specimens of O. tessellata 19 mm disk diameter as well as a paralectotype of O. tessellata (USNM 9070) 21 mm disk diameter have only about 110–120 dorsal plates on the disk (compare Figures 4B, 5D and 4A, 5A–C). The small paratypes of Ophioplinthus pseudotessellata sp. nov. (disk diameter 7 mm) have 100 dorsal disk plates, whereas small Ophioplinthus tessellata (disk diameter 10 mm) have 70–75 dorsal disk plates. To avoid further confusion between these two closely related species we designate here a lectotype of O. tessellata – specimen YPM 39095. Paralectotypes are correspondingly YPM 9246 (nine specimens), YPM 39096 (one specimen) and USNM 8094 (three specimens), USNM 8124 (three specimens), USNM 9070 (one specimen), USNM 9172 (one specimen), USNM 11522 (one specimen), USNM 11523 (three specimens), USNM 14993 (one specimen), USNM 21231 (two specimens), USNM 24735 (one specimen), USNM 27888 (one specimen). The only other species of Ophioplinthus known from the North Atlantic is O. abyssorum (Lyman, 1883) (B). This species (including variety O. abyssorum var. africana Madsen, 1947) differs clearly from Ophioplinthus pseudotessellata sp. nov. in a significantly smaller number of dorsal plates (50–60 in syntype 11.5 mm disk diameter), the shape of the radial shields, the widely rhombic shape of the dorsal arm plates, not fragmented oral and radial shields, and much less developed tentacle scales at the second tentacle pore. Some species of the genus Ophioplinthus, e.g. the type species O. medusa as well as O. clasta, and O. madseni are similar to O. pseudotessellata sp. nov. in having very narrow dorsal arm plates but differ considerably in a number of other characters (e.g. by general appearance of the disk and shape of the dorsal arm plates in O. medusa, thick skin of the disk of O. clasta and well-developed tentacle pores in O. madseni).
Ophiomusium lymani Wyville Thompson, 1873
(H)
Ophiomusium lymani Wyville Thomson 1873: 174; Paterson Citation1985: 147–8, Figure 58. Ophiomusa lymani: Hertz Citation1927a: 103–5.
Material
RV G.O. Sars, MAR-ECO cruise, St. 44/369, five specimens; St. 68/384, one specimen; St. 70/385, 14 specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 316, 11 specimens; St. 317, nine specimens; St. 319, 20 specimens; St. 354, three specimens; St. 385, one specimen; St. 415, 89 specimens; St. 418, 26 specimens; St. 445, 36 specimens; St. 446, 11 specimens; St. 464, seven specimens; St. 465, five specimens; St. 478, one specimen; St. 489, one specimen; St. 492, two specimens; St. 499, two specimens.
Distribution
Atlantic, Pacific and Indian Oceans. North Atlantic records: from West Indies to Davis Strait and from south-west Iceland to south of Cape Blanc, at 651–4829 m (Paterson Citation1985). Present study: the Reykjanes Ridge, the Charlie-Gibbs Fracture Zone, the Mid-Atlantic Ridge north of the Azores, at 1535–2440 m.
Family Ophiacanthidae
Ophiacantha aculeata Verrill, 1885
(Figures 7A, 10)
Ophiacantha aculeata: Verrill 1885a: 153; Verrill Citation1885b: 547; Koehler Citation1914: 74–7, Plate 11, Figures 1, 2; Paterson Citation1985: 38–9, .
Material
RV G.O. Sars, MAR-ECO cruise, St. 40/367, two specimens; St. 50/373, one specimen; St. 52/374, one specimen; RV Akademik Mstislav Keldysh, fourth cruise, St. 316-5, 108 specimens; St. 317, three specimens; St. 445, two specimens; St. 499, 172 specimens.
Type and historically important material studied. USNM 6610, 11 specimens identified by M. Rathbun, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
USNM 12669, lectotype, here designated, specimen identified by R. Koehler, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
USNM 27900, one specimen identified by A.E. Verrill, Steamer Albatross, cruise of 1886, St. 2725, Virginia Beach, 36°34′00′′N, 73°48′00′′W, depth 2513 m.
USNM 1100627, three paralectotypes, specimens identified by R. Koehler, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
Distribution
North Atlantic: from off Virginia to New England, east of Rockall Trough, Bay of Biscay, Iberian Basin, at 2500–3584 m (Verrill 1885a,b; Paterson Citation1985). Present study: the Reykjanes Ridge, Mid-Atlantic Ridge north of the Azores, at 1650–2979 m.
Remarks
See discussion on Ophiacantha fraterna below.
Ophiacantha anomala G.O. Sars, Citation1871
(B–J)
Figure 7. (A) Ophiacantha aculeata Verrill, 1885, ZMMU D-641, RV Akademik Mstislav Keldysh, St. 499, disk diameter 9 mm. (B, C, E–J) Ophiacantha anomala G.O. Sars, Citation1871: (B) ZMMU D-661, six-armed specimen, RV Akademik Mstislav Keldysh, St. 464, disk diameter 9 mm; (C) ZMMU D-658, five-armed specimen, RV Akademik Mstislav Keldysh, St. 464, disk diameter 9 mm; (E) ZMMU D-659, seven-armed specimen, RV Akademik Mstislav Keldysh, St. 478, disk diameter 3.1 mm; (F) ZMMU D-658, five-armed specimen, RV Akademik Mstislav Keldysh, St. 464, scanning electron micrographs of oral structures; (G) ZMMU D-659, six-armed specimen, RV Akademik Mstislav Keldysh, St. 478, scanning electron micrographs of oral structures; (H, J) ZMMU D-659, six-armed specimen, RV Akademik Mstislav Keldysh, St. 478, scanning electron micrographs of spicules from dorsal side of the disk; (I) ZMMU D-658, five-armed specimen, RV Akademik Mstislav Keldysh, St. 464, scanning electron micrographs of spicules from dorsal side of the disk. (D) Syntype MCZ 1942 of Ophiacantha cuspidata Lyman, 1878, HMS Challenger, St. 344, Ascension I., depth 764 m, disk diameter 10 mm. Dorsal and ventral views are shown in (A–E). Scale bars: 1 mm (F); 600 µm (G); 150 µm (H, J); 200 µm (I). Photographs: Tanya Korshunova (A–C, E); Alexander Martynov (D, F–J).
Ophiacantha anomala: G.O. Sars 1871: 12; Paterson Citation1985: 24–5, 34, .
Ophiacantha cuspidata: Lyman 1878: 143, Plate 10, Figures 248–250, syn. nov.
Material
RV G.O. Sars, MAR-ECO cruise, St. 65/382, two specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 385, 17 specimens; St. 418, one specimen; St. 446, four specimens; St. 464, 15 specimens; St. 478, 55 specimens.
Distribution
Ascension I., 785 m (Lyman 1878, type locality of O. cuspidata), North Atlantic: from Florida to south-west Iceland and from east Iceland to Norway and Rockall Trough, at 141–1500 m (Paterson Citation1985) and off Morocco, 1750 m (Bartsch Citation1987, as O. cuspidata). Present study: the Reykjanes Ridge, the Charlie-Gibbs Fracture Zone at 607–1915 m.
Remarks
Mortensen (Citation1933) mentioned two findings of five-armed O. anomala collected off southern Iceland and from Trondheimsfjord. Moreover, Mortensen also mentioned six-armed juveniles in the bursae of the five-armed specimens. Despite this, Paterson (Citation1985) strictly distinguished five- and six-armed species of the genus Ophiacantha in the North Atlantic. New material from the Reykjanes Ridge (geographically close to one of Mortensen's specimens) includes apparently one species, though with a different number of arms – five or six. The general appearance, shape of the spicules of the disk, oral shield and oral papillae, and tentacle scale are the same in five- and six-armed specimens (compare C, F, I and 7B, G, H). Both five- and six-armed specimens fit the diagnosis of O. anomala well. At the same time, five-armed O. cuspidata are very similar to O. anomala and five-armed specimens of O. anomala have never been mentioned as a synonym of O. anomala. We examined two paratypes of O. cuspidata from the Museum of Comparative Zoology (Harvard University) (D). They do not differ from the five-armed O. anomala, therefore O. cuspidata is considered here as a synonym of O. anomala. We also discovered one seven-armed juvenile of O. anomala (E). The six-armed specimens differ from the five-armed specimens in slightly narrower jaws and oral shields because development of the additional arm results in tighter packing of the oral structures.
Ophiacantha fraterna Verrill, 1885
(Figures 8A, B, F, H, 9, 11A)
Ophiacantha fraterna: Verrill Citation1885b: 545; Koehler Citation1914: 83–6, Plate 11, Figures 5, 6.
Ophiacantha bidentata var. fraterna Mortensen Citation1933: 21–2, Figures 6, 7.
? Ophiacantha hibernica: Farran Citation1913: 39, Figures 10, 11.
= Ophiacantha bidentata auct. non Retzius, 1805.
Material
RV G.O. Sars, MAR-ECO cruise, St. 60/379, one specimen; St. 68/384, one specimen; St. 70/385, one specimen; RV Akademik Mstislav Keldysh, fourth cruise, St. 316, 72 specimens; St. 317, three specimens; St. 415, 18 specimens; St. 418, two specimens; St. 445, 19 specimens; St. 464, three specimens; St. 478, one specimens; St. 499, 10 specimens.
