Abstract
The Icelandic pectinoid fauna is reviewed, based on material from the benthic survey programme BIOICE and 17 species are recorded. Similipecten oskarssoni is proposed as a replacement name for Pecten groenlandicus var. minor Locard, Citation1898 (Propeamussiidae), which is considered a valid species. Lectotypes are designated for Pecten groenlandicus var. minor and Pecten frigidus Jensen, Citation1904 (Pectinidae). Cyclopecten ambiannulatus Schein, Citation1989, Parvamussium propinquum (Smith, 1885), Catillopecten eucymatus (Dall, Citation1898), and Aequipecten opercularis (Linnaeus, Citation1758) were previously not known from the Icelandic fauna. All species are figured and described and distinguishing characters are discussed. The distribution in Scandinavian and North Atlantic waters is revised based on the collections in SMNH. The low frequency of juvenile specimens of A. opercularis, Mimachlamys varia and Pecten maximus may indicate that Scandinavian populations are the result of very occasional recruitment.
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
Introduction
Iceland occupies a zoogeographic key position in the North Atlantic (Ekman Citation1935; Dahlgren et al. Citation2000), on the Greenland–Scotland Ridge that separates the Arctic abyssal basins from the North-Atlantic deep sea (Bouchet & Warén Citation1979). At the same time, the ridge forms a connection between the European and the American more shallow faunas, with Iceland and the Faroes as ‘stepping stones’. (For further references, see Gudmundsson Citation1998 and Dahlgren et al. Citation2000.) The Icelandic fauna is therefore of considerable interest for evaluating the possibilities for genetic communication between East and West Atlantic populations. The presence of a species in Iceland enhances the possibilities for genetic communication between American and European populations. The crab Lithodes maia occurs in the eastern and western Atlantic, and Iceland. Another well-known decapod, Homarus, does not occur in Iceland, and the East and West Atlantic populations have diverged to H. gammarus and H. americanus (Harding et al. Citation1997; Kit & Kornfield Citation1998).
On the other hand, Iceland is distant enough from Europe to cut off much of the genetic communication by planktonic larvae, and it is only a few decades since the snail Hydrobia ventrosa (found 1971, Oskarsson et al. Citation1977), and the bivalves Cerastoderma edule (found 1948), and Mya arenaria (found 1958, Oskarsson Citation1982), were naturalized there, probably spread by human activities. This suggests that the capacity of dispersal of these species is insufficient for the step to Iceland. The periwinkle Littorina littorea has dispersed to Iceland after the latest glaciation (Leifsdóttir & Simonarson Citation2001), despite seemingly favourable environmental conditions, and is the only mollusc absent in Iceland but present in both North America and northern Norway; additional support for the opinion that it was transferred to the western Atlantic by human activities. (See also Wares et al. Citation2002, who erroneously believed that it had occurred in Iceland during the Atlantic Stade, ca. 8000 years ago.)
Iceland is also a southern outpost of the arctic fauna, with many classical arctic species like Yoldia limatula, Thracia myopsis, several Buccinum, Colus, and Boreotrophon, although the most high-arctic species like Portlandia arctica, Macoma loveni and Pandora glacialis do not reach it. The arctic influence is most important along the eastern and north-eastern coasts (Thorson Citation1941; Madsen Citation1949).
Another element, the Mid-Atlantic Ridge fauna, i.e. species occurring along the ridge, sometimes only on its peaks, seems not to be represented among the pectinoids. The pectinoids of this fauna, south of Iceland, were treated by Dijkstra & Gofas (Citation2004).
The Greenland–Scotland Ridge separates the Arctic abyssal basins from the abyssal parts of the North Atlantic. The ridge usually reaches up to a minimum depth of 300–400 m, but between the Faroes and Shetland it is transected by a pass which allows communication at a depth of ca. 750 m. Here, the current is a persistent outflow of cold (ca. –0.5 to –1.5°C), low-salinity, arctic bottom water. Such water is also to a lesser extent washing over the ridges and the Denmark Strait between Greenland and Iceland (Hansen et al. Citation2001). Since all the arctic abyssal species which can be carried out with the bottom currents have lecithotrophic larvae, this means that the communication between these abyssal faunas becomes unidirectional, as emphasized by Bouchet & Warén (Citation1979).
The Icelandic pectinoid fauna was most recently listed by Ockelmann (Citation1959) and Oskarsson (Citation1982); the latter, however, only including the uppermost bathyal and more shallow species. Ockelmann left two species unidentified/undescribed: Cyclopecten sp. aff. imbrifer (here considered a synonym of Cyclopecten hoskynsi) and Arctinula sp. b (here considered a valid species, and renamed Similipecten oskarssoni). The access to new material from a great range of depths and localities (see ) presented an opportunity to address these and other taxonomic problems within the superfamily. Together with the recent review of pectinoids from North Atlantic seamounts (Dijkstra & Gofas Citation2004) this gives a total coverage of the Atlantic pectinoids north of ca. 40°N; offshore, south to ca. 30°N.
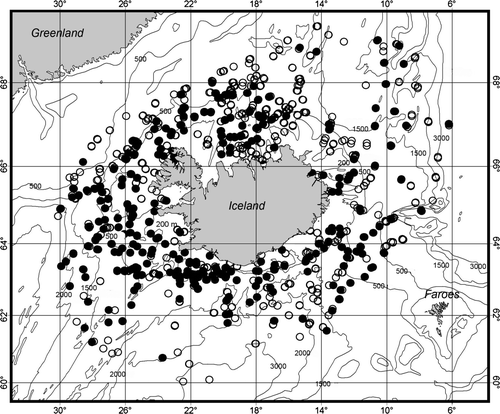
Map 1. Distribution of BIOICE stations; dots mark collecting operations yielding pectinoids, circles, no pectinoids.
In general, the classification and terminology follow Waller (Citation1972, Citation1984, Citation1991, Citation1993, Citation2006) and Clemam (Check List of European Marine Mollusca: www.mnhn.fr/mnhn/bimm/clemam/). Some recent suggestions in generic nomenclature have not been implemented, awaiting additional genetic support. All types listed have been examined by Dijkstra.
In order to build a better foundation for the future classification of the Pectinoidea, we have illustrated much of the variation in micro-sculpture and larval shells among the species, which has been sadly neglected previously. We have also emphasized illustrations of the poorly known species and young specimens.
Abbreviations and acronyms used in the text
BMNH, The Natural History Museum, London; HD, H.H. Dijkstra private pectinoid collection, Sneek, Netherlands; HL, Hans Larsen, Reykjavik; IMNH, Icelandic Museum of Natural History, Reykjavik; JB, Jón Bógason, Kópavogur; KBIN, Koninklijk Belgisch Instituut voor Natuurwetenschappen, Brussels; LSL, Linnean Society, London; MNHN, Muséum national d'Histoire naturelle, Paris; MOM, Musée Océanographique, Monaco; NNM, National Museum of Natural History, Naturalis, Leiden; NMW, National Museum & Gallery, Cardiff; SMNH, Swedish Museum of Natural History, Stockholm; USNM, US National Museum of Natural History, Washington, DC; ZMA, Zoological Museum, University of Amsterdam; ZMC, Zoological Museum, University of Copenhagen; ZMO, Zoological Museum, University of Oslo. Spm, pair of (joint) valves; v, valve(s); C, convexity (depth) of articulated shell; H, height (dorsal–ventral measurement); L, length (anterior–posterior measurement); Ø (used in pictures for practical reasons), largest diameter of figured specimen.
Material and methods
The samples were collected around Iceland during the BIOICE project (Benthic Invertebrates of Icelandic waters), from the research vessels Bjarni Sæmundsson, Håkon Mosby and Magnus Heinason (Gudmundsson Citation1998) from 1992 to 2006. Pectinoids were found in 356 samples (of a total of 1033 benthic samples) at depths between 19 and 3007 m (79 hauls more shallow than 100 m average depth and 150 hauls deeper than 1200 m; see ). The sampling devices were a modified RP sled (Rothlisberg & Pearcy Citation1976; Brattegard & Fosså Citation1991), Sneli sled (Sneli Citation1998), Agassiz trawl, and triangular dredge. The RP sled is towed along the bottom and collects animals, which are stirred up by turbulence in front of the sampler. The net of the sled has 0.5 mm mesh in the cod end. The Sneli sled ploughs into the sediment and works well on soft and mixed substrates, where the more fragile RP sled cannot be towed. The samples were sieved through a series of sieves down to 0.5 mm mesh width, fixed in 4% buffered formalin and later transferred to 80% ethanol. All samples were sorted under a stereomicroscope at the Sandgerdi Marine Centre.
We have in some cases supplemented the BIOICE material with information from the large private collection of Jón Bógason, Kópavogur (JB). His material originates from collaboration with fishing boats and from his work at the Marine Research Institute, Reykjavik. Most of his collection is now in IMNH. Further information on distribution comes from Hans Larsen (HL), volunteer at the Icelandic Museum, who sorted and made the initial identifications of most molluscs from the BIOICE programme.
The BIOICE material is deposited in IMNH, with some voucher specimens in the private pectinoid reference collection of the senior author, SMNH, and at ZMA.
Propeamussiids are often difficult to work on with regular methods for shell studies. The shells are fragile and as noticed by Jensen (Citation1912), this has given many authors an impression that the right valve is smaller than the left one. They are actually rather much the same size, but the contraction when preserved causes the margin of the right valve to break off commarginally, so neatly so it is hardly to be noticed as damage. Figures E and B show specimens where this is visible; Figure B shows a specimen reduced in size in this way.
The shells are also difficult to clean for examination of surface structures. Normal cleaning with diluted bleach destroys the organic matrix and causes the outer layers to peel; 3% hydrogen peroxide and a trace of sodium hydroxide was more successful. Ultrasonic cleaning can only be used with the more sturdily built shallow-water species. Soaking with a neutral detergent, weak heating (<50°) and a fine paintbrush turned out the best way for cleaning.
Dimensions of larval shells were based on measurement of its length in 1–4 specimens with SEM and additional checks under stereomicroscope with an accuracy of ±10 µm, but no statistics were kept.
The total list of pectinoid records on which this paper is based is available as supplementary online material at http://www.informaworld.com/mpp/uploads/dijkstraeta_suppmat.doc. This list is the base for the distributional Maps 2–9.
Maps 2–5. Distribution of pectinoids collected by BIOICE. (2) Propeamussium lucidum, dots; Hyalopecten frigidus, triangles; Cyclopecten hoskynsi, squares. (3) Parvamussium propinquum, triangles; Pseudamussium peslutrae, dots. (4) Palliolum striatum, triangles; Pseudamussium sulcatum, dots. (5) Similipecten greenlandicus, dots; S. oskarssoni, triangles.
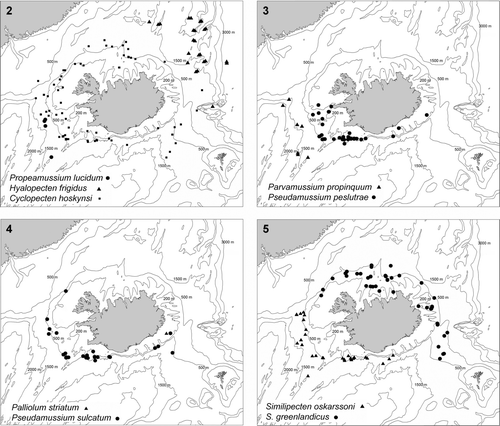
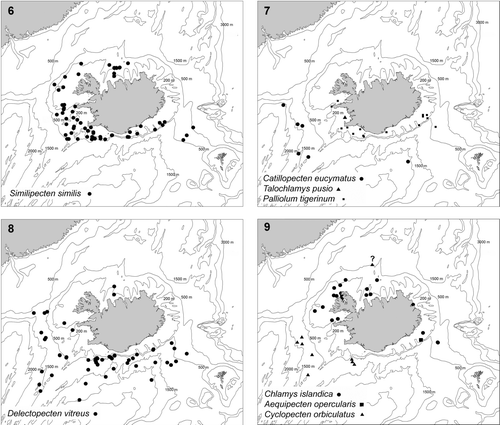
Maps 6–9. Distribution of pectinoids collected by BIOICE. (6) Similipecten similis. (7) Catillopecten eucymatus, dots; Talochlamys pusio, triangles; Palliolum tigerinum, squares. (8) Delectopecten vitreus, dots. (9) Chlamys islandica, dots; Aequipecten opercularis, square; Cyclopecten ambiannulatus, triangles.
All North Atlantic material of Pectinoidea in SMNH, upon which much of the information on distribution is based, is now available via the Global Biodiversity Information Facility (GBIF; http://www.gbif.net).
A complete set of high-resolution colour plates can be obtained from A. Warén ([email protected]).
Taxonomy
Superfamily Pectinoidea Rafinesque, 1815
Family Propeamussiidae Abbott, 1954
Genus Propeamussium De Gregorio, 1884
Propeamussium lucidum (Jeffreys in Thomson, 1873)
(Figures A,B, F, E,F, I,J, I,J)
Pleuronectia lucida Jeffreys in Thomson (1873: 464, fig. 78a [original description]).
Amussium hypomeces Dautzenberg & Fischer (Citation1897: 195, pl. 5, figs 11 and 12). Type material: Syntypes, MOM 28 0059–63, 28 0066, 28 070, MNHN (no register number), KBIN, Dautzenberg colln I.G. 10591, NMW Z.1955.158.02237. Type locality: The Azores, 800–1385 m depth. For additional data see Dautzenberg and Fischer (1897:140–1).
Amussium lucidum; Jensen (Citation1912:37, pl. 2, figs 3a–c [description, distribution]).
Propeamussium centobi Schein (1989:93, pl. 8, figs 1–6). Type material: Holotype, MNHN (no register number). Type locality: North of Bay of Biscay, 4729.6'N, 0904.5'W, 2878 m depth.
Propeamussium lucidum; Morton & Thurston (Citation1989:471–96, figs. 1–18 [functional morphology]); Dijkstra & Goud (Citation2002:34, figs 1–4, 39, 40 [references, synonymy, description, distribution, discussion]).
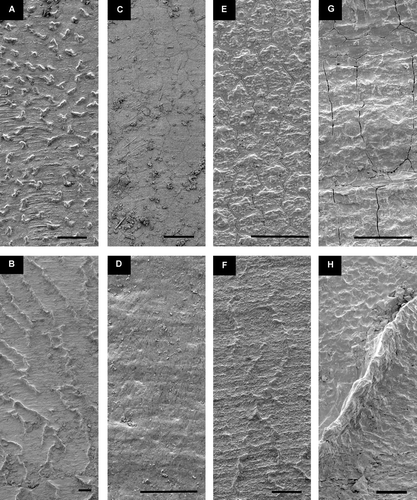
Figure 1. Thin-shelled deep-sea Pectinoidea. (A,B) Propeamussium lucidum, Ø 5.0 and 4.7 mm, BIOICE#2701, off western Iceland, depth 1121 m (SMNH 55517). (C,D) Catillopecten eucymatus, (C) Ø 4.5 mm, BIOICE#2914, off western Iceland, depth 2005 m (SMNH 55526). (D) Ø 3.6 mm, BIOICE#2707, off southwestern Iceland, depth 1407 m (SMNH 55521). (E,F) Cyclopecten hoskynsi, Ø 6.0 and 5.5 mm, BIOICE#2268, off southwestern Iceland, depth 450 m (SMNH 55534). (G,H) Cyclopecten ambiannulatus, Ø 5.2 and 4.6 mm, BIOICE#2707, off southwestern Iceland, depth 1407 m (SMNH 55519). (I,J) Parvamussium propinquum, Ø 9.0 mm, BIOICE#2706, off southwestern Iceland, depth 1406 m (SMNH 55512). (K,L) Hyalopecten frigidus, Ø 9.0 and 10 mm, BIOICE#3214, NE of Iceland, 3000–3003 m (SMNH 59237).

Figure 2. Larval right valves. Scale lines 50 µm. Arrows indicate demarcation of prodissoconch 1. The scale line is valid for length only; the height may differ slightly because of tilt. (A) Hyalopecten frigidus, BIOICE#3214, NE of Iceland, 3000–3003 m (SMNH 59237). (B) Similipecten oskarssoni, BIOICE#2401, off southwestern Iceland (SMNH 55525). (C) Similipecten similis, BIOICE#2393, off southwestern Iceland (SMNH 55515). (D) Similipecten greenlandicus, BIOICE#2047, off northeastern Iceland (SMNH 55520). (E) Parvamussium propinquum, BIOICE#2706, off southwestern Iceland (SMNH 55512). (F) Propeamussium lucidum, BIOICE#2701, off western Iceland (SMNH 55517). (G) Catillopecten eucymatus, BIOICE#2914, off western Iceland (SMNH 55526). (H) Cyclopecten hoskynsi, BIOICE#2268, off southwestern Iceland (SMNH 55534). (I) Cyclopecten ambiannulatus, BIOICE#2707, off southwestern Iceland (SMNH 55519). (J) Delectopecten vitreus, BIOICE#3067, off southeastern Iceland (SMNH 55531).
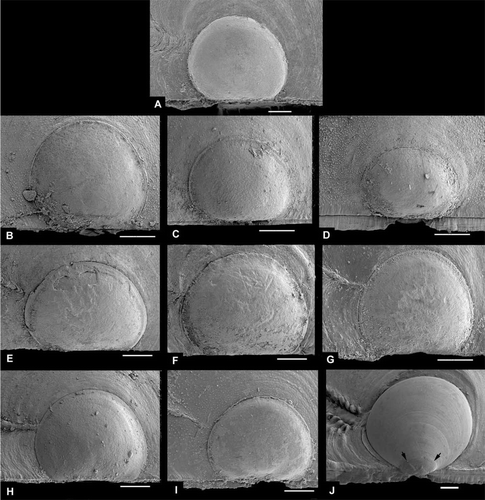
Figure 3. Internally ribbed propeamussiids (all broken at margin). (A–D) Parvamussium propinquum, external and internal view, Ø 9.0 mm, BIOICE#2706, off southwestern Iceland, depth 1406 m (SMNH 55512). (E,F) Propeamussium lucidum, external and internal view, Ø 6.1 mm, BIOICE#2701, off western Iceland, depth 1121 m (SMNH 55517).
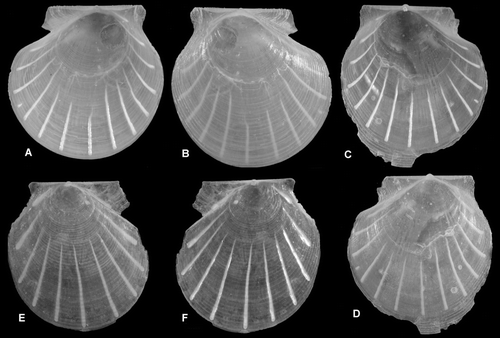
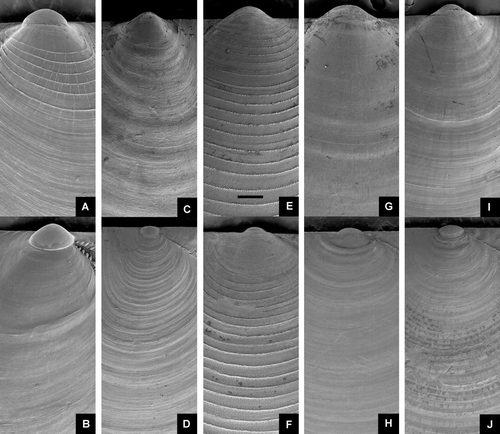
Figure 4. Umbonal area of smooth or concentrically sculptured Pectinoidea. Left valve above. All at same magnification. Scale line (see Figure E) 250 µm. (A,B) Delectopecten vitreus, BIOICE#3067, off southeastern Iceland, depth 1729 m (SMNH 55531). (C,D) Catillopecten eucymatus, BIOICE#2707, off southwestern Iceland, depth 1407 m (SMNH 55521), (E,F) Cyclopecten ambiannulatus, BIOICE#2707, off southwestern Iceland, depth 1407 m (SMNH 55519). (G,H) Parvamussium propinquum, BIOICE#2706, off southwestern Iceland, depth 1406 m (SMNH 55512). (I,J) Propeamussium lucidum, BIOICE#2701, off western Iceland, depth 1121 m (SMNH 55517).
Type material
Lectotype, BMNH 85.11.5.590, designated by Dijkstra & Gofas (Citation2004:36), paralectotypes, USNM 62336, 62346, 62348, BMNH 77.11.28.8, BMNH 85.11.5.156.
Type locality
Off the British Isles, 1019 m depth. For additional data see Warén (Citation1980:43).
Distribution
Northern to tropical eastern Atlantic (64N–15N), south of Greenland, southwest of Iceland and south of the Greenland–Scotland Ridge. Bathymetric range from ca. 785 m (Dijkstra & Goud Citation2002) to 4453 m (Morton & Thurston Citation1989). BIOICE material alive at four stations in 1042–1327 m ().
Description
Shell (Figures A,B, E,F) up to ca. 15 mm high, fragile, usually transparent, rarely opaque, slightly higher than long, left valve slightly more convex than right valve, equilateral (anterior and posterior end of disc of similar shape); anterior auricles larger than posterior ones. Prodissoconch (Figures F), length ca. 200–230 µm, white.
Left valve almost smooth, sometimes irregularly sculptured with weak radial and/or commarginal striae or delicate lirae (Figures I, I). Right valve with regularly spaced, commarginal lirae (Figures J, J). Anterior auricle dorsally spiny. Internal radial ribs generally 9, rarely 8 or 10, with a few rudimentary interstitial and 2 auricular ones. Byssal notch small, ctenolium (series of teeth along the edge of the shell, close to the anterior ears) lacking.
Remarks
Propeamussium lucidum seems to be scarce off Iceland (Ockelmann Citation1959), but rather common in the temperate and subtropical regions of the eastern Atlantic (Salas Citation1996; Dijkstra & Goud Citation2002; Dijkstra & Gofas Citation2004). Its supposed occurrence in the western Atlantic seems to be based on Jeffrey's old record (see below).
The BIOICE specimens are very similar to the type material. Propeamussium lucidum varies in sculpture on the left valve, usually very weak radial striae, sometimes with a few radial ribs anteriorly and posteriorly. A weak reticulated sculpture is formed when both radial and commarginal striae are developed. Specimens from the temperate East Atlantic are nearly smooth (as in the types), whereas more southern specimens are weakly sculptured (P. hypomeces morphs: weak commarginal sculpture, and P. centobi morphs with weak radial sculpture or weak reticulate sculpture; based on examination of specimens in BMNH, MNHN, NNM). For discussion of geographical morphs see Dijkstra & Goud (Citation2002) and Dijkstra & Gofas (Citation2004).
Jeffreys’ (Thomson Citation1873:465) figures of Pleuronectia lucida belong to two species: the North Atlantic P. lucidum (Figure 78a, lv), and Propeamussium pourtalesianum (Dall, 1886) from the Gulf of Mexico (Figure78b, rv). A more precise locality ‘Gulf of Mexico, off Alligator Bank, 156 fms’ of the latter specimen was given by Jeffreys (Citation1876:425) and the identification by Jeffreys (in Thomson 1873) was followed by Dall (Citation1881:117), but in 1886 Dall introduced a new species name Amusium (Propeamussium) pourtalesianum for Jeffreys’ ‘Pleuronectia lucida’ from the western Atlantic. Propeamussium pourtalesianum differs from P. lucidum by having a more circular disc (P. lucidum is elongate), 10 instead of 9 internal ribs and a coloured left valve instead of a colourless or transparent one.
Several variations were described by Jeffreys (Citation1879:562) and Locard (1898:407), but they are individual and geographic. Amussium lucidum var. striata Jeffreys, 1879 was based on specimens of the same appearance as P. centobi with ‘distinct longitudinal striae’. We consider it a synonym of P. lucidum.
Schein (1989) gave a length of 170 µm for prodissoconch 2, but both our measurements (ca. 220 µm) and those in Morton & Thurston (Citation1989:476), 200–230 µm, differ considerably. It is obvious that Schein's figure is strongly tilted and this may explain the discrepancy.
Genus Parvamussium Sacco, 1897
Parvamussium propinquum (Smith, 1885)
(1I,J, fig 2E, fig 3A–D, 4G,H, 8C,D)
Amussium propinquum Smith (1885:314, pl. 23, figs 7, 7a,b [original description]).
Amussium sublucidum Dautzenberg & Fischer (1897:193, pl. 5, figs 9, 10). Type material: Syntypes, MOM 28 0088, 28 0090–93, MNHN (no register number), KBIN, Dautzenberg colln I.G. 10591. Type locality not specified: The Azores, 1165–1557 m depth. For additional data see Dautzenberg & Fischer (1897:194).
Parvamussium propinquum; Dijkstra & Goud (Citation2002):38, figs 14–20 [synonymy, description, distribution]); Dijkstra & Gofas (Citation2004):38, figs 3A–E).
Type material
Holotype, BMNH 1887.2.9.3325.
Type locality
The Azores, 1829 m depth.
Distribution
Tropical to temperate eastern Atlantic (ca. 40N–20N), now extended to Iceland (ca. 65N) with a depth range from 790 m (Dijkstra & Goud Citation2002) to 4060 m (Locard 1898). BIOICE material alive at 6 stations in 1295–2007 m ().
Description
Shell (Figures I,J, A–D) up to ca. 7 mm high, fragile, transparent to opaque, nearly circular, somewhat longer than high, slightly drawn out posteroventrally, left valve slightly more convex than right valve; auricles small, nearly equal in size, umbonal angle ca. 125. Prodissoconch (Figures E), length ca. 200 µm, colour whitish.
Left valve (Figures G, C) smooth at central part of disc with fine closely spaced, radial riblets and commarginal lamellae near the margin.
Right valve (Figures H, D) sculptured with regularly arranged commarginal lirae. Hinge line rather long and straight and very weakly sculptured with close-set lamellae. Inside of valve usually with 12 short and narrow radial riblets, sometimes with several rudimentary interstitial ones. Ribs commence near the central part and extend to the submarginal area. Byssal notch well-developed, ctenolium lacking.
Remarks
The Icelandic specimens have a somewhat weaker sculpture than the type material and usually lack the radial sculpture on the left valve. Adult specimens from the Azores often have smaller, intercalated, internal riblets, which are not present in Icelandic specimens.
Parvamussium propinquum differs from Propeamussium lucidum in having a circular circumference (P. lucidum is generally higher than long), larger anterior auricles, a well-developed byssus notch (in P. lucidum, weak and shallow), and in having more numerous internal ribs (in P. propinquum 12, with several rudimentary ones commencing when half-grown; in P. lucidum usually only 9, developing early).
Genus Cyclopecten Verrill, Citation1897
Cyclopecten hoskynsi (Forbes, Citation1844)
(Figures E,F, H, 5B,C, 9C,D, 10G,H)
Pecten hoskynsi Forbes (1844:192 [original description]).
Pecten imbrifer Lovén (1846:185). Type material: Syntypes, SMNH 1999, 2000. Type locality: Finmarken, Norway.Pecten mamillatus M. Sars, Citation1869:256 nom. nud. (fide Friele Citation1873:300).
Pecten pustulosus Verrill (Citation1873:14). Type material: Holotype, USNM 48764. Type locality: Near St. George's Bank, mud, 274 m depth.
Pecten hoskynsi var. major Leche (Citation1878:35 [not Pecten major Young & Bird, 1828]). Type material: Many syntypes in SMNH. Type locality: Not specified, Kara Sea, east to 65°E, 50–225 m depth (not Leche [Citation1883:452] = Parvamussium alaskensis (Dall, 1871).
Pecten lucidus Haren Noman (Citation1881:2, pl. 1, figs 5–8 [not Pecten lucidus Goldfuss, 1836]). Two syntypes, NNM 99593.Type locality not specified: Barents Sea, 219–384 m depth.
Cyclopecten subimbrifer Verrill & Bush in Verrill (1897:84, 92). Type material: Syntypes, USNM 48762, 48766. Type locality: Off New England, 221–571 m depth.
Pecten imbrifer var. lamellosa Posselt & Jensen (Citation1898:13, pl. 1, fig. 1f). Type material: Holotype, SMNH 3802. Type locality: Greenland, Baffin Bay, 415 m depth.
Pecten imbrifer; Jensen (Citation1912:25, pl. 2, figs 1 and2); (var. major).
Pecten imbrifer var. minor Jensen (1912:25 [infrasubspecific], fig. 2a,b [references, Icelandic distribution]).
Pecten imbrifer var. minor; Madsen (Citation1949:35 [Icelandic distribution]).
Propeamussium (Cyclopecten) imbriferum; Ockelmann (Citation1959:66, pl. 2, fig. 1 [references, distribution, biology, discussion]).
Cyclopecten hoskynsi; Soot-Ryen (Citation1958:13 [anatomy]); Lucas (Citation1979:16, figs [synonymy]); Smriglio & Mariottini (Citation1990:4, 5, pl. 2, figs 3a–e); Dijkstra & Goud (Citation2002:41, figs 13, 21–26 [references, distribution, description, discussion]).
Figure 5. Pectinoidea spp. (A) Delectopecten vitreus, unusually strong sculpture, Ø 6.7 mm, N of Norway, 72° 10’ N, 20° 37’ E, depth 360–415 m (SMNH 56311). (B,C) Cyclopecten hoskynsi, Ø 16.1 mm, BIOICE#2758, off northern Iceland, depth 786 m (SMNH 55511). (D,E) Delectopecten vitreus, smooth ‘abyssorum’ form, Ø 8.3 mm, BIOICE#2424, off southwestern Iceland, depth 495 m (SMNH 55513). (F,G) Hyalopecten frigidus, Ø 22 mm, off northern Norway, 73° 28’ N, 10 °00’ E, depth 2346 m (SMNH 55536). (H,I) Cyclopecten ambiannulatus, Ø 4.3 and 5.6 mm (same specimen), BIOICE#2707, off southwestern Iceland, depth 1407 m (SMNH 55519).