Figure 8. (A) Ophiacantha fraterna Verrill, 1885, ZMMU D-635, RV Akademik Mstislav Keldysh, St. 415, disk diameter 5.7 mm; (B) Ophiacantha fraterna Verrill, 1885, YPM 7241, specimen identified by A.E. Verrill, Steamer Albatross, St. 2562, off New Jersey, depth 2609 m, disk diameter 5.5 mm; (C) Ophiacantha bidentata (Retzius, 1805), syntype of Ophiacantha spinulosa Müller & Troschel, 1842, SMNH-Type-3289, Svalbard, West Spitsbergen, probably collected by Sven Lovén, no date, no depth, disk diameter about 15 mm; (D) Ophiacantha bidentata (Retzius, 1805), syntype of ‘Ophiocoma’ arctica Müller & Troschel, 1842, SMNH-Type-5144, Svalbard, West Spitsbergen, probably collected by Sven Lovén, no date, no depth, disk diameter 10 mm; (E) Ophiacantha bidentata (Retzius, 1805), ZMMU D-727, RV Sergei Vavilov, Cruise 13, St. 1132, Barents Sea, 70°22.6′N, 55°37.1′E, depth 80–109 m, disk diameter 6.5 mm; (F) Ophiacantha fraterna Verrill, 1885, ZMMU D-635, RV Akademik Mstislav Keldysh, St. 415, disk diameter 7.2 mm, scanning electron micrograph of the spicules from dorsal side of the disk; (G) Ophiacantha bidentata (Retzius, 1805), ZMMU D-728, RV Sergei Vavilov, Cruise 11, St. 1035, Barents Sea, 70°22′N, 57°39.1′E, depth 70–90 m, disk diameter 7.5 mm, scanning electron micrograph of the spicules from dorsal side of the disk; (H) Ophiacantha fraterna Verrill, 1885, ZMMU D-635, RV Akademik Mstislav Keldysh, St. 415, scanning electron micrograph of the oral structures and proximal arm segments; (I) Ophiacantha bidentata (Retzius, 1805), ZMMU D-750, RV Sevastopol, cruise 5, St. 1238, 71°00′N, 12°56′W, depth 380 m, scanning electron micrograph of the oral structures and proximal arm segments. Dorsal and ventral views are shown in (A–E). Scale bars: 150 µm (F, G); 1 mm (H, I). Photographs: Tanya Korshunova (A, E); Alexander Martynov (B, F–I); Sabine Stöhr (C, D).
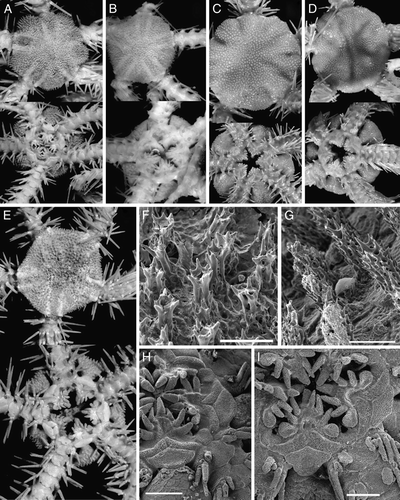
Figure 9. Ophiacantha fraterna Verrill, 1885. (A) USNM 7905, specimen identified by A.E. Verrill, Steamer Albatross, cruise of 1884, St. 2206, New Jersey, Hudson Canyon, 39°35′00′′N, 71°24′30′′W, depth 1907 m, disk diameter 6.5 mm; (B) lectotype USNM 33816, specimen identified by R. Koehler, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m, disk diameter 6.5 mm; (C) USNM 24783, specimen identified by A.E.Verrill as Ophiacantha aculeata, Steamer Albatross, cruise of 1883, St. 2042, Massachusetts, SE of Nantucket Shoals, 39°33′00′′N, 68°26′45′′W, depth 2844, disk diameter 7.5 mm. Dorsal and ventral views are shown. Photographs: Cynthia Ahearn.
Type and historically important material studied. Ophiacantha spinulosa Müller & Troschel, 1842: SMNH-Type-3289, syntype, Svalbard, West Spitsbergen.
‘Ophiocoma’ arctica Müller & Troschel, 1842: SMNH-Type-5144, syntype, Svalbard, West Spitsbergen.
Ophiacantha fraterna Verrill, 1885: USNM 7905, specimen identified by A.E. Verrill, Steamer Albatross, cruise of 1884, St. 2206, New Jersey, Hudson Canyon, 39°35′00′′N, 71°24′30′′W, depth 1907 m.
USNM 11451, three specimens identified by A.E. Verrill, Steamer Albatross, cruise of 1885, St. 2532, Massachusetts, SE of Georges Bank, 40°34′30′′N, 66°48′00′′W, depth 1289 m.
USNM 33816, lectotype, here designated, one specimen identified by R. Koehler, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
USNM 1100626, paralectotypes, seven specimens identified by R. Koehler, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
Re-identified specimens of O. fraterna removed from O. aculeata USNM lots: USNM 24783, one specimen, re-identified by A.V. Martynov, Steamer Albatross, cruise of 1883, St. 2042, Massachusetts, SE of Nantucket Shoals, 39°33′00′′N, 68°26′45 W, depth 2844 m.
USNM 1100630, one specimen removed from USNM 6610 O. aculeata lot, re-identified by A.V. Martynov, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
YPM 7679, two specimens labelled as ‘Ophiacantha aculeata, Type’, actually belong to Ophiacantha fraterna, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m.
Distribution
Reliable records from south-east of New England (Verrill Citation1885b), from the Reykjanes Ridge, the Charlie-Gibbs Fracture Zone and the Mid-Atlantic Ridge south of the Charlie-Gibbs Fracture Zone (present study) at 1237–2440 m, from northern Rockall Trough (Tyler & Gage Citation1982). Specimens from BIOGAS, at least those occurring below 1500 m, identified as O. bidentata by Paterson (Citation1985), also correspond to O. fraterna.
Remarks
Since 1885 it has been known that there is a deep-water Atlantic species similar to the shallow-water Ophiacantha bidentata. Verrill (1885b, 1899) was the first to separate it as Ophiacantha fraterna. This decision was supported only by Koehler (Citation1914). Most authors maintained O. fraterna just as a variety of O. bidentata (Farran Citation1913; Grieg Citation1921; Mortensen Citation1933). Dyakonov (Citation1954) considered deep-water specimens of O. bidentata under the same name as shallow-water specimens, but did not mention in the synonymy the name fraterna even as a variety. Baranova & Kunzevich (Citation1969) indicated O. fraterna as a synonym of O. bidentata. In the study of reproductive biology of deep-water Ophiacantha bidentata, Tyler & Gage (Citation1982) demonstrated the hermaphroditic nature of deep-water specimens, whereas the shallow-water O. bidentata was known as gonochoristic (Kaufman Citation1974; Tyler & Gage Citation1982). Tyler & Gage (Citation1982) and Gage et al. (1983) concluded that the deep-water populations might represent a distinct species. However, Paterson (Citation1985) did not find any reasons to maintain a separate deep-water species and also included O. fraterna into the synonymy of O. bidentata.
The present study confirms the view of Verrill (1885b) and Koehler (Citation1914) that the deep-water Atlantic species O. fraterna differs from O. bidentata. Ophiacantha fraterna is well distinguished from O. bidentata by the following characters: (1) spicules of the disk of O. fraterna have a rosette of denticles at the top whereas in O. bidentata they are more club-shaped and acute at the top with denticles mainly along the sides of the spicules (compare F and 8G); (2) the outer oral papilla in O. fraterna is always narrow, whereas O. bidentata typically has a widened outer oral papilla (compare H and 8I); (3) the tentacle scale of O. fraterna is more or less narrow, acute, not widened, whereas O. bidentata has a widened tentacle scale (compare A, H and 8E, I); (4) Ophiacantha fraterna is hermaphroditic, whereas O. bidentata is gonochoristic. In addition, O. fraterna is smaller than O. bidentata.
All these characters are enough to keep the species separate. Since Retzius’ types of O. bidentata in Lund University are not traceable (Lars R. Lundqvist, personal communication), for comparative purposes in the present study we examined syntypes of the two species that had been regarded as junior synonyms of O. bidentata. These are Ophiacantha spinulosa Müller & Troschel, 1842 (SMNH-Type-3289) (C) and ‘Ophiocoma’ arctica Müller & Troschel, 1842 (D) (SMNH-Type-5144) as well as O. bidentata from distant regions, i.e. from Norway, the Barents Sea, the Sea of Okhotsk and the Sea of Japan. All these specimens demonstrated clear differences from deep-water Atlantic O. fraterna.
Ophiacantha fraterna and O. aculeata – two common North Atlantic species – have often been found together in considerable quantity at the same stations. Both species were described by Addison Emery Verrill in two successive works (Verrill 1885a,b). While for Ophiacantha fraterna already in the first description (Verrill Citation1885b), a diagnosis sufficient for the separation from other species even without illustrations was provided, the first description of O. aculeata (Verrill, Citation1885a) is very short and ambiguous. However, already in the same year as the first description, Verrill (Citation1885b) published an extended description of O. aculeata, where diagnostic features of this species were expressed clearly. According to Verrill (1885b), O. fraterna is characterized by very small, rough disk spinelets terminated by several minute sharp thorns, and about three rather long, acute and spiniform oral papillae, the distalmost of which does not differ from the others. These features agree well with the specimens used in the present study. Verrill did not give the exact station where O. fraterna was discovered, but it is known that for both of Verrill's works of 1885 material collected during the cruise of the RV Albatross in 1883 was used (Verrill 1885a,b). In addition, records of the depth accompany the first description of O. fraterna – from 908 to 1608 fathoms. Thus we can estimate the approximate number of stations that can be included into the type series. Specimens of O. fraterna from the Yale Peabody Museum (e.g. YPM 7241) originated from the cruise of the Albatross of 1885 and therefore cannot be syntypes. At the same time, USNM keeps some specimens of O. fraterna which, although not labelled as syntypes, were collected during the cruise of the Albatross of 1883, at St. 2105, depth 2551 m (B). These specimens were identified by Rene Koehler and used by him for the re-description of O. fraterna (Koehler, Citation1914). In the latter work Koehler mentioned that these specimens were compared with specimens identified by Verrill from Albatross St. 2573 in 1885. Data of the label of the eight ethanol specimens from the lot USNM 33816 (1883, St. 2105) agree well with those mentioned in Verrill's first description. All eight specimens display essentially the same structure of the disk spinelets and oral papillae and agree well with the first description of O. fraterna. It is necessary to highlight that the type series of both O. fraterna and O. aculeata were never located. From all the available materials from Yale Peabody Museum and the Smithsonian Institution, only the above-mentioned specimens from Albatross cruise 1883 lot USNM 33816 may represent syntypes of O. fraterna. All other specimens, even those identified personally by Verrill, originated from cruises of the Albatross in other years. In summary, despite some potential problems with the decision, we prefer to consider lot USNM 33816 as syntypes of O. fraterna.