Type material
Syntype, National Museums of Scotland, Edinburgh, reg. no. 1976.5.31904.
Type locality
Off Piscopi [= Tilos], Sporades, Greece, 366 m depth (restricted by Smaldon et al. Citation1976:51).
Distribution
Arctic to tropical, eastern and western Atlantic, from Svalbard and northern Greenland southwards to Massachusetts (USA) (Abbott Citation1974:445, as Cyclopecten pustulosus) in the West Atlantic and southwards to the Cape Verde Islands in the East Atlantic, in 82–2064 m (Dijkstra & Goud Citation2002:41; Locard 1898). BIOICE material alive at 77 stations in 125–1314 m ().
Description
Shell (Figures E,F, B,C) up to ca. 24 mm high, usually smaller, fragile, transparent to opaque, somewhat higher than long, flattened, left valve slightly more convex than right valve, anterior auricles larger and longer than posterior ones. Prodissoconch (Figure H), length ca. 180 µm, colour whitish.
Left valve (Figures C, G) sculptured with broad, radially arranged vesicles and small secondary interstitial vesicles commencing in late growth stage or, usually damaged, so only curved lamellae remain, sculpture sometimes nearly absent. Commarginal lamellae usually present near ventral margin, sometimes absent. Anterior auricle with prominent commarginal lamellae and some radial rows of vesicles, posterior auricle similarly but more weakly sculptured.
Right valve (Figures D, H) with closely spaced commarginal lamellae. Auricles rather small, anterior one more prominent sculptured with small vesicles or lamellae. Internally sometimes with rudimentary riblets commencing in late growth stage and extending towards the submarginal area. Byssal notch small, ctenolium lacking.
Remarks
Arctic and northern specimens (SMNH, ZMC), have usually been identified as Cyclopecten imbrifer, and resemble Mediterranean C. hoskynsi in shape and sculpture, although larger (up to ca. 24 mm in height in high arctic areas, ca. 15 mm in northern Iceland; continuously smaller southwards, usually 6–7 mm in the Mediterranean). We cannot find any additional difference and the variation in sizes looks like a cline with BIOICE material of intermediate size. The names Cyclopecten pustulosus and Cyclopecten subimbrifer were based on specimens from the northwestern Atlantic with unusually prominent vesicles on the ribs in pustulosus and weak concentric lamellae and small vesicles in subimbrifer (see Jensen Citation1912 and Schiøtte Citation1989). Ockelmann (Citation1959) separated four species in this group, Cyclopecten imbrifer (arctic), C. subimbrifer (West Atlantic), C. pustulosus (West Atlantic) and Cyclopecten sp. A. (from the Faroes), but also SEM examination of sculpture and larval shells from extreme localities failed to reveal any persisting differences.
Among congeneric species only those from the northern hemisphere have vesicles on the left valve. Cyclopecten carlottensis Bernard, Citation1968, known from the north-eastern Pacific, differs from C. hoskynsi in having coarser commarginal lamellae on the right valve and more closely set rows of pustules on the left one. Also the rows of pustules are usually more prominent than in C. hoskynsi. This species is morphologically very similar to the north-western Atlantic form of C. hoskynsi named C. pustulosus (now considered a synonym of hoskynsi). Another resembling species is Cyclopecten davidsoni (Dall, Citation1897), from Alaska, distinguishable from C. hoskynsi by having a more sturdily built shell, often abruptly increasing the convexity of the valves close to the margin (flattened in C. hoskynsi), and by having narrower radial rows of crowded vesicles.
Cyclopecten ambiannulatus Schein, 1989
(Figures G,H, I, E,F, H,I, E,F)
Cyclopecten ambiannulatus Schein (1989:86, pl. 4, figs 1–9, 11–12).
Type material
Holotype (livetaken) H 6.8 mm, L 6.7 mm, and 3 paratypes (livetaken), MNHN [no register number].
Type locality
North of Bay of Biscay, 4732.8’ N, 835’ W, 2126 m depth, alive, BIOGAS station 1, BIOGAS IV, DS 63, 01.Feb.1974.
Material examined
Iceland: BIOICE 2257, 6315.00′ N, 2629.57′ W, 1209–1212 m, clay and shell sand, 5 spm (IMNH); 2406, 6259.40′ N, 2146.10′ W, 934 m, 3 spm + 1 v (IMNH); 2410, 6251.90′ N, 2143.30′ W, 1074–1075 m, silty sand, 21 spm (IMNH/18, ZMA/3); 2412, 6244.50′ N, 2133.80′ W, 1170–1174 m, silt, 2 spm (IMNH); 2697, 6410.40′ N, 2743.80′ W, 1042 m, 11 spm + 3 v (IMNH/12, SMNH/2); 2704, 6350.80′ N, 2742.00′ W, 1295 m, 18 spm + 1 v (IMNH/17, ZMA/2); 2707, 6355.60′ N, 2817.20′ W, 1407–1408 m, silty sand, 18 spms + 1 v (SMNH/16, ZMA/3); (2740, 6738.38'N, 2015.80'W, 503-512 m, silt, 1 spm (IMNH); 3187, 6209.04′ N, 2700.74′ W, 1326–1327 m, 30 spm (IMNH).
Distribution
Off western Iceland (62–67N, 20–28W), living in 503–1408 m (Map 9). A single specimen is supposed to come from station 2740, but considering the locality, so far further north than the others, it must be considered uncertain, perhaps mislabelled.
Description
Shell (Figures G,H, H,I) up to ca. 7 mm high, fragile, transparent to opaque, circular, left valve slightly more convex than right valve, anterior auricles slightly larger and more demarcated from disc than posterior ones; umbonal angle ca. 110; internal ribs lacking, colour whitish. Prodissoconch (Figure I), length 185 µm, colour white.
Both valves (Figures E,F, E,F) sculptured with uniform and closely spaced commarginal lamellae (ca. 10 per mm on the central part of the disc), commencing in early growth stage directly below the prodissoconch boundary and extending towards the ventral margin. Granular intercalated micro-sculpture. Similar sculpture on the auricles, except anterior auricle of right valve, which is almost smooth. Posterior auricle of right valve continuous with disc, anterior auricle demarcated. Hinge line straight. Byssal notch moderately deep, byssal fasciole narrow, suture somewhat folded, ctenolium lacking.
Discussion
The BIOICE specimens resemble Cyclopecten ambiannulatus Schein, 1989, known from the temperate eastern Atlantic (44–58N). Both species are somewhat similar in shape, size and colour, but differ in sculpture. Cyclopecten orbiculatus has more closely and regularly spaced commarginal lamellae on the left valve, and lacks interstitial radial sculpture. Cyclopecten ambiannulatus has widely spaced lamellae in young specimens and closely spaced ones with delicate interstitial radial lines in adult and subadult specimens (Schein 1989:pl. 6, figures 1–5).
Cyclopecten cincinnatus Dijkstra & Gofas, Citation2004, known from the Mid-Atlantic seamounts (30–34N) differs from C. ambiannulatuus in having much more widely spaced commarginal lamellae on the central part of the left valve (ca. 2 per mm, in C. ambiannulatus 8–10), and in having prominent commarginal lamellae on the anterior auricle of the left valve (in C. ambiannulatus very weak or lacking).
Some specimens of Cyclopecten hoskynsi (‘var. lamellosus’ in Jensen Citation1912) have prominent commarginal lamellae also on the left valve, but these are undulating, widely spaced and irregularly developed, and originate from incompletely formed vesicles. Such specimens can be distinguished from C. ambiannulatus by C. hoskynsi having very small posterior shell ears.
Genus Similipecten Winckworth, 1932
Similipecten greenlandicus (CitationSowerby II, 1842)
(Figures 2D, 6A,B,D–F, 9G,H, 10A,B)
Pecten greenlandicus Sowerby II (1842:57, pl. 13, fig. 40 [original description]).
Pecten groenlandicus [sic]; Jensen (Citation1912:30 [references, Icelandic distribution]); Madsen (Citation1949:36 [references, Icelandic distribution]).
Propeamussium (Arctinula) groenlandicum [sic]; Ockelmann (Citation1959:68, pl. 2, fig. 2 [references, distribution, biology]).
Cyclopecten (Delectopecten) greenlandicus; Grau (Citation1959:53, pl. 20 [references, description]).
Delectopecten greenlandicus; Lubinsky (Citation1980:28, pl. 5, fig. 7 [distribution]).
Similipecten greenlandicus; Knudsen (Citation1985:104 [distribution]).
Arctinula greenlandica; Richling (Citation2000:46, text fig. 49 [distribution]).
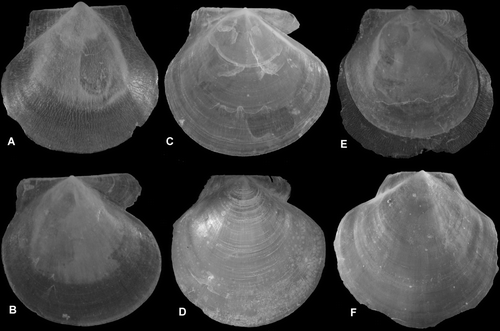
Figure 6. Similipecten spp. (A,B, E) S. greenlandicus, BIOICE#2047, off northeastern Iceland, depth 272 m (SMNH 55520). (A) Left valve, Ø 4.8 mm. (B) Right valve, Ø 4.2 mm (of same specimen). (E) Complete specimen, showing the partly and concentrically broken right valve contained in the left one. (C) S. similis, Ø 6.2 mm, BIOICE#2393, off southwestern Iceland, depth 291 m (SMNH 55515). (D,F) S. greenlandicus, Ø 30 mm, Svalbard, Vest Spitsbergen, depth 38–55 m (SMNH 55510).
Type material
Not found; not in The Hancock Museum at Newcastle upon Tyne (UK) (L. Jessop, personal communication) or in BMNH.
Type locality
Greenland.
Distribution
Arctic region except the Chukchi Sea (north of Bering Sea); in the North Atlantic southwest to the Gulf of St. Lawrence (Clarke Citation1974; Lubinsky Citation1980) southeast to northern Norway (Lofoten), south-eastern Iceland and the Faeroes (Madsen Citation1949; Petersen Citation1968). Living in 4 to ca. 600 m, on fine sediments. Some very deep records (2000 m in Knudsen Citation1985; Richling Citation2000) probably originate from ice drift. BIOICE material alive at 55 stations in 107–1048 m, only two records below 600 m (Map 5).
Description
Shell (Figures A,B,D–F) up to ca. 30 mm high, fragile, almost circular (adult specimens slightly longer than high), weakly convex, left valve somewhat more convex than right valve, equilateral, anterior auricles larger than posterior ones, both nearly equal in length, umbonal angle ca. 120, Greyish semitransparent, internal ribs lacking. Prodissoconch (Figure D), length 160–170 µm, colour white.
Left valve (Figures G, G) with delicate antimarginal, micro-sculpture of irregularly branching and anastomosing raised lines and commarginal growth lines near ventral margin. Auricles with growth lines and slightly raised laterally, anterior auricle more demarcated from disc than posterior one.
Right (Figures H, H) valve smooth with commarginal growth lines. Anterior auricle with closely spaced growth lines and strongly demarcated from disc, anterior margin curved, posterior auricle continuous with disc. Byssal notch small, byssal fasciole narrow, ctenolium lacking.
Remarks
High arctic specimens are much larger, up to 32.5 mm in height in Svalbard (Knipowitsch Citation1901) and 35 mm in East Greenland (Ockelmann Citation1959), than specimens from Iceland and northern Norway (up to 12 mm in height), but they cannot otherwise be separated.
Jeffreys (1879:560) considered certain small, smooth specimens from off western Europe to be young S. greenlandicus; Locard (1898:399) introduced a varietal name (var. minor); and Jensen (1912:32) considered such specimens as a ‘dwarf-form’. Ockelmann (Citation1959:72) had the opinion that the arctic and more southern specimens differed anatomically. Moreover, arctic and West European specimens differ in sculpture of the left valve, which in S. greenlandicus has an antimarginal micro-sculpture of irregular raised lines, while southern specimens have a much weaker sculpture, restricted to the umbo and visible mainly in well preserved specimens. The almost smooth specimens are here considered a distinct species and renamed Similipecten oskarssoni.
Similipecten oskarssoni nom. nov.
(Figures B, B–F, E,F, E,F)
Pecten groenlandicus; Jeffreys (1879:560 [not P. greenlandicus Sowerby]).
Pecten groenlandicus var. minor Locard (1898:399 [original description]).
Propeamussium sp. b Ockelmann (Citation1959:72).
Similipecten similis (Laskey); Schein (1989:96 [pro parte]).
Figure 7. Similipecten spp. (A) Similipecten similis, Ø 3.6 mm, BIOICE#2393, off southwestern Iceland, depth 291 m (SMNH 55515). (B–F) S. oskarssoni, (B,C) Ø 3.75 and 2.3 mm, BIOICE#2401, off southwestern Iceland, depth 520 m (SMNH 55525). (D–F), (D) left side; (E) right side of same specimen, Ø 5.1 mm; (F) right side, not broken, Ø 5.7 mm, BIOICE#2401, off southwestern Iceland, depth 520 m (SMNH 55518). (G–I) S. similis, (G,H) left side, varying colour pattern, Ø 7.1 and 7.9 mm; (I) right side of H; BIOICE#2710, off western Iceland, depth 220 m (SMNH 55516).
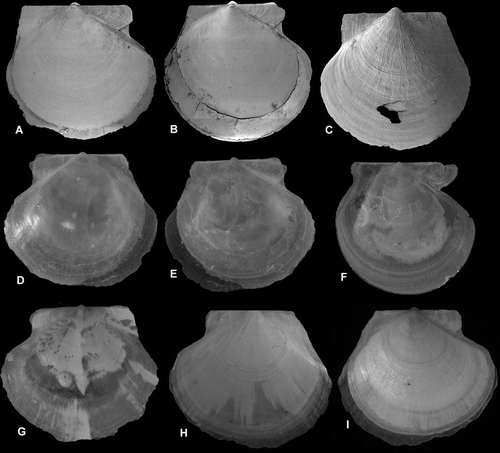
Figure 8. Detail of sculpture, mid-shell of half-grown specimens. Left valve above. Scale lines 25 µm. (A,B) Catillopecten eucymatus, BIOICE#2914, off western Iceland, depth 2005 m (SMNH 55526). (C,D) Parvamussium propinquum, BIOICE#2706, off southwestern Iceland, depth 1406 m (SMNH 55512). (E,F) Cyclopecten ambiannulatus, BIOICE#2707, off southwestern Iceland, depth 1407 m (SMNH 55519). (G,H) Delectopecten vitreus, BIOICE#3067, off southeastern Iceland, depth 1729 m (SMNH 55531). (I,J) Propeamussium lucidum, BIOICE#2701, off western Iceland, depth 1121 m (SMNH 55517).