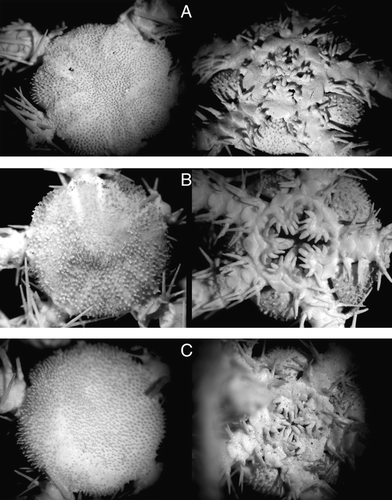
Contrary to O. fraterna, the situation with O. aculeata is more complex. In the first description Verrill (1885a) mentioned very slender rough disk spinelets and four to five rather slender acute oral papillae. As far as very slender disk spinelets clearly point to O. aculeata, the ‘rather slender, acute’ oral papillae may apply equally to both O. aculeata and O. fraterna. However, in the extended description (Verrill Citation1885b) of O. aculeata, Verrill unambiguously mentioned the long slender disk spinelets (including specially highlighting that the spinelets are longer and more slender than in Ophiacantha bidentata) together with broad and flat distalmost oral papillae. Since these very broad distalmost oral papillae have an acute tip, it probably explains why he mentions the ‘all acute oral papillae’ in the first description. Thus, both the first and following descriptions of O. aculeata agree in the presence of ‘long slender disk spinelets’, the features characteristic exactly for O. aculeata and not for O. fraterna (). Koehler (Citation1914) performed a detailed study of both Ophiacantha aculeata and O. fraterna including descriptions of specimens either identified by Verrill or brought to Koehler from the same stations as those of the first description. Koehler's re-description of O. aculeata was based on non-identified specimens from St. 2105, Albatross cruise of 1883, and on specimens identified by Verrill from St. 2725, Albatross cruise of 1886. Current usage of the name O. aculeata (e.g. Paterson Citation1985) is thus based on Verrill's (1885b) extended description without illustrations and the re-description by Koehler (Citation1914) including illustrations of the above-mentioned specimens. We have studied all available specimens of O. aculeata from various Albatross cruises. However, after examination of two specimens (YPM 7679) labelled as ‘Ophiacantha aculeata, Type’ (A) it was discovered that they do not belong to the species O. aculeata. Short disk spinelets and clearly slender oral papillae, all similar in width, including the distalmost papilla, of both YPM 7679 specimens unambiguously point to O. fraterna. Further study of old specimens of O. aculeata from USNM also discovered a similar contradiction of the current usage of the name ‘O. aculeata’ including usage of this name in Verrill's descriptions and own identifications of Verrill. For instance, a specimen from St. 2042, Albatross cruise 1883 (USNM 24783) undoubtedly belongs to O. fraterna, but was identified by Verrill as O. aculeata (C). At the same time, a specimen from St. 2725, Albatross cruise 1886, identified by Verrill as O. aculeata (A), actually belongs to this species. Since the two specimens labelled as ‘O. aculeata, Type’ (YPM 7679) and actually belonging to the species O. fraterna originated from the same station (Albatross, St. 2105, 1883) as recorded for O. aculeata in the first description (Verrill 1885a), further usage of these exemplars as types will lead to confusion between two common North Atlantic deep-water species of Ophiacantha. As was already mentioned above, Verrill (1885a,b) in two works published in the same year unambiguously differentiated the species, O. aculeata and O. fraterna. Thus we consider the discovered confusion with identifications of O. aculeata by Verrill as secondary. It may be explained by further incorrect determination of various specimens, some of which then were designated as a ‘type’. To preserve the current usage of the species names O. aculeata and O. fraterna, which are based on correct interpretations of Verrill's first descriptions (1885a,b), Verrill's extended description of O. aculeata (1885b) and re-descriptions of both species by Koehler (Citation1914), and to avoid further confusion, we here designate lectotypes for both species. For O. fraterna the designated lectotype is selected from syntypes of the lot USNM 33816 (B), for O. aculeata the designated lectotype is selected from syntypes of the lot USNM 12669 (B). Paralectotypes of O. fraterna are seven specimens, USNM 1100626, paralectotypes of O. aculeata are three specimens, USNM 1100627 correspondingly.
Ophiacantha simulans Koehler, 1896
(B)
Figure 10. Ophiacantha aculeata Verrill, 1885. (A) USNM 27900, specimen identified by A.E. Verrill, Steamer Albatross, cruise of 1886, St. 2725, Virginia Beach, 36°34′00′′N, 73°48′00′′W, depth 2513 m, disk diameter 13 mm; (B) lectotype USNM 12669, specimen identified by R. Koehler, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m, disk diameter 12 mm; (C) paralectotype USNM 1100627, same data as in the lectotype, disk diameter ca. 14 mm. Dorsal and ventral views are shown. Photographs: Cynthia Ahearn.
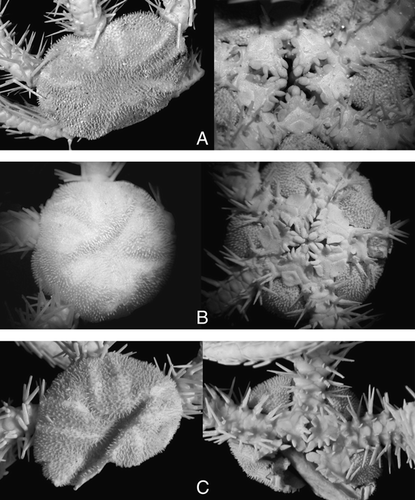
Figure 11. (A) Ophiacantha fraterna Verrill, 1885, specimen labelled as ‘Ophiacantha aculeata, Type’, YPM 7679, Steamer Albatross, cruise of 1883, St. 2105, off Delaware Bay, 37°50′00′′N, 73°03′50′′W, depth 2551 m, disk diameter 6 mm; (B) Ophiacantha simulans Koehler, 1896, ZMMU D-640, RV Akademik Mstislav Keldysh, St. 464, disk diameter 6 mm; (C) Ophiacantha spectabilis G.O. Sars, Citation1871, ZMMU D-660, RV Akademik Mstislav Keldysh, St. 464, disk diameter ca. 11 mm; (D) Ophiacantha veterna Koehler, 1907, ZMMU D-657, RV Akademik Mstislav Keldysh, St. 464, disk diameter 7.5 mm. Dorsal and ventral views are shown. Photographs: Tanya Korshunova.
Ophiacantha simulans: Koehler Citation1896a: 82, Plate 4, Figures 37, 38; Paterson Citation1985: 39–40, .
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 415, one specimen; St. 464, two specimens.
Distribution
The North Atlantic, in the west – off Carolinas, in the east – from Iceland to the Azores, at 1575–3018 m (Paterson Citation1985) and of Morocco, at the depth 1750 m (Bartsch Citation1987). Present study: the Reykjanes Ridge, at 1535–1785 m.
Ophiacantha spectabilis G.O. Sars, Citation1871
(C)
Ophiacantha spectabilis: G.O. Sars 1871: 10; Paterson Citation1985: 41, .
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 464, one specimen.
Distribution
The North Atlantic: off Nova Scotia, south of Iceland, Norway, south-west Ireland, Bay of Biscay, at depths 145–1700 m (Paterson Citation1985). Present study: Reykjanes Ridge, at 1670–1750 m.
Ophiacantha veterna Koehler, 1907
(D)
Ophiacantha veterna: Koehler Citation1907b: 41.
Ophiacantha enopla veterna: Paterson Citation1985: 37, .
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 464, three specimens.
Distribution
Prior to this study O. veterna was recorded only from the Bay of Biscay and south of the Azores and Madeira at 101–2300 m (Koehler Citation1907b; Paterson Citation1985; Stöhr & Segonzac Citation2005). Now the range is extended to the north to the Reykjanes Ridge, at 1670–1750 m.
Remarks
Paterson (Citation1985) considered O. veterna as a subspecies of O. enopla Verrill, 1985. We believe that characters distinguishing O. enopla enopla from O. enopla veterna listed by Paterson are enough to maintain them as separate species. Especially characteristic is the shape of tentacle scales in O. veterna: these are very small compared to those of O. enopla and are missing on distal arm segments.
Ophiocamax patersoni sp. nov.
(Figures 13D, 14, 15C, F, 16H, I)
Material
RV G.O. Sars, MAR-ECO cruise, St. 52/374, holotype (ZMBN 77854, dried), one paratype (ZMBN 77855, dried), St. 50/373, 10 paratypes (ZMBN 77856, in ethanol), two paratypes (ZMMU D-748, dried), St. 72/386, one paratype (ZMBN 77857, in ethanol).
Etymology
This species is named in honour of Gordon L.J. Paterson (The Natural History Museum, London) for his important contribution in studying North Atlantic Ophiuroidea.
Description of the holotype
The disk has diameter 17.5 mm, is domed and circular, but indented interradially to various degrees – from minor to considerable. The disk plates are numerous, different in size and shape, from small to moderate, irregular-rhomboidal and -polygonal, bearing one or few long spines. Some spines bearing at about mid-length a few irregularly placed short thorns, whereas others lack distinct thorns. Proximal to the radial shields the disk plates are larger and fewer, whereas in the centre of the disk and in the interradii the plates are smaller. Radial shields (2–3 mm in length) are relatively small, about one-sixth of the disk diameter, irregularly triangular in outline, proximally narrowed and obtuse pointed, distally widened, joined at almost their length, but in two interradii have irregular row of spinelets in-between. At the distal border of the radial shields and between the first dorsal arm plate there is a dense irregular row of spines. The interradii are swollen, ventrally covered by spines; close to the oral shields the interradial scales bear short tubercle-like spines or are naked. Each jaw bears two or three irregularly placed narrow spiniform apical papillae. The latter are hardly distinguished from teeth and from adjacent lateral oral papillae. Nevertheless, the first pair of the lateral papillae is quite distinct. Other papillae are placed along jaws in two irregular rows – one internal and one external. At least two pairs placed dorsally in the middle of the jaws. The distal papillae are similar to the proximal ones in shape, differing only in length, but forming a semicircular comb around the second tentacle pore. The first papillae are pointed, narrow. Sometimes additional papillae are placed between apical and first papillae. The shape of the oral shield can be described as wide-arrowhead with straight rather long distal part attached to the interradius and triangular proximal part, completely separated from the first lateral arm plate by the wide adoral shields. The madreporic oral shield is swollen and has lost its usual shape. One to five rough spindle-shaped papillae (in one interradius these papillae probably are lost) are placed on the proximal edge of the madreporite. Two oral shields also have two tubercles at their distal edge. Arm length is about four times the disk diameter. The dorsal arm plates are broad, fan-shaped, rather large, entirely separated by the lateral arm plates, even on most basal segments. The proximal edge of the dorsal arm plate is almost straight throughout the length of the arm, whereas on the basal segments this edge is slightly triangular. The distal edge of the dorsal arm plate is semicircular and distinctly covered with sparse quite long thorns. These thorns are present already on most basal segments and placed in few irregular rows – one more close to the distal edge, whereas others are placed farther from the edge of the dorsal arm plate. The proximal area of the dorsal arm plate is also covered sparsely with short thorns or spiny small tubercles. On the basal segments these proximal thorns are more developed, whereas towards the distal end of the arm they tend to be reduced. The middle area of the dorsal arm plates has few and weakly developed thorns or is fully devoid of them. Most distal dorsal arm plates have ca. five blunt tubercle-shaped distal thorns and few prominent needle-shaped thorns on the middle line of the lateral plates. Arms have distinct nodes and form a strong lateral ridge on which the spine arm articulation ridges are placed. On most proximal segments there are eight or nine spines, in the middle of the arm – six or seven, distally – five or six. The dorsalmost spine is shortest, the second slightly longer and the third is longest and strongest (about four segments in length). The next spine is considerably shorter and following spines are gradually reduced in size. Sparse quite long thorns cover the dorsal spines including the longest, whereas the more ventrally placed spines have a rough surface with some short denticles. The first ventral arm plate is rather small, trapezoid. The second ventral arm plate is very remarkable – distally rhomboid, proximally with two semicircular lateral wings corresponding to the large basal tentacle pore. The third and fourth ventral arm plates are also distally rhomboid (about two times wider than long) and proximally show traces of the above described structure, which disappears towards the middle of the arm and again becomes more developed on more distal segments. Further ventral arm plates towards the middle of the arm have a triangular proximal lobe, while they are polygonal with rounded angles distally. Some segments have nearly ovoid ventral arm plates. The distalmost segments have a rather trapezoid distal part of the ventral arm plate. Ventral plates throughout the length of the arm are separated. The tentacle pores are larger and more conspicuous proximally than towards the distal end of the arm, where they become small and inconspicuous. Basally around the tentacle pores at least four tentacle scales are placed. The basal tentacle scales are large, narrowly conical, with widened base and gradually sloped towards top. Towards the distal end of the arm the number of tentacle scales reduced to two and then to one. About at mid-length of the arm the tentacle scales became spindle-shaped, and at the distalmost segments they turn into a rather short tubercle.