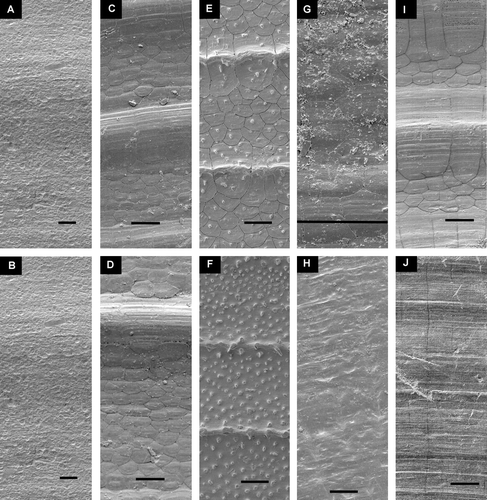
Figure 9. Umbonal area of smooth or radially sculptured Pectinoidea. Left valve above. All at same magnification. Scale line (see Figure E) 250 µm. (A,B) Hyalopecten frigidus, BIOICE#3214, NE of Iceland, depth 3000–3003 m (SMNH 59237). (C,D) Cyclopecten hoskynsi, BIOICE#2268, off southwestern Iceland, depth 450 m (SMNH 55534). (E,F) Similipecten oskarssoni, BIOICE#2401, off southwestern Iceland, depth 520 m (SMNH 55525). (G,H) Similipecten greenlandicus, BIOICE#2047, off northeastern Iceland, depth 272 m (SMNH 55520). (I,J) Similipecten similis, BIOICE#2393, off southwestern Iceland, depth 291 m (SMNH 55515).
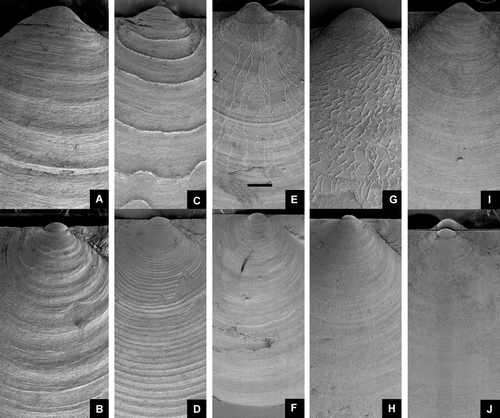
Figure 10. Detail of sculpture, mid-shell of half-grown specimens, A–D, Left valve above, E–H, right valve above. Scale lines 25 µm. (A,B) Similipecten greenlandicus, BIOICE#2096, off northern Iceland, depth 300 m (SMNH 56408). (C,D) Similipecten similis, BIOICE#2393, off southwestern Iceland, depth 291 m (SMNH 55515). (E,F) Similipecten oskarssoni, BIOICE#2401, off southwestern Iceland, depth 520 m (SMNH 55525). (G,H) Cyclopecten hoskynsi, BIOICE#2268, off southwestern Iceland, depth 450 m (SMNH 55534).
Type material
Lectotype of Pecten groenlandicus var. minor (spm: H 6.2, L 6.5, C 1.6 mm), 5 paralectotypes (3 spm + 2 v) in MNHN [no register number], here designated.
Type locality
Bay of Biscay, 647–1190 m depth.
Etymology
Named after Ingimar Óskarsson (1892–1981), honorary doctor at the Icelandic University, and an initiator of high level amateur natural history in Iceland.
Distribution
Western and south-western Iceland to the Bay of Biscay, probably also off West Africa but the paralectotypes supposed to originate from such localities, have unclear locality data or are lost. BIOICE material alive at 33 stations in 291–1408 m, on fine sediments (Map 5).
Description
Shell (Figures B–F) up to ca. 10 mm high, fragile, transparent, weakly convex, equivalve, nearly circular (somewhat longer than high), equivalve and equilateral, anterior auricles slightly longer than posterior ones and somewhat more demarcated from disc, umbonal angle ca. 120, internal ribs lacking. Prodissoconch (Figure B), length 175 µm, colour white.
Both valves smooth (Figures E,F, E,F), except the umbonal area of the left valve, which has a sculpture similar to S. greenlandicus, but more delicate (Figure E). Posterior auricle more continuous with disc than anterior one. Anterior auricle of right valve larger and more curved laterally than anterior one of left valve. Byssal notch moderately small, byssal fasciole very weak, ctenolium lacking.
Remarks
The type material of Pecten groenlandicus var. minor consists of two lots, one labelled ‘Talisman 1883, dragage 34’, from 112 m depth off northwestern Morocco. A second lot consists of 3 spm and 4 v from ‘Travailleur 1880, dragages 2, 7, 90, 12, 14, 16 and 20’, from the Bay of Biscay, in 697–1190 m. A third lot with a single specimen and the same station numbers has obviously been separated later from this and glued to a piece of cardboard. Two lots listed by Locard (1898) are missing. The specimen from dragage 34 seems to be mislabelled, already by or before Locard, since the depth is much smaller than expected. We have therefore selected a specimen from the mixed lot as lectotype. A restriction of the type locality, more detailed than Bay of Biscay, does not seem necessary. All specimens in identifiable condition belong to S. oskarssoni and the characteristic, irregular lines can be seen on the umbo of the left valve.
Similipecten oskarssoni can be confused with S. greenlandicus of similar size. Both species occur in Iceland and live in similar habitats, but with no overlap in distribution (Map 5). Similipecten oskarssoni has only a few irregular radial lines on the umbo of the left valve, visible at 50× with a good stereomicroscope, whereas S. greenlandicus has a similar but much coarser micro-sculpture and S. similis is smooth (Figures G,H, I). For further differences see . Schein (1989:96) regarded this species as a ‘forme incolore’ of Similipecten similis.
Table I. Morphologic characters of Similipecten spp.
According to the IZCN (1999:50, art. 45.6.4) the name Pecten groenlandicus var. minor Locard, 1898 may be considered subspecific, but the subspecies name is preoccupied by Pecten maximus var. minor Locard, Citation1888. Therefore the epithet is replaced with Similipecten oskarssoni nom. nov. (ICZN, Citation1999:63, art. 60.3).
Similipecten similis (Laskey, Citation1811)
(Figures C, 6C, 7A,G–I, 9I,J, 10C,D)
Pecten similis Laskey (1811:387, pl. 8, fig. 8 [original description]); Jensen (Citation1912:29 [Icelandic distribution]); Madsen (Citation1949:36 [Icelandic distribution]).
Similipecten similis; Lucas (Citation1979:3, figs [synonymy]); Dijkstra & Goud (Citation2002:44, figs 33–38 [synonymy, description, distribution]); Dijkstra & Gofas (Citation2004:53, fig. 14A–E).
Type material
Not searched for by the present authors. Types of Laskey are supposed to be in the Hunterian Museum, Glasgow.
Type locality
Firth of Forth, Scotland.
Distribution
North-eastern Atlantic (70N–15N), bathymetric range usually 25–500 m, living on fine silt to sand. A long-lived and frequently cited record from 3.6 m (Forbes & Hanley Citation1849:293) represents an exception. In Scandinavia it occurs more shallow, from ca. 25–50 m (SMNH). Found at 76 BIOICE stations in 104–1000 m, usually more shallow than 300 m (Map 6).
Description
Shell (Figures C, A,G–I) up to ca. 7 mm high, weakly convex, right valve slightly more convex than left one, semi-opaque, slightly longer than high; posterior auricles larger and longer than anterior ones; umbonal angle ca. 120. Colour strongly variable whitish, mottled with red and yellowish brown. Prodissoconch (Figure C), length 145 µm, colour white.
Both valves smooth (Figures I,J, C,D), sometimes weakly sculptured with delicate growth lines. Anterior auricles weakly sculptured with closely spaced commarginal lirae or lamellae; posterior auricles nearly smooth. Hinge line straight, except near the antero-dorsal margin where it is somewhat curved. No internal riblets, except two small auricular ones (sometimes nearly lacking). Byssal notch small, byssal fasciole very narrow or nearly absent, ctenolium lacking.
Remarks
Juvenile specimens of Similipecten oskarssoni could be confused with S. similis, but the sculpture of the umbo of the left valve in oskarssoni is visible already at a size of 2 mm. For further differences see .
Genus Catillopecten Iredale, 1939
Catillopecten eucymatus (Dall, 1898)
(Figures 1C, D, 2G, 4C,D, 8A,B)
Pecten fragilis Verrill (1897:81 [not Jeffreys 1876]).
Pecten eucymatus Dall (1898:754 [based on Pecten fragilis Verrill 1897, not Jeffreys, 1876]).
Bathypecten eucymatus; Schein (1989:81, pl. 4, figs 1–8, pl. 5, figs 1, 3, 5 [synonymy, description, distribution]).
Catillopecten eucymatus; Dijkstra & Gofas (Citation2004:55, fig. 15A–F [references, description, distribution]).
Type material
Syntypes, USNM 35566.
Type locality
Western Atlantic, North Carolina, off Cape Hatteras, 1542 m depth.
Distribution
Northern and temperate western and eastern Atlantic, from off southern Iceland to Cape Hatteras (North Carolina, USA), the Azores, and southwest of Ireland into the Bay of Biscay, living in bathyal to abyssal depths on soft sediments. Depth range 1057–4829 m (Verrill 1897:81; Schein 1989:81). Found at 5 BIOICE stations in 1295–2007 m (Map 7).
Catillopecten eucymatus is new to the Icelandic fauna.
Description
Shell (Figures C,D), up to ca. 10 mm high, fragile, translucent, unusually flat, left valve slightly more convex than right valve, circular, almost equilateral, auricles nearly equal in size, anterior auricles somewhat demarcated from disc and posterior auricles continuous with disc, umbonal angle ca. 110, colour whitish, internal ribs lacking. Prodissoconch (Figure G), length 175–180 µm, colour white.
Both valves (Figures C,D, A,B) smooth with no trace of macro- and micro-sculpture; commarginally undulated, especially central part of left valve; right valve less so. Delicate irregularly spaced, commarginal lamellae on auricles of left valve, somewhat more prominent on anterior one. Commarginal growth lines and very fine commarginal lamellae near ventral margin of right valve. Anterior auricle of right valve with minute commarginal striae, posterior ones smooth. Hinge line straight. Byssal notch moderately deep, byssal fasciole and ctenolium lacking.
Remarks
Dall (1898:754) considered the specimens which Verrill (1897) had identified as Hyalopecten fragilis to belong to a distinct species and named them Pecten eucymatus.
Catillopecten eucymatus and Similipecten oskarssoni are both fragile, strongly flattened, and circular in circumference. Catillopecten eucymatus differs, however, by having commarginal undulations (lacking in S. oskarssoni) and a smaller and more oblong anterior auricle in the right valve (broader and somewhat curved in S. oskarssoni).
Cragg (Citation2006) reported a presence of prodissoconch 2 based on Schein's (1989) plate 5, figure , although Schein did not consider the narrow zone surrounding the prodissoconch in the figured specimen a true prodissoconch 2. We consider this narrow zone the normal feature of a lecithotrophic larva mature for metamorphosis.
For a discussion on the generic names Bathypecten and Catillopecten see Dijkstra and Gofas (Citation2004) and Dijkstra and Marshall (Citation2008).
Family Pectinidae Rafinesque, 1815
Genus Hyalopecten Verrill, 1897
Hyalopecten frigidus (Jensen, Citation1904)
(Figures 1K,L, 2A, 5F,G, 9A,B)
Pecten frigidus Jensen (Citation1904:305–07, fig. [original description]); Jensen (Citation1912:33, pl. 1. figs 7a–f [description, distribution]).
Hyalopecten frigidus; Bouchet & Warén (Citation1979:216, figs 31 and 32 [distribution]); Richling (Citation2000:47, fig. 80 [references]).
Type material
Lectotype (here designated), the specimen figured by Jensen (Citation1904:figs; 1912:pl. 1, figs 7a,b,e), ZMC BIV-442. Approximately 300 paralectotypes (‘Ingolf’ stations 102–105, 110–113, 117–120, 125).
Type locality
Northeast of Iceland, 1847 m depth.
Distribution
North of the Greenland–Scotland Ridge in the lower bathyal and abyssal parts, sometimes attached to shells and other objects (Jensen Citation1912:33; Bouchet & Warén Citation1979:216). Found alive in 23 samples NE of Iceland, at 1456–3006 m depth, NE off Iceland (Map 2).
Description
Shell (Figures 1K,L, 5F,G) up to ca. 30 mm high, fragile, opaque, drawn out anteriorly, weakly inflated, higher than long, left valve slightly more convex than right valve, anterior margin more circular than posterior one, anterior auricles slightly larger and longer than posterior ones in young specimens, almost equal in adult specimens, colour whitish. Prodissoconch (Figure A), length 220 µm, colour white.
Both valves regularly, commarginally undulated and sculptured with numerous fine, close set radial lirae (Figures A,B), somewhat coarser on the left valve. Auricles small, anterior one set off, posterior one continuous with the disc. Hinge line straight. Byssal notch deep, byssal fasciole broad, ctenolium weak.
Remarks
This species closely resembles Hyalopecten pudicus (Smith, Citation1885) from south of the Greenland–Scotland Ridge. Hyalopecten frigidus is flatter, has somewhat broader undulations and the radial ribs are weaker, almost absent on the right valve. Hyalopecten pudicus also has a reticulate sculpture on the auricles, which have radial striae in H. frigidus.
A single fragment of Hyalopecten pudicus was recovered by the INGOLF Expedition from a depth of 2050 m, west of Cape Farewell (Greenland, 61° 44'N 30° 29'W; Jensen Citation1912:38). It can be expected also closer to Iceland and we refer to Dijkstra and Gofas (Citation2004) for description and figures.
Genus Delectopecten Stewart, 1930
Delectopecten vitreus (Gmelin, Citation1791)
(Figures 2J, 4A,B, 5A,D,E, 8G,H)
Ostrea vitrea Gmelin (1791:3328 [based on Chemnitz 1784:pl. 67, fig. 637a] [original description]).
Pecten vitreus var. laevigata Lovén (1846:31 [not Goldfuss 1833]). Syntypes, SMNH 4217, 8 v. Type locality Scandinavia.
Pecten vitreus var. abyssorum Lovén in Jeffreys (Citation1869:169).Type material not seen (supposed to be in USNM). Type locality: Not given by Jeffreys.
Pecten abyssorum Lovén in Sars (Citation1878:22, pl. 2, figs 6a–c [figs and distribution]).
Pseudamussium gelatinosum Mabille & Rochebrune (Citation1889:H–126). Type material not searched for (supposed to be in MNHN). Type locality: ‘Baie Orange’ [Orange Bay, Beagle Channel, Tierra del Fuego].
Pecten chaperi Dautzenberg & H. Fischer (1897:190, pl. 5, figs 5–8). Syntypes, MOM 28 148–9. Type locality: Azores, 845–1600 m depth.