Paratype variations
The smallest paratype with disk diameter 8.5 mm (radial shields approximately 0.8–0.9 mm) is different compared to the large specimen disk spine structure. These spines, both in the centre and periphery have a well-developed basal rosette of four or five long thorns and several also have long denticles towards the top of the spine. At the distal border of the radial shields similar spines are placed, with a basal rosette of denticles. The radial shields are very short and irregularly square in outline, entirely joined in some radii, whereas in others slightly divergent proximally. The dorsal arm plates, especially on the seven or eight proximal segments at their distal edge heavily covered by long spines (15–20 spines on the mentioned segments), which are arranged in two irregular rows. Towards the middle of the arm the spines become more tubercle-shaped and reduced in number. The oral papillae are more irregular than in the holotype, the single large, and small additional rows of papillae encircling the second tentacle pores are especially well developed. The additional papillae are placed on the adoral shields instead of on the oral shield, and the shape of the papillae are approaching the shape of the disk spines but more massive and with lesser developed rosette of denticles. The oral shields are more or less hourglass-shaped. The tentacle scales on all segments, including the proximalmost, are spiniform and not massive conical. At least one tentacle scale has a wide base contrasting with its narrow remaining part. The tentacle scales are rough, some have distinct thorns. Proximally there are seven to nine spines, five or six towards the distal end of the arm. The largest paratype of 19.5 mm disk diameter (radial shields 3–4 mm in length) has disk spines that are roughly spiniform but without thorns. Only a few spines in the interradii have two or three blunt thorns. The radial shields are triangular, some with obtuse proximal angle, whereas others have an acute angle. The radial shields are in contact for most of their length, but in two radii there is a distinct narrow insertion plate in-between. Proximally the radial shields are divergent. All dorsal arm plates bear on their distal edge a set of, to a various degree developed, blunt short spines (on proximal segments) or tubercles, 15–20 in number on proximal segments. Few short sharp spines or tubercles are placed also proximally on the dorsal arm plate. The dorsal arm plates are rhomboidal, wider than long. Towards the middle of the arm the dorsal plates became wider and shorter. All dorsal arm plates are not contiguous but the basalmost plates lie very close to each other due to somewhat dorsally shifted arms.
The oral papillae have a more irregular appearance than in the holotype, but they also have two distinct irregular rows around the tentacle pore (laterally they are adjoining true tentacle scales). The tentacle scales at the second and third segments are very long, narrow, up to five in number, with a smooth surface. The oral shield is shaped with a proximal wide triangular part with slightly convex sides, and a triangular lobe of considerable size. At the lateral sides of the distal edge of the triangular lobe there are usually one or two pairs of long narrow papillae. The distal part of the oral shield is short and wide.
Distribution
The Charlie-Gibbs Fracture Zone, the Mid-Atlantic Ridge north of the Azores, 2522–2979 m.
Remarks
For comparative purposes we studied type material of various species of the genus Ophiocamax as well as non-type specimens of O. dominans Koehler, Citation1906 identified by Ilse Bartsch. Since species of the genus Ophiocamax are difficult to distinguish, we illustrate here the above-mentioned material. From the geographically closest species O. dominans Koehler, Citation1906 (known presently only off Sahara, and never reported from the Reykjanes Ridge), Ophiocamax patersoni sp. nov. clearly differs in the following characters. Disk plates are more numerous, radial shields are relatively small (from one-fifth to one-tenth of the disk diameter in Ophiocamax patersoni sp. nov. and from one-half to three-quarters in O. dominans; the latter according to Paterson (Citation1985) (compare Figures 12A, 15A and 13D, 14A, B, 15C, F). In the specimens identified by Bartsch we found the length of the radial shield to the disk diameter to be one-third to one-fifth, thus not considerably different from Ophiocamax patersoni sp. nov. Disk spines of Ophiocamax patersoni sp. nov are narrow elongated cones, almost devoid of the long thorns in large specimens (few can be found in the interradial area), whereas in large specimens of O. dominans most disk spines presented massive spines with conspicuous thorns. The distal edges of the radial shields of Ophiocamax patersoni sp. nov. are heavily covered with long spines (C), whereas O. dominans has radial shields almost devoid of spines at their distal edge (A). The dorsal arm plates of Ophiocamax patersoni sp. nov. are rhomboidal, slightly convex at their proximal edge, and form an obtuse angle at the distal edge, wider than long, not contiguous even on most basal segments (Figures 13D, 14A–C, 15C, F) (O. dominans has dorsal arm plates clearly triangular, distal edge convex, proximal edge represented by an acute angle, clearly contiguous at the basal segments and still nearly in contact (Figures 12A, 15A).) In Ophiocamax patersoni sp. nov. most dorsal arm plates have distal edges heavily covered with relatively long spines or obtuse tubercles (Figures 14C–E, 15C, F), whereas in O. dominans they are entirely devoid of spines (A), except for the distal part of the arms where two tiny spines appear (according to Bartsch Citation1987). The shape of the oral shield of Ophiocamax patersoni sp. nov. is an arrow-head (proximal part is large, acutely triangular with slightly concave lateral sides) (H, I), whereas O. dominans has the oral shield irregularly hour-glass-shaped (A). Another Atlantic species, O. fasciculata Lyman, 1883 from the Caribbean region, also differs considerably from Ophiocamax patersoni sp. nov. by a number of features: dorsal arm plates clearly triangular, distal edge convex, proximal edge represented by an acute angle, clearly contiguous on the basal segments and still nearly in contact, most dorsal arm plates entirely devoid of spines (Figures 12B, 15B), oral shields with large ovoid proximal part with small triangular lobe and short wide distal edge (B), apical and oral papillae are very irregular, tentacle scales with considerably widened base (Figures 12B, 16B). The only other Caribbean species, O. hystrix Lyman, 1878 (and very similar to this species Ophiocamax austera Verrill, Citation1899), has slightly rugose dorsal and lateral arm plates (D), but in other characters it differs greatly from Ophiocamax patersoni sp. nov.: distinctly lobed disk, deeply notched interradially thus forming a very small central area (C), disk covered with short massive conical stumps or blunt tubercles, trapezoid dorsal arm plates contiguous basally (D), oral papillae placed proximally in two distinct clusters (C), ovoid proximal part of the oral shield with small triangular proximal lobe and distinct swollen tubercle on distal edge (C). The North Atlantic Ophiocamax patersoni sp. nov. is by general appearance and shape and structure of the dorsal arm plates even more similar to Antarctic species of the genus – O. gigas Koehler, 1901 and especially to O. drygalskii Hertz, 1927. Ophiocamax gigas differs from Ophiocamax patersoni sp. nov. in the numerous short spinelets of the disk having few thorns at the top (A), dorsal arm plates with convex proximal edge and distal edge forming an obtuse angle covered by a single regular row of rounded tubercles (E), the oral shield heavily covered with thorny spines (E) and tentacle scales with few denticles at the top. Ophiocamax drygalskii Hertz, 1927 differs from Ophiocamax patersoni sp. nov. in the numerous short spinelets of the disk having few denticles at the top or along their sides (even in large specimens) (B, C), proximal edge of the dorsal arm plate distinctly triangular – from acute to obtuse angle, nearly contiguous on basal segments (Figures 13B, C, 15G, H), oral shield hour-glass-shaped or with very elongated proximal part (F, G). It is necessary to note that in the studied type specimens of O. drygalskii the shape of the oral shield is quite variable. Different shapes of the oral shields were observed even within single specimen (madreporic oral shield is not considered). At the same time, Ophiocamax patersoni sp. nov. does not display such polymorphism and all studied large specimens have essentially the same shape of the oral shield, well distinguished from O. drygalskii by this character. Ophiocamax applicatus Koehler, Citation1922 described off Tasmania, Australia is considerably larger than Ophiocamax patersoni sp. nov. and has proximally contiguous triangular dorsal arm plates. Ophiocamax brevicetra Baker, 1974, off Otago, New Zealand, differs from Ophiocamax patersoni sp. nov. in shape of disk spinelets, structure of spines, shape of thorns at distal edge of the dorsal arm plates and in shape of the oral shield.
Finally, the common Indo-West Pacific deep-water species O. vitrea, which was recently shown as being very polymorphic (O'Hara & Stöhr Citation2006), is well distinguished from Ophiocamax patersoni sp. nov. by the shape and structure of the disk spines and spines of the arm (Figures 12D, 15I), numerous irregular oral papillae and the shape of the oral shield (D).
Thus, in the North and Middle Atlantic there are several locally distributed, but well-distinguished species – in the Caribbean region O. fasciculata and O. hystrix; off Northern Africa O. dominans; and on the Charlie-Gibbs Fracture Zone and the Mid-Atlantic Ridge north of the Azores Ophiocamax patersoni sp. nov.
Ophiolimna bairdi (Lyman, 1883)
(A)
Figure 12. (A) Ophiocamax dominans Koehler, Citation1906, ZSM 20043260, RV Meteor, Cruise 36, St. 98/ES 148, off Spanish Sahara, 25°40.8′N, 16°02.0′W, depth 883–992 m, disk diameter 11.1 mm; (B) Ophiocamax fasciculata Lyman, 1883, holotype MCZ 2082, Blake, St. 209, off Martinique, depth ca. 344 m, disk diameter 14 mm; (C) Ophiocamax hystrix Lyman, 1878, holotype MCZ 2084, Blake, off Havana, Cuba, depth ca. 318 m, disk diameter 13 mm; (D) Ophiocamax vitrea Lyman, 1878, syntype MCZ 2096, Challenger, St. 219, off Admiralty Is., depth ca. 273 m, disk diameter 17 mm. Dorsal and ventral views are shown. Photographs: Tanya Korshunova (A); Alexander Martynov (B–D).
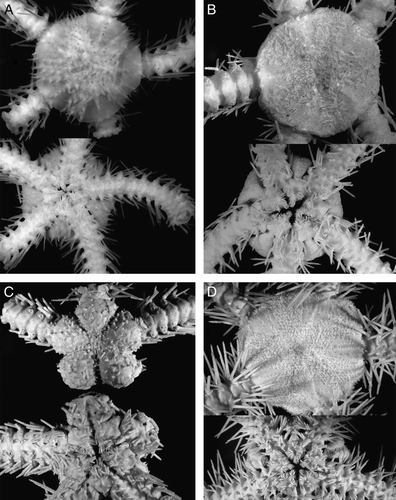
Figure 13. (A) Ophiocamax gigas Koehler, 1901, IORAS 5.1, RV Akademik Kurchatov, Cruise 11, St. 881, 57°8′S, 26°42′W, depth 932 m, disk diameter 34 mm; (B) Ophiocamax drygalskii Hertz, 1927, syntype ZSM 20001015/2, Deutsche Südpolar-Expedition, 65°31′S, 85°14′W, depth 2450 m, disk diameter 19.8 mm; (C) Ophiocamax drygalskii Hertz, 1927, syntype ZSM 20001015/1, 65°31′S, 85°14′W, depth 2450 m, disk diameter 20.1 mm; (D) Ophiocamax patersoni sp. nov., holotype ZMBN 77854, MAR-ECO St. 52/374, disk diameter 17.5 mm. Dorsal and ventral views are shown. Photographs: Alexander Martynov (A, D); Tanya Korshunova (B, C).

Figure 14. Ophiocamax patersoni sp. nov. (A) Paratype ZMMU D-748, MAR-ECO St. 50/373, disk diameter 19.5 mm, dorsal and ventral views; (B) paratype ZMMU D-748, MAR-ECO St. 50/373, disk diameter 8.5 mm, dorsal and ventral views; (C) holotype ZMBN 77854, scanning electron micrograph of dorsal arm plate, basal segments; (D) holotype ZMBN 77854, scanning electron micrograph of dorsal arm plate, distal segments; (E) paratype ZMMU D-748, disk diameter 8.5 mm, scanning electron micrograph of dorsal arm plate, basal segments (enlarged view of spines also shown); (F) paratype ZMMU D-748, disk diameter 8.5 mm, scanning electron micrograph of arm spine articulation ridges, middle segments; (G) paratype ZMMU D-748, disk diameter 19.5 mm, scanning electron micrograph of tentacle scale, basal segments; (H) paratype ZMMU D-748, disk diameter 19.5 mm, scanning electron micrograph of ventral arm spine, middle segments; (I) paratype ZMMU D-748, disk diameter 19.5 mm, scanning electron micrograph of basal part of third arm spine, middle segments. Scale bars: 300 µm (C–E, G–I).; 100 µm (F). Photographs: Alexander Martynov.
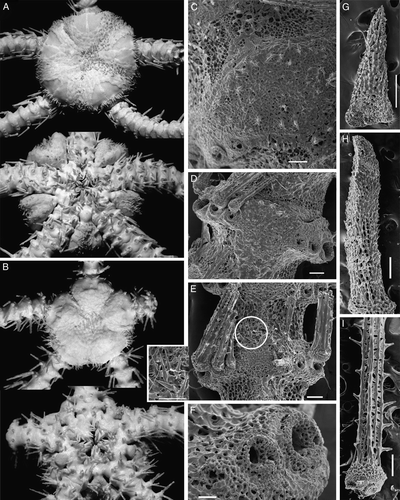
Figure 15. Segment of the disk and basal part of the arm of various Ophiocamax species. (A) Ophiocamax dominans Koehler, Citation1906, ZSM 20043261, RV Meteor, Cruise 36, St. 98/AT 149, off Spanish Sahara, 25°31′05′′N, 16°02′00′′W, depth 658–888 m, disk diameter 16.5 mm; (B) Ophiocamax fasciculata Lyman, 1883, paratype MCZ 4613, Blake, St. 145, 147, 148, off St Kitts, 378–491 m, disk diameter 18 mm; (C) Ophiocamax patersoni sp. nov., paratype ZMMU D-748, MAR-ECO St. 50/373, disk diameter 19.5 mm; (D) Ophiocamax hystrix Lyman, 1878, holotype MCZ 2084, Blake, off Havana, Cuba, depth ca. 318 m, disk diameter 13 mm; (E) Ophiocamax gigas Koehler, 1901, RV Akademik Kurchatov, Cruise 11, St. 881, 57°8′S, 26°42′W, depth 932 m, disk diameter 34 mm; (F) Ophiocamax patersoni sp. nov., paratype ZMMU D-748, MAR-ECO St. 50/373, disk diameter 8.5 mm; (G) Ophiocamax drygalskii Hertz, 1927, syntype ZSM 20001015/1, 65°31′S, 85°14′W, depth 2450 m, disk diameter 18.1 mm; (H) Ophiocamax drygalskii Hertz, 1927, syntype ZSM 20001015/2, Deutsche Südpolar-Expedition, 65°31′S, 85°14′W, depth 2450 m, disk diameter 20.9 mm; (I) Ophiocamax vitrea Lyman, 1878, syntype MCZ 2096, Challenger, St. 219, off Admiralty Is., depth ca. 273 m, disk diameter 17 mm. Photographs: Tanya Korshunova (A, G, H); Alexander Martynov (B–F, I).
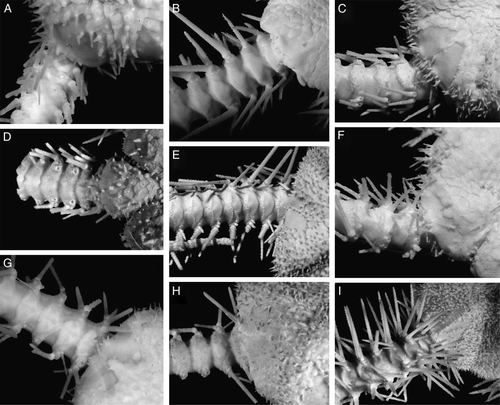
Figure 16. Oral frame showing shape of the oral shield of various Ophiocamax species. (A) Ophiocamax dominans Koehler, Citation1906, ZSM 20043261, RV Meteor, Cruise 36, St. 98/AT 149, off Spanish Sahara, 25°31.5′N, 16°02.0′W, depth 658–888 m. (B) Ophiocamax fasciculata Lyman, 1883, paratype MCZ 2083, Blake, St. 295, off Barbados, ca. 328 m. (C) Ophiocamax hystrix Lyman, 1878, holotype MCZ 2084, Blake, off Havana, Cuba, depth ca. 318 m. (D) Ophiocamax vitrea Lyman, 1878, syntype MCZ 2096, Challenger, St. 219, off Admiralty Is., depth ca. 273 m. (E) Ophiocamax gigas Koehler, 1901, RV Akademik Kurchatov, Cruise 11, St. 881, 57°08′S, 26°42′W, depth 932 m. (F, G) Ophiocamax drygalskii Hertz, 1927, syntype ZSM 20001015/2, Deutsche Südpolar-Expedition, 65°31′S, 85°14′W, depth 2450 m. (H, I) Ophiocamax patersoni sp. nov.: (H) holotype ZMBN 77854, MAR-ECO St. 52/374; (I) paratype ZMMU D-748, MAR-ECO St. 50/373, disk diameter 19.5 mm. Scale bars: 1 mm. Photographs: Tanya Korshunova (A, F, G); Alexander Martynov (B–E, H, I).
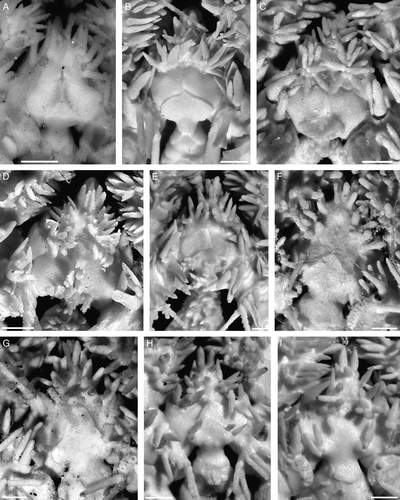
Ophiacantha bairdi: Lyman 1883: 256, Plate 5, Figures 70–72.
Ophiolimna bairdi: Verrill 1899: 40, 44; Paterson Citation1985: 60, Figure 24.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 316-5, 56 specimens; St. 415, one specimen; St. 445, three specimens; St. 478, one specimen; St. 499, 10 specimens.
Distribution
Arctic Ocean; North Atlantic: in the west – south of Martha's Vineyard, east – from the Rockall Trough to south of Portugal, at 620–2600 (Dyakonov Citation1954; Paterson Citation1985). Present study: the Reykjanes Ridge, at 1535–2440 m.