Cyclopecten (Delectopecten) vitreus; Grau (Citation1959:47, pl. 18, fig. [synonymy]).
Delectopecten vitreus; Abbott (1974:446); Lucas (Citation1979:9, 10, 18, figs); Wagner (1991:13, text fig. 5 [synonymy, references]); Dijkstra & Kilburn (Citation2001:270, figs 3 and 4); Dijkstra & Goud (Citation2002:48, figs 42–44 [description, distribution]); Dijkstra & Gofas (Citation2004:59, fig. 18A–I [synonymy, references, distribution]).
Delectopecten vitreus var. abyssorum (Lovén); Schein (1989:75, pl. 2, figs 1–6).
Type material
Syntypes, ZMC BIV 54-55.
Type locality
‘Oceano septentrionali’ [North Atlantic Ocean].
Distribution
Atlantic Ocean, from north of Norway, 72°10'N (SMNH 56311) around Iceland, and the Labrador Sea, to the Antarctic; in Scandinavia more shallow, ca. 50–1000 m (SMNH), further south deeper, from 322 m (east of Gibraltar) to 4255 m (near the Azores, Schein 1989). The only littoral–sublittoral record from Greenland, ‘off Sukkertoppen’ (Posselt & Jensen 1898) is based on a misidentification of Palliolum striatum (SMNH 56310). Off the Icelandic northern coast Delectopecten vitreus is not found deeper than ca. 200 m, while in the south it occurred down to 1952 m. Delectopecten vitreus seems to prefer current-swept bottoms and is often attached to dead skeletons of gorgonians and scleractinia. Delectopecten vitreus is often inhabited by the large parasitic foraminiferid Hyrrokin sarcophaga Cedhagen, 1994, which penetrates the shell and sends pseudopodia into the host tissues. Found at 59 BIOICE stations in 165–1952 m (Map 8), mainly S and W of Iceland.
Description
Shell (Figures A,D,E) up to ca. 20 mm high, transparent to opaque, circular to somewhat oval in shape, valves nearly equiconvex, anterior auricles slightly larger than posterior ones, nearly equal in length, umbonal angle 110, colour whitish. Prodissoconch 1 (Figure J), length 100 µm, prodissoconch 2, length 315 µm, colour white or brown.
Both valves (Figures A,B, 8G,H) may be smooth (var. abyssorum) or, more commonly, covered with antimarginal micro-sculpture (Palliolum-like) and commarginal rows of small vesicles. When commarginal scales are damaged, minute commarginal ridges are formed by their bases. Posterior auricles continuous with disc and similarly sculptured as disc. Anterior auricle of right valve demarcated from disc and sculptured with 3–5 radial riblets. Hinge line straight. Byssal notch well-developed, byssal fasciole rather broad. Active ctenolium well-developed.
Remarks
Delectopecten vitreus is variable in convexity and in sculpture, sometimes nearly smooth; sometimes with prominent, concentrically arranged scales. The specimens from Iceland are usually smooth and transparent, occasionally with a radial micro-sculpture.
Pecten vitreus var. abyssorum and Chlamys chaperi are based on sculptural variations for which we have failed to see any geographic pattern and we believe it to be phenotypic variations. Pseudamussium gelatinosum Mabille & Rochebrune, 1889 (from off southern Argentina) was synonymized by Grau (Citation1959:48) and we cannot question this from morphological evidence. HD has also examined indistinguishable specimens from off Namibia. Such a wide distribution is, however, exceptional and suggests that more than one species may be involved, especially since the distribution seems not to be continuous.
Genus Palliolum Monterosato, 1884
Palliolum striatum (Müller, Citation1776)
(Figures 11C,D, 13G, 14E,F, 15E,F, 16E,F)
Pecten striatus Müller (1776:248 [original description]); Müller (Citation1788:26, pl. 60, figs 3–5 [subsequent description]). Pecten striatus Müller; Jensen (Citation1912:24 [Icelandic distribution]); Madsen (Citation1949:34 [Icelandic distribution]).
Palliolum striatum (Müller); Abbott (1974:446, fig. 5165); Wagner (1991:9, text fig. 3, pl. 2, figs 2, 4 [synonymy, references].
Figure 11. Young specimens of Pectinidae. Scale lines A, C, E, 50 µm, others 500 µm. (A,B) Palliolum incomparabile, Sweden, Bohuslän, Koster area, depth 35 m (SMNH 55523). (C,D) Palliolum striatum, western Norway, Korsfjord, depth 300–200 m (SMNH 19999). (E,F) Palliolum tigerinum, western Norway, Raunefjord, depth 65–70 m (SMNH 20064). (G) Aequipecten opercularis, western Norway, Raunefjord, depth 12–7 m (SMNH 20511). (H) Aequipecten opercularis, Italy, Sicily, Brucoli, depth 20 m (SMNH 56412). (I) Chlamys islandica, BIOICE#2537, off northwestern Iceland, depth 44 m (SMNH 55503).

Type material
Probably lost (not in ZMC, T. Schiøtte, personal communication).
Type locality
Although Müller (1776) did not indicate a locality for this species, it is most likely that the type material was collected from the Oslofjord (Norway), at that time belonging to Denmark (see Soot-Ryen Citation1963:111).
Distribution
North-western Atlantic from Newfoundland southwards to Massachusetts (USA), and north-eastern Atlantic from Hammerfest in Norway at 70.6°N, southwards into the Mediterranean Sea. A single record from south-western Greenland (Sukkertoppen, 27–36 m, 23 Sept 1870, SMNH 56310) earlier identified as Delectopecten vitreus (Posselt & Jensen 1898, see D. vitreus) must be considered uncertain although the label seems to be in order. All around Iceland, except the eastern and south-eastern coasts. Living in 25–400 m (HL), on coarse sediments, gravel, and shells, byssally attached to hard substrates. Found during BIOICE at a single station in 139–141 m (Map 4).
Description
Shell (Figures 15E,F,16E,F) up to ca. 25 mm high (usually smaller), thin, flattened, circular to more oval in shape, nearly equiconvex, slightly higher than long, anterior auricles larger and longer than posterior ones, umbonal angle ca. 95, colour strongly variable with blotches or streaks. Prodissoconch 1, length 90 µm, prodissoconch 2, length 215 µm (Figure G), colour brown.
Left valve (Figures C,D) covered with a coarsely granular antimarginal micro-sculpture and numerous, regularly spaced, scaly riblets (up to ca. 70). Anterior (large) and posterior (small) auricles indistinctly demarcated from disc, with similar micro-sculpture and 4–7 radial riblets.
Right valve with a more delicate antimarginal micro-sculpture and 3–5 anterior and posterior radial riblets. Auricles of similar sculpture, anterior ones with 7–10, posterior ones with 3–5 radial riblets. Hinge line nearly straight, somewhat raised laterally. Byssal notch deep, byssal fasciole broad. Active ctenolium well-developed with 5–7 teeth along suture.
Remarks
Juveniles (< 4 mm) (Figures E,F) may easily be confused with juveniles of Palliolum incomparabile (Risso 1826). However, P. striatum has small, radially arranged scales on the left valve, whereas P. incomparabile (Figures C,D) has a more uniform and delicate antimarginal sculpture on both valves. The small scales appear at a size around 1.5 mm, but at this size, only their bases are developed. The right valves of the two species can hardly be distinguished, even in adult specimens. Also P. tigerinum may be very similar, but its radial micro-sculpture is usually slightly stronger, also before the radial ribs appear.
Icelandic specimens agree well with topotype material (ZMO, SMNH) from the Oslofjord. To find only a single specimen in the BIOICE material is surprising since the species is not rare in private collections; perhaps it has a more shallow distribution than the BIOICE dredgings.
Palliolum incomparabile (Risso, Citation1826) (= Palliolum furtivum (Lovén, Citation1846)) is not known from Iceland or the Faroes, but occurs from ca. 69° N in Norway (Brattegard & Holthe Citation1997) and southwards along the European coast in 20–300 m depth (Dijkstra & Goud Citation2002, SMNH). It is figured (Figures 11A,B, 13I, 14C,D, 15C,D, 16C,D) for comparison.
Palliolum tigerinum (Müller, 1776)
(Figures 11E,F, 13H, 14A,B, 15A,B, 16A,B)
Pecten tigerinus Müller (1776:248 [original description]); Müller (1788:26, pl. 60, figs 6–8 [subsequent description]).
Pecten triradiatus Müller (1776:248 [original description]); Müller (1788:25, pl. 60, figs 1, 2 [subsequent description]); Soot-Ryen (Citation1963:111, pl. 13, upper and lower figs). Type material: Probably lost (not in ZMC, T. Schiøtte, personal communication). Type locality: Oslofjord (see also P. striatus).
Pecten tigerinus; Jensen (Citation1912:22 [Icelandic distribution]); Madsen (Citation1949:34 [Icelandic distribution]); Oskarsson (Citation1982:51, fig. 32 [Icelandic distribution]).
Palliolum tigerinus; Wagner (1991:10, text fig. 4, pl. 2, fig. 5 [references, synonymy, description, distribution]).
Type material
Probably lost (not in ZMC, T. Schiøtte, personal communication).
Type locality
See Palliolum striatum.
Distribution
Eastern Atlantic, from northern, western, southern and south-eastern Iceland and all the Norwegian coast (absent in the White Sea) southwards into the Mediterranean and to the Cape Verde Islands. It normally occurs in 20–400 m depth on muddy, sandy or rocky bottoms (SMNH). Found during BIOICE at 16 stations in 64–356 m, but the general distribution is more shallow than these finds indicate (HL).
Description
Shell (Figures 15A,B, 16A,B) up to ca. 30 mm high, rather sturdy, higher than long, equivalve and equilateral, anterior auricles much larger and longer than posterior ones, left valve somewhat more convex than right one, umbonal angle ca. 85. Colour usually based on purplish brown, often mottled with red, white, and yellow, but also pure yellow, white or red specimens occur. Prodissoconch 1, length 100 µm, prodissoconch 2, length 265 µm (Figure H), colour brown.
Both valves (Figures E,F) sculptured with antimarginal micro-sculpture and radial macro-sculpture of up to ca. 80, sometimes weak or even lacking riblets. Auricles with radial ribs (anteriorly 4–5, posteriorly 5–8). Byssal notch shallow, byssal fasciole narrow. Active ctenolium well-developed.
Remarks
Icelandic specimens are similar to Norwegian ones (SMNH, ZMC), where radially sculptured specimens dominate, including the more unusual variant triradiatus (with three prominent radial ridges), contrasting specimens from the Irish Sea (HD, ZMA), which are smooth. Mature shells are sometimes swollen, with a distinct increase in convexity close to the margin. All these variations have been observed from Iceland.
Palliolum tigerinum may be difficult to separate, especially when young (Figures A,B), from P. striatum (Figures C,D) and P. incomparabile (Figures E,F). Helpful features are that the anterior auricle has a much stronger cancellate sculpture (in addition to the regular micro-striae covering the shell) and the posterior ear is disproportionally short. The height of the shell more distinctly exceeds its length, than in striatum and incomparabile.
Genus Pseudamussium Mörch, 1853
Pseudamussium peslutrae (Linnaeus, Citation1771)
(Figures 12H, 13B, 17C,D, 18E,F)
Ostrea peslutrae Linnaeus (1771:547); Dijkstra (Citation1999:427, figs 7E,F [lectotype]).
Pecten septemradiatus Müller, 1776; Jensen (Citation1912:20, pl. 1, figs 6a–c [var. scaber] [distribution]); Madsen (Citation1949:33 [distribution]).
Pseudamussium septemradiatum; Wagner (1991:14, pl. 3 fig. 1, text-fig. 6 [synonymy, references]).
Pseudamussium peslutrae; Dijkstra & Goud (Citation2002:53, fig. 50 [description, distribution]).
Figure 12. Young specimens of Pectinidae. Scale lines 0.5 mm. (A) Talochlamys pusio, Strait of Gibraltar, Ceuta, depth 20–40 m, (SMNH 59345). (B) Talochlamys pusio, Acores, Sao Miguel, depth 15–22 m (SMNH 59346). (C) Talochlamys pusio, western Norway, Korsfjord, 276–256 m (SMNH 55505). (D) Talochlamys multistriata, Strait of Gibraltar, Ceuta, depth 9–10 m (SMNH 56215). (E) Talochlamys multistriata, Strait of Gibraltar, Ceuta, depth 16–25 m (SMNH 57226). (F) Mimachlamys varia, Sweden, Bohuslän, Gullmarsfjord (SMNH 54319). (G) Pseudamussium sulcatum, western Norway, Korsfjord, depth 260–250 m (SMNH 20130). (H) Pseudamussium peslutrae, BIOICE#2393, off southwestern Iceland, depth 291 m (SMNH 55502). (I) Mimachlamys varia, Strait of Gibraltar, Ceuta, depth 16–25 m (SMNH 20577).

Figure 13. Larval right valves. Scale lines 50 µm. Arrows indicate demarcation of prodissoconch 1. The scale line is valid for length only; the height may differ slightly because of tilt. (A) Pseudamussium sulcatum, western Norway, Korsfjord, depth 260–250 m (SMNH 20130). (B) Pseudamussium peslutrae, BIOICE#2393, off southwestern Iceland (SMNH 55502). (C) Chlamys islandica, 55503, BIOICE#2537, off northwestern Iceland (SMNH 55503). (D) Aequipecten opercularis, Italy, Sicily, Brucoli, depth 20 m (SMNH 56412). (E) Mimachlamys varia, Sweden, Bohuslän, Koster area, depth 1–2 m (SMNH 20569). Prodissoconch 1 too corroded to indicate. (F) Talochlamys pusio, western Norway, Korsfjord, depth 276–256 m (SMNH 55505). (G) Palliolum striatum, western Norway, Korsfjord, depth 300–200 m (SMNH 19999). (H) Palliolum tigerinum, western Norway, Raunefjord, depth 65–70 m (SMNH 20064). (I) Palliolum incomparabile, Sweden, Bohuslän, Koster area, depth 35 m (SMNH 55523).
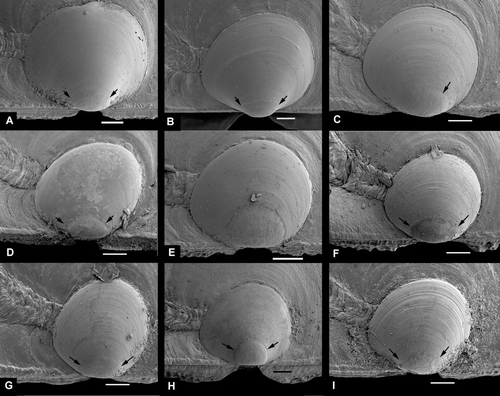
Figure 14. Palliolum spp., young specimens. (A,B) P. tigerinum, Ø 4.0 mm, western Norway, Raunefjord, depth 65–70 m (SMNH 20064). (C,D) P. incomparabile, Ø 5.0 mm, Sweden, Bohuslän, Gullmarsfjord (SMNH 55958). (E,F) P. striatum, Ø 5.2 mm, western Norway, Korsfjord, depth 300–200 m (SMNH 19999).
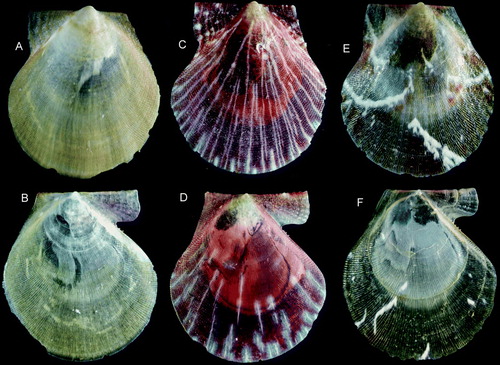
Figure 15. Palliolum spp., adult specimens. (A,B) P. tigerinum, Ø 18.9 mm, off southeastern Iceland, 63° 54’ N, 15° 11.5’ W, 134 m (IMNH 8670, SMNH 55496). (C,D) P. incomparabile, Ø 14.6 and 13.4 mm, Sweden, Bohuslän, Koster area (SMNH 55497). (E,F) P. striatum, Ø 12.3 mm, BIOICE#2388, off southwestern Iceland, depth 171 m (SMNH 55508).

Figure 16. Palliolum spp., adult specimens. (A,B) P. tigerinum, Ø 12.3 mm, BIOICE#2710, off western Iceland, depth 220 m (SMNH 55507). (C,D) P. incomparabile, Ø 14.6 and 13.4 mm, Sweden, Bohuslän, Koster area (SMNH 55497). (E,F) P. striatum, Ø 18.2 mm, Sweden, Bohuslän, Koster area (SMNH 55494).
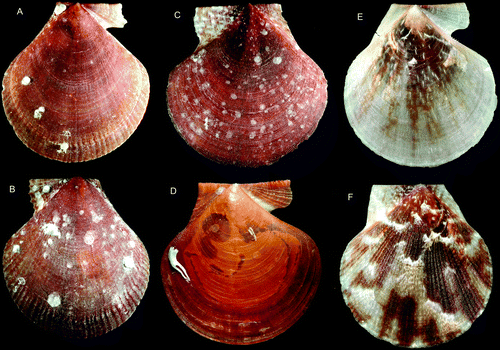
Figure 17. Pseudamussium spp., young specimens. (A,B) P. sulcatum, Ø 4.0 mm, western Norway, Korsfjord, depth 260–250 m (SMNH 20130). (C,D) P. peslutrae, Ø 3.8 mm, BIOICE#2393, off southwestern Iceland, depth 291 m (SMNH 55502).

Figure 18. Pseudamussium spp. (A,B) P. sulcatum, Ø 25 mm, BIOICE#2305, off southwestern Iceland, depth 406 m (SMNH 55524). (C,D) P. sulcatum, Ø 16 mm, western Norway, Hjeltefjord, depth 100–180 m (SMNH 20143). (E,F) P. peslutrae, Ø 18.8 mm, Sweden, Bohuslän, Koster area (SMNH 55495).

Type material
Lectotype, LSL, designated by Dijkstra (Citation1999:427).
Type locality
Off Largs, Ayrshire, Scotland (Dijkstra Citation1999).
Distribution
The eastern Atlantic from western, southern and south-eastern Iceland and the whole Norwegian coast (absent in the White Sea) to Morocco, and into the western Mediterranean Sea, living sublittorally to bathyally, on fine sediments. Depth range usually 50–500 m, occasionally recorded much deeper (mostly empty shells). BIOICE material alive from 35 stations in 146–993 m (Map 3).
Description
Shell (Figures E,F) up to ca. 65 mm high, rather thin, right valve slightly more convex than left one, almost circular, auricles nearly equal in size, umbonal angle ca. 95. Colour reddish-brownish, mottled with darker or lighter shades; right (lower) valve paler, interior glossy white. Prodissoconch 1, length 100 µm, prodissoconch 2, length 320 µm (Figure B), colour brown.
Both valves (Figure E,F) with 5–10 equidistant radial plicae, on right valve broader than on left one; secondary radial scaly riblets (sometimes lacking), and microscopic radial striae cover all surface; microsculpture of incremental lamellae in early growth stage. Auricles short, anterior ones slightly longer than posterior ones, with radial ridges and dorsal scales; antimarginal microsculpture weak or lacking. Hinge line straight. Byssal notch and active ctenolium weakly developed.
Remarks
Pseudamussium peslutrae is rather variable in form, macro- and micro-sculpture. Specimens from the boreal region are larger (up to ca. 65 mm high) and more sharply sculptured than those from further south, which have an evenly undulated shell, rather than groups of ribs forming a ‘mega-sculpture’. Young specimens (Figures 12H, 17A,B) closely resemble P. sulcatum (Figures 12G, 17C,D), which is more coarsely sculptured, but safe identification needs access to a good growth series.
Pseudamussium sulcatum (Müller, 1776)
(Figures 12G, 13A, 17A,B, 18A–D)
Pecten sulcatus Müller (1776:248 [original description]).
Ostrea arata Gmelin (1791:3327). Based on Müller's (1776) description and takes the same type locality and type material.
Pecten bruei Payraudeau (Citation1826:78, figs 10–14). Type material: Syntypes, MNHN [no register number]. Type locality: ‘Les golfes de Sagone, d'Ajaccio, de Valinco, de Porto-Vecchio’ (Corsica).
Pecten idaeus CitationReeve (1853:species 153 [unpaginated], pl.33, fig. 153). Type material: Holotype, BMNH 19990407. Type locality: Unknown.
Pecten aratus; Jensen (Citation1912:19, pl. 1, figs 5a–e [Icelandic distribution]); Madsen (Citation1949:32 [distribution]).
Karnekampia sulcata; Wagner (Citation1988:41, 43 fig. 2); Wagner (1991:19, pl. 3, fig. 4, text-fig. 9 [synonymy, references]); Waller (Citation2006:17).
Pseudamussium sulcatum; Waller (Citation1991:18, 35); Dijkstra & Goud (Citation2002:57, fig. 54 [description, distribution]); Dijkstra & Gofas (Citation2004:62, fig. 19A–D [synonymy, distribution]
Type material
Pecten sulcatus: probably lost, not traced in ZMC (T. Schiøtte, personal communication).
Type locality
See Palliolum striatum.
Distribution
The eastern Atlantic, all around Iceland (except the north-eastern coast, HL), the Norwegian coast from Andfjorden (69°10'N; SMNH), southwards to the Madeira Archipelago, and into the Mediterranean Sea. It lives in sublittoral to upper bathyal depths on current-swept bottoms, byssally attached to rocks or amongst gravel and/or rubble. It is one of the most common bivalves on North-east Atlantic Lophelia reefs, where it lives attached to coral debris. BIOICE material alive at 31 stations in 98–922 m (Map 4).
Description
Shell (Figures A–D) up to ca. 30 mm high, weakly convex, left valve somewhat more convex than right one, slightly higher than long, anterior auricles much larger and longer than posterior ones, umbonal angle ca. 90, colour variable, usually uniform deep red, orange, yellow or rarely white, occasionally with commarginal stripes. Prodissoconch 1, length 110 µm, prodissoconch 2, length 310 µm (Figure A), colour white.
Left valve (Figures A,C) sculptured with 15–40, irregularly spaced, primary ribs, often hollow sometimes more solid, with tubercles, spines or lamellae. Intercalated secondary radial riblets commence on central part of disc. Primary and secondary ribs sometimes of almost equal size and spacing.
Right valve (Figures B,D) with clustered or equally spaced ribs. Antimarginal micro-sculpture present laterally and on auricles, absent from anterior auricle of right valve. Auricles with 3–6 radial riblets and commarginal riblets in early growth stage on anterior auricle of left valve. Hinge line straight. Byssal notch moderately deep, byssal fasciole broad. Active ctenolium well-developed with 4–6 teeth.
Remarks
Jensen (Citation1912) and Madsen (Citation1949) recorded Pseudamussium sulcatum from Iceland under the absolute synonymous name Pecten aratus.
Pecten bruei was based on Mediterranean specimens with more developed groups of ribs. This form intergrades with Atlantic sulcata, and we can see no possibility to separate them morphologically (see Dijkstra & Gofas Citation2004). For separation from P. peslutrae, see that species.
Genus Chlamys Röding, 1798
Chlamys islandica (Müller, 1776)
(Figures 11I, 13C, 19A,B, 20D,E)
Pecten islandicus Müller (1776:248 [original description]).
Chlamys islandica; Abbott (1974:444, pl. 19, fig. 5147); Wagner (Citation1991:25, text fig. 13, pl. 4, figs 1–3 [synonymy, references, description]).
Figure 19. Young pectinids. (A,B) Chlamys islandica, Ø 3.6 mm, BIOICE#2537, off northwestern Iceland, depth 44 m (SMNH 55503). (C,D) Mimachlamys varia, Ø 4.5 mm, Sweden, Bohuslän, Koster area (SMNH 20571). (E,F) Talochlamys pusio, Ø 3.4 mm, western Norway, Korsfjord, northwest of Marsteinen, depth 276–256 m (SMNH 55505).

Figure 20. Pectinidae spp. (A) Talochlamys pusio, Ø 23.3 mm, western Norway, 39–41 m (SMNH 55498). (B,C) Talochlamys pusio, Ø 29.1 mm, western Norway, Bergen (SMNH 55500). (D,E) Chlamys islandica, Ø 14.7 mm, BIOICE#2941, off northwestern Iceland, depth 156 m (SMNH 55492). (F,G) Mimachlamys varia, Ø 7.0 mm, Sweden, Bohuslän, Gullmarsfjord (SMNH 55943). (H,I) Mimachlamys varia, Ø 16.0 mm, Norway, Raunefjord, intertidal (SMNH 20567). (J,K) Aequipecten opercularis, Ø 15.2 mm, Sweden, Bohuslän, Koster area (SMNH 55493).

Type material
Probably lost, not in ZMC (T. Schiøtte, personal communication).
Type locality
Iceland.
Distribution
Circumpolar except Beaufort Sea, Chukchi Sea and the entrance of Bering Strait (not in the North Pacific). In the north-eastern Atlantic it reaches southwards to ca. 69°N in Norway, the Faeroes, all of Iceland and Greenland. In the western Atlantic it reaches Massachusetts. In Norway there are scattered more southern relict pockets, south to the Bergen area, a reminiscence of its more southern distribution during the glaciation. Chlamys islandica lives in littoral to uppermost bathyal depths, byssally attached to gravel, stones and rocks. All around Iceland, except the south-eastern coast. BIOICE material alive from 20 stations in 19–327 m (Map 9), but the large populations occur in 50–300 m depth off eastern and western Iceland (HL).
Description
Shell (Figures D,E) up to at least 140 mm high, but generally smaller than 90 mm, sturdy, elongate to nearly circular; weakly inflated, of almost the same convexity, anterior auricles much larger and longer than posterior ones, umbonal angle ca. 82–90. Colour highly variable, orange–red (common), pink, yellow, purple, brown or white (rare), sometimes with patterns of concentric coloured bands, often with 3 or 5 light radiating streaks. Prodissoconch 1, length 100 µm, prodissoconch 2, length 250 µm (Figure C), colour white.
Both valves sculptured with numerous irregularly spaced, small scaly radial riblets of variable prominence; usually 5 ribs are distinctly stronger. At 2 mm shell height ca. 20 ribs commence and extend to the ventral margin, increasing with intercostal riblets to ca. 140 in adult specimens. Right valve with 10–15 riblets less than left valve. A shagreen micro-sculpture is usually present at a size of 5–40 mm.
Anterior auricles with ca. 12 scaly radial riblets in left valve; the ribs are weaker and fewer (4–6) on the right valve. Posterior auricles with ca. 12 weak radial riblets on left valve, ca. 10 on right valve. Hinge line straight. Byssal notch moderately deep; byssal fasciole rather broad. Active ctenolium well-developed with 4–6 teeth.
Remarks
Chlamys islandica is a highly variable species. The shape varies from much higher than long to nearly circular, sometimes weakly undulated radially; the sculpture from delicate to coarse with single or clustered ribs; the shagreen microscuplture may occur all over the shell or is only developed in early growth stages. Maximum size in Iceland ca. 125 mm, but unconfirmed reports indicate a diameter exceeding 145 mm in Labrador (Nain).
Small specimens (Figures 11I, 19A,B) may resemble Mimachlamys varia (not present in Iceland), but that species (Figures 12F, 13E, 19C,D, 20F–I) has more numerous ribs when young. Large specimens of M. varia have fewer ribs and lack the shagreen sculpture present in C. islandica.
Mimachlamys varia is known in Norway from Lofoten (ca. 68°N) and southwards (Brattegard & Holthe Citation1997, SMNH).
Chlamys islandica is the subject of commercial fishery in Iceland and Norway, and has a delicious taste. In 1987 the Norwegian fishery peaked at 45,000–50,000 tons, but due to over-fishing it dropped abruptly. It is now strictly regulated and all fields are closed at present. The fishery in Iceland has dropped from 17,000 tons in 1984, to ca. 6000 tons in 2002. Rearing of larvae and artificial habitats are being explored in Norway.
Genus Talochlamys Iredale, 1929
Talochlamys pusio (Linnaeus, 1758)
(Figures 12A–C, 13F, 19E,F, 20A–C)
Ostrea pusio Linnaeus (1758:698, no. 169 [original description]); Dijkstra (Citation1999:417, figs 4c–f [synonymy]).
Hinnites (Hinnites) distortus; Wagner (1991:32, text-fig. 17 [synonymy, references]).
Crassadoma pusio; Waller (Citation1993:215, figs 5b,e,h,k,l [type data]).
Talochlamys pusio; Dijkstra & Goud (Citation2002:69, figs 73 and 74 [distribution]).
Type material
Lectotype in LSL, designated by Waller (Citation1993:215). See also Dijkstra (Citation1999:418–19).
Type locality
‘O. australiore’. Waller (Citation1993:215) designated a slightly more precise locality: ‘North-eastern Atlantic’.
Distribution
Northern to tropical eastern Atlantic from south-western Iceland and northern Norway (69.5°N, Høisæter Citation1986) southwards to the Ivory Coast (Waller Citation1993:216), also occurring in the western Mediterranean Sea and the Azores (Dijkstra & Goud Citation2002:69). It is very rare in the Skagerrak, common throughout the British Isles, the Channel Islands and Azores, rare in the western Mediterranean Sea and Canary Islands. This species lives in littoral and sublittoral depths; from a size of 10–20 mm it is cemented with the right valve to hard substrates. In Scandinavia frequent in empty shells of Arctica islandica. During BIOICE a single specimen was found alive in 31–33 m (Map 7), but the species has also been taken in shallow water at Reykjanes (SMNH) and washed ashore in other localities at the south-western coast (HL).