Figure 17. (A) Ophiolimna bairdi (Lyman, 1883), ZMMU D-648, RV Akademik Mstislav Keldysh, St. 499, disk diameter 11 mm. (B) Histampica duplicata (Lyman, Citation1875), ZMMU D-729, RV Akademik Mstislav Keldysh, St. 445, disk diameter 5.5 mm. (?) Ophiactis abyssicola (M. Sars, Citation1861), ZMMU D-634, RV Akademik Mstislav Keldysh, St. 478, five-armed specimen, disk diameter 6.5 mm. (D) Ophiactis abyssicola (M. Sars, Citation1861), MAR-ECO, St. 60/379, six-armed specimen, disk diameter 5 mm. (E) Ophiactis abyssicola (M. Sars, Citation1861), MAR-ECO, St. 60/379, seven-armed specimen, disk diameter 4.8 mm. (F, G) Ophiothamnus chariis H.L. Clark, Citation1915, MAR-ECO, St. 60/379: (F) scanning electron micrograph of the ventral side; (G) scanning electron micrograph of the spicule of the disk. (H) Ophiochiton ternispinus Lyman, 1883, ZMMU D-743, RV Akademik Mstislav Keldysh, St. 445, disk diameter 5.5 mm. Dorsal and ventral views are shown in (A–E, H). Scale bars: 600 µm (F); 60 µm (G). Photographs: Tanya Korshunova (A, B, H); Konstantin Tabachnik (C–E); Alexander Martynov (F, G).
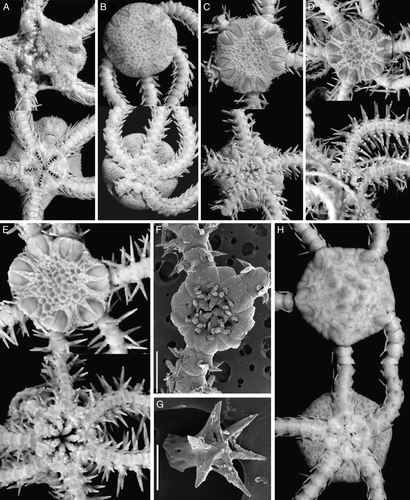
Family Ophiactidae
Histampica duplicata (Lyman, Citation1875)
(B)
Amphiura duplicata: Lyman 1875: 19–20, Text figure 87, Plate 5, Figure 78.
Ophiactis duplicata: Lütken & Mortensen 1899: 142–3, Plate 6, Figures 1–3.
Amphiactis duplicata: Matsumoto Citation1915: 66–7.
Histampica duplicata: A.M. Clark Citation1970: 73–4.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 418, two specimens.
Distribution
The Caribbean, the Pacific Ocean. North Atlantic records: Bay of Biscay to north Africa, at 125–2870 m (Paterson Citation1985). Present study: the Reykjanes Ridge, at 1815–1915 m.
Ophiactis abyssicola (M. Sars, Citation1861)
(C–E)
Amphiura abyssicola: M. Sars 1861: 18.
Ophiactis abyssicola: Ljungman Citation1867: 324; Paterson Citation1985: 76–8, Figure 32.
Ophiactis poa: Lyman 1882: 119.
Ophiactis echinata: Koehler Citation1898: 48.
Ophiactis corallicola: Koehler Citation1896a: 75.
Material
RV G.O. Sars, MAR-ECO cruise, St. 42/368, one specimen; St. 53/375, one specimen; St. 60/379, 25 specimens; St. 62/380, 13 specimens; St. 65/382, five specimens; St. 70/385, seven specimens; RV Akademik Mstislav Keldysh, fourth cruise, St. 319, 13 specimens; St. 364, five specimens; St. 385, 56 specimens; St. 415, three specimens; St. 418, one specimen; St. 445, 40 specimens; St. 446, three specimens; St. 464, one specimen; St. 465, 65 specimens; St. 478, one specimen; St. 489, 15 specimens.
Distribution
North Atlantic: Davis Strait, off Iceland, Faeroe Islands, from Norway to southern Africa, at 121–4721 m (Paterson Citation1985). Present study: the Reykjanes Ridge, at 607–2107 m.
Remarks
Mortensen (Citation1933) recorded six- and seven-armed specimens of this species as a rare abnormality. This feature was confirmed in the present study (C–E). Other characters are essentially the same as in common five-armed specimens. In our material this species was found associated with the sponge Hertwigia falcifera Schmidt, 1880.
Ophiothamnus chariis H.L. Clark, Citation1915
(F, G)
Ophiacantha gracilis Verrill Citation1885b: 548.
Ophiothamnus gracilis: Verrill 1899: 41.
Ophiothamnus chariis H.L. Clark Citation1915: 208; Mortensen Citation1933: 44–5, Figure 27.
Material
RV G.O. Sars, MAR-ECO cruise, St. 60/379, one specimen.
Distribution
Off Nova Scotia, 400–1561 m (Verrill Citation1885b) and Davis Strait, at 700–1055 m (Mortensen Citation1933). Present study: the Reykjanes Ridge, at 1237–1296 m. This species was omitted by Paterson (Citation1985) and recorded here for the first time after 70 years.
Family Ophiochitonidae
Ophiochiton ternispinus Lyman, 1883
(H)
Ophiochiton ternispinus Lyman 1883: 255–6, Plate 5, Figures 67–69; Paterson Citation1985: 96–7, Figure 39.
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 316, one specimen; St. 413, one specimen; St. 445, five specimens; St. 446, one specimen; St. 489, one specimen.
Distribution
North Atlantic: in the west – from the Gulf of Mexico to Davis Strait, in the east – from south-eastern Iceland to south of the Azores, at 425–2220 m. Present study: the Reykjanes Ridge, at 1550–2200 m.
Family Ophiolepididae
Ophiophyllum nesisi sp. nov.
()
Material
RV Akademik Mstislav Keldysh, fourth cruise, St. 464 holotype (ZMMU D-746, dried), St. 492, one paratype (ZMMU D-747, dried).
Etymology
This species is named in honour of the prominent cephalopod taxonomist Kir Nesis, whose early works were devoted to the North Atlantic benthic fauna.
Figure 18. Ophiophyllum nesisi sp. nov. (A) Holotype ZMMU D-746, RV Akademik Mstislav Keldysh, St. 464, disk diameter 7 mm, dorsal and ventral views; (B) segment of the disk and the arm of the holotype; (C) scanning electron micrograph of the lateral arm plate of the holotype; (D) scanning electron micrograph of the arm spine articulation ridges of the holotype; (E) scanning electron micrograph of the ventral spine from distal arm segments of the holotype; (F) scanning electron micrograph of the dorsal spine of the paratype, RV Akademik Mstislav Keldysh, St. 492; (G) scanning electron micrograph of the spine from the disk fringe of the paratype. Scale bars: 200 µm (C); 100 µm (D, F, G); 120 µm (E). Photographs: Alexander Martynov.

Description of the holotype
The disc diameter of the holotype is 7 mm. It is subcircular, moderately swollen and distinctly concave in the interradii. The dorsal disk surface is clearly divided into two areas – a concave, oblique peripheral area including large radial shields and few (including at least one large) inserted plates between it and a slightly depressed central area covered with numerous (about 110 in number) small, almost uniform in size overlapping irregular-rounded plates. The plates are elevated towards forming tubercle-shaped structures. There are no conspicuous primary plates. A single subcentral plate is slightly larger and more elevated than the neighbouring ones. The surface of the disk plates is smooth. The radial shields are large, elongated narrowly and cup-shaped in outline, proximally obtuse, distally slightly widened. The radial shields are entirely separated from each other by one to two narrow slightly carinated plates. In the interradii the radial shields are separated by one large trapezoid plate distally and few irregularly square plates proximally. The surface of the radial shields is smooth. In the interradii at the border of the disk there is a fringe comprised of a single row of flattened spines. In two interradii the spine fringe is almost completely lost. The jaws are dorsally concave, each bearing three short spiniform irregular-appearing apical papillae of different length and six to eight pairs of rectangular lateral oral papillae. The lateral papillae are formed along the entire jaw edges without gap. The outermost papillae are wide, often with a sharpened edge directed towards the mouth slit. The middle papillae are irregularly rectangular, whereas the most proximal papillae are more spiniform or tubercle-shaped. Both outermost and middle papillae at some jaws have a general appearance as a single block-like scalloped structure. There is a single long spiniform tooth on each short dental plate. The oral shield is large, irregularly rhomboidal in shape, wider than long. In one interradius the oral shield is fragmented by two middle insertion plates. Ventrally there are few irregularly polygonal plates proximally in contact with the oral shield and distally with the spine fringe. The adoral shields are long narrow bands thickened where they are in contact, about five times as long as wide. Genital slits are short, extending from second to third ventral arm plates, in some interradii quite distinct, whereas in others tightly closed and inconspicuous. All arms are broken, maximal number of the saved segments is nine. The saved segments have a quite massive general appearance. Together with the adjacent parts of the disk spine fringe the arms appear conical. The dorsal arm plates are small, narrowly hexagonal in shape and distinctly carinated, contiguous throughout the saved segments of the arms. Distal edges of the dorsal plates are concave. The lateral plates are abruptly sloping giving the arms a high and carinated general appearance. Spines are placed only at the ventral border of the lateral plates. There are two spines: a spiniform one is placed more dorsally, while another is wide and flat, and placed almost ventrally at nearly the same level as the ventral plates. On basal segments the ventral spines are wider than on more distal segments. The ventral spine has a concave surface and few ridges. Ventrally the dorsal plates are placed distinctly obliquely in relation to the ventral plates. The first ventral arm plate is rather square with slightly concave lateral sides. Other ventral arm plates have quite straight proximal and distal edges and deeply concave lateral sides. All ventral arm plates are contiguous. The tentacle pores are large, bordered by deep incisions in the ventral arm plates and oblique lateral arm plates. Tentacle scales are entirely reduced, but at a few basal segments there are two to five thickened, rarely even formed very small, short tubercle-like tentacle scales.
Paratype variation
The single paratype is generally similar to the holotype in size and structure, but differs by more separated, not block-shaped middle oral papillae and the presence of several (the holotype has a single) spiniform teeth.
Distribution
Reykjanes Ridge, at 1670–1895 m.