Description
Shell (Figures A–C) up to ca. 40 mm high, usually less than 30 mm, sturdily built. Before permanent attachment at a size of 10–20 mm, the shape is higher than long, inflated, right valve slightly more convex than left one. After attachment the right valve takes its shape after the substrate and may be higher than long or longer than high with a concave, flat or convex left (upper) valve. Anterior auricles larger and longer than posterior ones, umbonal angle ca. 82–90. Colour variable, whitish, creamy, yellow, orange, red or brown, colour pattern dotted, blotched, or with zigzag lines. Prodissoconch 1, length 85 µm, prodissoconch 2, length 230 µm (Figure F), colour white.
In the non-cemented stage (Figures 12A–C, 19E,F) both valves have numerous, irregularly spaced, squamous radial riblets of varying strength, commencing with ca. 10 at 5 mm shell height, gradually increasing in number to ca. 70 near the ventral margin in adult specimens. Anterior auricles larger than posterior ones, with 6–9 radial riblets on left valve, 5 on right valve and posterior auricles with 6–8 on left, 6 on right. Micro-sculpture granular on umbo, later radially striated between the ribs. Shagreen or reticulated micro-sculpture lacking. Byssal notch relatively deep, byssal fasciole moderately broad. Active ctenolium well-developed with 6–9 teeth.
Remarks
Free-living juveniles of Talochlamys pusio (Figures 12A–C) and T. multistriata (Poli, Citation1791) (Figures 12D,E), the latter never cemented, are hard to distinguish and often misidentified. Adult specimens can be differentiated by a secondary calcareous layer inside the umbo, present in T. pusio and lacking in T. multistriata. Moreover, T. pusio has a stronger micro-sculpture on the umbo of the left valve, than T. multistriata. These characters can be observed in the pre-cemented Chlamys-resembling stage. No intermediates have been found (Waller Citation1993). Small specimens may be imagined to closely resemble Mimachlamys varia, but that species has a much finer sculpture at the same size (Figures 12F,I, 19C,D) and its ribs are of uniform size, not alternating broader and more narrow ones, as in T. pusio. Mimachlamys varia is not known from Iceland or the Faeroes, but occurs in Norway, north to 64° N (Trondheimsfjord; Høisæter Citation1986, SMNH), so it is figured here for comparison (Figures H,I).
Genus Aequipecten Fischer, 1886
Aequipecten opercularis (Linnaeus, 1758)
(Figures 11G,H, 13D, 20J,K)
Ostrea opercularis Linnaeus (1758:698, no. 171); Dijkstra (Citation1999:421, figs 7A–D [lectotype]).
Aequipecten opercularis; Wagner (1991:40, pl. 6 figs 1–5, text fig. 22 [synonymy, references]); Dijkstra & Goud (Citation2002:70, fig. 76 [description, distribution]).
Type material
Lectotype in LSL, designated by Dijkstra (Citation1999:423).
Type locality
‘O. meridionali’, restricted to Brittany (France) by Dijkstra (Citation1999:422).
Distribution
Throughout the eastern Atlantic along the European coasts from south-eastern Iceland and Norway south of Lofoten (Høisæeter Citation1986; Senja at 69.5° N, valves in SMNH) to the Canary Islands and the Mediterranean Sea. As an adult it lives free, in colonies on sandy or silty sediments in the littoral to sublittoral zone. Juveniles are usually byssally attached. A single BIOICE specimen was taken alive in 134–141 m (Map 9), and the species was previously not known in Iceland.
Description
Shell (Figures J,K) usually up to ca. 60 mm high, occasionally 100 mm; weakly inflated, left valve slightly more convex than right valve, somewhat longer than high, equilateral. Periphery slightly drawn out posteriorly. Anterior auricles somewhat larger and longer than posterior ones. Umbonal angle ca. 105°. Colour extremely variable; uniformly coloured or with blotches. Prodissoconch 1, length 110 µm, prodissoconch 2, length 250 µm (Figure D), colour white.
Both valves with 18–25, regularly arranged, radial, scaly ribs, secondary radial, scaly riblets and commarginal lamellae on and between primary ribs, more prominent on left valve. Hinge line straight, byssal notch moderately deep and small, byssal fasciole rather narrow. Active ctenolium with 3–6 teeth.
Remarks
Petersen (Citation1968:12) commented that Aequipecten opercularis was not known from Iceland, but is common around the Faeroes, where it forms the basis for a strictly regulated fishery. Young specimens (Figures G,H) resemble those of Talochlamys pusio and Mimachlamys varia, but are proportionally lower and flatter and the size of the ribs is more uniform.
Discussion
About 450 species of Pectinoidea, 150 Propeamussiidae and 300 Pectinidae are known worldwide. About 80 species, 25 propeamussiids and 55 pectinids occur north of the Equator in the Atlantic. Pectinidae is probably one of the best known bivalve families, but still new species are being described regularly.
A total of 17 species, represented by ca. 8000 specimens, were collected at 356 stations around Iceland, an average of ca. 25 specimens per station or ca. 10 specimens based on the total number of stations. Their bathymetrical range is intertidal to more than four km depth. Two species seem more rare than they actually are, namely Palliolum striatum and Talochlamys pusio, because their normal depth range is more shallow than the working range of the BIOICE programme, mainly deeper than 100 m. Aequipecten opercularis, from the littoral to upper bathyal zone seems to be an occasional guest.
The poverty of the pectinoid fauna north of Iceland in bathyal depths is striking. Hyalopecten frigidus is the only species known below 1000 m depth. The Ingolf expedition took about 300 shells and specimens, from ca. 1050 to 2400 m, in a temperature of ca. –0.6 to –1°C (Jensen Citation1912). More than 300 specimens were then found living at 8 out of 16 stations in this depth range; at 4 only valves were found. During 1991 to 2004 it was found at 24 stations NE of Iceland, at 1456–3006 m water depth, and at a temperature range from –0.7 to –0.9°C. In these samples it is one of the most common bivalves.
North of Iceland and more shallow, Cyclopecten hoskynsi dominates in 300–1000 m. At ca. 600 m, Similipecten greenlandicus and S. similis are added. Cyclopecten hoskynsi and S. similis occur considerably deeper south of Iceland, down to 1400 m. It seems thus that low water temperatures constitute a limiting factor north of Iceland.
South of Iceland, a slightly impoverished North Atlantic bathyal fauna is encountered. Hyalopecten pudicus and Parvamussium fenestratum were not found, although they probably occur there.
We were a little surprised when examining the collections at SMNH for Scandinavian juveniles of certain species, but could hardly find a single small specimen of Mimachlamys varia, Aequipecten opercularis and Pecten maximus. The collections there include material collected by Lovén in 1840–1860 and he also searched for juvenile specimens. The same has been the intention of AW, and other, more rare species like Talochlamys pusio were well represented by small specimens in the collections (as are most other Scandinavian bivalves). In the Mediterranean and West European material, such juveniles were very common for all pectinids. If the observation on the rarity of juvenile Mimachlamys varia, Aequipecten and Pecten, is correct, a possible explanation may be that they are non-reproducing pseudo-populations and that they are maintained by an inflow of larvae from the south. Certainly this is the origin of the single Icelandic specimen of Aequipecten orpercularis which has a stable population in the Faroes.
As a consequence of its reflection in the larval shell, much has been speculated about the connections between types of larval development, dispersal and distribution among marine molluscs. Examination of larval shells is easier in pectinids than in most other bivalves since the animals live on the bottom, not in it, and therefore abrasion against sediment and corrosion by drops in pH are avoided, and the larval shells stay in good shape. summarizes the type of larval development and gives the size of the parts of the larval shell.
Table II. Distribution and larval shell data on Northeast Atlantic Pectinoidea. AA, Amphiatlantic; EA, East Atlantic incl. Iceland. Colour of larval shell indicated by w, white, b, brownish. In Delectopecten vitreus we have seen both brown and white larval shells. Figure for Pecten maximus based on Cragg (2006).
There may be a phylogenetic constraint on the type of larval development, since all propeamussiids investigated here have lost planktotrophy; that was also the case in their slightly more southern relatives discussed by Dijkstra & Gofas (Citation2004) and may be a trait of the family. Hyalopecten frigidus forms an exception among the Pectinidae and follows Thorson's (Citation1951) rule, that deep-sea animals predominantly have lecithotrophic development (not correct for gastropods: Rex & Warén Citation1982). All other pectinids have a shallower upper distributional limit and have planktotrophic development.
Furthermore, the relation between the size of prodissoconch 2 and 1 reflects the length of the planktotrophic life. In the species discussed here the ratio varies between 3.4 and 2.0. Cragg (Citation2006), table 2.1) presented such relations for 34 pectinoids, of which 6 were in the range 1.13–1.02, the others higher than 1.8. We believe the group of six unexpectedly low values to be based on erroneous original data.
The fact that only some of the North European pectinids with planktotrophic larvae occur in Iceland is not reflected in the size of the larval shell; it is of similar size as in those that do not reach Iceland. Three species of Palliolum occur in Scandinavia, all with similar size of prodissoconch 1 and 2; actually P. striatum, which occurs on both sides of the Atlantic, has the smallest larval shell. It should also be considered that a short or no dispersal phase may be favourable for maintaining small, isolated populations (Johannesson Citation1988). Such populations are, however, probably more vulnerable for temporary environmental changes in their habitat since they do not regularly communicate with more distant and perhaps unchanged places.
The zoogeographic connections of the Icelandic pectinoid fauna are clearly European, seven of the seventeen species occur in Europe but not in the western Atlantic; no species has only West Atlantic affinity. This fits with the current ideas about the zoogeography of the North Atlantic (see Gudmundsson Citation1998), for molluscs recognized already by Mörch (Citation1868) and confirmed by Thorson (Citation1941) and Madsen (Citation1949), although Dahlgren et al. (Citation2000) found more connection between Icelandic and American populations of the bivalve Arctica islandica than with European ones. Probably the long coastline with subarctic conditions, from Greenland to Massachusetts, forms a barrier for the more shallow species that cannot rely on a deeper continuous distribution along the Greenland–Scotland Ridge.
Editorial responsibility: Christoffer Schander
Supplementary Data
Download Zip (33.9 KB)Acknowledgements
For the loan of material and access to the mollusc collections we are much indebted to P. Bouchet (MNHN), J. Knudsen and T. Schiøtte (ZMC), and K.M. Way (BMNH). Hans Larsen, Reykjavik, is thanked for his management of the mollusc collections of the BIOICE programme, preliminary identifications, and labeling. He and Jón Bógason also gave Warén access to their unpublished notes on the Icelandic mollusc fauna. Dr Jon-Arne Sneli, Trondheim, helped with information on commercial pectinid fishery.
Notes
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
References
- Abbott , RT . 1974 . American Seashells , 2nd edn , New York : Van Nostrand Reinhold .
- Bernard , FR . 1968 . Cyclopecten carlottensis, a new species of Pectinidae from the northeastern Pacific . Journal of the Fisheries Research Board of Canada , 25 : 1509 – 10 .
- Bouchet , P and Warén , A . 1979 . The abyssal molluscan fauna of the Norwegian Sea and its relation to other faunas . Sarsia , 64 : 211 – 43 .
- Brattegard , T and Fosså , JH . 1991 . Replicability of an epibenthic sampler . Journal of the Marine Biological Association of the UK , 71 : 153 – 66 .
- Brattegard T , Holthe T 1997 . Distribution of marine, benthic macro-organisms in Norway . Research Report for DN 1997-1. Directorate for Nature Management, Norway .
- Clarke , AH . 1974 . Molluscs from Baffin Bay and the northern North Atlantic Ocean. National Museum of Natural Sciences . Publications in Biological Oceanography , 7 : i – xii . 1–23
- Cragg , SM . 2006 . “ Development, physiology, behavoiur and ecology of scallop larvae ” . In Scallops: Biology, Ecology and Aquaculture , Edited by: Shumway , SE and Parsons , GJ . 45 – 122 . Amsterdam : Elsevier .
- Dahlgren , TG , Weinberg , JR and Halanych , KM . 2000 . Phylogeography of the ocean quahog (Arctica islandica): Influences of paleoclimate on genetic diversity and species range . Marine Biology , 137 : 487 – 95 .
- Dall , WH . 1881 . Reports on the results of dredging […] ‘Blake’ […] XV. Preliminary report on the Mollusca . Bulletin of the Museum of Comparative Zoology , 9 ( 2 ) : 33 – 144 .
- Dall , WH . 1897 . New west American shells . Nautilus , 11 : 85 – 6 .
- Dall , WH . 1898 . Contributions to the Tertiary fauna of Florida […], Part 4. I. Prionodesmacea, II. Teleodesmacea . Transactions of the Wagner Free Institute of Science of Philadelphia , 3 : i – viii . 571–947
- Dautzenberg , P and Fischer , H . 1897 . Campagnes scientifiques de S.A. le Prince Albert Ier de Monaco, dragages effectués par l’ ‘Hirondelle’ et par la ‘Princesse-Alice’ (1888–1896) . Mémoires de la Société Zoologique de France , 10 : 139 – 234 .
- Dijkstra , HH . 1999 . Type specimens of Pectinidae (Mollusca: Bivalvia) described by Linnaeus (1758–1771) . Zoological Journal of the Linnean Society , 125 : 383 – 443 .
- Dijkstra , HH and Gofas , S . 2004 . Bathyal Pectinoidea (Bivalvia: Propeamussiidae, Pectinidae) from some northeastern Atlantic seamounts . Sarsia , 89 : 33 – 78 .
- Dijkstra , HH and Goud , J . 2002 . Pectinoidea (Bivalvia, Propeamussiidae & Pectinidae) collected during the Dutch CANCAP and MAURITANIA expeditions in the south-eastern region of the North Atlantic Ocean . Basteria , 66 : 31 – 82 .
- Dijkstra , HH and Kilburn , RN . 2001 . The family Pectinidae in South Africa and Mozambique (Mollusca: Bivalvia: Pectinoidea) . African Invertebrates , 42 : 263 – 321 .
- Dijkstra , HH and Marshall , BA . 2008 . The recent Pectinoidea of the New Zealand region (Mollusca: Bivalvia: Propeamussiidae, Pectinidae and Spondylidae) . Molluscan Research , 28 : 1 – 88 .
- Ekman , S . 1935 . Tiergeographie des Meeres , Leipzig : Akademische Verlagsgesellschaft .
- Forbes , E . 1844 . Report on the Mollusca and Radiata of the Aegean Sea, and on their distribution, considered as bearing on geology . Report of the British Association for the Advancement of Science , 1843 : 130 – 93 .
- Forbes , E and Hanley , S . 1849 . A History of British Mollusca , Vol. 2 , London : J. van Voorst . (1849–1850)
- Friele , H . 1873 . Oversigt over de i Bergens omegn forekommende skaldækte mollusker. Forhandlinger i Videnskabs-Serlskabet . Christiania , 1873 : 269 – 312 .
- Gmelin , JF . 1791 . Caroli Linnaei Systema Naturae per Regna Tria Naturae […] Editio Decima Tertia, Aucta, Reformata, Vermes Testacea. Vol.1, pt. 6. Lipsiae: Georg. Emanuel . Beer. , 1 : 3021 – 910 .
- Grau , G . 1959 . Pectinidae of the eastern Pacific. Alan Hancock Foundation . Pacific Expeditions , 23 : 1 – 308 .
- Gudmundsson , G . 1998 . Distributional limits for Pyrgo species at the biogeographic boundaries of the arctic and the North Atlantic Boreal regions . Journal of Foraminiferal Research , 28 : 240 – 56 .
- Hansen , B , Turrell , WR and Østerhus , S . 2001 . Decreasing overflow from the Nordic seas into the Atlantic Ocean through the Faroe Bank channel since 1950 . Nature , 411 : 927 – 30 .
- Harding , GC , Kenchington , EL , Bird , CJ , Pezzack , DS and Landry , DC . 1997 . Relationships among subpopulations of American lobster (Homarus americanus) as revealed by random amplified polymorphic DNA . Canadian Journal of Fisheries and Aquatic Sciences , 54 : 1762 – 71 .
- Haren Noman D van . 1881 . Die Lamellibranchiaten gesammelt während der Fahrten des ‘Willem Barents’, 1878 und 1879 . Archiv für Zoologie, Supplement-Band I, Zoologischen Ergebnisse der zwei in der Jahren 1878 und 79 ausgeführten Fahrten des Schoners ‘Willem Barents’. Leiden: E.J. Brill and Leipzig: C.F. Winter. 51 pages .
- Høisæter , T . 1986 . An annotated check-list of marine molluscs of the Norwegian coast and adjacent waters . Sarsia , 71 : 73 – 145 .
- ICZN . 1999 . International Code of Zoological Nomenclature (4th ed) . London : The International Trust for Zoological Nomenclature .
- Jeffreys , JG . 1869 . British Conchology, or an Account of the Mollusca which now Inhabit the British Isles and the Surrounding Seas , Vol. 5 , London : J. van Voorst .
- Jeffreys , JG . 1876 . New and peculiar Mollusca of the Pecten, Mytilus and Arca families, procured in the Valorous expedition . Annals and Magazine of Natural History. ser. 4, , 18 : 424 – 36 .
- Jeffreys , JG . 1879 . On the Mollusca procured during the ‘Lightning’ and ‘Porcupine’ expeditions 1868–70. (II). Proceedings of the Zoological Society of London . for , 1879 : 553 – 588 .
- Jensen , AS . 1904 . Pecten frigidus, nomen pectini profundorum maris polaris incolae novum datum . Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening i Kjøbenhavn , 1904 : 305 – 11 .
- Jensen , AS . 1912 . Lamellibranchiata, Part I . The Danish-Ingolf Expedition , 2 ( 5 ) : 1 – 119 .
- Johannesson , K . 1988 . The paradox of Rockall: Why is a brooding gastropod (Littorina saxatilis) more widely dispersed than one having a planktonic larval dispersal stage (L. littorea)? . Marine Biology , 99 : 507 – 13 .
- Kit , Y and Kornfield , I . 1998 . Phylogenetic relationships of clawed lobster genera (Decapoda: Nephropidae) based on mitochondrial 16S rRNA gene sequences . Journal of Crustacean Biology , 18 : 138 – 46 .
- Knipowitsch , N . 1901 . Zoologische Ergebnisse der Russischen Expeditionen nach Spitzbergen. Mollusca und Brachiopoda. I. […] . Ezhegodnik Zoologicheskago Muzeya Imperatorskoi Akademii Nauk. , 6 : 435 – 558 .
- Knudsen , J . 1985 . Abyssal Mollusca of the Arctic Ocean . Journal of Conchology , 32 : 97 – 107 .
- Laskey , J . 1811 . Account of North British Testacea . Memoirs of the Wernerian Natural History Society , 1 : 370 – 417 .
- Leche , W . 1878 . Fversigt öfver de af svenska expeditionerna till Novaja Semlja och Jenissej 1875 och 1876 insamlade hafs-mollusker . Kongliga Svenska Vetenskaps-Akademiens Handlingar , 16 ( 2 ) : 1 – 86 .
- Leche , W . 1883 . fversigt öfver de af Vega-Expeditionen insamlade Arktiska hafsmollusker. I. Lamellibranchiata . Vega-Expeditionens Vetenskapliga lakttagelser , 3 : 435 – 53 .
- Leifsdóttir , ÓE and Simonarson , LA . 2001 . Vard fjörudoppa (Littorina littorea) til á Ìslands Færeyjarhryggnum sudaustur af Íslandi . Natturufrædingurinn , 70 : 97 – 110 .
- Linnaeus , C . 1758 . Systema naturae per regna tria naturae […] I, editio decima, reformata , Holmiae : Salvii .
- Linnaeus , C . 1771 . Mantissa Plantarum altera Generum editionis VI. & Specierum editionis II. […] , Holmiae : Salvii .
- Locard , A . 1888 . Monographie des espèces appartenant au genre Pecten . Contributions à la faune malacologique Française , 11 : 1 – 159 .
- Locard , A . 1898 . Expéditions scientifiques du ‘Travailleur’ et du ‘Talisman’, 1880–1883. Mollusques testacés , Vol. 2 , 517 – 1044 . Paris : Masson .
- Lovén S. 1846 . Nordens Hafs-Mollusker. Index Molluscorum. Litora Scandinaviae occidentalia habitantium . Öfversigt af Kongliga Vetenskaps-Akademiens Förhandlingar 3:134–61 (meeting of May), 182–204 (meeting of June). The proceedings of these meetings were published less than two months after the meeting, considering that October meetings 1845–65 which have a printing date, are dated the year they were held; November meetings next year. The paper was also published as reprint with a new title page and paginated pp. 1–50 and dated Holmiae, P.A.Nordstedt et Fili, 1846 .
- Lubinsky , I . 1980 . Marine bivalve molluscs of the Canadian central and eastern Arctic: Faunal composition and zoogeography . Canadian Bulletin of Fisheries and Aquatic Sciences , 207 : i – vi . 1–111
- Lucas , M . 1979 . The Pectinoidea of the European coasts . La Conchiglia , 11 : 15 – 18 .
- Mabille , J and Rochebrune , AT . 1889 . Mollusques. Mission Scientifique du Cap Horn 1882–1883 , Vol. 6 , Paris : Imprimeurs Gauthier, Villars et fils .
- Madsen , FJ . 1949 . “ Marine Bivalvia ” . In The Zoology of Iceland Edited by: Fridriksson , A and Tuxen , SL . Vol. 4 , 1 – 116 .
- Mörch , OAL . 1868 . Faunula molluscorum Islandiae. Oversigt over Islands blöddyr . Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening i Kjøbenhavn , 0 ( 11–13 ) : 185 – 227 .
- Morton , B and Thurston , MH . 1989 . The functional morphology of Propeamussium lucidum (Bivalvia: Pectinacea), a deep-sea predactory scallop . Journal of Zoology , 218 : 471 – 96 .
- Müller , OF . 1776 . Zoologiae Danicae Prodomus, […] , Havniae : Hallageriis .
- Müller , OF . 1788 . Zoologia Danica, […] , Vol. 2 , Havniae : N. Möller .
- Ockelmann , WK . 1959 . The zoology of East Greenland. Marine Lamellibranchiata . Meddelelser om Grønland , 122 ( 4 ) : 1 – 256 .
- Oskarsson , I , Ingólfsson , A and Gardarsson , A . 1977 . Stranddoppa (Hydrobia ventrosa) á Íslandi . Natturufrædingurinn , 47 : 8 – 15 .
- Oskarsson I. 1982 . Skeldyrafana Islands . Reykjavik : Prentsmidjan Leiftur HF . 351 pages. [This book contains ‘Samlokur i sjo’ (Marine bivalves) (1st ed. 1952, 2nd ed. 1964) and ‘Saesniglar med skel’ (Marine gastropods with a shell) (1st ed. 1962) with a supplement.]
- Payraudeau BC. 1826 . Catalogue descriptif et méthodique des Annelides et des Mollusques de l'Ile de Corse . Imprimerie de J. Tastu . 218 pages .
- Petersen , GH . 1968 . Marine Lamellibranchiata . Zoology of the Faroes , # ( 1 ) : 1 – 80 .
- Poli , J . 1791 . Testacea utriusque Siciliae eorumque historia et anatome , Parmae : Regio Typographeio . Vol 2.
- Posselt , HJ and Jensen , A . 1898 . Grønlands Brachiopoder og Bløddyr . Meddelelser om Grønland , 23 : i – xix . 1–298
- Reeve L. 1852–53 . Conchologia Iconica: or, Illustrations of the shells of molluscous animals, vol. 8, Monograph of the genus Pecten . London : Lovell Reeve. No pagination , 35 plates .
- Rex , MA and Warén , A . 1982 . Planktotrophic development in deep-sea prosobranch snails from the western North Atlantic . Deep-Sea Research (2A) , 29 : 171 – 84 .
- Richling , I . 2000 . Arktische Bivalvia – eine taxonomische Bearbeitung auf Grundlage des Materials der Expeditionen Transdrift 1 und ARK IX/4 (1993) in das Laptevmeer . Schriften zur Malakozoologie , 15 : 1 – 93 .
- Risso , A . 1826 . Histoire naturelle des principales productions de l’ Europe Méridionale et particulièrement de celles des environs de Nice et des Alpes maritimes. 4 , Paris : Levrault .
- Rothlisberg , PC and Pearcy , WG . 1976 . An epibenthic sampler used to study ontogeny of vertical migration of Pandalus jordani (Decapoda, Caridea) . Fisheries Bulletin , 74 : 994 – 7 .
- Salas , C . 1996 . Marine bivalves from off the southern Iberian Peninsula collected by the Balgim and Fauna 1 expeditions . Haliotis , 25 : 33 – 100 .
- Sars , GO . 1878 . Bidrag om kundskaben om Norges arktiske fauna. I. Mollusca regionis arcticae Norwegiae , Christiania : Christiania. AW Brögger .
- Sars , M . 1869 . Fortsatte bemærkninger over det dyriske livs utbredning i havets dybde . Forhandlinger i Videnskabs-Selskabet Kristiania , 1868 : 246 – 75 .
- Schein , E . 1989 . Pectinidae (Mollusca, Bivalvia) bathyaux et abyssaux des campagnes BIOGAS (Golfe de Gascogne) systématique et biogéographie . Annales de l'Institut Océanographique Nouvelle Série , 65 : 59 – 125 .
- Schiøtte , T . 1989 . Marine Mollusca from Jørgen Brønlund Fjord, North Greenland, including the description of Diaphana vedelsbyae n.sp. Meddelelser om Grønland . Bioscience , 28 : 1 – 24 .
- Smaldon G , Heppell D , Watt KR . 1976 . Type specimens of invertebrates (excluding insects) held at the Royal Scottish Museum, Edinburgh . Royal Scottish Museum Information Series, Natural History 4 : I – iv , 1–118 .
- Smith , EA . 1885 . Report on the Lamellibranchiata collected by H.M.S. ‘Challenger’ during the years 1873–1876. Challenger Reports . Zoology , 13 ( 35 ) : 1 – 341 .
- Smriglio , C and Mariottini , P . 1990 . Descrizione di una nuova specie di Pectinidae (Rafinesque, 1815) per il Mar Mediterraneo […] . Bollettino Malacologico , 26 : 1 – 18 .
- Sneli , JA . 1998 . A simple benthic sledge for shallow and deep-sea sampling . Sarsia , 83 : 69 – 72 .
- Soot-Ryen , T . 1958 . Pelecypods from East Greenland . Skrifter fra Norsk Polarforskningsinstitutt , 113 : 1 – 32 .
- Soot-Ryen , T . 1963 . Notes on the species of Pecten described by O.F. Müller, 1776 . Proceedings of the Malacological Society of London , 35 : 111 – 12 .
- Sowerby , GB. II . 1842 . Thesaurus Conchyliorum, or figures and descriptions of recent shells. Part 2. Monographs of the genera Pecten and Hinnites , 45 – 80 . London : Sowerby .
- Thomson , CW . 1873 . The Depth of the Sea. […] , London : Macmillan and Co .
- Thorson , G . 1941 . Marine Gastropoda Prosobranchiata . Zoology of Iceland , 4 ( 60 ) : 1 – 150 .
- Thorson , G . 1951 . The Godthaab expedition 1928. Scaphopoda, Placophora, Solenogastres, Gastropoda, Prosobranchiata, Lamellibranchiata . Meddelelser om Grønland , 81 ( 2 ) : 1 – 117 .
- Verrill AE . 1873 . Results of recent dredging expeditions on the coast of New England . American Journal of Science ( 3 ) 5 : 1 – 16 , 98–106 .
- Verrill , AE . 1897 . A study of the family Pectinidae with a revision of the genera and subgenera . Transactions of the Connecticut Academy of Arts and Science , 10 : 41 – 95 .
- Wagner , HP . 1988 . The status of four scallop species (Mollusca; Bivalvia; Pectinidae), with description of a new genus . Basteria , 52 : 41 – 4 .
- Wagner , HP. Undated . 1991 . Review of the European Pectinidae. Overzicht van de Europese Pectinidae (Mollusca: Bivalvia) . Vita Marina , 41 : 1 – 48 .
- Waller , TR . 1972 . The Pectinidae (Mollusca: Bivalvia) of Eniwetok Atoll, Marshall Islands . The Veliger , 14 : 221 – 64 .
- Waller , TR . 1984 . The ctenolium of scallop shells: functional morphology and evolution of a key family-level character in the Pectinacea (Mollusca, Bivalvia) . Malacologia , 25 : 203 – 19 .
- Waller , TR . 1991 . “ Evolutionary relationship among commercial scallops (Mollusca: Bivalvia: Pectinidae) ” . In Scallops, Biology, Ecology and Aquaculture , Edited by: Shumway , SE . 1 – 73 . Amsterdam : Elsevier .
- Waller , TR . 1993 . The evolution of ‘Chlamys’ (Mollusca: Bivalvia: Pectinidae) in the tropical western Atlantic and eastern Pacific . American Malacological Bulletin , 10 : 195 – 249 .
- Waller , TR . 2006 . “ New phylogenies of the Pectinidae (Mollusca: Bivalvia): Reconciling morphological and molecular approaches ” . In Scallops: Biology, Ecology and Aquaculture , Edited by: Shumway , SE and Parsons , GJ . 1 – 44 . Amsterdam : Elsevier .
- Warén , A . 1980 . Marine Mollusca described by John Gwyn Jeffreys, with the location the type material . Conchological Society of Great Britain and Ireland Special Publication , 1 : 1 – 60 .
- Wares , JP , Goldwater , DS , Kong , BY and Cunningham , CW . 2002 . Refuting a controversial case of a human-mediated marine species introduction . Ecology Letters , 5 : 577 – 84 .