Remarks
A species named after the Atlantic, Ophiophyllum atlanticum Stöhr & Segonzac, Citation2005, was recently described from a depth of 4078 m from the Mid-Atlantic Ridge in the Central Atlantic (Stöhr & Segonzac Citation2005). Before the discovery of that species, only a single record of the genus was known from the Atlantic, the Caribbean O. petilum Lyman, 1878, but that record was dubious, because the species had originally been described from the Pacific Ocean (Clark Citation1915). Despite that Ophiophyllum nesisi sp. nov. like O. atlanticum was also discovered from the Mid-Atlantic Ridge (but considerably more northerly at almost half the depth), it differs clearly from the latter in a number of characters. In Ophiophyllum nesisi sp. nov., basal segments of arms together with adjacent parts of the disk clearly form a cone in outline whereas the corresponding parts of the arms in O. atlanticum have a cylindrical appearance. The dorsal arm plates of O. atlanticum are triangular, with straight distal edge, whereas in Ophiophyllum nesisi sp. nov. they are polygonal, with distinctly concave distal edge. The ventral spine articulation ridges of the lateral arm plates have a distinctly subparallel appearance with two openings in Ophiophyllum nesisi sp. nov., whereas O. atlanticum has a semicircular ridge with hardly conspicuous openings. The dorsal small spine of Ophiophyllum nesisi sp. nov. is almost straight and narrow, whereas O. atlanticum has a distinctly conical dorsal spine with wide base. The oral shield, despite its variability, is large rhomboidal in Ophiophyllum nesisi sp. nov. and rather small and rounded pentagonal in O. atlanticum, and, finally, the appearance of the oral frame is very different in Ophiophyllum nesisi sp. nov. and O. atlanticum – in the latter the middle oral papillae are rounded and there is a distinct gap devoid of papillae between middle and apical papillae, whereas Ophiophyllum nesisi sp. nov. has square or block-shaped middle oral papillae and no gap. Holotype and paratype of Ophiophyllum nesisi sp. nov. are originated from two different stations but show essentially the same features. Ophiophyllum nesisi sp. nov. is more similar to the New Zealand species O. teplium McKnight, 2003 than to its Atlantic congeners. From O. teplium, Ophiophyllum nesisi sp. nov. differs clearly in its elongated cup-shaped and not triangular radial shields, the rhomboidal and not ovoid oral shield, fewer (six to eight in Ophiophyllum nesisi sp. nov. and seven to ten in O. teplium) and often block-shaped oral papillae. Type species of the genus, O. petilum Lyman, 1878 differs from Ophiophyllum nesisi sp. nov. by absence of the small dorsal arm spine, and even if the dorsal spine is considered easily lost, it differs clearly in its flat, thin not domed disk (Lyman 1882: 102 in his description of O. petilum specially highlighted this as ‘disk flat and very thin’, given thus the genus name Ophiophyllum). Genital slits of O. petilum are relatively long and extending from the oral shield to the fringe of disk spines, whereas in Ophiophyllum nesisi sp. nov. they are short and extending from adoral shield to lateral plate of the third segment. Arms of O. petilum have cylindrical appearance (conical in Ophiophyllum nesisi sp. nov.) and considerably fewer segments within the disk (two in O. petilum and four in Ophiophyllum nesisi sp. nov.). Ophiophyllum petilum has short and wide ventral side of the lateral arm plate, whereas in Ophiophyllum nesisi sp. nov. it is narrow, relatively long and oblique. In Ophiophyllum petilum there is a clear gap between spines of the adjacent segments, whereas in Ophiophyllum nesisi sp. nov spines are overlapping. First ventral arm plate is triangular in Ophiophyllum petilum and square with concave lateral sides in Ophiophyllum nesisi sp. nov., same length as width (or wider) of the other ventral arm plates in Ophiophyllum petilum and distinctly longer than broad in Ophiophyllum nesisi sp. nov. Shape of distalmost and other oral papillae is similar in Ophiophyllum petilum, whereas Ophiophyllum nesisi sp. nov. has a markedly different distalmost oral papilla in shape and size. Ophiophyllum concinnus Litvinova, 1981 is considerably different from Ophiophyllum nesisi sp. nov. in having a flattened disk, not clearly divided into central and peripheral areas, a cylindrical appearance of the arms, triangular dorsal arm plates, a distinct gap between spines of the adjacent segments, less differentiated oral papillae and a small rounded oral shield. Ophiophyllum borbonica Vadon & Guille, 1984 is clearly different from Ophiophyllum nesisi sp. nov. in having larger disk plates, a rounded shape of the radial shields, considerably fewer block-shaped oral papillae, and two distinct tentacle scales. Ophiophyllum novacaledoniae Vadon, 1991 has fewer dorsal disk plates compared to the distinct primary plates, large triangular radial shields, triangular dorsal arm plates, and two distinct tentacle scales. All these characters are enough to separate the latter species from Ophiophyllum nesisi sp. nov. Finally O. marginatum A.H. Clark, 1918 is clearly separated from Ophiophyllum nesisi sp. nov. by the presence of only ventral arm spines and two distinct tentacle scales.
Editorial responsibility: Christoffer Schander
Acknowledgements
We are greatly indebted to Tanya Korshunova (Institute of Higher Nervous Activity and Neurophysiology RAS, Moscow) for preparing most of the plates and digital images. Andrey V. Gebruk (P.P. Shirshov Institute of Oceanology) and Endre Willassen and Jon Kongsrud (Zoological Museum, University of Bergen) made it possible for one of the authors (N.M. Litvinova) to visit the Bergen Museum and examine the MAR-ECO ophiuroids. Special thanks to Frederick J. Collier (Museum of Comparative Zoology, Harvard University) and Martha Collier who made possible the visit of one of the authors (A.V. Martynov) to the Museum of Comparative Zoology to work on the ophiuroid type material. Our sincere gratitude goes to Cynthia Ahearn (National Museum of Natural History, Smithsonian Institution), who kindly lent H.L. Clark's ophiuroid type materials to us and generously helped with photographing numerous type specimens. We are especially grateful to Sabine Stöhr (Swedish Museum of Natural History) for improvement of the English, suggestions on the manuscript and making photographs of the type specimens of the Müller and Troschel ophiacanthids. We are grateful to Eric A. Lazo-Wasem (Peabody Museum of Natural History, Yale University) for discussion, loan of Verrill's ophiacanthid type specimens and making photographs of some types. We deeply appreciate Jon Kongsrud (Zoological Museum, University of Bergen), who kindly made photographs of the type series of Ophioplinthus pseudotessellata sp. nov. Konstantin R. Tabachnik (P.P. Shirshov Institute of Oceanology) kindly photographed ophiuroid specimens from the MAR-ECO expedition. We are thankful to Jessica D. Cundiff (Museum of Comparative Zoology, Harvard University) for photographs of some ophiuroid types. Many thanks to Jens M. Bohn and Eva Lodde (Zoologische Staatssammlung, München) for help during the visit of one of the authors to ZSM. Some ophiacanthid types were photographed during the visit of one of the authors (A.V. Martynov) to the Museum of Comparative Zoology, Harvard University, supported by The Ernst Mayr Travel Grant in Animal Systematics. Andrey V. Gebruk and Alan J. Southward (Marine Biological Association, Plymouth) kindly improved the English. Three anonymous referees kindly helped to improve the manuscript. This work was an element of MAR-ECO, a field project under the Census of Marine Life programme.
Related Research Data
References
- Baranova ZI , Kunzevich ZV . 1969 . Ophiuroidea . In: Zenkevich LA Biology of the Pacific Ocean, 2 . Moscow : Nauka . p 115 – 24 . Russian .
- Bartsch , I . 1982 . Ophiuroidea (Echinodermata) from the Patagonian shelf . Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut , 79 : 211 – 50 .
- Bartsch , I . 1987 . Notes on Ophiuroidea (Echinodermata) from the northeastern Atlantic Ocean. I. Ophiacanthidae . Spixiana , 10 ( 2 ) : 115 – 30 .
- Bartsch , I . 1991 . Schlangensterne (Ophiuroidea) in NO-Atlantik, im der ibero-marokkanisch-mauretanischen Region . Spixiana , 14 ( 1 ) : 95 – 112 .
- Bergstad , OA and Gebruk , AV . 2008 . Approach and methods for sampling of benthic fauna on the 2004 MAR-ECO expedition to the Mid-Atlantic Ridge . Marine Biology Research , 4 : 160 – 163 .
- Bernasconi , I and D'Agostino , MM . 1975 . Equinodermos antarticos. III. Ofiuroideos. 1). Ofiuroideos del extremo norte de la Peninsula Antartica. Revista Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’ . Hidrobiologia , 4 ( 2 ) : 80 – 133 .
- Cherbonnier , G and Sibuet , M . 1972 . Résultats scientifiques de la campagne Noratlante: Astérides et Ophiurides. Bulletin du Muséum National d'Histoire Naturelle, Paris, Série 3 . Zoologie , 76 ( 102 ) : 1333 – 94 .
- Clark , AM . 1970 . Notes on the Amphiuridae (Ophiuroidea) . Bulletin of the British Museum (Natural History) Zoology , 41 : 3 – 81 .
- Clark , HL . 1911 . North Pacific ophiurans in the collection of the United States National Museum . Bulletin of the United States National Museum , 75 : 1 – 302 .
- Clark , HL . 1915 . Catalogue of recent ophiurans based on collections of the Museum of Comparative Zoology. Memoirs of the Museum of Comparative Zoology . Harvard , 25 ( 4 ) : 165 – 376 .
- Dyakonov AM. Ophiury (Zmeekhvostki) Morei SSSR. Opredeliteli po faune SSSR. 1954 Vol. 55 . Moscow: Zoological Institute. 135 p. Russian.
- Farran , GP . 1913 . The deep water Asteroidea, Ophiuroidea and Echinoidea of the West coast of Ireland. Scientific Investigations, 1912, Fisheries Branch, Department of Agriculture for Ireland . Dublin , 6 : 1 – 66 .
- Fell , HB. 1961 . The fauna of the Ross Sea. Part 1. Ophiuroidea . New Zealand Department of Scientific and Industrial Research Bulletin , 142 : 9 – 79 .
- Gage , JD , Pearson , M , Clark , AM , Paterson , GLJ and Tyler , PA . 1983 . Echinoderms of the Rockall Trough and adjacent areas . Bulletin of the British Museum (Natural History) Zoology , 45 ( 5 ) : 263 – 308 .
- Grieg , JA . 1921 . Echinodermata. Report on the Scientific Results of the Michael Sars North Atlantic Deep-Sea Expedition 1910 . Bergen , 3 ( 2 ) : 1 – 47 .
- Hertz , M . 1927a . Die Ophiuroiden der deutscher Tiefsee-Expedition . Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer Valdivia 22 , 1898–1899 ( 3 ) : 59 – 122 .
- Hertz , M . 1927b . Die Ophiuroiden der Deutschen Sudpolar-Expedition 1901–1903 . Deutsche Sudpolar-Expedition 1901–1903 , 19 ( Zoologie 11 ) : 1 – 56 .
- Hunter , RL . 2007 . Morphological cladistic analysis of Ophiurolepis Matsumoto, 1915 (Ophiurida: Ophiuridae) from the Southern Ocean . Zootaxa , 1401 : 33 – 51 .
- Kaufman , ZS . 1974 . Sexual cycles and gametogenesis of invertebrates from the White Sea . Issledovaniya po Faune Morei , 13 ( 21 ) : 191 – 271 .
- Koehler , R . 1895 . Dragages profonds exécutés à bord du Caudan dans le Golfe de Gascogne. Rapport préliminaire sur les échinoderms . Revue Biologique du Nord de la France , 7 : 439 – 96 .
- Koehler , R . 1896a . Résultats scientifiques de la campagne du Caudan dans le Golfe de Gascogne. Echinoderms. Annales de l . Université de Lyon , 26 : 33 – 122 .
- Koehler , R . 1896b . Note préliminaire sur les ophiures recueillis pendant les campagnes de la Princesse Alice . Mémoires de la Société Zoologique de France , 9 : 203 – 13 .
- Koehler , R . 1897 . Echinodermes recueillis par l'Investigator dans l'Océan Indien. Les Ophiures de mer profonde . Annales des Sciences Naturelles Zoologie , 8 ( 4 ) : 277 – 372 .
- Koehler , R . 1898 . Echinides et Ophiures provenant des campagnes du yacht l'Hirondelle . Résultats des Campagnes Scientifiques Accomplies par le Prince Albert I Monaco , 12 : 1 – 78 .
- Koehler , R . 1901a . Note préliminaire sur quelques Ophiures nouvelles provenant des campagnes de la Princesse Alice . Bulletin de la Société Zoologique de France , 26 : 222 – 31 .
- Koehler R. 1901b . Échinides et ophiures. Résultats du Voyage du S.Y. Belgica en 1897–1898–1899 , Rapports Scientifiques, Zoologie 3 – 42 .
- Koehler , R . 1906 . Description des Ophiures nouvelle recueillies par le Travailleur et Talisman pendant les campagnes 1880, 1881, 1882 et 1883 . Mémoires de la Société Zoologique de France , 19 : 5 – 34 .
- Koehler , R . 1907a . Ophiures . Expéditions Scientifiques du Travailleur et du Talisman , 7 : 245 – 311 .
- Koehler , R . 1907b . Note préliminaire sur quelque Astéries provenant des campagnes de la Princesse Alice. Bulletin de l . Institut Océanographique , 99 : 1 – 47 .
- Koehler , R . 1908 . Astéries, Ophiures et Échinides de l'Expédition Antarctique Nationale Écossaise . Transactions of the Royal Society of Edinburgh , 46 ( 3 ) : 529 – 649 .
- Koehler , R . 1909 . Echinoderms provenant des campagnes du yacht Princesse Alice . Résultats des Campagnes Scientifiques Accomplies par le Prince Albert I Monaco , 34 : 1 – 317 .
- Koehler , R . 1911 . Astéries, Ophiures et Echinides . British Antarctic Expedition 2 , 1907–1909 ( 4 ) : 25 – 66 .
- Koehler , R . 1914 . A contribution to the study of ophiurans of the United States National Museum . Bulletin of the United States National Museum , 84 : 1 – 173 .
- Koehler , R . 1922 . Ophiuroidea. Scientific Reports . Australasian Antarctic Expedition 8 , 1911–1914 ( 2 ) : 1 – 98 .
- Koehler , R . 1923 . Astéries et Ophiures . Further Zoological Results of the Swedish Antarctic Expedition , 1 ( 1 ) : 5 – 145 .
- Litvinova NM. 1971 . The brittle stars of the genus Amphiophiura of the Pacific and Indian Oceans collected by Soviet expeditions on R/V ‘Vityaz’ and ‘Akademik Kurchatov’ . Trudy Instituta Okeanologii 92 : 298 – 316 . Russian.
- Ljungman , AV . 1867 . Ophiuroidea viventia huc usque cognita enumerat . Ofversigt af Kungliga Svenska Vetenskapsakademiens Handlingar Stockholm , 23 ( 9 ) : 303 – 36 .
- Lütken , CF and Mortensen , T . 1899 . Reports on an exploration off the west coasts of Mexico, Central and South America, and off the Galapagos Islands, in charge of Alexander Agassiz by the United States Fish Commission steamer Albatross, during 1891. 25. The Ophiuridae. Memoirs of the Museum of Comparative Zoology . Harvard , 23 ( 2 ) : 97 – 207 .
- Lyman , T . 1875 . Ophiuridae and Astrophytidae. Zoölogical Results of the Hassler Expedition, Illustrated Catalogue of the Museum of Comparative Zoölogy . Harvard , 8 ( 2 ) : 4 – 34 .
- Lyman , T . 1878 . Ophiuridae and Astrophytidae of the exploring voyage of H.M.S. Challenger. Bulletin of the Museum of Comparative Zoology . Harvard , 5 ( 7 ) : 65 – 168 .
- Lyman , T . 1882 . Report on the Ophiuroidea. Report on the Scientific Results of the Voyage of H.M.S. Challenger 1873–1876 . Zoology , 5 ( 14 ) : 1 – 386 .
- Lyman , T . 1883 . Report on the Ophiuroidea. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Caribbean Sea (1878–1879), and on the east coast of the United States, during the summer of 1880, by the U.S. coast survey steamer ‘Blake’. Bulletin of the Museum of Comparative Zoölogy . Harvard , 10 ( 6 ) : 227 – 87 .
- Madsen , FJ . 1967 . Ophiuroidea. BANZ Antarctic Research Expedition 1929–1931, Reports . Series B (Zoology and Botany) , 9 ( 3 ) : 123 – 45 .
- Madsen , FJ . 1977 . The Ophioleucidae (Ophiuroidea) . Galathea Report , 14 : 109 – 22 .
- McKnight , DG . 1967 . Echinoderms from Cape Hallett, Ross Sea . New Zealand Journal of Marine and Freshwater Research , 1 : 314 – 23 .
- Matsumoto , H . 1915 . A new classification of the Ophiuroidea . Proceedings of the Academy of Natural Sciences of Philadelphia , 67 : 43 – 92 .
- Menzies RJ George RY Rowe GT 1973 . Abyssal Environment and Ecology of the World Ocean . New York : John Wiley Interscience . 590 p.
- Mortensen , T . 1913 . Some new echinoderms from Greenland . Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening i Kjøbenhavn , 66 : 37 – 43 .
- Mortensen , T . 1933 . Ophiuroidea . Danish Ingolf Expedition , 4 ( 8 ) : 5 – 121 .
- Mortensen , T . 1936 . Echinoidea and Ophiuroidea . Discovery Reports , 12 : 199 – 348 .
- O'Hara TD , Stöhr S . 2006 . Deep water Ophiuroidea (Echinodermata) of New Caledonia: Ophiacanthidae and Hemieuryalidae . In: B Richer De Forges , JL Justine Tropical Deep-Sea Benthos. 24. Mémoires du Muséum National d. Histoire Naturelle 193 : 33 – 141 .
- Paterson , GLJ . 1985 . The deep-sea Ophiuroidea of the North Atlantic Ocean . Bulletin of the British Museum (Natural History) Zoology , 49 ( 1 ) : 1 – 162 .
- Paterson , GLJ , Tyler , PA and Gage , JD . 1982 . The taxonomy and zoogeography of the genus Ophiocten (Echinodermata: Ophiuroidea) in the North Atlantic Ocean . Bulletin of the British Museum (Natural History) Zoology , 43 ( 3 ) : 109 – 28 .
- Sars , GO . 1871 . Nye Echinodermer fra den Norske kyst . Videnskabelige Selskabs Forhandlingar , 1871 : 1 – 31 .
- Sars M . 1861 . Oversigt af Norges Echinodermer . Christiania (Norway) . 160 p.
- Smirnov IS. 2006 . Taxonomic list of the ophiures of the Southern Ocean [online]. ECOANT . Available from: www.zin.ru .
- Stöhr , S . 2003 . A new fissiparous amphiurid brittlestar (Echinodermata: Ophiuroidea) from southwest of Iceland . Sarsia , 88 : 373 – 8 .
- Stöhr , S and Segonzac , M . 2005 . Deep-sea ophiuroids (Echinodermata) from reducing and non-reducing environments in the North Atlantic Ocean . Journal of the Marine Biological Association of the United Kingdom , 85 : 383 – 402 .
- Studer , T . 1876 . Über Echinodermen aus dem antarktischen Meere und zwei neue Seeigel von den Papua-Inseln, gesammelt auf der Reise S.M.S. Gazelle um die Erde. Monatsberichte der Königlichen Preussischen Akademie der Wissenschaften . Berlin , 1876 : 452 – 65 .
- Svoboda , A , Stepanjants , SD and Smirnov , IS . 1995 . Zwei polare Hydractiniden-Arten (Hydroida, Cnidaria) als Symbionten nahe verwandter Schlangensterne (Ophiolepididae, Echinodermata) – ein okologisches Beispiel fur Bipolaritat . Berichte zur Polarforschung , 155 : 86 – 9 .
- Tyler , PA , Emson , RH , Sumida , P and Howell , K . 2005 . Ophiuroid distribution at sublittoral and bathyal depths round the Faroe Islands, NE Atlantic Ocean . BIOFAR Proceedings , 2005 : 175 – 94 .
- Tyler , PA and Gage , JD . 1982 . The reproductive biology of Ophiacantha bidentata (Echinodermata: Ophiuroidea) from the Rockall Trough . Journal of the Marine Biological Association of the United Kingdom , 62 : 45 – 55 .
- Verrill , AE . 1884 . Notice on the remarkable marine fauna occupying the outer banks off the southern coast of New England, No. 10 . American Journal of Science , 28 : 213 – 20 .
- Verrill , AE . 1885a . Notice of the remarkable marine fauna occupying the outer banks off the southern coast of New England, No. 11 . American Journal of Science , 29 : 149 – 57 .
- Verrill , AE . 1885b . Results of the explorations made by the Steamer Albatross off the northern coast of the United States in 1883 . Reports of the United States Commission on Fisheries , 1883 : 503 – 699 .
- Verrill , AE . 1894 . Descriptions of new species of starfishes and ophiurans (with a revision of certain species formerly described; mostly from the collections made by United States Commission of Fisheries) . Proceedings of the United States National Museum , 17 : 245 – 97 .
- Verrill , AE . 1899 . Report on the Ophiuroidea; collected by the Bahama Expedition from the University of Iowa in 1893 . Bulletin from the Laboratories of Natural History of the State University of Iowa , 5 ( 1 ) : 1 – 86 .
- Wyville Thomson C. 1873 . The Depths of the Sea . London : Macmillan . 527 p.